

Abstract
Background. Antrodia cinnamomea is an edible fungus commonly used in Asia as a well-known medicinal herb capable of treating drug intoxication and liver cancer. Methods. This study evaluated the anticancer activity of its biotechnological product, mycelial fermentation broth (AC-MFB) on hepatocellular carcinoma (HCC) by tetrazolium-based colorimetric assay in vitro and syngeneic Balb/c 1MEA.7R.1 tumor implantation model in vivo. Given that cancer stem cell characteristics, such as angiogenesis, invasiveness, and migration, are known to cause recurrence, we further evaluated the effect of AC-MFB on cellular viability inhibition of HCC cells, angiogenic activity and migration of endothelial cells, and the release of proangiogenic factors from HCC cells. Results. We found that AC-MFB markedly inhibited the growth of HCC without hepatic enzyme abnormality. This anti-HCC activity was validated by growth-inhibitory effects on both cultured murine 1MEA.7R.1 and human HA22T/VGH HCC cells. For cancer stem cell characteristics, AC-MFB inhibited the cellular viability, migration, and tube formation activity of EA. hy926 and SVEC4-10 endothelial cells. Production of extracellular vascular endothelial growth factor and intracellular hypoxia-inducible factor-1 alpha from HCC cells was suppressed by AC-MFB. Conclusion. Antrodia cinnamomea could inhibit the growth and cancer stem cell characteristics of HCC cells.
1. Background
Antrodia cinnamomea, also known as niu-chang-chih, Taiwanofungus camphorate or Antrodia camphorata, is a fungus indigenous to Taiwan which grows on decayed Cinnamomum kanehirae [1–3]. In folk medicine, it is well known for its antidotal and antitumoral functions as it has been traditionally used to treat toxicities caused by food, alcohol, and drugs as well as diarrhea, abdominal pain, hypertension, itchy skin, and tumors [4, 5].
A. cinnamomea has been shown to exhibit anticancer properties [6–9] with many studies aimed at exploring the exact bioactive compounds [10–14]. For example, Chen et al. reported that the oral administration of A. cinnamomea fruiting bodies significantly increased the life span of ATCC BNL IMEA.7R.1 hepatoma-bearing mice [15]. It has also been reported that polysaccharides from A. cinnamomea mycelial extract could inhibit angiogenic activities in endothelial cells (ECs) [16–18].
To avoid contamination in the wild source and to expand the production of this fungus for food use, A. cinnamomea mycelial fermentation broth (AC-MFB) produced using biotechnology has been developed by our group and others. Although AC-MFB is not used as natural form, it could be regarded as a modern formulation of this herbal drug. However, the anticancer activity and safety of these biotechnological products have not been extensively examined.
We previously purified a series of compounds from the fruiting body of A. cinnamomea, including 24-methylenelanosta-7,9(11)-diene-3b,15adiol-21-oic acid (designated herein as MMH01), which possesses cytotoxicity against human leukemia and pancreatic cancer cells [19]. In the present study, we investigated the anticancer effect and safety of AC-MFB with a syngeneic hepatocellular carcinoma (HCC) implantation model. The effect of AC-MFB against cancer stem cell characteristics in HCC and endothelial cells was also evaluated.
2. Methods
2.1. Plants Materials, Fermentation, and Characterization
A. cinnamomea, strain B137, identified by fungi specialist Dr. T. T. Chang (Taiwan Forestry Research Institute, Taipei, Taiwan), was maintained on potato dextrose agar pasteurized Petri dishes and transferred to a fresh medium at 1-month intervals. Mycelial agar discs (8 pieces, 0.5 cm each) were obtained by a sterilized tip and used as the inoculum in a shake flask preculture. The preculture medium (LM-B) consisted of the following components (mg/mL): glucose 30, sucrose 20, yeast extract 15, peptone 13, MgSO4 0.3, KH2PO4 0.3, and K2HPO4 0.3 with an initial condition of pH 4.0. For the preculture, a 200 mL medium was prepared in a 500 mL flask and inoculated, followed by a 7-day incubation at 28°C on a rotary shaker (100 rpm). For a series of experiments, a 100 mL medium (LM-B) was prepared in a 500 mL flask and inoculated with mycelium suspension (6%) from the preculture broth, followed by a 14-day incubation at 28°C on a rotary shaker (100 rpm). The fermentation product was then harvested and poured through a nonwoven fabric on a 20-mesh sieve to separate the deep-red fermented culture broth and the mycelia, followed by centrifugation at 3000 g for 10 min and then by passage through a filter of 0.22 μm pore size. The culture broth (1.0 L) was then concentrated into a colloid (47.6 mg) under vacuum and stored at −30°C before the analysis. The AC-MFB colloid was redissolved and directly taken into the study without further extraction. The polysaccharide components in AC-MFB were used in quantity analysis for standardization in this study. The phenol-sulfuric acid assay, a colorimetric method, was employed to determine the total concentration of carbohydrates present in AC-MFB [20].
2.2. In Vivo Tumorigenesis and Hepatic Toxicity Assay in Mice
A subcutaneous allogenic mice animal model was used for tumorigenesis assay. Specific pathogen-free male BALB/c mice (4 weeks old, 25–28 gm) were obtained from the National Laboratory Animal Center (Taipei, Taiwan) and were maintained in pathogen-free conditions. They were kept in our animal facility for at least two weeks before use. All of the mice were used at the age of 6–8 weeks. All animals were cared for following the Guide for the Care and Use of Laboratory Animals (NIH publication no. 85-23, revised in 1985). The mice were subcutaneously injected with 1 × 105 1MEA.7R.1 HCC cells over the flank. The mice were fed orally either with 50 mg/mL AC-MFB 200 μL or with a vehicle of normal saline 200 μL (N = 6 for each group) for 28 days. The tumor size of each mouse was measured by a caliper and calculated by the formula: L × W 2/2, where W was the shortest dimension and L was the longest dimension in centimeters. The tumor size was estimated every 2 to 3 days after the tumor size increased to 0.1 mL in size. Tumor growth curves were plotted as the mean tumor volume relative to the treatment group.
Hepatic enzyme alanine aminotransferase (ALT) is found primarily in the cytoplasm of hepatic cells which catalyses gluconeogenesis from noncarbohydrate sources and is an important marker for liver injury [21, 22]. Elevation of the serum concentrations of this marker implied disruption of plasma membrane integrity, which eventually led to leakage of the enzymes into the blood circulation [23]. ALT was used as a biochemical marker for hepatic damage assay in this study. Animals were fed with different doses of AC-MFB by gastric tube for each condition for 28 days (Table 1); each group had 6 mice. Blood samples were collected for the assays of ALT on the first day as a control and on the third day and the 28th day for acute and chronic hepatic toxicity assays, respectively. Serum activities of ALT were determined using the colorimetric method [24].
Table 1.
Effect of AC-MFB on ALT levels in mice.
| Treatment group | Dose (g/Kg BW) | Day 0 | Day 3 | Day 28 |
|---|---|---|---|---|
| Normal saline 200 μL | 0.0 | 75 ± 20 | 96 ± 62 | 60 ± 2 |
| 50 mg/mL 200 μL | 0.4 | 49 ± 12 | 48 ± 11 | 44 ± 4 |
| 100 mg/mL 200 μL | 0.8 | 111 ± 52 | 60 ± 12 | 126 ± 42 |
| 250 mg/mL 200 μL | 2.0 | 78 ± 42 | 44 ± 10 | 44 ± 2 |
| 500 mg/mL 200 μL | 4.0 | 73 ± 36 | 93 ± 42 | 77 ± 22 |
| 500 mg/mL 300 μL | 6.0 | 130 ± 95 | 56 ± 15 | 160 ± 79 |
All values represent mean ± SD. Normal value of mouse ALT: 28–132 U/L.
N = 6 for each group.
The protocol used in this study was approved by the Institutional Animal Care and Use Committee, Chang Gung University, Taiwan (IACUC approval no. CGU10-143).
2.3. In Vitro Cytotoxicity Assays
The in vitro cytotoxicities of the AC-MFB on both HCC cells and ECs were examined using a modified MTT assay [25]. The human ECs EA. hy926 (a gift from Dr. Cora-Jean S. Edgell, Pathology Department, University of North Carolina), human HCC cell HA22T/VGH [26], and the BALB/c murine BNL 1MEA.7R.1 (BNL) HCC cells were grown in DMEM, containing 584 mg/mL L-glutamine and 10% FBS. The murine SV40-transformed mouse ECs (SVEC4-10, ATCC no. CRL-2181) were maintained in DMEM with 10% FBS.
2.4. In Vitro Antiangiogenic Assays
The antiangiogenic activity was investigated by the capability of endothelial cell migration and tube formation on Matrigel in vitro.
Endothelial cell migration was determined by means of a wound-healing assay. ECs were seeded into 6-well tissue culture dishes and cultured in medium containing 10% FBS to confluent cell monolayers, which were then carefully wounded using sterile 200 μL pipette tips with all cell debris removed by phosphate buffered saline (PBS). The cells were then incubated for another 24 h in medium with or without AC-MFB treatment. The cells that had migrated across the edge of the wound were photo recorded under an inverted phase-contrast microscope (Nikon, Tokyo, Japan), and the cellular migration was determined by measuring the widths of the wounds. The differences in the width of the wounds were measured with Adobe Photoshop version 5.5 (Adobe Systems Incorporated, USA). Each data point was compared with its own control present in the control dish at 24 h and interpreted as a percentage of wound migration inhibition.
Matrigel (BD BioCoat Angiogenesis System, Franklin Lakes, NJ, USA) was employed to promote the differentiation of ECs into capillary tube-like structures. A total of 75 μL of thawed Matrigel was added to 96-well tissue culture plates, followed by incubation for 60 min at 37°C to allow polymerization. Then, 4 × 104 quiescent ECs were seeded on the gels in medium supplemented without FBS in the presence or absence of AC-MFB, followed by incubation at 37°C in 5% CO2 for 12 h. Tube formation was examined by an inverted phase-contrast microscope (Nikon, Tokyo, Japan). The number of the tubes was counted with Adobe Photoshop version 5.5 (Adobe Systems Incorporated, USA). Each data point was compared with its own control (without AC-MFB) at 12 h and interpreted as a percentage of tube formation inhibition.
2.5. Inhibition of Proangiogenic Factors from Hepatic Cancer Cells
Secretion of VEGF and HIF-1α by HCCs was stimulated by 2 h incubation with 200 μM desferrioxamine (DFO) (Sigma, St. Louis, MO, USA) to mimic hypoxia. VEGF levels of HA22T and 1MEA.7R.1 HCCs were measured by Human VEGF ELISA kit no. DY293B and Mouse VEGF ELISA no. DY493, respectively. The HIF-1α was measured using a solid-phase enzyme immunoassay (“sandwich ELISA,” NOVUS and NeoMarkers, Inc.). Each measurement was done in triplicate, and the data were calculated and normalized to total protein in the culture media.
2.6. Statistical Analysis
Data from three separate experiments were expressed as a mean ± standard deviation (SD). Statistical comparisons of the results were made using analysis of variance [27] as indicated. Significant differences (P < 0.05) between the means of the two test groups were analyzed by Tukey's test.
3. Results
3.1. Characterization and Stability of AC-MFB
Although the bioactive ingredients of AC-MFB remain to be determined, the amount of polysaccharides in a concentrated AC-MFB colloid was used as the standard of quality control to characterize each batch of product. The phenol-sulfuric acid method showed that there were 0.341 mg polysaccharides in 47.6 mg AC-MFB colloid from 1.0 L fermentation broth. The polysaccharides shared 0.7% weight in the concentrated cultured broth colloid. There was no significant change in the amount of polysaccharides in AC-MFB throughout the 12 months of storage, indicating it is a product with long-term stability.
3.2. AC-MFB Inhibited Tumor Growth in Mice without Hepatic Enzyme Abnormality
In BALB/c mice bearing 1MEA.7R.1 liver cancer cells, oral intake of 200 μL AC-MFB (50 mg/mL) for 28 days significantly inhibited the tumor growth in comparison with the vehicle control (P < 0.05) (Figure 1).
Figure 1.
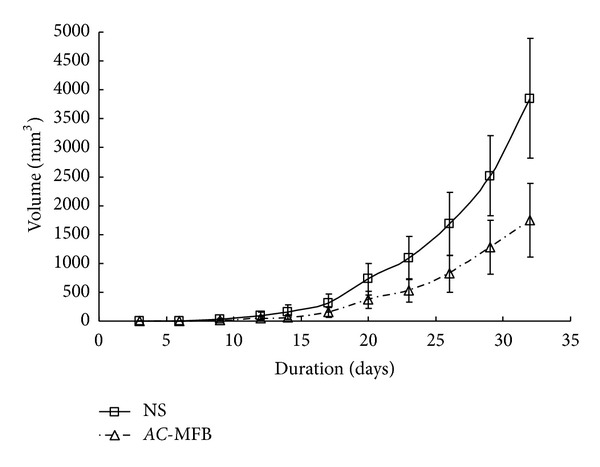
Antitumor effect of AC-MFB on BALB/c liver cancer. Effect of AC-MFB on growth of 1MEA.7R.1 liver cancer cells in BALB/c mice. Mice were treated with oral AC-MFB (50 mg/mL) or normal saline for 28 days. *P < 0.05.
Mice serum ALT levels at 3 days and 28 days after treatment were examined for estimation of liver toxicity. As shown in Table 1, there was no statistically significant difference between serum ALT levels of AC-MFB and control groups after 3 days and 28 days of AC-MFB oral intake.
3.3. AC-MFB Inhibited Cellular Viability of HCC Cells and ECs
As shown in Figures 2 and 3, AC-MFB inhibited the viability of HCC cells (human HA22T/VGH, murine 1MEA.7R.1) and ECs (EA. hy926, SVEC4-10) in both dose-dependent and time-dependent parameters.
Figure 2.
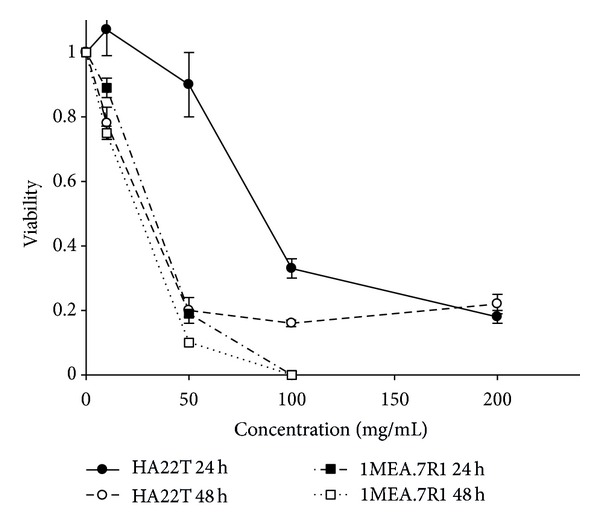
Cytotoxicity of AC-MFB on hepatocellular carcinoma cell. Effect of AC-MFB on cellular viability of HA22T and 1MEA.7R.1 HCC cells by MTT assay. Data from three separate experiments were expressed as percentages of untreated cells after 24 and 48 h.
Figure 3.
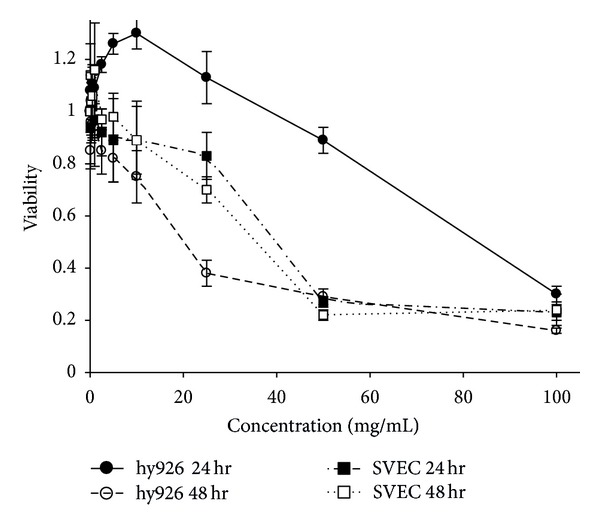
Cytotoxicity of AC-MFB on endothelial cell. Effect of AC-MFB on cellular viability of EA. hy926 and SVEC4-10 ECs by MTT assay. Data from three separate experiments were expressed as percentages of untreated cells after 24 and 48 h.
3.4. AC-MFB Inhibited Endothelial Cell Migration and Tube Formation Activity
The migration and tube formation activity of ECs are important in vitro endpoints for angiogenesis. As demonstrated in Figure 4, AC-MFB profoundly inhibited the migration of EA. hy926 and SVEC4-10 cells in a concentration-dependent manner. The IC50 values of migration inhibition of EA. hy926 and SVEC4-10 were 8.8 ± 1.6 mg/mL and 3.0 ± 0.3 mg/mL, respectively. The tube formation activity in these two endothelial cell lines was also inhibited by treatment using AC-MFB with IC50 ranging from 1.4 to 3.1 mg/mL (Figure 5).
Figure 4.

Migration inhibition of AC-MFB on endothelial cell. Effect of AC-MFB on migration of EA. hy926 ((a)–(d)) and SVEC4-10 ((e)–(h)) ECs by in vitro wound-healing migration assay. Data from three separate experiments were expressed as mean inhibition percentages compared with untreated cells after 24 h treatment.
Figure 5.

Tube formation inhibition of AC-MFB on endothelial cell. Effect of AC-MFB on tube formation activity of EA. hy926 ((a)–(d)) and SVEC4-10 ((e)–(h)) ECs. Data from three separate experiments were expressed as mean inhibition percentages compared with untreated cells after 12 h treatment.
3.5. AC-MFB Suppressed Production of VEGF and Downregulated Intracellular HIF-1α Levels in HA22T/VGH and 1MEA.7R.1 Cell Lines
Given that VEGF plays a critical role in angiogenesis, we investigated whether AC-MFB could modulate the secretion of VEGF from HCC cells. As shown in Figure 6, AC-MFB treatment suppressed the DFO-stimulated VEGF secretion in both HCC cell lines, but only the difference in HA22T/VGH cells reached statistical significance (P < 0.05).
Figure 6.
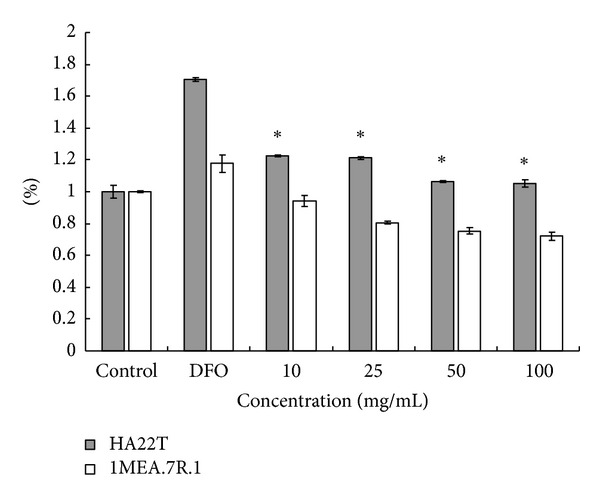
AC-MFB inhibits VEGF from hepatocellular cancer cell. Effect of various concentrations of AC-MFB on extracellular VEGF levels in culture of HA22T and 1MEA.7R.1 HCC cells. Cells were pretreated with 200 μM DFO for 2 h followed by AC-MFB for 6 h. All the data are normalized to the control group (AC-MFB 0.0 mg/mL). Data from three independent experiments in triplicate were expressed as mean ± standard error of mean; *P < 0.05 versus 200 μM DFO.
Hypoxia-inducible factor-1α (HIF-1α) is a protein that regulates gene transcription through binding to hypoxia response elements, resulting in upregulation of VEGF and thereby enhancing angiogenesis. AC-MFB treatment significantly downregulated the DFO-stimulated intracellular HIF-1α levels in both HCC cell lines (P < 0.05) (Figure 7).
Figure 7.
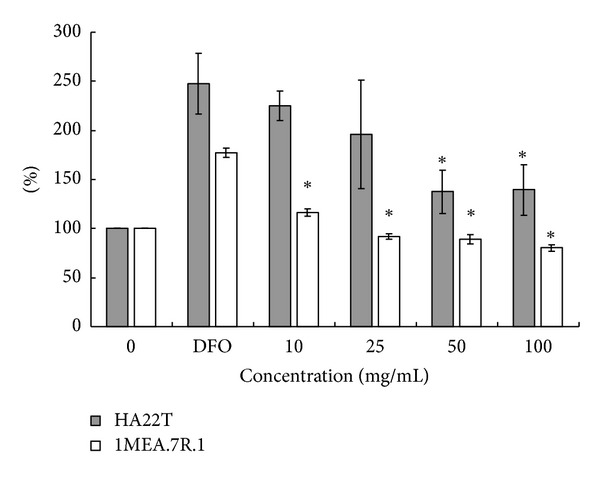
AC-MFB inhibits HIF-1α from hepatocellular cancer cell. Effect of various concentrations of AC-MFB on intracellular HIF-1α levels in culture of HA22T and 1MEA.7R.1 HCC cells. Cells were pretreated with 200 μM DFO for 2 h followed by AC-MFB for 2 h. All the data were normalized to the control group (AC-MFB 0 mg/mL). Data from three independent experiments in triplicate were expressed as mean ± standard error of mean; *P < 0.05 versus 200 μM DFO.
4. Discussion
In this study, we examined the in vivo activity of AC-MFB first. Since the animal model showed it was effective without major toxicity, we next evaluated the in vitro effect of AC-MFB on both HCC cells and endothelial cells to explore possible targets. Based on the antiangiogenesis activity of AC-MFB, we further assessed the modulation of angiogenic factors by AC-MFB.
The results showed that AC-MFB could inhibit the viability of HCC cells (human HA22T/VGH, murine 1MEA.7R.1) and ECs (EA. hy926, SVEC4-10) in both dose-dependent and time-dependent parameters in vitro (Figures 1 and 2). The AC-MFB could also inhibit the migration and tube formation activities in both ECs (Figures 4 and 5). In vivo, oral intake of AC-MFB inhibited the tumor growth in BALB/c mice bearing 1MEA.7R.1 liver cancer cells without hepatic enzyme abnormality (Figure 3, Table 1). In bimolecular activity, AC-MFB could suppress the secretion of VEGF and downregulate the HIF-1α level (Figures 6 and 7).
The A. cinnamomea mycelial fermentation broth, a biotechnological product serving as a medicinal herb, could inhibit the growth of hepatocellular carcinoma in vivo without liver toxicity. Further investigation to clarify the antitumoral mechanisms revealed an inhibitory activity on angiogenesis and the release of related proangiogenic factors VEGF and HIF-1α, indicative of inhibitory activity related to cancer stem cell characters.
Natural A. cinnamomea is very rare and slowly growing. Moreover, the host wood, Cinnamomum kanehirae, is a local species that is getting scarce and difficult to find in the forest. Therefore, the extracts of the fruiting body are difficult to obtain for use as a medicinal herb. Through biotechnological production in a fermentation broth and the examination of its bioactivity against HCC, a scientific basis for AC-MFB to serve as a medicinal herb in adjunct cancer treatment was found.
Hepatocellular carcinoma is one of the most frequently occurring malignancies with poor prognosis in Taiwan [28]. A. cinnamomea has attracted great attention due to its hepatoprotective effect and antitumoral activity against HCC [4, 7, 9]. Experimental and clinical data had indicated that HCC tumor progression is associated with angiogenesis and that an increase in microvascular density is associated with a poor prognosis [29]. Given that HCC is characterized by hypervascularity [30], antiangiogenesis might be one of the primary treatment approaches [31].
Our study showed that not only did AC-MFB have antitumoral activity against HCC cells in vitro, it also delayed 1MEA.7R.1 HCC growth without hepatic enzyme abnormality in vivo. In other words, AC-MFB might be a potential therapeutic agent or an adjuvant regimen for treatment of HCC.
Furthermore, AC-MFB inhibited the cellular events related to cancer stem cells, such as angiogenesis, migration, and HIF-α expression. For antiangiogenesis, our study also showed that AC-MFB could inhibit the cellular viability, migration, and tube formation of ECs and decreased both the extracellular VEGF and the intracellular HIF-1α levels of HCC cells under hypoxia. These findings suggest that the antiangiogenic activity of AC-MFB might involve inhibition of HIF-1α transcription and downregulation of VEGF expression in HCC cells. Such claims could be supported by previous investigations. For example, Yang reported that fractionated polysaccharides from A. cinnamomea mycelia, with molecular weight (MW) > 100 kDa, are antiangiogenic in vitro and ex vivo, and the effects are likely through immunomodulation [17]. Cheng et al. not only reported that polysaccharides extracted from fermented A. cinnamomea mycelia could inhibit VEGF receptor phosphorylation and angiopoietin-2 protein expression but also found the chemical structure of the polysaccharide [16, 18]. Shih et al. also reported that extracted polysaccharides from the fruiting body of A. cinnamomea with a molecular weight between 2693 and 2876 kDa had a critical inhibitory effect on tube formation without cytotoxicity [32]. Polysaccharides were also proved to elicit an antitumoral effect by promoting a Th1-dominant state and killer activities [6]. The polysaccharides only shared 0.7% weight of our concentrated cultured broth colloid; soother lanostane-type components like MMH01 isolated from A. cinnamomea could have anticancer activity [33]. Taken together, polysaccharides may not be the major bioactive component in AC-MFB. Instead, the amount of exopolysaccharide could serve as a good standardization index for each batch of fermented products.
Our study is the first one demonstrating the antiangiogenic activity of A. cinnamomea mycelial fermentation broth, not from extracts of fruiting body. It indicates that medicinal herbal products of A. cinnamomea may possess the original bioactivity against HCC and angiogenesis.
5. Conclusion
The A. cinnamomea mycelial fermentation broth, a biotechnological product serving as a medicinal herb, could inhibit growth of hepatocellular carcinoma in vivo. Furthermore, AC-MFB could inhibit the cellular events related to cancer stem cells, such as angiogenesis, migration, and HIF-α expression in HCC.
Authors' Contribution
Y.-M. Liu and Y.-K. Liu contributed to this work equally.
Conflict of Interests
The authors declare that they have no competing interests in the publication of the paper.
Acknowledgments
The authors thank the great help from Ms. S. L. Chou and Dr. P. I. Huang from Cancer Center of Taipei Veterans General Hospital, Taipei, Taiwan, for their great assistance in this study.
Abbreviations
- AC-MFB:
An tr od ia cinnamomea mycelial fermentation broth
- HCC:
Hepatocellular carcinoma
- ECs:
Endothelial cells
- MTT:
Modified tetrazolium-based colorimetric assay (3-(4,5-dimethylthiazol)-2,5-diphenyl tetrazolium bromide assay
- VEGF:
Vascular endothelial growth factor
- HIF-1α:
Hypoxia-inducible factor-1α
- ALT:
Alanine aminotransferase.
References
- 1.Wu SH, Ryvarden L, Chang TT. Antrodia cinnamomea (“niu-chang-chih”), new combination of a medicinal fungus in Taiwan. Botanical Bulletin of Academia Sinica. 1997;38:273–275. [Google Scholar]
- 2.Chang TT, Chou WN. Antrodia cinnamomea sp. nov. on Cinnamomum kanehirai in Taiwan. Mycological Research. 1995;99(6):756–758. [Google Scholar]
- 3.Zang M, Su CH. Ganoderma comphoratum, a new taxon in genus Ganoderma from Taiwan. China Acta Botanica Yunnanica. 1990;12:395–396. [Google Scholar]
- 4.Tsai ZT, Liaw SL, editors. The Use and the Effect of Ganoderma. Taichung, Taiwan: Sheng-Yun Publisher; 1982. [Google Scholar]
- 5.Wang BC, Huang RC. The development and marketing of Ganoderma lucidum and Antrodia camphorata . Food Industry. 2002;34(5):3–14. [Google Scholar]
- 6.Liu JJ, Huang TS, Hsu ML, et al. Antitumor effects of the partially purified polysaccharides from Antrodia camphorata and the mechanism of its action. Toxicology and Applied Pharmacology. 2004;201(2):186–193. doi: 10.1016/j.taap.2004.05.016. [DOI] [PubMed] [Google Scholar]
- 7.Hsu YL, Kuo YC, Kuo PL, Ng LT, Kuo YH, Lin CC. Apoptotic effects of extract from Antrodia camphorata fruiting bodies in human hepatocellular carcinoma cell lines. Cancer Letters. 2005;221(1):77–89. doi: 10.1016/j.canlet.2004.08.012. [DOI] [PubMed] [Google Scholar]
- 8.Hseu YC, Yang HL, Lai YC, Lin JG, Chen GW, Chang YH. Induction of apoptosis by Antrodia camphorata in human premyelocytic leukemia HL-60 cells. Nutrition and Cancer. 2004;48(2):189–197. doi: 10.1207/s15327914nc4802_9. [DOI] [PubMed] [Google Scholar]
- 9.Hsu YL, Kuo PL, Cho CY, et al. Antrodia cinnamomea fruiting bodies extract suppresses the invasive potential of human liver cancer cell line PLC/PRF/5 through inhibition of nuclear factor κB pathway. Food and Chemical Toxicology. 2007;45(7):1249–1257. doi: 10.1016/j.fct.2007.01.005. [DOI] [PubMed] [Google Scholar]
- 10.Yang SW, Shen YC, Chen CH. Steroids and triterpenoids of Antrodia cinnamomea—a fungus parasitic on Cinnamomum micranthum . Phytochemistry. 1996;41(5):1389–1392. [Google Scholar]
- 11.Shen YC, Yang SW, Lin CS, Chen CH, Kuo YH, Chen CF. Zhankuic acid F: a new metabolite from a formosan fungus Antrodia cinnamomea . Planta Medica. 1997;63(1):86–88. doi: 10.1055/s-2006-957614. [DOI] [PubMed] [Google Scholar]
- 12.Hseu YC, Chang WC, Hseu YT, et al. Protection of oxidative damage by aqueous extract from Antrodia camphorata mycelia in normal human erythrocytes. Life Sciences. 2002;71(4):469–482. doi: 10.1016/s0024-3205(02)01686-7. [DOI] [PubMed] [Google Scholar]
- 13.Hsiao G, Shen MY, Lin KH, et al. Antioxidative and hepatoprotective effects of Antrodia camphorata extract. Journal of Agricultural and Food Chemistry. 2003;51(11):3302–3308. doi: 10.1021/jf021159t. [DOI] [PubMed] [Google Scholar]
- 14.Geethangili MT, Tzeng YM. Review of pharmacological effects of Antrodia camphorata and its bioactive compounds. Evidence-Based Complementary and Alternative Medicine. 2011;2011:17 pages. doi: 10.1093/ecam/nep108.212641 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chen YY, Chang HH, Lee HY, Sheu F. Immunomodulatory and anti-tumor effects of oral administration with Antrodia cinnamomea fruiting bodies in BALB/c mice. Taiwanese Journal of Agricultural Chemistry and Food Science. 2008;46(2):87–95. [Google Scholar]
- 16.Cheng JJ, Huang NK, Chang TT, Wang DL, Lu MK. Study for anti-angiogenic activities of polysaccharides isolated from Antrodia cinnamomea in endothelial cells. Life Sciences. 2005;76(26):3029–3042. doi: 10.1016/j.lfs.2004.11.023. [DOI] [PubMed] [Google Scholar]
- 17.Yang CM, Zhou YJ, Wang RJ, Hu ML. Anti-angiogenic effecct and mecchanisms of polysaccharides from Antrodia cinnamomea wiith different molecular weights. Journal of Ethnopharmacology. 2009;123(3):407–412. doi: 10.1016/j.jep.2009.03.034. [DOI] [PubMed] [Google Scholar]
- 18.Cheng JJ, Lu MK, Lin CY, Chang CC. Characterization and functional elucidation of a fucosylated 1,6-α-d-mannogalactan polysaccharide from Antrodia cinnamomea . Carbohydrate Polymers. 2011;83(2):545–553. [Google Scholar]
- 19.Chen YJ, Chou CJ, Chang TT. Compound MMH01 possesses toxicity against human leukemia and pancreatic cancer cells. Toxicology In Vitro. 2009;23(3):418–424. doi: 10.1016/j.tiv.2009.01.011. [DOI] [PubMed] [Google Scholar]
- 20.Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. Colorimetric method for determination of sugars and related substances. Analytical Chemistry. 1956;28(3):350–356. [Google Scholar]
- 21.Satyapal US, Kadam VJ, Ghosh R. Hepatoprotective activity of livobond a polyherbal formulation against CCl4 induced hepatotoxicity in rats. International Journal of Pharmacology. 2008;4(6):472–476. [Google Scholar]
- 22.Limdi JK, Hyde GM. Evaluation of abnormal liver function tests. Postgraduate Medical Journal. 2003;79(932):307–312. doi: 10.1136/pmj.79.932.307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Johnston DE. Special considerations in interpreting liver function tests. The American Family Physician. 1999;59(8):2223–2230. [PubMed] [Google Scholar]
- 24.Reitman S, Frankel S. A colorimetric method for the determination of serum GOT and GPT. American Journal of Clinical Pathology. 1957;28:57–60. doi: 10.1093/ajcp/28.1.56. [DOI] [PubMed] [Google Scholar]
- 25.Plumb JA, Milroy R, Kaye SB. Effects of the pH dependence of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide-formazan absorption on chemosensitivity determined by a novel tetrazolium-based assay. Cancer Research. 1989;49(16):4435–4440. [PubMed] [Google Scholar]
- 26.Lin YM, Hu CP, Chou CK, et al. A new human hepatoma cell line: establishment and characterization. Chinese Journal of Microbiology and Immunology. 1982;15(3):193–201. [PubMed] [Google Scholar]
- 27.El Bahi S, Caliot E, Bens M, et al. Lymphoepithelial interactions trigger specific regulation of gene expression in the M cell-containing follicle-associated epithelium of Peyer’s patches. The Journal of Immunology. 2002;168(8):3713–3720. doi: 10.4049/jimmunol.168.8.3713. [DOI] [PubMed] [Google Scholar]
- 28.Teo EK, Fock KM. Hepatocellular carcinoma: an Asian perspective. Digestive Diseases. 2001;19(4):263–268. doi: 10.1159/000050692. [DOI] [PubMed] [Google Scholar]
- 29.Folkman J. Tumor angiogenesis: therapeutic implications. The New England Journal of Medicine. 1971;285(21):1182–1186. doi: 10.1056/NEJM197111182852108. [DOI] [PubMed] [Google Scholar]
- 30.Mc Cuskey RS, editor. The Hepatic Microvascular System. New York, NY, USA: Raven Press; 1994. [Google Scholar]
- 31.Ribatti D, Vacca A, Nico B, Sansonno D, Dammacco F. Angiogenesis and anti-angiogenesis in hepatocellular carcinoma. Cancer Treatment Reviews. 2006;32(6):437–444. doi: 10.1016/j.ctrv.2006.06.002. [DOI] [PubMed] [Google Scholar]
- 32.Shih CC, Lu MK, Cheng JJ, Danny LW. Antiangiogenic activities of polysaccharides isolated from medicinal fungi. FEMS Microbiology Letters. 2005;249(2):247–254. doi: 10.1016/j.femsle.2005.06.033. [DOI] [PubMed] [Google Scholar]
- 33.Shen CC, Kuo YC, Huang RY, Lin LC, Chou CJ. New ergostane and lanostane from Antrodia camphorata . The Journal of Chinese Medicine. 2003;14:247–258. [Google Scholar]