

Abstract
The large-scale use of antivirals during influenza pandemics poses a significant selection pressure for drug-resistant pathogens to emerge and spread in a population. This requires treatment strategies to minimize total infections as well as the emergence of resistance. Here we propose a mathematical model in which individuals infected with wild-type influenza, if treated, can develop de novo resistance and further spread the resistant pathogen. Our main purpose is to explore the impact of two important factors influencing treatment effectiveness: i) the relative transmissibility of the drug-resistant strain to wild-type, and ii) the frequency of de novo resistance. For the endemic scenario, we find a condition between these two parameters that indicates whether treatment regimes will be most beneficial at intermediate or more extreme values (e.g., the fraction of infected that are treated). Moreover, we present analytical expressions for effective treatment regimes and provide evidence of its applicability across a range of modeling scenarios: endemic behavior with deterministic homogeneous mixing, and single-epidemic behavior with deterministic homogeneous mixing and stochastic heterogeneous mixing. Therefore, our results provide insights for the control of drug-resistance in influenza across time scales.
Introduction
Rapid antigenic evolution in the influenza virus increases the likelihood of emergence of novel strains, against which little to no immunity may exist in the host population [1]–[4]. In this scenario, if vaccines are not yet available or non-pharmaceutical interventions have limited impact on disease containment, antiviral treatment plays a crucial role in the control of the disease [3], [5]–[7]. A critical constraint in the deployment of antivirals agents (e.g., M2 inhibitors and neuraminidase inhibitors [8]) is the evolution of highly transmissible drug-resistant mutants [9]. Resistance decreases the effectiveness of chemotherapy in infected patients, prolonging recovery or leading to outright treatment failure [10]. Epidemics of untreatable strains have the potential to cause major morbidity and mortality [11]–[14], with significant economic costs for both the individual and for society writ large [15]. Consequently, public health policy has a growing need to understand the key factors that lead to the rise and spread of resistance, and to devise strategies that amplify the effectiveness of existing drugs, while halting the spread of resistance [16]–[19].
In addition to important precautionary measures, such as improvement of hospital counter-infection methods and regulation of antiviral use, mathematical models can be used to explore plausible competition scenarios between sensitive and resistant strains and the impact of treatment strategies on these dynamics [17], [19]–[22]. Previous models of the development of resistance of influenza to antiviral agents have focused on efforts to minimize the fraction of drug-resistant infections during an epidemic outbreak [5]–[7], [17] and to give recommendations that inform policy [8], [15], [18], [23]–[25]. However, the study of the long-term (endemic) dynamics of drug-resistance has received less attention [26].
The present work assesses the effectiveness of treatment at minimizing the total number of infections while halting the spread of drug-resistance, both from an endemic and a single-epidemic perspective. We focus our attention on two points: i) the relative transmissibility of the drug-resistant strain with respect to the wild-type (drug-sensitive) strain, and ii) the frequency of de novo resistance. Point i) is related to the fitness cost associated with the evolution of drug resistance, reflected in a reduced transmissibility of the drug-resistant pathogen relative to its wild-type counterpart [8], [27]. Recent evidence has demonstrated, however, that this reduction in fitness may be limited due to compensatory mutations which can restore fitness without loss of resistance-conferring genes [9], [28]. Point ii) represents the probability that treatment leads to resistance within the treated host (de novo). Both quantities are crucial in the population dynamics of drug-resistance, specially due to their variability within different epidemiological settings [8]. Nonetheless, their combined effect on the effectiveness of treatment regimes during influenza pandemics is not fully understood [5].
We build on a previous model [17] to examine these issues in the long-term (endemic disease prevalence) as well as in the short-term (single-epidemic). Lipsitch et al. [17] observed that intermediate levels of antiviral use are indicated to reduce the attack rate during an influenza pandemic. Complementing these results, we find that to effectively reduce the endemic levels of the wild-type and resistant strains, treatment regimes (i.e., treated fraction) should be at intermediate levels if the resistant strain is highly transmissible and de novo resistance is rare. However, if resistance comes with a high fitness cost and de novo resistance is frequent, then higher levels of antiviral use may be preferable. In the single epidemic case we compare our optimal treatment regime with that of [17], showing that their relative effectiveness also depends on the strains’ relative transmissibility and the frequency of de novo resistance. Moreover, we demonstrate the applicability of our optimal treatment regimes by evidencing its effectiveness at quelling the spread of resistance when considering the effects of the stochasticity inherent to the transmission dynamics and the complex contact structure in the population.
Methods
Model Formulation
We extend a version of the model in [17] to include demography (see Figure 1). Susceptible hosts,  , enter the population at a per-capita rate
, enter the population at a per-capita rate  and die at rate equal to
and die at rate equal to  , keeping the total population size,
, keeping the total population size,  , constant. Susceptible individuals can be infected by pathogens either sensitive or resistant to the available antiviral (this model does not include superinfection with both strains). A fraction
, constant. Susceptible individuals can be infected by pathogens either sensitive or resistant to the available antiviral (this model does not include superinfection with both strains). A fraction  of patients infected with the wild-type strain are treated, and a fraction
of patients infected with the wild-type strain are treated, and a fraction  of those treated develop resistance de novo. Therefore, individuals infected with the wild-type strain are either untreated (
of those treated develop resistance de novo. Therefore, individuals infected with the wild-type strain are either untreated ( ), effectively treated (
), effectively treated ( ), or resistant to treatment (
), or resistant to treatment ( ). Infection with a resistant strain is either developed de novo or acquired from another resistant-infected individual. Susceptible individuals become infected at a rate proportional to the densities of susceptible and infected individuals, and to the transmission rates of each class,
). Infection with a resistant strain is either developed de novo or acquired from another resistant-infected individual. Susceptible individuals become infected at a rate proportional to the densities of susceptible and infected individuals, and to the transmission rates of each class,  , and
, and  , respectively. Untreated, treated, and resistant infected individuals recover at per-capita rates
, respectively. Untreated, treated, and resistant infected individuals recover at per-capita rates  , and
, and  , respectively. We assume no disease-induced mortality, and that the pathogen induces sterilizing immunity [21], [29].
, respectively. We assume no disease-induced mortality, and that the pathogen induces sterilizing immunity [21], [29].
Figure 1. Compartmental Model for Eqs. (1)–(5).

The relative transmissibility of the resistant strain is defined as  . Successfully treated individuals: 1) are not more infectious:
. Successfully treated individuals: 1) are not more infectious:  , where
, where  is the reduction in viral shedding [16], [30], [31], and 2) recover faster:
is the reduction in viral shedding [16], [30], [31], and 2) recover faster:  , where
, where  is the increase in recovery rate [8], [16], [32].
is the increase in recovery rate [8], [16], [32].
The ordinary differential equation (ODE) model describing these dynamics is
| (1) |
| (2) |
| (3) |
| (4) |
| (5) |
with forces of infection  and
and  . Note, we are modeling densities (i.e.,
. Note, we are modeling densities (i.e.,  ). In what follows, let
). In what follows, let  be the total per-capita rate out of class
be the total per-capita rate out of class  , i.e.,
, i.e.,  ,
,  , and
, and  .
.
Reproduction numbers
The basic reproduction number,  , is the average number of secondary cases produced by a typical infected individual in a completely susceptible population. We find
, is the average number of secondary cases produced by a typical infected individual in a completely susceptible population. We find  for each strain using the Next Generation Operator (NGO) method [33]. The non-zero eigenvalues of the NGO matrix
for each strain using the Next Generation Operator (NGO) method [33]. The non-zero eigenvalues of the NGO matrix
| (6) |
| (7) |
are the reproduction number of the wild-type and resistant strains, respectively. Detailed derivations can be found in the Supporting Information (Text S1).
Results
Fixed Points and Bifurcation Analysis
The system (1)–(5) has three fixed points (FPs): 1) a disease free equilibrium (DFE); a FP where only the resistant strain persists (RFP); and a coexistence FP in which both strains coexist (CFP). Conceptually, these FPs represent: 1) eradication of both resistant and wild-type strain, eradication of the wild-type strain when treatment and/or relative transmissibility are high enough to allow persistence of the resistant strain; and coexistence of both strains due to low treatment and/or low fitness of the resistant strain, where typically the resistant strain persists at low levels. The FPs are:
DFE:
| (8) |
RFP:
| (9) |
CFP:
| (10) |
where
| (11) |
The recovered class fraction in each case is given by 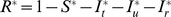 . Comparing the susceptible steady states in (9) and (10) suggests that for the RFP, prevalent infections are attributable to the resistant strain, whereas for the CFP, the reproduction number of the wild-type strain determines how prevalent the disease is.
. Comparing the susceptible steady states in (9) and (10) suggests that for the RFP, prevalent infections are attributable to the resistant strain, whereas for the CFP, the reproduction number of the wild-type strain determines how prevalent the disease is.
To be biologically significant (BS) the steady states have to lie in the set
The RFP is BS if  . For the CFP,
. For the CFP,  must hold so that
must hold so that  . This also implies that the numerator of
. This also implies that the numerator of  in (11) is positive. For
in (11) is positive. For  to be non-negative, the denominator of
to be non-negative, the denominator of  must be positive, i.e.,
must be positive, i.e.,  , which implies
, which implies  . For
. For  and
and  to be non-negative
to be non-negative  must hold. Therefore, the CFP is BS if
must hold. Therefore, the CFP is BS if
| (12) |
Thus, the two strains coexist if the wild-type strain is transmissible enough to be able to spread, and also more transmissible than the resistant strain.
Stability of fixed points
For the stability analysis of the FPs we study the eigenvalues of the matrix in the linearized system around the FPs: equilibria that have eigenvalues with negative real part are stable, whereas equilibria that have eigenvalues with positive real part are unstable [34]. We present here the results of the analysis; detailed analytic derivations can be found in the Text S1.
As expected, the DFE is globally stable if  and
and  . The RFP is locally stable if
. The RFP is locally stable if  . While determining the stability of the CFP is not analytically tractable, (12) states that the CFP is BS if
. While determining the stability of the CFP is not analytically tractable, (12) states that the CFP is BS if  and
and  . Thus, the conditions in (12) imply that neither the DFE nor the RFP are stable. We then conjecture that the CFP is BS and globally stable if (12) holds. Epidemiological arguments and numerical integrations support this hypothesis.
. Thus, the conditions in (12) imply that neither the DFE nor the RFP are stable. We then conjecture that the CFP is BS and globally stable if (12) holds. Epidemiological arguments and numerical integrations support this hypothesis.
Bifurcation analysis
Depending on  and
and  , the system has one, two, or three BS FPs. Figure 2 features all four stability regions described above in the (
, the system has one, two, or three BS FPs. Figure 2 features all four stability regions described above in the ( ) and the (
) and the ( ) parameter space. The boundary of these regions can be found by solving
) parameter space. The boundary of these regions can be found by solving  yielding
yielding
| (13) |
and
| (14) |
Figure 2. Stability regions in the ( ,
,  ) and (
) and ( ) parameter space.
) parameter space.

Coexistence 2FP (CFP stable, DFE unstable); Coexistence 3FP (CFP stable, DFE unstable, RFP unstable); Resistance (DFE unstable, RFP stable). When the system crosses any of the region boundaries it experiences a transcritical bifurcation.
The boundaries are shown in Figure 2, where  is the red-dashed line,
is the red-dashed line,  is the dashed and horizontal line, and
is the dashed and horizontal line, and  is the dashed and vertical line. The intersection of these curves (black dot) represents the overall disease threshold: any increase in
is the dashed and vertical line. The intersection of these curves (black dot) represents the overall disease threshold: any increase in  or decrease in
or decrease in  away from this intersection would result in an epidemic. Moreover, Eq. (13) shows that, for appropriate parameter values, increasing
away from this intersection would result in an epidemic. Moreover, Eq. (13) shows that, for appropriate parameter values, increasing  or decreasing
or decreasing  , decreases the
, decreases the  -coordinate (
-coordinate ( ) of the overall disease threshold point. Thus, increasing the recovery rate or decreasing the transmission rate of those treated, represents an epidemiological trade-off: it jointly expands the “DFE” and the “Resistance” stability regions, making it more likely for the system to either stay disease-free or give rise to prevalent resistance (see Figure S4, S5, S6, S7, S8 in Text S1 for details).
) of the overall disease threshold point. Thus, increasing the recovery rate or decreasing the transmission rate of those treated, represents an epidemiological trade-off: it jointly expands the “DFE” and the “Resistance” stability regions, making it more likely for the system to either stay disease-free or give rise to prevalent resistance (see Figure S4, S5, S6, S7, S8 in Text S1 for details).
Optimal Treatment Regimes
The main goal of this work is to derive treatment regimes (i.e., treated fractions) that minimize the wild-type infections while restraining the spread of resistance. From the CFP in (10), it is clear that for very low treatment levels, the wild-type strain is prevalent in the population, and the resistant strain prevalence stays at minimal levels [10], i.e.,  . Additionally, treatment will reduce the viral shedding (
. Additionally, treatment will reduce the viral shedding ( ) and increase the recovery rate by
) and increase the recovery rate by  , implying that
, implying that  . Thus, treating a larger proportion of the population will reduce the number of wild-type infected. However, it will also increase the number of de novo resistant cases, as well as the pool of susceptibles for the resistant strain to spread in.
. Thus, treating a larger proportion of the population will reduce the number of wild-type infected. However, it will also increase the number of de novo resistant cases, as well as the pool of susceptibles for the resistant strain to spread in.
These observations intuitively suggest that an effective treatment strategy should minimize  by increasing
by increasing  , while keeping
, while keeping  . Formally,
. Formally,
| (15) |
Since  does not depend on
does not depend on  (Eq. (7)), and assuming
(Eq. (7)), and assuming  , (15) can be solved by reducing
, (15) can be solved by reducing  until
until  . This equality yields
. This equality yields  as in (14), a linearly decreasing function of the relative transmissibility,
as in (14), a linearly decreasing function of the relative transmissibility,  (see Figure 2).
(see Figure 2).
However, (15) is inadequate since it does not consider the fitness advantage that development of de novo resistant cases give to the resistant strain. As  is the expected number of new cases produced by a typical infected person in a susceptible population, this quantity can be considered a measure of the fitness of a pathogen at the population level. Additionally, in our model,
is the expected number of new cases produced by a typical infected person in a susceptible population, this quantity can be considered a measure of the fitness of a pathogen at the population level. Additionally, in our model,  is directly related to the within-host fitness of the resistant pathogen. The overall fitness of the resistant strain is the added contributions of the fitness at the population and the within-host level. To estimate this overall fitness, assume, for the sake of clarity,
is directly related to the within-host fitness of the resistant pathogen. The overall fitness of the resistant strain is the added contributions of the fitness at the population and the within-host level. To estimate this overall fitness, assume, for the sake of clarity,  . Let also
. Let also  and
and  be the number of resistant and wild-type cases in the
be the number of resistant and wild-type cases in the  “epidemic generation” (with duration approximately
“epidemic generation” (with duration approximately  ) in a predominantly susceptible population. Defining
) in a predominantly susceptible population. Defining  and
and  as the overall fitness of the resistant and wild-type strains, respectively, we obtain (see Text S1 for details):
as the overall fitness of the resistant and wild-type strains, respectively, we obtain (see Text S1 for details):
| (16) |
where  is the Heaviside step function (
is the Heaviside step function ( if
if  , and
, and  otherwise), i.e., if
otherwise), i.e., if  , the wild-type strain goes extinct. It is then clear that
, the wild-type strain goes extinct. It is then clear that  has an additional contribution from the de novo cases. More importantly, from the sole comparison of the reproduction numbers we cannot infer properly which strain will dominate, nor can we devise effective treatment regimes.
has an additional contribution from the de novo cases. More importantly, from the sole comparison of the reproduction numbers we cannot infer properly which strain will dominate, nor can we devise effective treatment regimes.
A more appropriate way to optimize the treatment regime is attained by focussing on the fixed points (FPs). The system has two FPs where the disease is endemic (RFP and CFP). On the one hand, if the CFP is stable, the optimal treatment regime,  , is defined as the fraction treated that yields the minimum number of wild-type infected, while the resistant is kept at lower endemic levels than the wild-type. Formally,
, is defined as the fraction treated that yields the minimum number of wild-type infected, while the resistant is kept at lower endemic levels than the wild-type. Formally,
| (17) |
where  . The regime
. The regime  can then be found by solving for
can then be found by solving for  in
in  (see Eq. (19)).
(see Eq. (19)).
On the other hand, if the RFP is stable, then treatment will have no effect on the prevalence of the resistant strain since  is not a function of
is not a function of  . Two scenarios are then possible: (A)
. Two scenarios are then possible: (A)
 , or (B)
, or (B)
 . An assessment of these two scenarios yields our definition of overall optimal treatment regime
. An assessment of these two scenarios yields our definition of overall optimal treatment regime  : if (A) is true,
: if (A) is true,  will minimize the endemic levels of the wild-type strain, while keeping the resistant strain at comparatively low levels; if (B) holds,
will minimize the endemic levels of the wild-type strain, while keeping the resistant strain at comparatively low levels; if (B) holds,  will transition the system to the RFP stability region. Hence, in case (A) it is best to maintain the system within the CFP limits, whereas in (B) the RFP will be preferred. The latter can be achieved by increasing
will transition the system to the RFP stability region. Hence, in case (A) it is best to maintain the system within the CFP limits, whereas in (B) the RFP will be preferred. The latter can be achieved by increasing  beyond
beyond  . Formally,
. Formally,
 |
We now show that conditions (A) and (B) can be expressed in terms of our two key parameters: relative transmissibility,  , and the frequency of de novo resistance,
, and the frequency of de novo resistance,  . To find
. To find  we solve for
we solve for  in
in  , or,
, or,
| (18) |
Within the CFP limits, (19) indicates when the overall fitness of both strains are equal (notice the similarity of the left hand side and the right hand side of (19) with, respectively,  and
and  in (16), when
in (16), when  ). The explicit expression for
). The explicit expression for  is given in the Text S1. Noteworthy,
is given in the Text S1. Noteworthy,  is the only value of
is the only value of  in
in  for which
for which  . This claim is justified as follows:
. This claim is justified as follows:  is a monotonically decreasing function of
is a monotonically decreasing function of  in
in  , with
, with  and
and  . Additionally,
. Additionally,  is either increasing or concave in
is either increasing or concave in  (see Figure 3). In both cases,
(see Figure 3). In both cases,  and
and  intersect at only one point (green dots in Figure 3). See Text S1 and Figure S10 for analytic details.
intersect at only one point (green dots in Figure 3). See Text S1 and Figure S10 for analytic details.
Figure 3. The two possible monotonicity behaviors of  .
.

In black,  is concave for
is concave for  (solid lines) (
(solid lines) ( ), and monotonically increasing for
), and monotonically increasing for  (dashed lines) (
(dashed lines) ( ). Red lines are the corresponding
). Red lines are the corresponding  curves. The x-values of the green and gray dots represent
curves. The x-values of the green and gray dots represent  and
and  , respectively, while the x-value of the red dot represents
, respectively, while the x-value of the red dot represents  . As
. As  (or
(or  ) increases, the red dot moves rightward, surpassing the green dot (
) increases, the red dot moves rightward, surpassing the green dot ( ), and eventually surpassing the gray dot as well (
), and eventually surpassing the gray dot as well ( ). At this point, the system displays a transcritical bifurcation between the CFP and the RFP. Other parameters:
). At this point, the system displays a transcritical bifurcation between the CFP and the RFP. Other parameters:  . A large value of
. A large value of  was used to magnify the difference between the two cases.
was used to magnify the difference between the two cases.
It is easy to show that  (gray dots in Figure 3). However, also
(gray dots in Figure 3). However, also  , where
, where
| (19) |
Then, if  , the term
, the term  represents the treatment regime within the region of coexistence (CFP) for which the resistant strain is as prevalent as in the resistant-only stability region (RFP) (red dot in Figure 3). Additionally, it can be deduced from (20) that
represents the treatment regime within the region of coexistence (CFP) for which the resistant strain is as prevalent as in the resistant-only stability region (RFP) (red dot in Figure 3). Additionally, it can be deduced from (20) that
| (20) |
In the Text S1 we show that when (21) holds,  is concave for
is concave for  . The concavity of
. The concavity of  means, biologically, that the resistant strain prevalence is sustained largely by de novo resistant cases. Put differently,
means, biologically, that the resistant strain prevalence is sustained largely by de novo resistant cases. Put differently,  is not large enough for the resistant strain to self-sustain high levels of prevalence in the absence of treated wild-type infected.
is not large enough for the resistant strain to self-sustain high levels of prevalence in the absence of treated wild-type infected.
Recalling that  and
and  , if
, if  is concave for
is concave for  and
and  , then
, then  (condition (B)), indicating that the RFP is preferred over the CFP (solid curves in Figure 3). Furthermore, condition
(condition (B)), indicating that the RFP is preferred over the CFP (solid curves in Figure 3). Furthermore, condition  reduces to
reduces to  given that
given that  for
for  . If instead
. If instead  , then condition (A) applies and keeping the system in the CFP while applying a treatment regime
, then condition (A) applies and keeping the system in the CFP while applying a treatment regime  will be the best option (dashed curves in Figure 3). These observations along with expression (20) allow to restate conditions (A) and (B), and therefore the optimal treatment, in terms of
will be the best option (dashed curves in Figure 3). These observations along with expression (20) allow to restate conditions (A) and (B), and therefore the optimal treatment, in terms of  and
and  as
as
 |
In case (A), which corresponds to a high  and low
and low  scenario, the CFP is stable with the wild-type and the resistant strains kept at low levels. In case (B) (i.e., low
scenario, the CFP is stable with the wild-type and the resistant strains kept at low levels. In case (B) (i.e., low  and high
and high  ), shifting the stability to the RFP is preferable since only the resistant strain will persist at low levels (see Figure 3). In other words, if the resistant strain features high relative transmissibility and resistance is rare, the best treatment regime would be at intermediate levels
), shifting the stability to the RFP is preferable since only the resistant strain will persist at low levels (see Figure 3). In other words, if the resistant strain features high relative transmissibility and resistance is rare, the best treatment regime would be at intermediate levels  ; whereas if the opposite holds true, treating a larger fraction (
; whereas if the opposite holds true, treating a larger fraction ( ) of the infected population is preferred.
) of the infected population is preferred.
Recalling  and
and  are found from (19) and
are found from (19) and  , respectively, and noticing that as
, respectively, and noticing that as  expression (19) reduces to
expression (19) reduces to  , we conclude that
, we conclude that  as
as  . For this reason, when
. For this reason, when  is small
is small  , becomes a good treatment strategy if (A) holds. Moreover, as
, becomes a good treatment strategy if (A) holds. Moreover, as  , it is expected that (A) holds, at least in the epidemiologically interesting cases where
, it is expected that (A) holds, at least in the epidemiologically interesting cases where  will likely be greater than
will likely be greater than  (i.e., the resistant strain can emerge and spread in the population). In conclusion, when
(i.e., the resistant strain can emerge and spread in the population). In conclusion, when  is small, then
is small, then  is a good treatment regime to minimize both the wild-type and the resistant strains (green bands in Figure 4).
is a good treatment regime to minimize both the wild-type and the resistant strains (green bands in Figure 4).
Figure 4. Effectiveness of  for
for  and
and  .
.

Prevalences  and
and  are depicted in black and
are depicted in black and  in red, for two different treatment recovery benefits (
in red, for two different treatment recovery benefits ( , solid;
, solid;  , dashed). The RFP is unstable for
, dashed). The RFP is unstable for  (blue dashed line). Strain dominance transition at
(blue dashed line). Strain dominance transition at  (vertical dashed lines). Optimal treatment regimes (
(vertical dashed lines). Optimal treatment regimes ( ) in green bands. Parameters:
) in green bands. Parameters:  ,
,  ,
,  ,
,  ,
,  (see also Figure S9).
(see also Figure S9).
Despite the large uncertainties in the frequency of patients that develop de novo resistance [8],  can be assumed to be relatively small. Figure 4 evidences how, for
can be assumed to be relatively small. Figure 4 evidences how, for  (as in [17]),
(as in [17]),  is the optimal treatment fraction: it diminishes the prevalence of the sensitive strain as much as possible, while hindering the emergence of the resistant strain. For low levels of treatment the CFP is stable: the wild-type strain prevails and the resistant strain (
is the optimal treatment fraction: it diminishes the prevalence of the sensitive strain as much as possible, while hindering the emergence of the resistant strain. For low levels of treatment the CFP is stable: the wild-type strain prevails and the resistant strain ( ) remains at low levels. As soon as
) remains at low levels. As soon as  , the resistant strain out-competes the wild-type strain. Expectedly, as treatment further reduces the infectious period (i.e. larger
, the resistant strain out-competes the wild-type strain. Expectedly, as treatment further reduces the infectious period (i.e. larger  , dashed lines), increasing treatment reduces the wild-type strain prevalence more effectively. In this case, the optimal levels of treatment are lower. A similar behavior is obtained when, instead of increasing
, dashed lines), increasing treatment reduces the wild-type strain prevalence more effectively. In this case, the optimal levels of treatment are lower. A similar behavior is obtained when, instead of increasing  , we reduce
, we reduce  (reduction of viral shedding due to treatment).
(reduction of viral shedding due to treatment).
Frequency of de novo resistance and endemic levels of resistance
We have shown how the frequency of de novo resistance,  , plays a crucial role in devising effective treatment strategies. In addition, we find that smaller values of
, plays a crucial role in devising effective treatment strategies. In addition, we find that smaller values of  lead to more abrupt transitions from wild-type to resistant strains. In other words, the smaller the probability of developing de novo resistance, the faster the RFP gains stability when the system is close to the threshold
lead to more abrupt transitions from wild-type to resistant strains. In other words, the smaller the probability of developing de novo resistance, the faster the RFP gains stability when the system is close to the threshold  (Figure 5). Thus, for small
(Figure 5). Thus, for small  , the system becomes more sensitive to variations in
, the system becomes more sensitive to variations in  ,
,  ,
,  , and
, and  near this threshold. This represents a potentially dangerous scenario: if the likelihood of de novo resistance is small, a policy-maker might underestimate the prospects of resistance emergence and, consequently, increase treatment levels to eradicate the wild-type strain. However, if treatment is increased above
near this threshold. This represents a potentially dangerous scenario: if the likelihood of de novo resistance is small, a policy-maker might underestimate the prospects of resistance emergence and, consequently, increase treatment levels to eradicate the wild-type strain. However, if treatment is increased above  , an abrupt transition may occur to a state where only resistant strains persist.
, an abrupt transition may occur to a state where only resistant strains persist.
Figure 5. Resistant strain prevalence vs. treatment fraction.

Smaller  leads to more abrupt transitions from wild-type to resistant strains. Larger
leads to more abrupt transitions from wild-type to resistant strains. Larger  renders
renders  ineffective as a treatment regime.
ineffective as a treatment regime.
Mathematically, this “abrupt transition” can be justified as follows: if  , then
, then  ; hence, when
; hence, when  , there is a “singularity” for
, there is a “singularity” for  and
and  in the CFP (10). Biologically, it is clear from (16) that
in the CFP (10). Biologically, it is clear from (16) that  and
and  . That is, as
. That is, as  , the reproduction numbers become the overall fitnesses of the strains, and
, the reproduction numbers become the overall fitnesses of the strains, and  represents the condition for which both strains are equally fit. Thus, the resistant strain outcompetes the wild-type strain as
represents the condition for which both strains are equally fit. Thus, the resistant strain outcompetes the wild-type strain as  surpasses
surpasses  .
.
The Single Epidemic Case
Frequently, public health programs and interventions are designed to prevent the emergence of drug resistance within a single epidemic. To address this issue, we model a closed population (i.e.,  in model (1)–(5)), and examine again the role of i) the relative transmissibility (
in model (1)–(5)), and examine again the role of i) the relative transmissibility ( ) and ii) the frequency of de novo resistance (
) and ii) the frequency of de novo resistance ( ) on the effectiveness of treatment regimes. In this assessment we focus on the final epidemic size (FS), defined as the proportion of the population infected during the epidemic. As in [17], we introduce the following correction to our numerical integrations: if
) on the effectiveness of treatment regimes. In this assessment we focus on the final epidemic size (FS), defined as the proportion of the population infected during the epidemic. As in [17], we introduce the following correction to our numerical integrations: if  then
then  . This prevents spurious results induced by the transmission of “non-cases” (since
. This prevents spurious results induced by the transmission of “non-cases” (since  , initially
, initially  can only increase due to de novo resistant cases; given that
can only increase due to de novo resistant cases; given that  is continuous in the ODE framework, the condition above avoids that a fraction of a de novo resistant case can cause a direct resistant infection). Throughout this section the following parameters are fixed:
is continuous in the ODE framework, the condition above avoids that a fraction of a de novo resistant case can cause a direct resistant infection). Throughout this section the following parameters are fixed:  ,
,  ,
,  ,
,  and
and  .
.
Figure 6 shows a feature demonstrated previously [6], [17], [22]: the existence of an “optimal” level of treatment for which the total FS is minimized. We can readily see this minimum is a function of  : as
: as  increases, the dip in the combined FS curve vanishes. Furthermore, the treatment regimes that minimize the total FS, are not optimal in terms of avoiding the emergence of resistance. Let
increases, the dip in the combined FS curve vanishes. Furthermore, the treatment regimes that minimize the total FS, are not optimal in terms of avoiding the emergence of resistance. Let  . We find that
. We find that  , where as before
, where as before  satisfies
satisfies  . That is, the minimum in the FS is reached when resistance has already significantly spread in the population. Additionally, Figure 6 shows that for larger
. That is, the minimum in the FS is reached when resistance has already significantly spread in the population. Additionally, Figure 6 shows that for larger  (diamond curves),
(diamond curves),  represents a value of the treatment fraction for which the resistant strain has already spread considerably throughout the population. This suggests that, as in the endemic case, the effectiveness of
represents a value of the treatment fraction for which the resistant strain has already spread considerably throughout the population. This suggests that, as in the endemic case, the effectiveness of  depends on the frequency of de novo resistance: as
depends on the frequency of de novo resistance: as  increases, the validity of
increases, the validity of  becomes compromised (notice similarity in black curves of Figures 5 and 6).
becomes compromised (notice similarity in black curves of Figures 5 and 6).
Figure 6. Final epidemic size of both strains vs.  .
.

( as asterisks and
as asterisks and  as diamonds) Vertical dashed lines represent
as diamonds) Vertical dashed lines represent  ; higher
; higher  , lower
, lower  . Note that
. Note that  is effective in halting the spread of resistance but not in reducing the total FS. Also, for larger
is effective in halting the spread of resistance but not in reducing the total FS. Also, for larger  ,
,  loses its effectiveness in avoiding the spread of resistance. Other parameters:
loses its effectiveness in avoiding the spread of resistance. Other parameters:  .
.
Notice also in Figure 6 that the epidemic is eradicated if  exceeds
exceeds  (Eq. (13)), where
(Eq. (13)), where  . That is, when the treatment fraction is large enough to rapidly halt the spread of the wild-type strain, the resistant strain will not emerge. This is possible, in part, given our assumption that treatment is implemented early in the epidemic (i.e.,
. That is, when the treatment fraction is large enough to rapidly halt the spread of the wild-type strain, the resistant strain will not emerge. This is possible, in part, given our assumption that treatment is implemented early in the epidemic (i.e.,  is small). In conclusion, if
is small). In conclusion, if  is relatively small and treatment is put in place later in the epidemic or it cannot surpass
is relatively small and treatment is put in place later in the epidemic or it cannot surpass  , then
, then  will ensure minimal spread of the resistant strain.
will ensure minimal spread of the resistant strain.
We now wish to contrast the effectiveness of  and
and  as a function of the relative transmissibility
as a function of the relative transmissibility  , assuming relatively low frequency of de novo resistance
, assuming relatively low frequency of de novo resistance  . Figure 7 shows the FSs (due to resistant strain (black) and total (blue)) vs.
. Figure 7 shows the FSs (due to resistant strain (black) and total (blue)) vs.  for
for  and
and  . A treatment regime
. A treatment regime  would “prioritize” the avoidance of resistance, while compromising the reduction of the overall epidemic; conversely,
would “prioritize” the avoidance of resistance, while compromising the reduction of the overall epidemic; conversely,  will, by definition, “prioritize” the minimization of the total epidemic size, while disregarding the spread of resistance. As a result,
will, by definition, “prioritize” the minimization of the total epidemic size, while disregarding the spread of resistance. As a result,  is more effective than
is more effective than  at halting the spread of resistance in the population, whereas
at halting the spread of resistance in the population, whereas  is a better option to reduce the overall epidemic. Moreover, since
is a better option to reduce the overall epidemic. Moreover, since 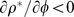 (see (14)), as
(see (14)), as  increases, a treatment regime
increases, a treatment regime  will systematically diminish the spread of resistance by reducing the treated fraction. Consequently, for higher
will systematically diminish the spread of resistance by reducing the treated fraction. Consequently, for higher  ,
,  will have minimal effects on reducing the total epidemic size (compare the diamond with the horizontal blue line, where no treatment is applied). Therefore, the decision to use
will have minimal effects on reducing the total epidemic size (compare the diamond with the horizontal blue line, where no treatment is applied). Therefore, the decision to use  or
or  as a treatment regime will mainly depend on how policy makers balance a larger epidemic produced largely by the wild-type strain, with minimal resistant cases (using
as a treatment regime will mainly depend on how policy makers balance a larger epidemic produced largely by the wild-type strain, with minimal resistant cases (using  , for which we have a better biological and mathematical understanding), versus a smaller overall epidemic with higher resistance incidence (using
, for which we have a better biological and mathematical understanding), versus a smaller overall epidemic with higher resistance incidence (using  ).
).
Figure 7. Final Sizes vs. relative transmissibility for  and
and  .
.

The figure shows the effectiveness of  and
and  vs. the relative transmissibility. For any value of
vs. the relative transmissibility. For any value of  ,
,  is more effective than
is more effective than  at avoiding the spread of resistance in the population (black diamond vs. black dashed curves). However,
at avoiding the spread of resistance in the population (black diamond vs. black dashed curves). However,  is more efficacious at reducing the overall epidemic (blue dashed vs. blue diamond). Solid line corresponds to
is more efficacious at reducing the overall epidemic (blue dashed vs. blue diamond). Solid line corresponds to  . Other parameters:
. Other parameters: 
To summarize, when the fraction of de novo resistant cases and the relative transmissibility are rather small,  constitutes a useful quantity for treatment policies in a single epidemic outbreak provided it can contain the overall epidemic while restraining the spread of resistance in the population.
constitutes a useful quantity for treatment policies in a single epidemic outbreak provided it can contain the overall epidemic while restraining the spread of resistance in the population.
Relative transmissibility and non-pharmaceutical interventions
It is likely that treatment alone cannot completely quell an emerging epidemic [35]. In such cases, non-pharmaceutical interventions (e.g., social distancing, case isolation, travel restrictions) could help to significantly mitigate the extent of the epidemic [3], [8], [16]. These can affect the transmissibility of the wild-type and the resistant strain while maintaining the relative transmissibility of the latter ( ). Here we investigate the competition dynamics between the wild-type and the resistant strain as a function of
). Here we investigate the competition dynamics between the wild-type and the resistant strain as a function of  , and the transmissibility of the wild-type strain
, and the transmissibility of the wild-type strain  (which varies due to non-pharmaceutical interventions) under different treatment regimes.
(which varies due to non-pharmaceutical interventions) under different treatment regimes.
The total FS is comprised by the resistant-strain cases (FS ) plus the wild-type cases (FS
) plus the wild-type cases (FS ). To determine the dominant strain, we compare FS
). To determine the dominant strain, we compare FS and FS
and FS . Figure 8 shows numerical results of FS
. Figure 8 shows numerical results of FS FS
FS in the (
in the ( ) parameter space (
) parameter space ( fixed). For instance, if FS
fixed). For instance, if FS FS
FS (gray-black region), the resistant strain is accountable for more cases than the wild-type strain. The wild-type dominated region is in red.
(gray-black region), the resistant strain is accountable for more cases than the wild-type strain. The wild-type dominated region is in red.
Figure 8. FS FS
FS in the (
in the ( ) parameter space.
) parameter space.

 is fixed, and
is fixed, and  (left to right). Gray-black regions are dominated by the resistant strain. As treatment increases the resistant strain 1) benefits from higher wild-strain transmissibility, 2) increases the range of relative transmissibility for which it can spread, and 3) expands the region in which it can extensively spread (black region).
(left to right). Gray-black regions are dominated by the resistant strain. As treatment increases the resistant strain 1) benefits from higher wild-strain transmissibility, 2) increases the range of relative transmissibility for which it can spread, and 3) expands the region in which it can extensively spread (black region).
The resistant strain can only spread in the  region for which the wild-type strain significantly spreads: notice in each graph, a vertical light-red region where only the wild-type strain minimally spreads, and to its right we see regions of coexistence. The value of
region for which the wild-type strain significantly spreads: notice in each graph, a vertical light-red region where only the wild-type strain minimally spreads, and to its right we see regions of coexistence. The value of  defining the split of these two regions,
defining the split of these two regions,  , is such that
, is such that  ; if
; if  , the wild-type strain will only generate few infections and consequently the resistant strain will mainly be in rare de novo resistant cases. A similar consideration was presented in Figure 6.
, the wild-type strain will only generate few infections and consequently the resistant strain will mainly be in rare de novo resistant cases. A similar consideration was presented in Figure 6.
In general, for lower  the resistant strain cannot spread, while the wild-type strain produces an increasingly larger number of infections as
the resistant strain cannot spread, while the wild-type strain produces an increasingly larger number of infections as  increases. As treatment (
increases. As treatment ( ) increases, the resistant-dominated region shifts to higher values of
) increases, the resistant-dominated region shifts to higher values of  , while expanding the range of
, while expanding the range of  for which it can significantly spread (darker areas).
for which it can significantly spread (darker areas).
These observations suggest that if the wild-type strain features relatively low transmission, the best strategy to contain both strains is to treat “hard and early”. However, if the transmissibility is higher and the fitness cost of resistance is low, then this strategy can have devastating consequences as the resistant strain can infect a large fraction of the population. This demonstrates the importance of effective non-pharmaceutical interventions that could reduce  .
.
For larger values of  an interesting process occurs. Starting from low
an interesting process occurs. Starting from low  the wild-type dominates. As
the wild-type dominates. As  increases – crossing the “vertical” null isocline where both FSs are equal – the resistant strain begins to prevail, until crossing the “slant” null isocline where the wild-type strain starts to regain its dominance. A possible explanation for this dominance shift is that as the wild-type strain becomes more transmissible, it depletes the pool of susceptibles too quickly, leaving the resistant strain with few individuals to infect once it emerges. However, for even larger
increases – crossing the “vertical” null isocline where both FSs are equal – the resistant strain begins to prevail, until crossing the “slant” null isocline where the wild-type strain starts to regain its dominance. A possible explanation for this dominance shift is that as the wild-type strain becomes more transmissible, it depletes the pool of susceptibles too quickly, leaving the resistant strain with few individuals to infect once it emerges. However, for even larger  and high
and high  , increasing
, increasing  also increases the FS of the resistant strain. In this scenario, the relative transmissibility is so high that even if the wild-type strain can spread rapidly, the resistant strain will eventually “catch up” and outcompete it.
also increases the FS of the resistant strain. In this scenario, the relative transmissibility is so high that even if the wild-type strain can spread rapidly, the resistant strain will eventually “catch up” and outcompete it.
Thus, when trying to predict the outcome of the competition dynamics between wild-type and resistant strains, knowing the relative transmissibility of the latter is not sufficient. One must also know the actual value of its transmissibility. In the endemic case, however, when treatment is fixed, the relative transmissibility completely determined which strain ultimately dominated (Figure 2). These considerations complement observations made in [17].
The Impact of Contact Structure
Two strong simplifications made in our model were to ignore the complex contact structure of human populations and the stochastic nature of the transmission and de novo resistance dynamics. While these assumptions allowed us to obtain closed-form solutions for effective treatment regimes, the social network underlying the epidemic process is known to have non-trivial effects on transmission dynamics [21], [22], [36], [37]. In this section we use a model equivalent to (1)–(5) that features contact structure [38] and stochasticity [39]. We again assume  , and utilize Monte-Carlo (MC) simulations to assess the effectiveness of
, and utilize Monte-Carlo (MC) simulations to assess the effectiveness of  (Eq. (14)) in single epidemic situations.
(Eq. (14)) in single epidemic situations.
To perform MC simulations of the model, we have generated networks of size  with fat-tailed degree distributions
with fat-tailed degree distributions  (distribution of number of contacts per individual, shown in Figure 9), via the Configuration Model algorithm [40]. For every generated network, a randomly chosen individual is infected with the wild-type strain and the dynamics are then simulated in discrete time:
(distribution of number of contacts per individual, shown in Figure 9), via the Configuration Model algorithm [40]. For every generated network, a randomly chosen individual is infected with the wild-type strain and the dynamics are then simulated in discrete time:
Figure 9. The fat-tailed degree distribution (contact per individual) with power-law tail and exponential cut-off.

Used to generate heterogeneous networks for the MC simulations.
each time step, every susceptible neighbor
 of every infectious individual
of every infectious individual  is infected with probability
is infected with probability  ;
;wild-type infections are treated with probability
 , leading to resistance-conferring mutation with probability
, leading to resistance-conferring mutation with probability  ;
;each time step every infectious individual
 recovers with probability
recovers with probability  (with
(with  ).
).
Figure 10 shows the variation in the final epidemic size (FS) of the system due to the contact heterogeneity and the inherent stochasticity of the disease and pathogen mutation processes. The worst-case scenarios (the highest FS obtained for a given value of  ) qualitatively follow the same behavior as the ODE model above (blue curves in Figure 6). More importantly, the predicted optimal treatment fraction
) qualitatively follow the same behavior as the ODE model above (blue curves in Figure 6). More importantly, the predicted optimal treatment fraction  provides a good approximation to what could be considered the best treatment plan, yielding the lowest total FS while halting the spreading of resistance (Figure 10, greener dots). As in the deterministic case, for
provides a good approximation to what could be considered the best treatment plan, yielding the lowest total FS while halting the spreading of resistance (Figure 10, greener dots). As in the deterministic case, for  , resistance spreads widely. Hence, when the frequency of de novo cases is small, the efficacy of the treatment fraction
, resistance spreads widely. Hence, when the frequency of de novo cases is small, the efficacy of the treatment fraction  to minimize both the epidemic size and the risks of resistance emergence, is robust to both the heterogeneity of population structure as well as the stochasticity of transmission and mutation dynamics.
to minimize both the epidemic size and the risks of resistance emergence, is robust to both the heterogeneity of population structure as well as the stochasticity of transmission and mutation dynamics.
Figure 10. Monte Carlo simulations on a network with heterogeneous contact structure.

( ). Every point represents one of over 10,000 simulations on networks of size 250 000, with color indicating the proportion of resistance in the FS (from black,
). Every point represents one of over 10,000 simulations on networks of size 250 000, with color indicating the proportion of resistance in the FS (from black,  wild-type, to green,
wild-type, to green,  resistant).
resistant).  (Eq. (14)) is shown in dashed black line. The effectiveness of
(Eq. (14)) is shown in dashed black line. The effectiveness of  is robust to stochasticity and heterogeneous contact structures. Other parameters:
is robust to stochasticity and heterogeneous contact structures. Other parameters:  and
and  , where
, where  is the average excess degree in the network [39].
is the average excess degree in the network [39].
Discussion
The rapid development of an effective vaccine against an emerging novel influenza virus presents considerable challenges. Thus, antiviral agents could play a central role as a first-line defense against emerging epidemics of influenza. The large-scale use of these drugs could, in turn, select for the evolution of drug-resistant strains [8], making the strategic distribution of antivirals essential in quelling the spread of drug-resistance while limiting the overall epidemic size. In this work we have discussed the influence of two key parameters on the effectiveness of treatment: the relative transmissibility of the drug-resistant strain ( ), and the frequency of de novo resistance (
), and the frequency of de novo resistance ( ). We extended a previous model [17] to include demography and performed analytical calculations of the reproductive numbers, stability of the fixed points, and conditions for the exclusion or coexistence of resistant and wild-type pathogen strains.
). We extended a previous model [17] to include demography and performed analytical calculations of the reproductive numbers, stability of the fixed points, and conditions for the exclusion or coexistence of resistant and wild-type pathogen strains.
In the endemic case we found that, depending on the values of  (or equivalently
(or equivalently  ) and
) and  , the optimal treatment regime will be at intermediate (case (A) for high
, the optimal treatment regime will be at intermediate (case (A) for high  and low
and low  ) or more extreme values (case (B) for low
) or more extreme values (case (B) for low  and high
and high  ). Intuitively it is clear that if the resistant strain is highly transmissible (high
). Intuitively it is clear that if the resistant strain is highly transmissible (high  ), then treatment should be moderate in order to limit the selective advantage of drug-resistant phenotypes. Conversely, if the resistant strain is weakly transmitted (low
), then treatment should be moderate in order to limit the selective advantage of drug-resistant phenotypes. Conversely, if the resistant strain is weakly transmitted (low  ), then more intense treatment regimes are preferred since resistance-only endemic levels will be relatively low. These recommendations are valid as long as infections with a wild-type or a resistant strain represent the same harm to the host (e.g., strains with similar infectious periods and virulence). In addition, we also remarked that when
), then more intense treatment regimes are preferred since resistance-only endemic levels will be relatively low. These recommendations are valid as long as infections with a wild-type or a resistant strain represent the same harm to the host (e.g., strains with similar infectious periods and virulence). In addition, we also remarked that when  is low, the optimal treatment regime can be approximated by
is low, the optimal treatment regime can be approximated by  . In the single epidemic case, numerical simulations also suggest that if
. In the single epidemic case, numerical simulations also suggest that if  and
and  are low,
are low,  is still a useful quantity when designing treatment strategies. However, in contrast to the endemic case, knowing the relative transmissibility of the resistant strain is not enough to predict the final outcome of the competition between the two strains. In this case, the strain that successfully spreads first has a significant impact on which strain infects more individuals during the epidemic. Our results also indicate that early and high treatment regimes are most effective at reducing the number of infections while hindering the rise of resistance, when the transmissibility of the wild-type strain is relatively low. This emphasizes the importance of non-pharmaceutical interventions aimed at reducing the transmission rate of the disease.
is still a useful quantity when designing treatment strategies. However, in contrast to the endemic case, knowing the relative transmissibility of the resistant strain is not enough to predict the final outcome of the competition between the two strains. In this case, the strain that successfully spreads first has a significant impact on which strain infects more individuals during the epidemic. Our results also indicate that early and high treatment regimes are most effective at reducing the number of infections while hindering the rise of resistance, when the transmissibility of the wild-type strain is relatively low. This emphasizes the importance of non-pharmaceutical interventions aimed at reducing the transmission rate of the disease.
Further, we showed that for small  , the parameter
, the parameter  is robust to the presence of contact heterogeneity and stochasticity, as it still minimizes both the epidemic size and the risks of resistance emergence. This reinforces the public health implications of the effective treatment expressions derived herein.
is robust to the presence of contact heterogeneity and stochasticity, as it still minimizes both the epidemic size and the risks of resistance emergence. This reinforces the public health implications of the effective treatment expressions derived herein.
An interesting similarity across time scales is the impact of the frequency of de novo resistance on  : as
: as  increases, the effectiveness of
increases, the effectiveness of  becomes compromised. While we give mathematical and biological arguments for this property, the inherent uncertainty in the empirical values of
becomes compromised. While we give mathematical and biological arguments for this property, the inherent uncertainty in the empirical values of  make this observation potentially relevant to the designing of treatment strategies [8].
make this observation potentially relevant to the designing of treatment strategies [8].
Our model, like any other, is not exempt of simplifying assumptions, or uncertainties about the model parameter values and transmission dynamics of wild-type and resistant strains. Thus, rather than providing specific quantitative recommendations for treatment policies, we emphasize the qualitative character of our observations. Moreover, we recognize that even if these uncertainties were resolved, we still face ethical issues when deciding to implement treatment policies based on our recommendations; e.g., treat only a certain fraction of those infected if relative transmissibility is high and de novo resistance is unlikely. This is a difficult case for the public health planner, and the choice is left to them. If relative transmissibility is low and de novo resistance is more likely, then our recommendations are less controversial: treat people as they come in based on their clinical profile. In terms of the assumptions made in our analysis, we considered that treatment and de novo resistance happen immediately after infection. In the Text S1 we present a model that features stage progressions (treatment and de novo resistance occur at certain rates rather than instantaneously) and show that its dynamics are analogous to those presented here (see Figure S1, S2, S3). We also assumed that the fraction of treated individuals can, with no regard to economic and social costs, attain any value between 0 and 1, and remain constant throughout time. This is generally not true as treatment availability and costs vary with time and socioeconomic context (models in [5], [6], [41], [42] explore different time-dependent treatment regimes). We have considered a model with equal birth and death rates, thus, it may also be important to study the impact of demographics on the effectiveness of treatment regimes, though less so in the single epidemic case. We have also excluded coinfection with both strains, which is known to affect the evolution of the influenza virus (e.g., viral reassortment [4]), and could in turn influence the development of drug-resistant phenotypes. We suspect that accounting for coinfection might lead to new and interesting dynamics.
Our results shed light on the epidemiological impact of the interplay between treatment regimes and relative transmissibility of a strain of influenza resistant to antiviral treatment and the frequency of de novo resistance, both aspects which are difficult to assess empirically. These findings could have important implications for the strategic distribution of antivirals in a population in response to the emergence of a novel influenza strain.
Supporting Information
Compartmental diagram for the analogous model.
(EPS)
Comparison of the two models in the endemic case.
(EPS)
Comparison of the two models in the single epidemic case.
(EPS)
Transcritical Bifurcation between DFE and CFP.
(EPS)
Transcritical Bifurcation between RFP and CFP.
(EPS)
Transcritical Bifurcation between the DFE and the RFP.
(EPS)
Transcritical Bifurcation between RFP and CFP.
(EPS)
Stability behavior of the system.
(EPS)
Prevalence as a function of  and
and  .
.
(EPS)
 for
for  and for
and for  .
.
(EPS)
Analytical derivation of reproduction numbers; analogous model; analytical derivations regarding the stability of the system; and analytical derivations regarding the optimal treatment regimes.
(PDF)
Acknowledgments
The authors would like to thank the reviewers for insightful comments that helped improve the manuscript. We also thank Andrés Gómez-Liévano for helpful discussions. Finally, we wish to thank the Santa Fe Institute and its Complex Systems Summer School at which this work was initiated.
Funding Statement
This work was supported by WAESOBD LSAMPBD NSF Cooperative Agreement HRD-1025879 (OPL); The Natural Sciences and Engineering Research Council of Canada (LHD); INET grant (no. IN01100005) (GMG); NSF Graduate Research Fellowship (grant no. DGE-0707427) (BMA). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Ferguson N, Galvani A, Bush R (2003) Ecological and immunological determinants of influenza evolution. Nature 422: 428–433. [DOI] [PubMed] [Google Scholar]
- 2. Bedford T, Rambaut A, Pascual M (2012) Canalization of the evolutionary trajectory of the human influenza virus. BMC biology 10: 38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Ferguson N, Cummings D, Cauchemez S, Fraser C, Riley S, et al. (2005) Strategies for containing an emerging influenza pandemic in southeast asia. Nature 437: 209–214. [DOI] [PubMed] [Google Scholar]
- 4. Nelson M, Holmes E (2007) The evolution of epidemic influenza. Nature reviews genetics 8: 196–205. [DOI] [PubMed] [Google Scholar]
- 5.Jaberi-Douraki M, Hefffernan J, Wu J, Moghadas S (2012) Optimal treatment profile during an influenza epidemic. Differential Equations and Dynamical Systems : 1–16.
- 6. Moghadas S (2008) Management of drug resistance in the population: influenza as a case study. Proceedings of the Royal Society B: Biological Sciences 275: 1163–1169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Alexander M, Bowman C, Feng Z, Gardam M, Moghadas S, et al. (2007) Emergence of drug resistance: implications for antiviral control of pandemic influenza. Proceedings of the Royal Society B: Biological Sciences 274: 1675–1684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Regoes R, Bonhoeffer S (2006) Emergence of drug-resistant influenza virus: population dynamical considerations. Science 312: 389–391. [DOI] [PubMed] [Google Scholar]
- 9. Weinstock DM, Zuccotti G (2009) The evolution of influenza resistance and treatment. JAMA 301: 1066–9. [DOI] [PubMed] [Google Scholar]
- 10. Levin B, Lipsitch M, Bonhoeffer S (1999) Population biology, evolution, and infectious disease: convergence and synthesis. Science 283: 806–809. [DOI] [PubMed] [Google Scholar]
- 11. Lipsitch M, Plotkin JB, Simonsen L, Bloom B (2012) Evolution, safety, and highly pathogenic influenza viruses. Science 336: 1529–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Herfst S, Schrauwen EJA, Linster M, Chutinimitkul S, de Wit E, et al. (2012) Airborne transmission of influenza a/h5n1 virus between ferrets. Science 336: 1534–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Russell CA, Fonville JM, Brown AEX, Burke DF, Smith DL, et al. (2012) The potential for respiratory droplet-transmissible a/h5n1 influenza virus to evolve in a mammalian host. Science 336: 1541–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Imai M, Watanabe T, Hatta M, Das SC, Ozawa M, et al. (2012) Experimental adaptation of an influenza h5 ha confers respiratory droplet transmission to a reassortant h5 ha/h1n1 virus in ferrets. Nature 486: 420–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Althouse BM, Bergstrom TC, Bergstrom CT (2010) A public choice framework for controlling transmissible and evolving diseases. Proc Natl Acad Sci U S A 107 Suppl 11696–701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Ferguson N, Cummings D, Fraser C, Cajka J, Cooley P, et al. (2006) Strategies for mitigating an influenza pandemic. Nature 442: 448–452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Lipsitch M, Cohen T, Murray M, Levin BR (2007) Antiviral resistance and the control of pandemic influenza. PLoS Medicine 4: e15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Wu JT, Leung GM, Lipsitch M, Cooper BS, Riley S (2009) Hedging against antiviral resistance during the next influenza pandemic using small stockpiles of an alternative chemotherapy. PLoS Med 6: e1000085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. zur Wiesch P, Kouyos R, Engelstädter J, Regoes R, Bonhoeffer S (2011) Population biological principles of drug-resistance evolution in infectious diseases. The Lancet infectious diseases 11: 236–247. [DOI] [PubMed] [Google Scholar]
- 20. Lipsitch M, Bergstrom CT, Levin BR (2000) The epidemiology of antibiotic resistance in hospitals: paradoxes and prescriptions. Proc Natl Acad Sci U S A 97: 1938–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Keeling MJ, Rohani P (2008) Modeling Infectious Diseases in Humans and Animals. Princeton University Press.
- 22. Althouse BM, Patterson-Lomba O, Goerg GM, Hébert-Dufresne L (2013) The timing and targeting of treatment in influenza pandemics inuences the emergence of resistance in structured populations. PLoS Computational Biology 9(2): e1002912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Schwartz B, Bell DM, Hughes JM (1997) Preventing the emergence of antimicrobial resistance. a call for action by clinicians, public health officials, and patients. JAMA 278: 944–5. [DOI] [PubMed] [Google Scholar]
- 24.WHO (2006) Who rapid advice guidelines on pharmacological management of humans infected with avian influenza a (h5n1) virus. Technical report,World Health Organization.
- 25. Fiore AE, Fry A, Shay D, Gubareva L, Bresee JS, et al. (2011) Antiviral agents for the treatment and chemoprophylaxis of influenza – recommendations of the advisory committee on immunization practices (acip). MMWR Recomm Rep 60: 1–24. [PubMed] [Google Scholar]
- 26. Qiu Z, Feng Z (2010) Transmission dynamics of an influenza model with vaccination and antiviral treatment. Bulletin of mathematical biology 72: 1–33. [DOI] [PubMed] [Google Scholar]
- 27. Lipsitch M (2001) The rise and fall of antimicrobial resistance. Trends Microbiol 9: 438–44. [DOI] [PubMed] [Google Scholar]
- 28. Handel A, Regoes R, Antia R (2006) The role of compensatory mutations in the emergence of drug resistance. PLoS computational biology 2: e137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Anderson R, May R (1992) Infectious Diseases of Humans: Dynamics and Control (Part 1, Chapters 4–7). Oxford Science Publications. Oxford University Press.
- 30. Lee N, Chan PKS, Hui DSC, Rainer TH, Wong E, et al. (2009) Viral loads and duration of viral shedding in adult patients hospitalized with influenza. J Infect Dis 200: 492–500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Ling LM, Chow AL, Lye DC, Tan AS, Krishnan P, et al. (2010) Effects of early oseltamivir therapy on viral shedding in 2009 pandemic influenza a (h1n1) virus infection. Clin Infect Dis 50: 963–9. [DOI] [PubMed] [Google Scholar]
- 32. Nicholson KG, Aoki FY, Osterhaus AD, Trottier S, Carewicz O, et al. (2000) Efficacy and safety of oseltamivir in treatment of acute influenza: a randomised controlled trial. neuraminidase inhibitor flu treatment investigator group. Lancet 355: 1845–50. [DOI] [PubMed] [Google Scholar]
- 33. Van den Driessche P, Watmough J (2002) Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Mathematical Biosciences 180: 29–48. [DOI] [PubMed] [Google Scholar]
- 34.Strogatz SH (1994) Nonlinear dynamics and Chaos: with applications to physics, biology, chemistry, and engineering. Addison-Wesley Pub.
- 35. Longini I, Nizam A, Xu S, Ungchusak K, Hanshaoworakul W, et al. (2005) Containing pandemic influenza at the source. Science 309: 1083–1087. [DOI] [PubMed] [Google Scholar]
- 36. Pastor-Satorras R, Vespignani A (2001) Epidemic spreading in scale-free networks. Phys Rev Lett 86: 3200–3. [DOI] [PubMed] [Google Scholar]
- 37. Read JM, Keeling MJ (2003) Disease evolution on networks: the role of contact structure. Proc Biol Sci 270: 699–708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Barabasi AL, Albert R (1999) Emergence of scaling in random networks. Science 286: 509–12. [DOI] [PubMed] [Google Scholar]
- 39. Newman MEJ (2002) Spread of epidemic disease on networks. Phys Rev E Stat Nonlin Soft Matter Phys 66: 016128. [DOI] [PubMed] [Google Scholar]
- 40.Newman M (2010) Networks: an introduction. Oxford University Press, Inc.
- 41. Hansen E, Day T (2011) Optimal antiviral treatment strategies and the effects of resistance. Proc Biol Sci 278: 1082–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Moghadas S, Bowman C, Röst G, Wu J (2008) Population-wide emergence of antiviral resistance during pandemic influenza. PLoS One 3: e1839. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Compartmental diagram for the analogous model.
(EPS)
Comparison of the two models in the endemic case.
(EPS)
Comparison of the two models in the single epidemic case.
(EPS)
Transcritical Bifurcation between DFE and CFP.
(EPS)
Transcritical Bifurcation between RFP and CFP.
(EPS)
Transcritical Bifurcation between the DFE and the RFP.
(EPS)
Transcritical Bifurcation between RFP and CFP.
(EPS)
Stability behavior of the system.
(EPS)
Prevalence as a function of and .
(EPS)
for and for .
(EPS)
Analytical derivation of reproduction numbers; analogous model; analytical derivations regarding the stability of the system; and analytical derivations regarding the optimal treatment regimes.
(PDF)