

The risk of hypoglycemia is one of the major obstacles in the optimum treatment in both type 1 (1,2) and type 2 (3) diabetes, as well as in patients in intensive care units (4). Furthermore, in addition to short-term risks, including death, hypoglycemia also produces long-term health consequences including a substantial risk of death from cardiovascular events (5). Thus therapies to minimize the risk of hypoglycemia could have a profound impact on public health (2). The main cause of hypoglycemia is a failure in the counterregulatory autonomic and endocrine responses that normally act to correct low blood glucose (2). Failure in counterregulatory responses is caused by antecedent hypoglycemia itself and is therefore referred to as hypoglycemia-induced autonomic failure (2). Counterregulatory responses are mediated by glucose-inhibited neurons in the ventromedial nucleus (VMN) of the hypothalamus (6,7). Specifically, inhibiting glucose metabolism in neurons in the VMN of the hypothalamus induces systemic counterregulatory responses (6), whereas inhibiting glutamate neurotransmission in Sf1 neurons (confined to the VMN) impairs counterregulatory responses (7).
Szepietowska et al. (8) now report an unexpected mechanism regulating counterregulatory responses that may also contribute to counterregulatory failure: the EphA5-ephrin signaling system. Ephrins were discovered in a search for tyrosine kinase receptor homologs that might contribute to oncogenesis (9) by screening for homology to the oncogene v-fps. The resulting gene was highly overexpressed in an erythropoietin-producing human hepatocellular carcinoma cell line, from which the gene, along with its homology to fps, derived its name. Further screening revealed several more homologs, until the Eph family became the largest known family of receptor-like tyrosine kinases, divided into two large classes, EphA and EphB. Endogenous ligands for the family were discovered using the ectodomains of two members of the family, which demonstrated that the endogenous ligands are also expressed on the cell surface and, in fact, only function when bound to the cell surface (10). Subsequent studies demonstrated that Ephrin/Eph signaling acts to guide axonal development via a repulsive mechanism (11) but also implicated a wide variety of cell-cell interactions in adults, including regulation of N-methyl-d-aspartate–dependent synaptic function (12).
Like other tyrosine kinase receptors, the Eph family autophosphorylate upon activation, which is required for signaling. However, a unique feature of Eph/Ephrin signaling is that, due to the membrane localization of the ligand, the signaling can occur in both directions, with the Eph signaling said to be in the “forward” direction and the Ephrin signaling said to occur in the “reverse” direction, and indeed the two directions often produce opposite effects. A particularly relevant example of this mechanism was the surprising discovery that EphA5/EphrinA5 signaling plays a key role in regulating insulin secretion from intact pancreatic islets (13). This study demonstrated that both EphA5 and its ligand EphrinA5 are expressed in pancreatic β-cells, and whereas the forward signaling of the EphA5 receptor inhibits basal insulin secretion in intact islets (at low glucose concentrations), the reverse signaling of the EphrinA5 ligand stimulates insulin secretion in intact islets at high glucose concentration (13). Furthermore, glucose induces dephosphorylation of the EphA5 receptor, an inactivating mechanism that is necessary for glucose-induced insulin secretion (13). This opposition between forward signaling and reverse signaling produces an either/or binary bistability that provides a “sticky” feed-forward additional level of regulation, a common feature of many metabolic processes, including the lac operon (14).
Glucose-sensing hypothalamic neurons and pancreatic β-cells sense glucose through mechanisms that specifically entail using the pancreatic form of glucokinase and glucose metabolism (15). Thus Szepietowska et al. hypothesized that hypothalamic EphA5/EphrinA5 signaling might play a role in mediating counterregulatory responses to hypoglycemia (8). These investigators now report that infusing Ephrin5A conjugates or enhancing Ephrin5A expression in the ventromedial hypothalamus enhances counterregulatory responses, whereas inhibition of Ephrin5A expression by short hairpin RNA interference inhibits counterregulatory responses (8).
Among the most compelling subsequent analyses will be the determination of the cellular identity of the cells expressing the relevant EphA5 and EprhinA5 ligand. Of particular relevance to this question, the serotype of adeno-associated virus used, adeno-associated virus 2, almost exclusively (>95%) transfects neurons (16). Therefore, even though Ephrin ligands are expressed in both neurons and glia, the EphrinA5 relevant to counterregulation is most likely neuronal. Although glial function is reduced by inhibition of EphrinA5, as indicated by reduced glutamine, this is likely a reflection of reduced release of glutamate, which is recycled by glia into glutamine. Furthermore, in the hypothalamus EphrinA5 is expressed mainly in medial arcuate neurons, which predominantly express neuropeptide Y (NPY)/Agouti-related peptide (AgRP), not in the VMN (17). Activation of EphA5 receptors, most likely in VMN neurons, enhances counterregulatory responses (8). Conversely, activation of EphA5 receptors (in β-cells) inhibits insulin secretion, but (reverse) EphrinA5 signaling enhances insulin secretion (13). Furthermore NPY preferentially inhibits glucose-inhibited neurons in the VMN (18). Since NPY is a signal for nutritional insufficiency, it is plausible that NPY neurons would inhibit glucose action as an additional layer of regulation to preserve blood glucose during nutritional insufficiency. On the other hand increased glucose inactivates EphA5 receptor action (probably by dephosphorylation), at least in β-cells (13), allowing the antagonistic reverse signaling (plausibly increased glucose metabolism) to dominate. Thus it is plausible that hypoglycemia increases EphA5 signaling in glucose-inhibited VMN neurons, leading to reduced glucose metabolism, thereby enhancing responsiveness to hypoglycemia. At the same time, reverse signaling of EphrinA5 in NPY neurons would be expected to increase glucose metabolism in these neurons, inhibiting NPY secretion, thus disinhibiting the same VMN neurons (Fig. 1). Such a feed-forward mechanism would have the effect of producing bistable states, which has indeed been demonstrated for NPY/AgRP neurons (19). We have suggested that a similar bistable state is produced by activation of hypothalamic peroxisome proliferator–activated receptor-α by hypoglycemia, and that the failure to produce this bistable state is associated with counterregulatory failure (20). Therefore it is plausible that failure in the forward signaling of EphA5 to reduce glucose metabolism in glucose-inhibited glutamatergic neurons may also contribute to counterregulatory failure and thus constitutes a novel target to improve counterregulatory responses in patients with diabetes.
FIG. 1.
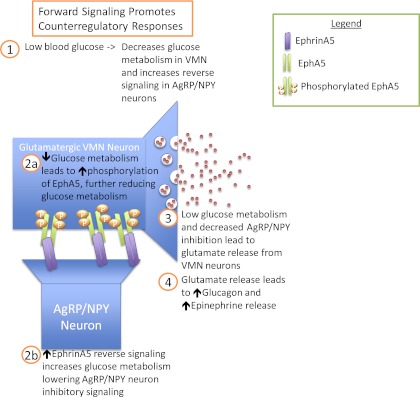
Hypothetical model by which the bidirectional signaling between EphA5 in VMN neurons and EphrinA5 in NPY neurons could produce bistability to enhance the robustness of counterregulatory responses to hypoglycemia.
ACKNOWLEDGMENTS
The Juvenile Diabetes Research Foundation funded studies relevant to this article. Research reported in this publication was supported by the National Institute on Aging of the National Institutes of Health under Award Number FAG042299A.
The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
No potential conflicts of interest relevant to this article were reported.
Footnotes
See accompanying original article, p. 1282.
REFERENCES
- 1.Cryer PE. Banting Lecture. Hypoglycemia: the limiting factor in the management of IDDM. Diabetes 1994;43:1378–1389 [DOI] [PubMed] [Google Scholar]
- 2.Cryer PE. Elimination of hypoglycemia from the lives of people affected by diabetes. Diabetes 2011;60:24-27. [DOI] [PMC free article] [PubMed]
- 3.Johnston SS, Conner C, Aagren M, Smith DM, Bouchard J, Brett J. Evidence linking hypoglycemic events to an increased risk of acute cardiovascular events in patients with type 2 diabetes. Diabetes Care 2011;34:1164–1170 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Scurlock C, Raikhelkar J, Mechanick JI. The economics of glycemic control in the ICU in the United States. Curr Opin Clin Nutr Metab Care 2011;14:209–212 [DOI] [PubMed] [Google Scholar]
- 5.Hsu P-F, Sung S-H, Cheng H-M, et al. Association of clinical symptomatic hypoglycemia with cardiovascular events and total mortality in type 2 diabetes: a nationwide population-based study. Diabetes Care 2013;36:894–900 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Borg WP, Sherwin RS, During MJ, Borg MA, Shulman GI. Local ventromedial hypothalamus glucopenia triggers counterregulatory hormone release. Diabetes 1995;44:180–184 [DOI] [PubMed] [Google Scholar]
- 7.Tong Q, Ye C, McCrimmon RJ, et al. Synaptic glutamate release by ventromedial hypothalamic neurons is part of the neurocircuitry that prevents hypoglycemia. Cell Metab 2007;5:383–393 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Szepietowska B, Zhu W, Czyzyk J, Eid T, Sherwin RS. EphA5-ephrinA5 interactions within the ventromedial hypothalamus influence counterregulatory hormone release and local glutamine/glutamate balance during hypoglycemia. Diabetes 2013;62:1282–1288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hirai H, Maru Y, Hagiwara K, Nishida J, Takaku F. A novel putative tyrosine kinase receptor encoded by the eph gene. Science 1987;238:1717–1720 [DOI] [PubMed] [Google Scholar]
- 10.Davis S, Gale NW, Aldrich TH, et al. Ligands for EPH-related receptor tyrosine kinases that require membrane attachment or clustering for activity. Science 1994;266:816–819 [DOI] [PubMed] [Google Scholar]
- 11.Drescher U, Kremoser C, Handwerker C, Löschinger J, Noda M, Bonhoeffer F. In vitro guidance of retinal ganglion cell axons by RAGS, a 25 kDa tectal protein related to ligands for Eph receptor tyrosine kinases. Cell 1995;82:359–370 [DOI] [PubMed] [Google Scholar]
- 12.Henderson JT, Georgiou J, Jia Z, et al. The receptor tyrosine kinase EphB2 regulates NMDA-dependent synaptic function. Neuron 2001;32:1041–1056 [DOI] [PubMed] [Google Scholar]
- 13.Konstantinova I, Nikolova G, Ohara-Imaizumi M, et al. EphA-Ephrin-A-mediated beta cell communication regulates insulin secretion from pancreatic islets. Cell 2007;129:359–370 [DOI] [PubMed] [Google Scholar]
- 14.Laurent M, Charvin G, Guespin-Michel J. Bistability and hysteresis in epigenetic regulation of the lactose operon. Since Delbrück, a long series of ignored models. Cell Mol Biol (Noisy-le-grand) 2005;51:583–594 [PubMed] [Google Scholar]
- 15.Yang XJ, Kow LM, Funabashi T, Mobbs CV. Hypothalamic glucose sensor: similarities to and differences from pancreatic beta-cell mechanisms. Diabetes 1999;48:1763–1772 [DOI] [PubMed] [Google Scholar]
- 16.Xu R, Janson CG, Mastakov M, et al. Quantitative comparison of expression with adeno-associated virus (AAV-2) brain-specific gene cassettes. Gene Ther 2001;8:1323–1332 [DOI] [PubMed] [Google Scholar]
- 17.Zarbalis K, Wurst W. Expression domains of murine ephrin-A5 in the pituitary and hypothalamus. Mech Dev 2000;93:165–168 [DOI] [PubMed] [Google Scholar]
- 18.Chee MJ, Myers MG, Jr, Price CJ, Colmers WF. Neuropeptide Y suppresses anorexigenic output from the ventromedial nucleus of the hypothalamus. J Neurosci 2010;30:3380–3390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yang Y, Atasoy D, Su HH, Sternson SM. Hunger states switch a flip-flop memory circuit via a synaptic AMPK-dependent positive feedback loop. Cell 2011;146:992–1003. [DOI] [PMC free article] [PubMed]
- 20.Poplawski MM, Mastaitis JW, Mobbs CV. Naloxone, but not valsartan, preserves responses to hypoglycemia after antecedent hypoglycemia: role of metabolic reprogramming in counterregulatory failure. Diabetes 2011;60:39–46 [DOI] [PMC free article] [PubMed] [Google Scholar]