

Abstract
Background
Salmonella enterica serovar Typhimurium is a major foodborne pathogen worldwide. S. Typhimurium encodes type III secretion systems via Salmonella pathogenicity islands (SPI), producing the major effector proteins of virulence. Previously, we identified two genes of Salmonella pyruvate metabolism that were up-regulated during chicken cell infection: pyruvate formate lyase I (pflB) and bifunctional acetaldehyde-CoA/alcohol dehydrogenase (adhE). We were therefore interested in examining the role these genes may play in the transmission of Salmonella to humans.
Methods
Mutant strains of Salmonella with single gene deletions for pflB and adhE were created. Invasion and growth in human HCT-8 intestinal epithelial cells and THP-1 macrophages was examined. Quantitative PCR was performed on 19 SPI-1 genes.
Results
In HCT-8 cells, both mutant strains had significantly higher intracellular counts than the wild-type from 4 to 48 h post-infection. Various SPI-1 genes in the mutants were up-regulated over the wild-type as early as 1 h and lasting until 24 h post-infection. In THP-1 cells, no significant difference in internal Salmonella counts was observed; however, SPI-1 genes were largely down-regulated in the mutants during the time-course of infection. We also found five SPI-1 genes - hilA, hilC hilD, sicP and rtsA - which were up-regulated in at least one of the mutant strains in log-phase broth cultures alone. We have therefore identified a set of SPI-1 virulence genes whose regulation is effected by the central metabolism of Salmonella.
Keywords: adhE, Metabolism, pflB, Pyruvate, Salmonella pathogenicity island, Salmonella Typhimurium, Virulence
Background
Salmonella serovars cause an array of diseases, ranging from gastroenteritis to systemic infections [1,2]. While there are thousands of characterized serovars, few have been demonstrated as virulent to humans [3,4]. Salmonella enterica serovar Typhimurium is one such strain, with a broad spectrum of hosts including pig, chicken and humans [1,5,6]. S. Typhimurium can be ubiquitous in the environment and within certain animal reservoirs; it can act as a commensal, produce an asymptomatic infection, and/or trigger disease in various hosts [7]. Serovar Typhimurium thus remains one of the top infectious food-borne pathogens worldwide.
The most common route of entry for S. Typhimurium is by oral transmission. Once contact is made with a human host through this route, the bacterium initiates infection by invading intestinal epithelial cells using a type III secretion system (T3SS) [8-10]. This system is encoded by a 40 kilobase region of the genome called Salmonella pathogenicity island (SPI) 1 [11-18]. The T3SS of S. Typhimurium consists of structural components (i.e. needle complex) that make up the apparatus used to inject effector proteins into the cytoplasm of host cells [12,19-21]. The effector proteins then promote infection by altering the physiology of the host cell. Specifically, Salmonella can seize the host cell machinery and induce rearrangements in the actin cytoskeleton and cause alterations in signal transduction that helps facilitate further systemic infection [22-24]. In mammals, the intracellular phase of infection proceeds in a coordinated effort with an additional type III secretion system encoded by SPI-2 [25-27]. Overall, the T3SS-1 encoded by SPI-1 promotes invasion of host intestinal epithelial cells [28,29], initiation of inflammation [27,30,31], and intracellular survival and persistence [32-34], while TTSS-2 encoded by SPI-2 is needed to promote intracellular growth [35-39].
While the T3SS and its genetic regulators are a central component in the ability of Salmonella to establish a systemic infection, the capability of Salmonella to invade and persist in a host has been partly linked to the central metabolism of the bacterium (reviewed in [40]) and environmental cues [41-44]. Alterations in Salmonella metabolic pathways, such as glycolysis and the tricarboxylic acid (TCA) cycle, using both in vitro and in vivo models have demonstrated the importance of factors outside of the T3SS in its pathogenicity [40]. For instance, deletions in the pyruvate dehydrogenase subunit E1 (aceE) gene of S. Enteritidis has been shown to attenuate the strain in a chicken model [45]. In murine models, the loss of functional phosphofructokinase (Pfk) activity prevents growth of S. Typhimurium [46], while deletions in other TCA cycle genes hyper-activate the mutants in macrophages though interestingly attenuates the response in BALB/c mice [47]. As the SPI genes and the T3SS have been extensively studied as key pathogen modulators in Salmonella, determining the ability of additional metabolic genes to affect pathogenicity remains to be explored.
In our previous study, we measured global gene expression of the Salmonella enterica serotype Typhimurium genome in chicken heterophils (data not shown). Two Salmonella genes, pyruvate formate lyase I (pflB) and bifunctional acetaldehyde-CoA/alcohol dehydrogenase (adhE), were significantly up-regulated after infection. Since these genes are not typical virulence genes but play a role in cellular metabolism (Figure 1), we sought to understand their role during infection. Therefore, the overall aim of this study is to further characterize the importance these Salmonella genes play in its pathogenesis. Deletion mutants of these two genes were created, and gentamicin protection assays and quantitative PCR of SPI-1 genes were performed on human epithelial and macrophage cell lines. The resulting experiments show that these two genes of the pyruvate metabolic pathway affect both infectivity of epithelial cells and SPI-1 gene expression.
Figure 1.
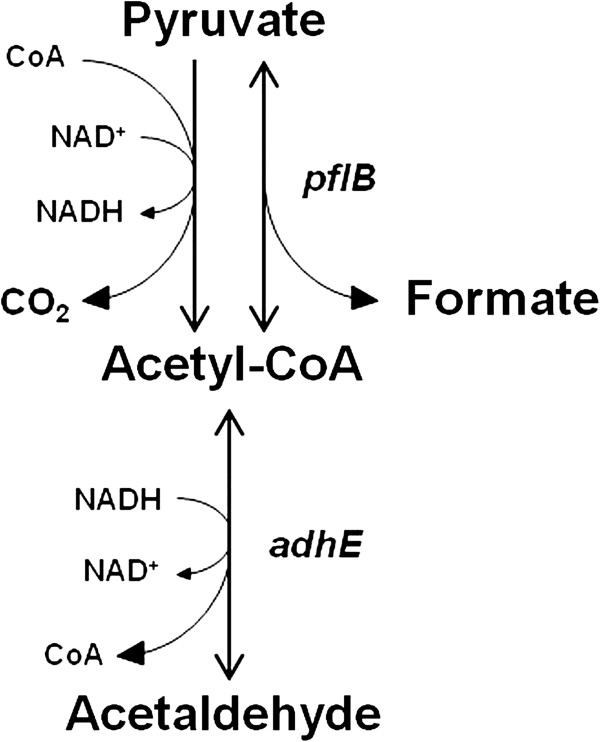
A simplified model of the pyruvate metabolism pathway in Salmonella Typhimurium. Genes producing metabolic enzymes (in italics) were deleted in the creation of mutant strains, including pyruvate formate lyase I (pflB) and bifunctional acetaldehyde-CoA/alcohol dehydrogenase (adhE).
Methods
Creation of Salmonella mutants
The Salmonella enterica serovar Typhimurium ATCC 14028 strain was used as the wild-type in this study. This strain was also used in the construction of mutant gene knock-out isolates. Two genes, pyruvate formate lyase I (pflB) and bifunctional acetaldehyde-CoA/alcohol dehydrogenase (adhE), produce enzymes of the pyruvate metabolic pathway as illustrated (Figure 1). To assess the effects of these genes during invasion, two mutant strains, ΔpflB::kan (S. Typhimurium gene number STM0973) and ΔadhE::kan (S. Typhimurium gene number STM1749), were created by the lambda red recombination method with adaptations as described by Santiviago et.al. [48]. Gene-specific primers used for the creation of each isolate are listed in Table 1. The recombination event was confirmed by polymerase chain reaction (PCR) using primers to the flanking genomic regions.
Table 1.
A list of the Salmonella primers used in this study
| Gene | Forward sequence (5′-3′) | Reverse sequence (5′-3′) | Locus or reference | Technique |
|---|---|---|---|---|
|
pflB |
GAAGGTAGGTGTTACATGTCCGAGCTT AATGAAAAGTTAGCCACAGTGCAGGCT GGAGCTGCTTC |
TTTCAGTCAAACCCATTACAT GGTCTGCGTGAAGGTACGAGT AATCATATGAATATCCTCCTTAG |
STM0973 |
Lambda red |
|
adhE |
TATCAGGAGAGCATTATGGCTGTTACT AATGTCGCTGAACTTAACGTGCAGGCT GGAGCTGCTTC |
AAAACAGATAACGAATTAAGC GGATTTTTTCGCTTTTTTCTC TGCCATATGAATATCCTCCTTAG |
STM1749 |
|
| 16S rRNA |
AGGCCTTCGGGTTGTAAAGT |
GTTAGCCGGTGCTTCTTCTG |
[49] |
qPCR |
|
hilA |
ATAGCAAACTCCCGACGATG |
ATTAAGGCGACAGAGCTGG |
[50] |
|
|
hilC |
CTCACCTCTTCAGCGGCCAGT |
CACCCGCAAATGGTCACAGGCT |
STM2867 |
|
|
hilD |
GGTAGTTAACGTGACGCTTG |
GATCTTCTGCGCTTTCTCTG |
[50] |
|
|
invC |
TCGGTCGATCGCTGCAA |
CCCTGGTCGCGAAAATATTC |
STM2894 |
|
|
invE |
CGAATGACGCCAGCTGTTC |
TGCGTCAGGCGTCGTAAA |
STM2897 |
|
|
invF |
TGAAAGCCGACACAATGAAAAT |
GCCTGCTCGCAAAAAAGC |
STM2899 |
|
|
invH |
GGTGCCCCTCCCTTCCT |
TGCGTTGGCCAGTTGCT |
STM2900 |
|
|
invJ |
CGCCGCCGGTTAATTG |
GCTTCCGCTGCAACCAA |
STM2892 |
|
|
orgA |
AGGCAGGGAGCCTTGCTT |
CCCTGATGCATTGCCAAAA |
STM2870 |
|
|
orgB |
ACCATCCCGAAACGCTTTTA |
TTGCCCCTCAGGCTTATCG |
STM2869 |
|
|
prgH |
TGAACGGCTGTGAGTTTCCA |
GCGCATCACTCTGACCTACCA |
STM2874 |
|
|
prgJ |
GGTCTATGGAAACGGACATTGTC |
CGCCGAACCAGAAAAAGC |
STM2872 |
|
|
prgK |
GGGTGGAAATAGCGCAGATG |
TCAGCTCGCGGAGACGATA |
STM2871 |
|
|
rtsA |
ACCCGTGGTGAGCTTGATGAGT |
CCTGTCCAGGTGGGGAGCAT |
SEE_03946 |
|
|
sicA |
GATGAGTCTCTGCGGGCAAA |
GCTCTGTCTCCGCCGTTTT |
STM2886 |
|
|
sicP |
AGATGATATCTGGTTATTGAACGGTATG |
CTGCCGCCAGATAGAATCG |
STM2879 |
|
|
sipA |
CGTCTTCGCCTCAGGAGAAT |
TGCCGGGCTCTTTCGTT |
STM2882 |
|
|
sipB |
GGCGCGCTGCTAACCAT |
TCGCCCCACCGGTAAAA |
STM2885 |
|
| sipC | ATCAGGCTGGTCGATTTACG | GTACGCCGCTACTCAGGAAC | [50] |
Cell culture
Human cell lines HCT-8 and THP-1 were used in this study. HCT-8 is an ileocecal colorectal adenocarcinoma epithelia cell line, while THP-1 is a monocytic leukemia cell line that can be stimulated to differentiate into macrophages. HCT-8 was grown using RPMI-1640 with 2.05 mmol/L L-Glutamine media (HyClone, Thermo Scientific, Rockford, IL) supplemented with 10% fetal bovine serum (Atlanta Biologicals, Lawrenceville, GA), 1% of 100 mmol/L sodium pyruvate (Thermo Scientific), and 1% of 10 000 U penicillin/10 mg streptomycin per mL (Thermo Scientific). THP-1 was grown in suspension using RPMI-1640 with 2.05 mmol/L L-Glutamine media (HyClone) supplemented with 10% fetal bovine serum (Atlanta Biologicals), 1% of 10,000 U penicillin/10 mg streptomycin per mL (Thermo Scientific), and 1.75 μL of 14.3 mM β-Mercaptoethanol (Sigma-Aldrich, St. Louis, MO). Both cell lines were cultured at 37°C in an air-jacketed incubator (NuAire, Plymouth, MN) with 5% CO2 and constant humidity.
Invasion assays
The infectivity of Salmonella on both HCT-8 and THP-1 was defined as the number of internal Salmonella cells per viable human cell at each unit of time. Internal Salmonella counts on both cell lines were determined using a gentamicin protection assay (GPA). Viable human cells were assayed by trypan blue exclusion. Invasion assays on each cell line were performed in at least three independent experiments, using triplicate wells at each time-point. Probabilities for statistical significance were calculated using Student’s t-test.
For HCT-8 cells, 2 × 105 cells were seeded in 12-well plates with 1 mL complete media (without antibiotics) and grown until confluent. Overnight ATCC 14028, ΔpflB::kan, and ΔadhE::kan strains of Salmonella grown at 37°C in Luria-Bertani (LB) broth were sub-cultured for 3 h in pre-warmed (37°C) LB broth before challenge to ensure log-phase cultures. HCT-8 cells were challenged with each Salmonella strain at a multiplicity of infection (MOI) of 100 by replacing media with Salmonella infectious media. Immediately upon challenge, plates were centrifuged at 500 × g for 5 min and placed in a CO2 incubator and allowed to incubate for 30 min. After this time, cells were washed with phosphate buffered saline (PBS, pH 7.4) and infectious media was replaced with 1 mL complete media supplemented with 100 μg/mL gentamicin sulfate (Mediatech, Manassas, VA). After 30 min, media was replaced with 2 mL complete media supplemented with 10 μg/mL gentamicin sulfate (Mediatech) and kept through the duration of the study. The first time-point of 1 h post-infection was collected by washing the infected HCT-8 cells with PBS and lysing with 1 mL PBS + 1% Triton X-100 (Sigma-Aldrich). Internalized Salmonella were then counted by serial dilutions on pre-warmed LB agar in triplicate. At the same time, infected HCT-8 cells were counted on a TC10 automated cell counter (Bio-Rad, Hercules, CA) using a 1:1 ratio of cells to 0.4% trypan blue dye (Bio-Rad). Additional time-points of 4, 18, 24, and 48 h post-infection were also collected.
For THP-1 cells, 8 × 105 THP-1 cells were seeded in 12-well plates with 1 mL complete media (without β-Mercaptoethanol or antibiotics) and 200 nmol/L phorbol 12-myristate 13-acetate (PMA; Millipore, Billerica, MA) for 24 h to allow differentiation of suspension monocytes to adherent macrophages. After 24 h, media was exchanged with pre-warmed media (without β-Mercaptoethanol or antibiotics) omitting PMA and cells were allowed to develop for an additional 24 h. Overnight cultures from ATCC 14028, ΔpflB::kan, and ΔadhE::kan strains of Salmonella were sub-cultured for 2.5 h in pre-warmed LB broth and then incubated with 10% human AB serum (Mediatech) for 30 min prior to challenge. THP-1 cells were challenged 48 h after seeding at a MOI10 from each Salmonella strain, and invasion was assessed of similarly as with HCT-8 challenge. To assess the role that opsonization may play on infectivity using these Salmonella strains, separate GPA experiments were also performed where strains were not incubated with human AB serum prior to challenge. Time-points of 1, 2.5, 4, and 6 h post-infection were collected from non-opsonized challenge experiments. Lower time-points and MOI were selected for non-opsonized challenge as the THP-1 cells were largely damaged and apopotic at MOI 100 and with longer incubations (data not shown), and has previously been observed [51].
Quantitative PCR
Real-time reverse-transcription quantitative PCR (RT-qPCR) was performed to assess mRNA expression levels of 19 selected SPI-1 genes. The primers used for RT-qPCR are listed in Table 1. SPI-1 transcripts were measured from both Salmonella grown in cultures and after invasion of host cells.
To generate complementary DNA (cDNA) for RT-qPCR from Salmonella cultures, 5 mL of ATCC14028, ΔpflB::kan, and ΔadhE::kan strains were grown overnight at 37°C with agitation, then sub-cultured 1:25 in pre-warmed LB broth for 4 h. The three strains were equalized by OD600 spectrophotometry and RNA was isolated by the RNeasy Protect Bacteria Kit (Qiagen, Valencia, CA) according to the manufacturer’s protocol. Potential contaminating genomic DNA was removed by the TURBO DNA-free™ kit (Ambion, Carlsbad, CA), as directed by the manufacturer. Reverse transcription reactions were then performed by the ThermoScript™ RT-PCR System for First-Strand cDNA Synthesis (Invitrogen, Carlsbad, CA) from 4 μg total RNA from each sample, using the supplied random hexamers as RT-primers, in 20 μL total volume reactions. Reactions for each sample were diluted 1:10 with sterile nanopure water and used as templates for RT-qPCR.
To generate cDNA for RT-qPCR from Salmonella after invasion of host cells, HCT-8 and THP-1 cells were again infected with ATCC 14028, ΔpflB::kan, or ΔadhE::kan strains by the GPA in additional independent experiments, with the following modifications. At each time-point post infection, cells were washed with PBS and lysed directly by the addition of 400 μL TRI-reagent (Ambion) to each well and gentle pipetting. Total RNA was then extracted using the RiboPure™-Bacteria Kit (Ambion) according to the manufacturer’s protocol. The TURBO DNA-free™ kit (Ambion) was used to remove potential genomic DNA contamination. Total RNA from each sample (representing one well of a 12-well plate) was quantified using a spectrophotometer (NanoDrop 1000, Wilmington, DE) and equimolar amounts from each of three replicate wells were pooled. RT-reactions were performed using the ThermoScript™ RT-PCR System for First-Strand cDNA Synthesis (Invitrogen) from 800 ng total RNA from each sample, using the supplied random hexamers as RT-primers, in 20 μL total volume reactions. Reactions for each sample were diluted 1:5 with sterile nanopure water and used as templates for RT-qPCR.
All RT-qPCR reactions were performed using the Power SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA), with modifications. A total reaction volume of 10 μL was used, to include: 5 μL of 2X SYBR Green, 2 μL of [1 μmol/L] forward primer, 2 μL of [1 μM] reverse primer, and 1 μL template cDNA. Quantitative PCR was performed on an ABI 7900HT system (Applied Biosystems), with cycling and dissociation curve parameters used according to the recommendations from the manufacturer. Each RT-qPCR reaction was performed using at least three technical replicates for each sample. After real-time PCR cycling, each product was assessed for quality and specificity by the corresponding dissociation curve. Raw data was edited using the SDS v2.4 Software (Applied Biosystems), and cycle threshold (Ct) values were recorded. Ct values were uploaded into the Relative Expression Software Tool (REST2009; Qiagen) [52] for comparisons. Data was considered significant at p-value < 0.05 after permutation testing using randomizations. The standard error of the mean was estimated from confidence intervals.
Results
Salmonella invasion of HCT-8 epithelial cells
After HCT-8 cells were challenged with either Salmonella wild-type or mutant strains, the number of intracellular bacteria as assessed by counting colony forming units (CFUs) on LB agar were averaged from repeated independent GPA experiments. At the earliest time post-infection of 1 h, there was no significant difference in average CFUs between the wild-type and mutant strains (Figure 2A). However, by 4 h, there was a significant increase in average CFUs in the mutant strains, ΔpflB::kan and ΔadhE::kan, compared to the wild-type strain, ATCC 14028 (Figure 2A). At 4 h, there were approximately 4–5 CFU of either mutant per 10 HCT-8 cells, while ATCC 14028 remained near 1 CFU/10 HCT-8 cells, thus representing an approximate 4–5 fold increase in Salmonella mutants over the wild-type. Increased numbers of mutant strains over the wild-type observed at 4 h post-infection also lasted through the duration of the study at 48 h post-infection, measured at increments of 18, 24, and 48 h (Figure 2B). At 18 h post-infection, there were 4.2 and 3.9 CFU per HCT-8 cell from ΔpflB::kan and ΔadhE::kan strains, respectively, while the wild-type ATCC 14028 strain remained at near 1 CFU/cell, thus representing an approximate 4-fold significant difference. Similarly at 24 h post-infection, there were 3.9 and 4.3 CFU per viable epithelial cell from ΔpflB::kan and ΔadhE::kan strains, respectively, while the wild-type again remained around 1 CFU/HCT-8 cell at this time-point. The wild-type strain ATCC 14028 was mostly consistent in the level of infectivity and growth measured by GPA with an average of 6.4 CFU per 10 HCT-8 cells over the 48 h course of study, ranging from 3 CFU/100 HCT-8 cells at 1 h to 1.3 CFU/HCT-8 cell at 48 h post-infection. However, in the mutant strains there was an approximate 8-fold significant increase in the number of CFUs between the 1 h and 4 h time-points and, likewise, an approximate 8-fold increase between the 4 h and 18 h time-points. Average CFU/HCT-8 cell was also increased in the mutants, with an average of 3.9 and 2.5 CFU/HCT-8 cell in ΔpflB::kan and ΔadhE::kan, respectively. CFUs of both mutant strains remained consistent between 18 h and 24 h post-infection; however, at 48 h post-infection a spike in ΔpflB::kan was observed (Figure 2B). At 4 h through 48 h post-infection, internal growth of the mutants was found to be highly significant (p-value < 0.01). Overall there was either a steady or an upward trend in the numbers of internalized Salmonella through the 48 h course of infection in epithelial cells with the magnitude of replication being significantly greater by the mutant strains beginning a 4 h post-infection (Figure 2).
Figure 2.

Invasion of HCT-8 cells by Salmonella Typhimurium strains. (A) Time-points of 1 h and 4 h post-infection; (B) Time-points of 18 h, 24 h, and 48 h post-infection. The X-axis represents time (h) while the Y-axis represents average colony forming units (CFU) of bacteria per viable HCT-8 cell. The different Salmonella strains are color-coded according to the legend. Asterisk indicates significance of the mutant strain from the wild-type at p < 0.01 assessed at each time.
Salmonella invasion of THP-1 macrophage cells
For challenge of THP-1 cells with Salmonella strains opsonized by treatment with human AB serum, time increments were chosen the same as with HCT-8 challenge. Again, average numbers of internalized Salmonella strains were assessed by counting CFUs. The numbers of internalized Salmonella ranged from approximately 1.5 to 21 CFU per viable THP-1 cell (Figure 3). ATCC 14028 wild-type strain had an average of 7.4 CFU/THP-1 cell, ΔpflB::kan strain had an average of 11.4 CFU/THP-1 cell, and ΔadhE::kan strain had an average of 9.9 CFU/THP-1 cell over the course of the 48 h study. There was a general trend that the mutant strains, in particular ΔpflB::kan, had higher CFUs per differentiated THP-1 cell than ATCC 14028. However, no significant difference in average CFU counts was observed between either of the mutant or wild-type strains after repeated GPA experiments. This was the case at all of the five time-points tested, from 1 h to 48 h post-infection (Figure 3). Overall, there was a bell-curved trend in the numbers of internalized Salmonella throughout this study, where bacteria peaked at 18 h post-infection, and then began to decline as the viable THP-1 cells declined (Figure 3).
Figure 3.

Invasion of THP-1 cells by opsonized Salmonella Typhimurium strains. Time-points of 1, 4, 18, 24, and 48 h post-infection are shown. The X-axis represents time (h) while the Y-axis represents average colony forming units (CFU) of bacteria per viable THP-1 cell. The different Salmonella strains are color-coded according to the legend.
For challenge of THP-1 cells with non-opsonized Salmonella strains, time-points of 1, 2.5, 4, and 6 h post-infection were selected to assess infectivity. The numbers of internalized Salmonella cells ranged from approximately 1.4 to 10.4 CFU per viable THP-1 cell (Figure 4). ATCC 14028 wild-type strain had an average of 3.3 CFU/THP-1 cell, ΔpflB::kan strain had an average of 5.4 CFU/THP-1 cell, and ΔadhE::kan strain had an average of 4.1 CFU/THP-1 cell over the course of the 6 h study. There was a general trend that the mutant strains, in particular ΔpflB::kan, had higher CFUs per differentiated THP-1 cell than ATCC 14028. Again however, no significant difference in average CFU counts was observed between either of the mutant or wild-type strains after repeated GPA experiments. This was the observation at any of the four time-points tested, from 1 h to 6 h post-infection (Figure 4). Overall, there was an upward trend in the numbers of internalized Salmonella throughout this study as viable THP-1 cells had not yet become noticeably apoptotic (Figure 3).
Figure 4.

Invasion of THP-1 cells by non-opsonized Salmonella Typhimurium strains. Time-points of 1, 2.5, 4, and 6 h post-infection are shown. The X-axis represents time (h) while the Y-axis represents average colony forming units (CFU) of bacteria per viable THP-1 cell. The different Salmonella strains are color-coded according to the legend.
After the THP-1 GPA trials, a direct comparison was made between 1 h and 4 h post-infection to assess the role opsonization may play between the Salmonella parent and mutant strains. For the opsonized challenge, ATCC 14028 had an average internalization of 7.3 and 7.2 CFU/THP-1 cell at 1 h and 4 h, respectively. ΔpflB::kan had an average internalization of 12.4 and 12.8 CFU/THP-1 cell at 1 h and 4 h, respectively. ΔadhE::kan had an average internalization of 7.7 and 10.6 CFU/THP-1 cell at 1 h and 4 h, respectively (Figure 3). For the non-opsonized challenge, ATCC 14028 had an average internalization of 1.4 and 3.6 CFU/THP-1 cell at 1 h and 4 h, respectively. ΔpflB::kan had an average internalization of 1.9 and 5.4 CFU/THP-1 cell at 1 h and 4 h, respectively. ΔadhE::kan had an average internalization of 1.4 and 4.0 CFU/THP-1 cell at 1 h and 4 h, respectively (Figure 4). Thus, larger average CFU counts were observed by challenge of THP-1 cells with opsonized bacteria.
SPI-1 gene expression in culture
To determine if the significant increase in numbers of Salmonella mutants to HCT-8 cells was potentially due to the type III secretion system, gene expression was measured in 19 SPI-1 genes (Table 1) by qPCR and compared to the expression in the wild-type strain. Differential regulation was measured from SPI-1 transcripts in log-phase cultures. A total of five genes were found significantly up-regulated in at least one of the mutant strains (Figure 5). In the ΔpflB::kan strain, hilA, hilD, rtsA, and sicP genes were up-regulated compared to ATCC 14028. Relative expression values ranged from 1.36 to 1.96 over the wild-type in genes found significant by qPCR (p < 0.05). In the ΔadhE::kan strain, hilA, hilC, hilD, and sicP genes were up-regulated compared to ATCC 14028. Relative expression values ranged from 1.64 to 2.58 over the wild-type in genes found significant by qPCR (Figure 5).
Figure 5.

Quantitative RT-PCR of Salmonella Typhimurium strains from cultures. Gene expression of select SPI-1 invasion genes measured by RT-qPCR from Salmonella mutants ΔpflB::kan and ΔadhE::kan and comparing expression to the wild-type strain. The 16S rRNA gene was used for normalization. Genes are listed along the X-axis while relative expression level is listed along the Y-axis. Asterisk indicates significance of the mutant strain from the wild-type at p < 0.05 assessed for each gene.
SPI-1 gene expression after invasion of HCT-8 epithelial cell line
Since a set of SPI-1 genes were up-regulated in cultures alone, we were interested to determine if a larger response involving a greater number of genes or a greater magnitude of the fold-change was observed through interaction with epithelial cells. The response of SPI-1 after infection with HCT-8 cells was determined by using qPCR on the same panel of 19 genes. SPI-1 genes were found to be up-regulated over the wild-type as early as 1 h post-infection (Table 2).
Table 2.
Quantitative RT-PCR of Salmonella during invasion of HCT-8 cells
| Time post-infection | Strain | Salmonella gene | Expression* | Regulation** |
|---|---|---|---|---|
| 1 h |
ΔpflB::kan |
hilC |
1.28 |
UP |
|
hilD |
1.60 |
|||
|
prgJ |
1.48 |
|||
|
prgK |
1.92 |
|||
|
sicA |
1.31 |
|||
| ΔadhE::kan |
hilC |
1.26 |
||
|
invE |
3.16 |
|||
|
invH |
1.56 |
|||
|
orgB |
1.25 |
|||
|
prgJ |
1.60 |
|||
|
prgK |
1.96 |
|||
|
sicA |
1.15 |
|||
| 4 h |
ΔpflB::kan |
rtsA |
2.52 |
UP |
|
sicA |
1.64 |
|
||
|
sipA |
1.88 |
|
||
| ΔadhE::kan |
hilA |
3.12 |
|
|
|
orgA |
3.70 |
|
||
|
orgB |
1.66 |
|
||
|
rtsA |
2.47 |
|||
|
sicA |
1.67 |
|
||
|
sicP |
1.56 |
|
||
|
sipA |
1.62 |
|
||
| 18 h |
ΔpflB::kan |
hilD |
2.15 |
UP |
|
prgH |
1.85 |
|||
|
prgJ |
3.08 |
|||
|
sicA |
2.78 |
|||
|
sipA |
1.88 |
|||
|
sipC |
3.21 |
|||
| ΔadhE::kan |
invC |
1.61 |
||
| 24 h | ΔpflB::kan |
prgH |
−1.92 |
DOWN |
|
prgJ |
−2.43 |
|||
|
prgK |
−1.75 |
|||
|
sicP |
−3.41 |
|||
|
sipA |
−3.36 |
|||
|
sipB |
−3.39 |
|||
| ΔadhE::kan |
invC |
−2.66 |
||
| sipA | −2.07 |
*Values listed are relative fold-change values as compared to the wild-type strain ATCC 14028; **All genes listed are significant at p-value < 0.05.
In ΔpflB::kan, five SPI-1 genes were up-regulated at 1 h, ranging from 1.28 to 1.92 fold change over the wild-type. At 4 h, three SPI-1 genes were up-regulated with relative expression values ranging from 1.64 to 2.52 fold over the wild-type. At 18 h post-infection, six SPI-1 genes were found to be up-regulated, with fold changes of 1.85 to 3.21 over the wild-type. In ΔadhE::kan, seven SPI-1 genes were up-regulated at 1 h post-infection, with relative expression values ranging from 1.15 to 3.16 fold over the wild-type. At 4 h, seven SPI-1 genes were up-regulated with relative expression values ranging from 1.62 to 3.70 fold over the wild-type. At 18 h post-infection, invC was up-regulated 1.61 fold in ΔadhE::kan over ATCC 14028. By 24 h post-infection, SPI-1 genes were either down-regulated in both mutant strains, or not significantly different from the wild-type strain.
SPI-1 gene expression after invasion of THP-1 macrophage cell line
Salmonella invasion gene expression from bacteria invaded into differentiated macrophages, THP-1 cell line, was also examined for non-opsonized wild-type and mutant strains. Similarly to non-opsonized Salmonella GPA, time-points of 1, 2.5, 4, and 6 h post-infection were chosen for testing of SPI-1 gene expression by qPCR.
In strain ΔpflB::kan, several SPI-1 genes were differentially expressed between the mutant and wild-type (Table 3). At 1 h, there was no significant differential gene expression. By 2.5 h, 6 SPI-1 genes were down-regulated with expression values ranging from −1.34 to −1.70, while invE was up-regulated 1.7-fold over the wild-type at the same time. At 4 h, hilA, hilD, invF, and prgJ were slightly up-regulated, with expression values of 1.18 to 1.72 fold over the wild-type. The largest degree of differential expression was observed at 6 h post-infection. A total of 17 out of the 19 SPI-1 genes tested (89%) were down-regulated, ranging from −1.3 to −2 fold change over the wild-type.
Table 3.
Quantitative RT-PCR of non-opsonized Salmonella strain ΔpflB::kan during invasion of THP-1 cells
| Time post-infection | Salmonella gene | Expression* | Regulation** |
|---|---|---|---|
| 1 h |
No change |
|
|
| 2.5 h |
invE |
1.71 |
UP |
|
invC |
−1.46 |
DOWN |
|
|
invH |
−1.41 |
||
|
rtsA |
−1.70 |
||
|
sicA |
−1.52 |
||
|
sicP |
−1.34 |
||
|
sipC |
−1.36 |
||
| 4 h |
hilA |
1.72 |
UP |
|
hilD |
1.36 |
||
|
invF |
1.19 |
|
|
|
prgJ |
1.18 |
|
|
| 6 h |
hilA |
−1.61 |
DOWN |
|
hilC |
−1.97 |
||
|
hilD |
−1.55 |
||
|
invC |
−1.54 |
||
|
invE |
−1.51 |
||
|
invF |
−1.52 |
||
|
invH |
−1.61 |
||
|
orgA |
−1.66 |
||
|
orgB |
−1.30 |
||
|
prgH |
−1.43 |
||
|
prgJ |
−1.46 |
||
|
rtsA |
−1.83 |
||
|
sicA |
−1.89 |
||
|
sicP |
−1.63 |
||
|
sipA |
−1.58 |
||
|
sipB |
−2.00 |
||
| sipC | −1.70 |
*Values listed are relative fold-change values as compared to the wild-type strain ATCC 14028; **All genes listed are significant at p-value < 0.05.
In strain ΔadhE::kan, SPI-1 gene expression was down-regulated at every time-point examined (Table 4). At 1 and 2.5 h post-infection, all genes tested were significantly down-regulated as compared to ATCC 14028. At 4 h post-infection, 14 of the 19 SPI-1 genes tested (74%) were down-regulated, with prgJ having the lowest magnitude of the fold-change at −1.67 and sipB having the highest magnitude of the fold-change at −4.44 under the wild-type. At 6 h post-infection, 16 of the 19 SPI-1 genes tested (84%) were down-regulated, with orgB having the lowest magnitude of the fold-change at −1.31 and sipB having the highest magnitude of the fold-change at −3.58 under the wild-type.
Table 4.
Quantitative RT-PCR of non-opsonized Salmonella strain ΔadhE:kan during invasion of THP-1 cells
| Time post-infection | Salmonella gene | Expression* | Regulation** |
|---|---|---|---|
| 1 h |
All genes tested were significantly down-regulated |
||
| 2.5 h |
All genes tested were significantly down-regulated |
||
| 4 h |
hilC |
−2.53 |
DOWN |
|
invC |
−2.63 |
||
|
invE |
−2.46 |
||
|
invF |
−2.14 |
||
|
invH |
−1.98 |
||
|
orgB |
−1.71 |
||
|
prgH |
−1.79 |
||
|
prgJ |
−1.67 |
||
|
prgK |
−1.91 |
||
|
rtsA |
−2.58 |
||
|
sicA |
−3.91 |
||
|
sicP |
−1.73 |
||
|
sipB |
−4.44 |
||
|
sipC |
−2.91 |
||
| 6 h |
hilA |
−2.08 |
DOWN |
|
hilC |
−2.76 |
||
|
hilD |
−1.56 |
||
|
invC |
−2.76 |
||
|
invE |
−2.07 |
||
|
invF |
−2.44 |
||
|
invH |
−2.30 |
||
|
orgA |
−2.36 |
||
|
orgB |
−1.31 |
||
|
prgH |
−1.71 |
||
|
rtsA |
−2.33 |
||
|
sicA |
−3.31 |
||
|
sicP |
−1.72 |
||
|
sipA |
−2.22 |
||
|
sipB |
−3.58 |
||
| sipC | −2.28 | ||
*Values listed are relative fold-change values as compared to the wild-type strain ATCC 14028; **All genes listed are significant at p-value < 0.05.
Discussion
SPI-1 of Salmonella enterica serovars is an important virulence factor that has been well-studied in several animal models and in disease outbreaks [53,54]. SPI-1-mediated infectivity of Salmonella Typhimurium was examined by creation of null mutations in genes that were up-regulated in our previous host-pathogen microarray study (data not shown). Those Salmonella genes, pyruvate formate lyase I (pflB) and bifunctional acetaldehyde-CoA/alcohol dehydrogenase (adhE), play a major role in the pyruvate metabolism of the organism (Figure 1). Here we show that these two genes can also affect virulence and SPI-1 gene expression.
For the HCT-8 intestinal epithelial cell line, internalized mutant Salmonella CFUs were significantly increased over the wild-type at 4 h post-infection and lasted through 48 h post-infection at which time the experiment was terminated (Figure 2). Since the T3SS encoded by SPI-1 is canonically involved in early contact with non-phagocytic cells such as fibroblasts and intestinal epithelia [27,30,31], a selected group of SPI-1 genes (Table 1) in mutant Salmonella were measured by qPCR and compared to the wild-type expression. Several SPI-1 genes were up-regulated after HCT-8 cell challenge, from 1 h post-infection lasting to 24 h post-infection when SPI-1 expression in the mutants decreased compared to the wild-type (Table 2). These up-regulated genes included hilC, hilD, rtsA, sipA, - all major genes involved in the early response to infection [55]. Some SPI-1 genes were up-regulated at multiple times post-infection and in both mutants, such as sicA, a gene encoding a complex chaperone protein required for Salmonella entry into host cells [56,57]. Overall, the results indicated that increased internalized growth of strains of S. Typhimurium deficient for transcripts of the pyruvate pathway in HCT-8 cells may in part be due to a differential regulation of genes in the T3SS.
Additional evidence in supporting this notion included qPCR results on SPI-1 genes in log-phase cultures alone. We found that the major regulators of SPI-1 were up-regulated when compared to the wild-type. These included hilA and hilD in both of the mutant strains, and hilC and rtsA in at least one of the mutant strains (Figure 5). HilA is the major regulator of SPI-1 invasion [41]. The hilA gene encodes an OmpR/ToxR family transcriptional regulator that activates the expression of invasion genes in response to both environmental and genetic factors [41]. Further, the proteins encoded by rtsA, hilC and hilD bind to a DNA region upstream of hilA and induce hilA expression [58-60]. Therefore, these genes could be expected to be induced at an early stage, as they are key modulators of invasive Salmonella infection and hilD appears to bridge communication between SPI-1 and SPI-2 regulation (reviewed in [55]). These results further indicate the increased virulence of the mutant strains to HCT-8 is occurring via a differential regulation of genes involved in the T3SS. Interestingly, the sicP gene was also up-regulated in both mutant strains measured from cell culture alone. SicP serves as a chaperone for the SPI-1 effector Salmonella protein tyrosine phosphatase (SptP). SptP has been shown to have several functions in the establishment of Salmonella infection, in both epithelial cells and macrophages [61,62]. While the gene expression of SptP was not examined, the up-regulation of the chaperone sicP transcripts could indicate early formation of SicP-SptP complexes even before contact with a host, as these two genes appear to be transcriptionally coupled [63]. The full significance of the gene expression response of sicP to alterations in the pyruvate metabolism pathway remains to be further explored.
We further examined the effects of the mutant strains on the infectivity of THP-1 macrophage cell line. We observed no significant difference in the internalization of Salmonella mutants over the wild-type strain, whether the invasion assays were using opsonized or non-opsonized bacteria (Figures 2 &3). Since we were interested in addressing the effects of our mutations on SPI-1-dependent virulence, we chose to examine SPI-1 gene expression using invasion assays with non-opsonized bacteria. Interestingly, while no significant internalization between strains was observed, SPI-1 gene expression was mostly down-regulated in the mutants after a challenge of THP-1 cells (Tables 3 &4). During the systemic phase of macrophage infection by Salmonella, SPI-1 expression is typically repressed to allow internal replication of the bacterium and suppress excess apoptosis [64]. Therefore, as differentiated THP-1 are phagocytic-type cells and infection is largely established via SPI2-encoded T3SS, low SPI-1 expression of invaded macrophages could therefore be expected based on previous efforts [65-70].
The specific Salmonella knockout genes and the role they may play in its pathogenesis were further examined. Our data supports the overall results of Huang et al. [50] that a pflB mutation in Salmonella can effect SPI-1 gene expression as measured from cultures alone. In addition to the previous work [50], we identified multiple SPI-1 genes that are differentially expressed both in cultures and at several time-points post-infection of an in vitro challenge using human HCT-8 cells. Knockout of the pflB gene likely produces an imbalance of metabolites in which the presence mimics the conditions in the host gastrointestinal tract [50]. We further show that another gene of the Salmonella pyruvate pathway (adhE) can have a similar effect on intestinal epithelia, suggesting that gut metabolites in addition to formate and/or other feedback mechanisms due to environmental cues may be at work. As AdhE is a multifunctional enzyme, another possibility for increased invasion of a ΔadhE mutant in addition to an SPI1-mediated virulence mechanism could be formulated. A lack of AdhE appears to stimulate type I fimbrial adhesion [71,72], important adhesions on the surface of Salmonella that may mediate attachment and internalization of bacteria [73]. Further experimentation would be warranted to determine the exact mechanism and co-factors involved with the pyruvate pathway involvement in Salmonella virulence.
For the generation of mutant strains, each recombination event was confirmed by PCR using primers to the flanking genomic regions. These confirmations, in addition to the evidence provided in this study, give confidence in the specificity of our deletions. By using lambda-red for site-specific mutagenesis, evidence suggests any potential downstream effects are reduced or eliminated with this system over that of random mutagenesis/transposons, including issues of polarity (e.g. [48]). We do acknowledge however that complementation assays to restore the phenotype could provide additional evidence, but these were not performed here.
We also acknowledge the role the host microbiome plays in Salmonella pathogenicity [74], particularly since microbiota may contribute to pyruvate pathway metabolites utilized in the establishment of infection [50]. The mammalian gastrointestinal tract contains high levels of short chain fatty acids as the result of the breakdown of foodstuffs by the digestive processes and the action of resident bacteria [75]. We therefore would be highly interested in future in vivo studies to help determine the mechanisms involved in the interplay between the host genetics, the gut environment, and the microbiome on Salmonella pathogenesis when dealing with mutant strains produced through metabolic pathway mutations.
Conclusions
Deletions of two genes, pflB and adhE, involved in the pyruvate metabolism pathway of Salmonella Typhimurium were shown to alter SPI-1 gene expression. In bacterial cell culture alone, five SPI-1 genes from the two mutant strains were up-regulated from the wild-type. In a challenge with human HCT-8 and THP-1 cell lines, SPI-1 gene expression was also significantly altered from the wild-type at various times post-infection. We therefore have identified a set of SPI-1 genes that crosstalk between Salmonella pyruvate metabolism and virulence. This work provides a further framework for examining the role that central metabolism plays in the infection of Salmonella Typhimurium.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
JA and XL designed assays and performed the necessary experiments to generate Salmonella mutants. JA, CC and CH performed cell culture and gene expression experiments. HZ conceived, planned and guided this study. JA and HZ drafted the manuscript. All authors read and approved the final manuscript.
Contributor Information
Jason Abernathy, Email: jwabernathy@ucdavis.edu.
Carolina Corkill, Email: ccorkil@tamu.edu.
Carolee Hinojosa, Email: caroleehinojosa@gmail.com.
Xianyao Li, Email: lxylly@gmail.com.
Huaijun Zhou, Email: hzhou@ucdavis.edu.
Acknowledgments
The authors would like to thank Dr. Helene Andrews-Polymenis at Texas A&M University for helpful discussions and technical assistance in the generation of Salmonella mutants. The authors also thank Drs. Palmy Jesudhasan and Suresh Pillai at Texas A&M University for use of equipment and providing some of the Salmonella qPCR primers used in this study. This work was supported in part by a USDA NIFA Postdoctoral Fellowship Grant (Award No. 2011-67012-30684).
References
- Ellermeier C, Slauch J. In: The Prokaryotes. Dworkin M, Falkow S, Rosenberg E, Schleifer K-H, Stackebrandt E, editor. New York: Springer; 2006. The Genus Salmonella; pp. 123–158. [Google Scholar]
- Raffatellu M, Tukel C, Chessa D, Wilson RP, Baumler AJ. In: Salmonella: Molecular Biology and Pathogenesis. Rhen M, Maskell D, Mastroeni P, Threlfall J, editor. Norfolk UK: Taylor & Francis; 2007. The Intestinal Phase of Salmonella Infections; pp. 31–52. [Google Scholar]
- Litrup E, Torpdahl M, Malorny B, Huehn S, Christensen H, Nielsen EM. Association between phylogeny, virulence potential and serovars of Salmonella enterica. Infect Genet Evol. 2010;10(7):1132–1139. doi: 10.1016/j.meegid.2010.07.015. [DOI] [PubMed] [Google Scholar]
- Brenner FW, Villar RG, Angulo FJ, Tauxe R, Swaminathan B. Salmonella nomenclature. J Clin Microbiol. 2000;38(7):2465–2467. doi: 10.1128/jcm.38.7.2465-2467.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fedorka-Cray PJ, Whipp SC, Isaacson RE, Nord N, Lager K. Transmission of Salmonella typhimurium to swine. Vet Microbiol. 1994;41(4):333–344. doi: 10.1016/0378-1135(94)90029-9. [DOI] [PubMed] [Google Scholar]
- Harvey PC, Watson M, Hulme S, Jones MA, Lovell M, Berchieri A Jr, Young J, Bumstead N, Barrow P. Salmonella enterica serovar typhimurium colonizing the lumen of the chicken intestine grows slowly and upregulates a unique set of virulence and metabolism genes. Infect Immun. 2011;79(10):4105–4121. doi: 10.1128/IAI.01390-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callaway T, Edrington T, Anderson R, Byrd J, Nisbet D. Gastrointestinal microbial ecology and the safety of our food supply as related to Salmonella. J Anim Sci. 2008;86(14 suppl):E163–E172. doi: 10.2527/jas.2007-0457. [DOI] [PubMed] [Google Scholar]
- Hansen-Wester I, Hensel M. Salmonella pathogenicity islands encoding type III secretion systems. Microbes Infect. 2001;3(7):549–559. doi: 10.1016/s1286-4579(01)01411-3. [DOI] [PubMed] [Google Scholar]
- Shea JE, Hensel M, Gleeson C, Holden DW. Identification of a virulence locus encoding a second type III secretion system in Salmonella typhimurium. Proc Natl Acad Sci USA. 1996;93(6):2593–2597. doi: 10.1073/pnas.93.6.2593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellermeier JR, Slauch JM. Adaptation to the host environment: regulation of the SPI1 type III secretion system in Salmonella enterica serovar Typhimurium. Curr Opin Microbiol. 2007;10(1):24–29. doi: 10.1016/j.mib.2006.12.002. [DOI] [PubMed] [Google Scholar]
- Morgan E. In: Salmonella: Molecular Biology and Pathogenesis. Rhen M, Maskell D, Mastroeni P, Threlfall J, editor. Norfolk UK: Taylor & Francis; 2007. Salmonella Pathogenicity Islands; pp. 67–88. [Google Scholar]
- Altier C. Genetic and environmental control of Salmonella invasion. J Microbiol. 2005;43(Spec No):85–92. [PubMed] [Google Scholar]
- Kimbrough TG, Miller SI. Contribution of Salmonella typhimurium type III secretion components to needle complex formation. Proc Natl Acad Sci USA. 2000;97(20):11008–11013. doi: 10.1073/pnas.200209497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimbrough TG, Miller SI. Assembly of the type III secretion needle complex of Salmonella typhimurium. Microbes Infect. 2002;4(1):75–82. doi: 10.1016/s1286-4579(01)01512-x. [DOI] [PubMed] [Google Scholar]
- Kubori T, Matsushima Y, Nakamura D, Uralil J, Lara-Tejero M, Sukhan A, Galan JE, Aizawa SI. Supramolecular structure of the Salmonella typhimurium type III protein secretion system. Science. 1998;280(5363):602–605. doi: 10.1126/science.280.5363.602. [DOI] [PubMed] [Google Scholar]
- Lee CA, Jones BD, Falkow S. Identification of a Salmonella typhimurium invasion locus by selection for hyperinvasive mutants. Proc Natl Acad Sci USA. 1992;89(5):1847–1851. doi: 10.1073/pnas.89.5.1847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mills DM, Bajaj V, Lee CA. A 40 kb chromosomal fragment encoding Salmonella typhimurium invasion genes is absent from the corresponding region of the Escherichia coli K-12 chromosome. Mol Microbiol. 1995;15(4):749–759. doi: 10.1111/j.1365-2958.1995.tb02382.x. [DOI] [PubMed] [Google Scholar]
- Sukhan A, Kubori T, Wilson J, Galan JE. Genetic analysis of assembly of the Salmonella enterica serovar Typhimurium type III secretion-associated needle complex. J Bacteriol. 2001;183(4):1159–1167. doi: 10.1128/JB.183.4.1159-1167.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collazo CM, Galan JE. Requirement for exported proteins in secretion through the invasion-associated type III system of Salmonella typhimurium. Infect Immun. 1996;64(9):3524–3531. doi: 10.1128/iai.64.9.3524-3531.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collazo CM, Galan JE. The invasion-associated type III system of Salmonella typhimurium directs the translocation of Sip proteins into the host cell. Mol Microbiol. 1997;24(4):747–756. doi: 10.1046/j.1365-2958.1997.3781740.x. [DOI] [PubMed] [Google Scholar]
- Cornelis GR. The type III secretion injectisome. Nat Rev Microbiol. 2006;4(11):811–825. doi: 10.1038/nrmicro1526. [DOI] [PubMed] [Google Scholar]
- Guiney DG, Lesnick M. Targeting of the actin cytoskeleton during infection by Salmonella strains. Clin Immunol. 2005;114(3):248–255. doi: 10.1016/j.clim.2004.07.014. [DOI] [PubMed] [Google Scholar]
- Galan JE. Salmonella interactions with host cells: type III secretion at work. Annu Rev Cell Dev Biol. 2001;17:53–86. doi: 10.1146/annurev.cellbio.17.1.53. [DOI] [PubMed] [Google Scholar]
- Abrahams GL, Hensel M. Manipulating cellular transport and immune responses: dynamic interactions between intracellular Salmonella enterica and its host cells. Cell Microbiol. 2006;8(5):728–737. doi: 10.1111/j.1462-5822.2006.00706.x. [DOI] [PubMed] [Google Scholar]
- McLaughlin LM, Govoni GR, Gerke C, Gopinath S, Peng K, Laidlaw G, Chien YH, Jeong HW, Li Z, Brown MD. The Salmonella SPI2 effector SseI mediates long-term systemic infection by modulating host cell migration. PLoS Pathog. 2009;5(11):e1000671. doi: 10.1371/journal.ppat.1000671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coombes BK, Brown NF, Valdez Y, Brumell JH, Finlay BB. Expression and secretion of Salmonella pathogenicity island-2 virulence genes in response to acidification exhibit differential requirements of a functional type III secretion apparatus and SsaL. J Biol Chem. 2004;279(48):49804–49815. doi: 10.1074/jbc.M404299200. [DOI] [PubMed] [Google Scholar]
- Hapfelmeier S, Stecher B, Barthel M, Kremer M, Muller AJ, Heikenwalder M, Stallmach T, Hensel M, Pfeffer K, Akira S. The Salmonella pathogenicity island (SPI)-2 and SPI-1 type III secretion systems allow Salmonella serovar typhimurium to trigger colitis via MyD88-dependent and MyD88-independent mechanisms. J Immunol. 2005;174(3):1675–1685. doi: 10.4049/jimmunol.174.3.1675. [DOI] [PubMed] [Google Scholar]
- Lostroh CP, Lee CA. The Salmonella pathogenicity island-1 type III secretion system. Microbes Infect. 2001;3(14–15):1281–1291. doi: 10.1016/s1286-4579(01)01488-5. [DOI] [PubMed] [Google Scholar]
- Schlumberger MC, Hardt WD. Salmonella type III secretion effectors: pulling the host cell’s strings. Curr Opin Microbiol. 2006;9(1):46–54. doi: 10.1016/j.mib.2005.12.006. [DOI] [PubMed] [Google Scholar]
- Coombes BK, Coburn BA, Potter AA, Gomis S, Mirakhur K, Li Y, Finlay BB. Analysis of the contribution of Salmonella pathogenicity islands 1 and 2 to enteric disease progression using a novel bovine ileal loop model and a murine model of infectious enterocolitis. Infect Immun. 2005;73(11):7161–7169. doi: 10.1128/IAI.73.11.7161-7169.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hapfelmeier S, Ehrbar K, Stecher B, Barthel M, Kremer M, Hardt WD. Role of the Salmonella pathogenicity island 1 effector proteins SipA, SopB, SopE, and SopE2 in Salmonella enterica subspecies 1 serovar Typhimurium colitis in streptomycin-pretreated mice. Infect Immun. 2004;72(2):795–809. doi: 10.1128/IAI.72.2.795-809.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brawn LC, Hayward RD, Koronakis V. Salmonella SPI1 effector SipA persists after entry and cooperates with a SPI2 effector to regulate phagosome maturation and intracellular replication. Cell Host Microbe. 2007;1(1):63–75. doi: 10.1016/j.chom.2007.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lawley TD, Chan K, Thompson LJ, Kim CC, Govoni GR, Monack DM. Genome-wide screen for Salmonella genes required for long-term systemic infection of the mouse. PLoS Pathog. 2006;2(2):e11. doi: 10.1371/journal.ppat.0020011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steele-Mortimer O, Brumell JH, Knodler LA, Meresse S, Lopez A, Finlay BB. The invasion-associated type III secretion system of Salmonella enterica serovar Typhimurium is necessary for intracellular proliferation and vacuole biogenesis in epithelial cells. Cell Microbiol. 2002;4(1):43–54. doi: 10.1046/j.1462-5822.2002.00170.x. [DOI] [PubMed] [Google Scholar]
- Buckner MM, Croxen MA, Arena ET, Finlay BB. A comprehensive study of the contribution of Salmonella enterica serovar Typhimurium SPI2 effectors to bacterial colonization, survival, and replication in typhoid fever, macrophage, and epithelial cell infection models. Virulence. 2011;2(3):208–216. doi: 10.4161/viru.2.3.15894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lilic M, Stebbins CE. Re-structuring the host cell: up close with Salmonella’s molecular machinery. Microbes Infect. 2004;6(13):1205–1211. doi: 10.1016/j.micinf.2004.08.001. [DOI] [PubMed] [Google Scholar]
- Patel JC, Galan JE. Manipulation of the host actin cytoskeleton by Salmonella–all in the name of entry. Curr Opin Microbiol. 2005;8(1):10–15. doi: 10.1016/j.mib.2004.09.001. [DOI] [PubMed] [Google Scholar]
- Ly KT, Casanova JE. Mechanisms of Salmonella entry into host cells. Cell Microbiol. 2007;9(9):2103–2111. doi: 10.1111/j.1462-5822.2007.00992.x. [DOI] [PubMed] [Google Scholar]
- Brown NF, Vallance BA, Coombes BK, Valdez Y, Coburn BA, Finlay BB. Salmonella pathogenicity island 2 is expressed prior to penetrating the intestine. PLoS Pathog. 2005;1(3):e32. doi: 10.1371/journal.ppat.0010032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dandekar T, Astrid F, Jasmin P, Hensel M. Salmonella enterica: a surprisingly well-adapted intracellular lifestyle. Front Microbiol. 2012;3:164. doi: 10.3389/fmicb.2012.00164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bajaj V, Lucas RL, Hwang C, Lee CA. Co-ordinate regulation of Salmonella typhimurium invasion genes by environmental and regulatory factors is mediated by control of hilA expression. Mol Microbiol. 1996;22(4):703–714. doi: 10.1046/j.1365-2958.1996.d01-1718.x. [DOI] [PubMed] [Google Scholar]
- Lee CA, Falkow S. The ability of Salmonella to enter mammalian cells is affected by bacterial growth state. Proc Natl Acad Sci USA. 1990;87(11):4304–4308. doi: 10.1073/pnas.87.11.4304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lundberg U, Vinatzer U, Berdnik D, von Gabain A, Baccarini M. Growth phase-regulated induction of Salmonella-induced macrophage apoptosis correlates with transient expression of SPI-1 genes. J Bacteriol. 1999;181(11):3433–3437. doi: 10.1128/jb.181.11.3433-3437.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernandez SB, Cota I, Ducret A, Aussel L, Casadesus J. Adaptation and preadaptation of Salmonella enterica to Bile. PLoS Genet. 2012;8(1):e1002459. doi: 10.1371/journal.pgen.1002459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pang E, Tien-Lin C, Selvaraj M, Chang J, Kwang J. Deletion of the aceE gene (encoding a component of pyruvate dehydrogenase) attenuates Salmonella enterica serovar Enteritidis. FEMS Immunol Med Microbiol. 2011;63(1):108–118. doi: 10.1111/j.1574-695X.2011.00834.x. [DOI] [PubMed] [Google Scholar]
- Bowden SD, Rowley G, Hinton JC, Thompson A. Glucose and glycolysis are required for the successful infection of macrophages and mice by Salmonella enterica serovar typhimurium. Infect Immun. 2009;77(7):3117–3126. doi: 10.1128/IAI.00093-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowden SD, Ramachandran VK, Knudsen GM, Hinton JC, Thompson A. An incomplete TCA cycle increases survival of Salmonella Typhimurium during infection of resting and activated murine macrophages. PLoS One. 2010;5(11):e13871. doi: 10.1371/journal.pone.0013871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santiviago CA, Reynolds MM, Porwollik S, Choi SH, Long F, Andrews-Polymenis HL, McClelland M. Analysis of pools of targeted Salmonella deletion mutants identifies novel genes affecting fitness during competitive infection in mice. PLoS Pathog. 2009;5(7):e1000477. doi: 10.1371/journal.ppat.1000477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee SH, Jung BY, Rayamahji N, Lee HS, Jeon WJ, Choi KS, Kweon CH, Yoo HS. A multiplex real-time PCR for differential detection and quantification of Salmonella spp., Salmonella enterica serovar Typhimurium and Enteritidis in meats. J Vet Sci. 2009;10(1):43–51. doi: 10.4142/jvs.2009.10.1.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang Y, Suyemoto M, Garner CD, Cicconi KM, Altier C. Formate acts as a diffusible signal to induce Salmonella invasion. J Bacteriol. 2008;190(12):4233. doi: 10.1128/JB.00205-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valle E, Guiney DG. Characterization of Salmonella-induced cell death in human macrophage-like THP-1 cells. Infect Immun. 2005;73(5):2835–2840. doi: 10.1128/IAI.73.5.2835-2840.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfaffl MW, Horgan GW, Dempfle L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002;30(9):e36. doi: 10.1093/nar/30.9.e36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibarra JA, Steele-Mortimer O. Salmonella–the ultimate insider. Salmonella virulence factors that modulate intracellular survival. Cell Microbiol. 2009;11(11):1579–1586. doi: 10.1111/j.1462-5822.2009.01368.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu Q, Coburn B, Deng W, Li Y, Shi X, Lan Q, Wang B, Coombes BK, Finlay BB. Salmonella enterica serovar Senftenberg human clinical isolates lacking SPI-1. J Clin Microbiol. 2008;46(4):1330–1336. doi: 10.1128/JCM.01255-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Srikanth CV, Mercado-Lubo R, Hallstrom K, McCormick BA. Salmonella effector proteins and host-cell responses. Cell Mol Life Sci. 2011;68(22):3687–3697. doi: 10.1007/s00018-011-0841-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tucker SC, Galan JE. Complex function for SicA, a Salmonella enterica serovar typhimurium type III secretion-associated chaperone. J Bacteriol. 2000;182(8):2262–2268. doi: 10.1128/jb.182.8.2262-2268.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaniga K, Tucker S, Trollinger D, Galan JE. Homologs of the Shigella IpaB and IpaC invasins are required for Salmonella typhimurium entry into cultured epithelial cells. J Bacteriol. 1995;177(14):3965–3971. doi: 10.1128/jb.177.14.3965-3971.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schechter LM, Lee CA. AraC/XylS family members, HilC and HilD, directly bind and derepress the Salmonella typhimurium hilA promoter. Mol Microbiol. 2001;40(6):1289–1299. doi: 10.1046/j.1365-2958.2001.02462.x. [DOI] [PubMed] [Google Scholar]
- Olekhnovich IN, Kadner RJ. DNA-binding activities of the HilC and HilD virulence regulatory proteins of Salmonella enterica serovar Typhimurium. J Bacteriol. 2002;184(15):4148–4160. doi: 10.1128/JB.184.15.4148-4160.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellermeier CD, Slauch JM. RtsA and RtsB coordinately regulate expression of the invasion and flagellar genes in Salmonella enterica serovar Typhimurium. J Bacteriol. 2003;185(17):5096. doi: 10.1128/JB.185.17.5096-5108.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin SL, Le TX, Cowen DS. SptP, a Salmonella typhimurium type III-secreted protein, inhibits the mitogen-activated protein kinase pathway by inhibiting Raf activation. Cell Microbiol. 2003;5(4):267–275. doi: 10.1046/j.1462-5822.2003.t01-1-00274.x. [DOI] [PubMed] [Google Scholar]
- Haraga A, Miller SI. A Salmonella enterica serovar typhimurium translocated leucine-rich repeat effector protein inhibits NF-kappa B-dependent gene expression. Infect Immun. 2003;71(7):4052–4058. doi: 10.1128/IAI.71.7.4052-4058.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Button JE, Galan JE. Regulation of chaperone/effector complex synthesis in a bacterial type III secretion system. Mol Microbiol. 2011;81(6):1474–1483. doi: 10.1111/j.1365-2958.2011.07784.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kage H, Takaya A, Ohya M, Yamamoto T. Coordinated regulation of expression of Salmonella pathogenicity island 1 and flagellar type III secretion systems by ATP-dependent ClpXP protease. J Bacteriol. 2008;190(7):2470–2478. doi: 10.1128/JB.01385-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cirillo DM, Valdivia RH, Monack DM, Falkow S. Macrophage-dependent induction of the Salmonella pathogenicity island 2 type III secretion system and its role in intracellular survival. Mol Microbiol. 1998;30(1):175–188. doi: 10.1046/j.1365-2958.1998.01048.x. [DOI] [PubMed] [Google Scholar]
- Deiwick J, Nikolaus T, Erdogan S, Hensel M. Environmental regulation of Salmonella pathogenicity island 2 gene expression. Mol Microbiol. 1999;31(6):1759–1773. doi: 10.1046/j.1365-2958.1999.01312.x. [DOI] [PubMed] [Google Scholar]
- Beuzon CR, Meresse S, Unsworth KE, Ruiz-Albert J, Garvis S, Waterman SR, Ryder TA, Boucrot E, Holden DW. Salmonella maintains the integrity of its intracellular vacuole through the action of SifA. EMBO J. 2000;19(13):3235–3249. doi: 10.1093/emboj/19.13.3235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eriksson S, Lucchini S, Thompson A, Rhen M, Hinton JC. Unravelling the biology of macrophage infection by gene expression profiling of intracellular Salmonella enterica. Mol Microbiol. 2003;47(1):103–118. doi: 10.1046/j.1365-2958.2003.03313.x. [DOI] [PubMed] [Google Scholar]
- Pfeifer CG, Marcus SL, Steele-Mortimer O, Knodler LA, Finlay BB. Salmonella typhimurium virulence genes are induced upon bacterial invasion into phagocytic and nonphagocytic cells. Infect Immun. 1999;67(11):5690–5698. doi: 10.1128/iai.67.11.5690-5698.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drecktrah D, Knodler LA, Ireland R, Steele-Mortimer O. The mechanism of Salmonella entry determines the vacuolar environment and intracellular gene expression. Traffic. 2006;7(1):39–51. doi: 10.1111/j.1600-0854.2005.00360.x. [DOI] [PubMed] [Google Scholar]
- Chuang YC, Wang KC, Chen YT, Yang CH, Men SC, Fan CC, Chang LH, Yeh KS. Identification of the genetic determinants of Salmonella enterica serotype Typhimurium that may regulate the expression of the type 1 fimbriae in response to solid agar and static broth culture conditions. BMC Microbiol. 2008;8:126. doi: 10.1186/1471-2180-8-126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umitsuki G, Wachi M, Takada A, Hikichi T, Nagai K. Involvement of RNase G in in vivo mRNA metabolism in Escherichia coli. Genes Cells. 2001;6(5):403–410. doi: 10.1046/j.1365-2443.2001.00430.x. [DOI] [PubMed] [Google Scholar]
- Guo A, Lasaro MA, Sirard JC, Kraehenbuhl JP, Schifferli DM. Adhesin-dependent binding and uptake of Salmonella enterica serovar Typhimurium by dendritic cells. Microbiology. 2007;153(Pt 4):1059–1069. doi: 10.1099/mic.0.2006/000331-0. [DOI] [PubMed] [Google Scholar]
- Sekirov I, Gill N, Jogova M, Tam N, Robertson M, de Llanos R, Li Y, Finlay BB. Salmonella SPI-1-mediated neutrophil recruitment during enteric colitis is associated with reduction and alteration in intestinal microbiota. Gut Microbes. 2010;1(1):30–41. doi: 10.4161/gmic.1.1.10950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lawhon SD, Maurer R, Suyemoto M, Altier C. Intestinal short-chain fatty acids alter Salmonella typhimurium invasion gene expression and virulence through BarA/SirA. Mol Microbiol. 2002;46(5):1451–1464. doi: 10.1046/j.1365-2958.2002.03268.x. [DOI] [PubMed] [Google Scholar]