

Abstract
The protective efficacy of DNA plasmids encoding avian infectious bronchitis virus (IBV) S1, N, or M protein was investigated in chickens. Chickens were inoculated monovalently (with plasmid pVAX1-16S1, pVAX1-16M, or pVAX1-16N alone) or multivalently (combination of the three different plasmids, pVAX1-16S1/M/N). A prime-boost immunization protocol against IBV was developed. Chickens were immunized with the multivalent DNA vaccine twice and then boosted with an inactivated vaccine once. Antibody titers of the chickens immunized with pVAX1-16S1/M/N were much higher than those of the monovalent groups (p < 0.01). A protective rate up to 90% was observed in the pVAX1-16S1/M/N group. The serum antibody titers in the prime-boost birds were significantly higher than those of the multivalent DNA vaccine group (p < 0.01) but not significantly different compared to the inactivated vaccine group at 49 days of age. Additionally, the prime-boost group also showed the highest level of IBV-specific cellular proliferation compared to the monovalent groups (p < 0.01) but no significant difference was found compared to the multivalent DNA vaccine group, and the prime-boost group completely protected from followed viral challenge.
Keywords: IBV, multivalent DNA vaccine, prime-boost
Introduction
Avian infectious bronchitis (IB) is a major disease in the poultry industry worldwide. This disease frequently occurs in vaccinated and non-vaccinated flocks, and has caused severe economic loss over the last few years [11-14,17,21]. Protection failure is mainly due to the numerous IB virus (IBV) serotypes and frequent emergence of new variants [23,26]. Given the economic effects of IBV on the commercial poultry industry, preventing infection with this virus has been continually pursued. Traditional IBV vaccines include inactivated or live attenuated vaccines [2-4]. However, both types of vaccines have unique disadvantages. Inactivated vaccines can induce the production of relatively high antibody titers but a very low level of cytotoxic T lymphocyte (CTL) responses [5]. Live attenuated vaccines can initiate humoral and cellular immune responses, but lead to the spread the live vaccine viruses [7,15]. It is therefore necessary to develop novel vaccines and/or optimize current vaccination procedures.
IBV belongs to the Coronaviridae family and contains a positive single-stranded RNA genome encoding four major structural proteins: a small envelope protein (E), integral membrane protein (M), phosphorylated nucleocapsid protein (N), and spike glycoprotein (S) [19]. The S protein is cleaved into two subunits (S1 and S2). The S1 protein is very important for inducing protective immunity and has been successfully used to construct IBV DNA vaccines [6,9]. The N protein is conserved and induces CTL as well as activated B cell responses, which are critical for preventing IBV infection in poultry [8,20]. The M glycoprotein can induce the production of detectable antibodies and delayed type hypersensitivity responses [8]. Hence, all of these proteins are primary targets for developing DNA vaccines to elicit immune responses.
In the present study, we evaluated the protective effect of three plasmids expressing the S1, N, and M proteins of the virulent IBVSX16 strain that we previously constructed [22]. Chickens were immunized monovalently with each individual plasmid (pVAX1-16S1, pVAX1-16M, and pVAX1-16N) or multivalently with a combination of the three different plasmids (pVAX1-16S1/M/N). To improve IBV vaccine efficacy, the chickens were immunized with a multivalent DNA vaccine followed by boosting with an inactivated IBV vaccine before being challenged with virulent IBV.
Materials and Methods
DNA vaccines, virus, and experimental animals
Plasmids pVAX1-16S1, pVAX1-16N, and pVAX1-16M encoding the S1, N, and M proteins of the virulent IBVSX16 strain, respectively, were described in our previous publication [22]. The virulent IBVSX16 strains used to challenge immunized chickens in this study were isolated from the kidneys of IB-infected chickens from the Shanxi province by Department of infectious Disease and immunology, College of Animal Science and Technology, Shanxi Agricultural University [21]. Virus stocks containing 1 × 103 egg infective dosage (EID50) of IBVSX16 with 100 µL were used to inoculate the allantoic cavities of 10-day-old specific pathogen free (SPF) embryonated chicken eggs (Shandong Specific-Pathogen-Free Chicken Research Center, China) that were then kept at 37℃ for 48 h. Allantoic fluid containing the virus was harvested after 48 h post-inoculation, stored at -80℃ until use. Chill eggs at 4℃ for at least 2 h to kill the embryo and to reduce the contamination of the allantoic fluid with blood during harvesting. Remove sticky tape and swab each egg with cotton wool soaked with 70% alcohol to disinfect and remove condensation from the shells. Dip the forceps or scissors in absolute alcohol and flame to sterilize. Remove the eggshell above the air space. Discard embryos that are visibly contaminated. Remove a sample of allantoic fluid from each egg. The 50% EID50/mL of the viral stocks were calculated as previously described by Reed and Muench [25]. Titer of the IBVSX16 strains was 109 EID50/mL. A total of 140 seven-day-old SPF chickens (Shandong Specific-Pathogen-Free Chicken Research Center, China) were housed in SPF environment at the Laboratory Animal and Resources Facility, Shanxi Agricultural University.
Inactivated vaccine
The inactivated vaccine was made by adding 37% formaldehyde (final concentration, 0.1%) to allantoic fluid containing IBVSX16 and incubating at 37℃ for 24 h. The inactivated vaccine 200 µL was inoculated into the allantoic cavity of 10-day-old SPF embryonated chicken eggs. The embryos were incubated at 37℃ and examined twice daily for their viability. The allantoic fluids were harvested after 72 h and two blind passages were conducted to examine the efficiency of IBVSX16 inactivation. One part of the inactivated allantoic fluid was then emulsified with two parts (v/v) of paraffin oil (Hangzhou Oil Refinery, China).
Immunization of chickens
Our animal research in our study had been approved by Shanxi Province Animal Disease Control Center (China). The plasmids used were amplified in Escherichia (E.) coli DH5α cells (TaKaRa, Japan), and extracted using a PureYield Plasmid Maxiprep System (Promega, USA). Seven-day-old chickens were randomly divided into seven groups of 20 chickens each and immunized intramuscularly on 7, 21, and 35 day-old, using different vaccination strategies (Table 1). Each group of chickens was injected with 100 µg (1 µg/µL) monovalent DNA vaccine (pVAX1-16S1, pVAX1-16M, and pVAX1-16N) respectively and 0.5 mL inactivated IBV vaccine. pVAX1-16S1/M/N was a multivalent DNA vaccine containing 100 µg of each plasmid (equivalent molar ratios for each DNA component) and therefore delivered the same dose of each expression plasmid targeting the S1, N, or M, respectively, as each monovalent vaccine. All the chickens were immunized intramuscularly with the vaccines.
Table 1.
Immunization schedule
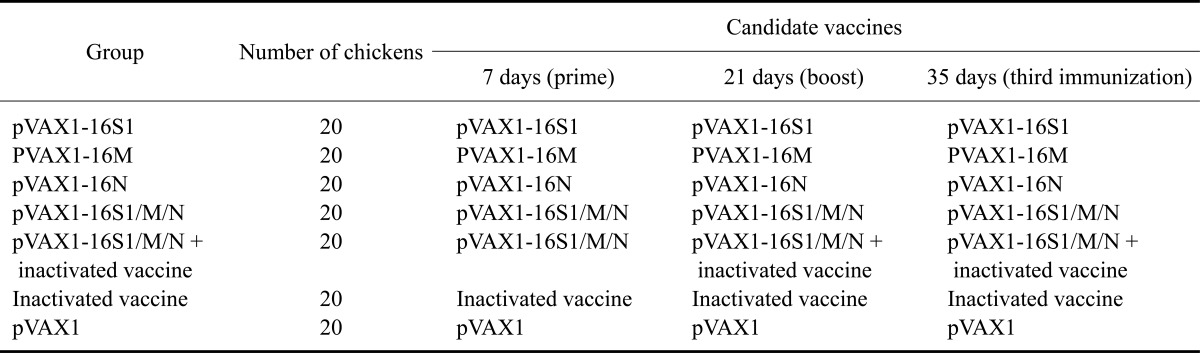
S1: spike protein, N: nucleocapsid protein, M: membrane protein.
Peripheral blood samples were also collected from five randomly selected chickens from each group from the jugular vein into heparinized capillary tubes (Laiwu Yaohua Pharmaceutical Packing, China) when the birds were 7, 21, 35, and 49 days old.
Detection of anti-IBV antibodies
Titers of antibodies against IBV in the serum samples were measured using an enzyme-linked immunosorbent assay (ELISA) kit (Idexx Laboratories, USA) according to the manufacturer's instructions. The optical density at 650 nm (OD650) was measured using a microplate reader (model 680; Bio-Rad, USA). Each serum sample, including the negative and positive controls, was analyzed in triplicate. Negative and positive sera for IBV were obtained from China Institute of Veterinary Drugs Control. Titers of virus neutralizing antibody against IBV were also measured as previously described [25]. Briefly, serial 2-fold dilutions of serum samples were mixed with 100 EID50 of IBV16 strain and kept at room temperature for 1 h. 200 µL the virus-serum mixtures in each dilution were inoculated into the allantoic cavity of 10-day-old SPF chicken embryos in ten replicates. The embryos were also inoculated with 100 EID50 alone in parallel. Seven days after inoculation, characteristic IBV lesions such as dwarfing, stunting, or curling of embryos were examined. The virus neutralizing antibody titer of each sample was recorded at the highest serum dilution value which protected 50% of the embryos from death.
Lymphocyte proliferation assay
A T cell proliferation assay was performed as previously described [1] with some modification. Peripheral blood mononuclear cells (PBMCs) were isolated by Lympholyte-Mammal (Cedarlane, Canada). Briefly, 2 mL freshly heparinized whole blood was diluted with Hanks' balanced salt solution in a ratio of 1:1 and carefully add on top of 2 mL lympholyte-mammal separation medium, centrifuge at 400 × g/min for 20 min, aspirate coat of leukocyte, add 4 mL KPMI 1640 (Invitrogen, USA), centrifuge at 400 × g/min for 20 min, discard supernatant and wash precipitation cell twice by the same method. The cells were resuspended in RPMI 1640 medium (Invitrogen, USA) supplemented with 5% fetal bovine serum (Invitrogen, USA), 100 µg/mL penicillin (Invitrogen, USA), and 100 µg/mL streptomycin (Invitrogen, USA) at a concentration of 1 × 107 cells/mL. The freshly isolated PBMCs (106 cells/mL) were seeded in U-bottomed 96-well microtiter plates (Costar; Corning, USA) and incubated with either 1 × 103 EID50 of IBVSX16 strain or 10 µg/mL concanavalin A (ConA; Sigma-Aldrich, USA) as a positive control, or RPMI-FBS alone in a final volume of 200 µL/well. Each blood sample was tested in triplicate. The microtiter plates were incubated for 48 h at 40℃ in a humidified atmosphere containing 5% CO2. After incubation, 10 µL of an MTT [3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] solution (5 mg/mL; Sigma, USA) was added into each well. The plates were then incubated at 40℃ for another 4 h. The culture medium was removed by aspiration, and 100 µL of dimethyl sulfoxide (Sigma, USA) were added to each well to lyse the cells. The absorbance of each well was measured at 570 nm (A570) with an ELISA microplate reader (model 680; Bio-Rad, USA). Mean A570 values were used as indicators of peripheral T lymphocyte proliferation.
Analysis of T lymphocyte subsets (CD3+, CD4+, and CD8+)
PBMCs were isolated from blood by lympholyte-mammal (Cedarlane, Canada) and adjusted to 1 × 107 cells / mL with RPMI 1640 medium. The samples (100 µL; 1 × 106 cells) was incubated with mouse anti-chicken CD4-PE (BD Biosciences, USA), mouse anti-chicken CD3-SPRD (BD Biosciences, USA), and mouse anti-chicken CD8-FITC (BD Biosciences, USA) primary antibodies (antibody final concentrations as 1.25 µg/100 µL) in the dark at 4℃ for 30 min. After the incubation, all samples were washed three times with cold PBS and then resuspended in 0.5 mL of PBS. The single-color positive controls were prepared by staining representative cells with mouse anti-chicken CD4-PE, mouse anti-chicken CD3-SPRD, and mouse anti-chicken CD8-FITC antibodies individually and used for estimation of proper compensation. The unstained cell sample was used as a negative control to adjust the correct voltage. The percentages of CD3+CD4+, and CD3+CD8+ lymphocytes were determined on flow cytometry (Beckman Coulter, USA).
Virus challenge
Three weeks after the third vaccination, all chickens were challenged with 103 EID50 of IBVSX16 through a nasal-ocular route. The chickens were observed daily for 14 days post-challenge. Deceased chickens were necropsied and evaluated for IBV infection. The surviving chickens were euthanized humanely by pectoral muscle injection of ketamine hydrochloride in 22 mg/kg of body weight 14 days post-infection. The trachea and kidneys were collected for virus detection. The number of IBV-positive chickens was confirmed by reverse transcriptase-polymerase chain reaction (RT-PCR). All procedures for RNA isolation and RT-PCR were previously described [15]. Viral RNA was isolated and purified from trachea and kidneys collected from chickens challenged with IBVSX16 using Trizpl reagents (Invitrogen, USA) according to the manufacturer's instructions. Amplification of the specific fragments of N gene by RT-PCR was performed. The RNA pellet was dissolved in 10 mL of DEPC-treated water. The first-strand cDNA was synthesized using the PrimeScript 1st Strand cDNA Synthesis Kit (Takara, Japan). The PCR reaction (50 µL) contained 1 µL of cDNA (about 100 ng), 5 µL of 10× PCR buffer, 4 µL of 2.5 mmol/L dNTP, 2 µL of 10 mmol/L of each of the two primers, 0.25 µL of 2 U/µL Taq DNA polymerase, and 37.75 µL of H2O. The PCR conditions for amplification were 94℃ for 5 min, 30 cycles of 94℃ for 40 sec, 60℃ for 40 sec, and 72℃ for 100 sec, followed by 72℃ for 10 min. The products were analyzed on 1.0% agarose gel. Mortality was calculated by dividing the number of dead chickens by the total number of chickens in each group. The level of protection was determined as a percentage by dividing the number of IBV-negative chickens by the total number of chickens of each group.
Statistical analysis
Statistical analysis was performed with SPSS (ver. 17.0; IBM, USA). Differences between the groups were analyzed with a one-way repeated measurement ANOVA and least significance difference test.
Results
Antibody responses to IBV in the chickens
The concentrations of serum anti-IBV antibodies were measured with a serum-neutralizing assay and an ELISA. Mean serum IBV-neutralizing antibody titers of the different groups immunized with different vaccines as shown in Fig. 1. Values are expressed as the log 10 inverse mean titer ± SD for 10 chickens in each group. Different letters above the columns indicate significant differences (p < 0.01) while identical letters indicate that there was no significant difference (p > 0.05) within each time point. The titers measured at each time point were significantly different in each group (p < 0.01). As expected, sera from the pre-primed chickens and birds immunized with the pVAX1 vector had no detectable levels of antibodies against IBV according to the serum-neutralizing assay (Fig. 1). Antibody titers for chickens immunized with pVAX1-16S1, pVAX1-16M, pVAX1-16N, or pVAX1-16S1/M/N increased from day 7 to day 49 (Fig. 1). Antibody titers of the multivalent pVAX1-16S1/M/N-immunized chickens were significantly higher than those of all chickens immunized with the monovalent DNA vaccine on days 21, 35, and 49 (p < 0.01). These results indicate that the triple-gene plasmid mix induced a greater antibody response than the single-gene plasmid.
Fig. 1.

Mean serum avian infectious bronchitis virus (IBV)-neutralizing antibody titers of the different groups immunized with different vaccines. Different letters above the columns indicate significant differences (p < 0.01) while identical letters indicate that there was no significant difference (p > 0.05) within each time point. All the mean serum IBV-neutralizing antibody titers between different time points were significantly different (p < 0.01).
Antibody titers of the pVAX1-16S1/M/N + inactive vaccine group were much higher than those of the pVAX1-16S1/M/N group (p < 0.01) on days 21 and 35. However, no significant difference (p > 0.05) in antibody titers between the two groups was observed on day 49. These findings were also confirmed with an ELISA assay as shown in Fig. 2. Data are expressed as the mean ± SD for 10 chickens in each group. Different letters above the columns indicate significant differences (p < 0.01) while identical letters indicate that there was no significant difference (p > 0.05) within each time point. The titers measured at each time point were significantly different in each group. (p < 0.01).
Fig. 2.

Mean ELISA antibody titers for the different groups immunized with different vaccines. Data are expressed as the mean ± SD for 10 chickens in each group. Each column is labeled with a letter. Different letters indicate that the differences between individuals in that day group in antibody titers are significant (p < 0.01). Absence of a letter indicates that no significant differences (p > 0.05) were observed between any of the time points. Although not indicated in the graph, the differences between corresponding treatments from different day groups are significant (p < 0.01).
Lymphocyte proliferation assay results
Kinetic changes in A570 values for the animals are shown in Fig. 3. Data are expressed as the mean OD570 values ± SD (n = 5). Columns are labeled with letters. Different letters indicate statistically significance differences (p < 0.01) between different treatments within the time point while columns with the same letters indicate that no significantly differences (p > 0.05) were observed. Absence of a letter indicates that there were no significant differences (p > 0.05) between any of the time points. Similar to the antibody responses we observed, PBMC proliferation in response to IBV and ConA stimulation was also observed in chickens immunized with pVAX1-16S1, pVAX1-16M, pVAX1-16N, and pVAX1-16S1/M/N on day 21 and further increased on day 35. A570 values for the monovalent and multivalent groups were much higher than those for the pVAX1 DNA vector control and inactivated vaccine groups when the birds were 35 days old. The A570 values for the chicken immunized with the pVAX1-16N construct were higher than those for birds immunized with the pVAX1-16S1 or pVAX1-16M constructs (p < 0.01). On day 49, proliferative responses of the pVAX1-16S1/M/N and pVAX1-16S1/M/N + inactivated vaccine groups were significantly higher than those of the other groups (p < 0.01). These results indicate that immunization with a combination of the three-gene DNA vaccine and an inactivated vaccine not only elicited the strongest antibody response, but also induced the highest IBV-specific cellular proliferation rates in the chickens. The responses of all experimental groups to stimulation with ConA were similar to ones provoked by IBV (data not shown).
Fig. 3.

Lymphocyte proliferation rates of chickens from the different groups administered different vaccines. Data are expressed as the mean OD570 values ± SD (n = 5). Columns are labeled with letters. Different letters indicate statistically significant differences (p < 0.01) between different treatments within the time point while columns with the same letters indicate that no significantly differences (p > 0.05) were observed. Absence of a letter indicates that there were no significant differences (p > 0.05) between any of the time points. Although not indicated in the graph, the differences between corresponding treatments from different day groups are significant (p < 0.01).
T lymphocyte subset analysis
Percentages of CD3+CD4+ and CD3+CD8+ T lymphocytes among the various groups are presented in Fig. 4. Columns are labeled with letters. Different letters indicate statistically significant differences (p < 0.01) while letters in common indicated that no significant differences (p > 0.01) were observed in each T cell subgroup. No significant difference in the two T lymphocyte subgroups was found between the control pVAX1 and inactivated vaccine group. In contrast, a significant difference in the percentage of CD3+CD4+ and CD3+CD8+ T lymphocytes (p < 0.01) was observed between the pVAX1-16S1, pVAX1-16M, pVAX1-16N, and pVAX1-16S1/M/N groups compared to the pVAX1 group. Levels of CD3+CD4+ and CD3+CD8+ T lymphocytes were significantly higher in the pVAX1-16N group than in the pVAX1-16S1 or pVAX1-16M groups (p < 0.01). The highest percentage of CD3+CD4+ and CD3+CD8+ T lymphocytes were detected in the pVAX1-16S1/M/N group and pVAX16S1/M/N + vaccinated group.
Fig. 4.

Percentages of CD3+CD4+ and CD3+CD8+ T lymphocytes in the different groups of chickens. Columns are labeled with letters. Different letters indicate statistically significant differences (p < 0.01) while letters in common indicated that no significant differences (p > 0.01) were observed in each T cell subgroup.
Protection against IBV challenge
To evaluate the protective efficacy of the DNA vaccines and prime-boost vaccination strategy we developed, all chickens were challenged with virulent IBVSX16 virus. Mortality and protection rates for the different groups are shown in Table 2. The number of chickens positive for IBV was determined by RT-PCR analysis of birds that died from viral infection and ones that were euthanized. Mortality was determined as the number of dead chickens divided by the total number of chickens in each group. Protection rate was measured as a percentage of the number of chickens negative for IBV over the total number of chickens. Chickens in the pVAX1-inocluated group developed clinical signs of infection including coughing, nasal discharge, and dyspnea. Mortality rate of the pVAX1-injected groups was 35% 14 days post-challenge. In contrast, the protection rates were 75% for the pVAX1-16S1 group, 60% for the pVAX1-16M group, 80% for the pVAX1-16N group, 90% for the pVAX1-16S1/M/N group, 80% for birds given the inactivated vaccine (80%), and 100% for the pVAX1-16S1/M/N + inactivated vaccine animals. The mortality rate was 5% for the pVAX1-16S1/M/N group, 10% for the inactivated vaccine group, and 0% for birds treated with pVAX1-16S1/M/N + inactivated vaccine.
Table 2.
Mortality and protection rates of the different groups following challenge with the virulent IBVSX16 strain

Compared to the pVAX1-16S1/M/N-immunized group, chickens in only the inactivated vaccine group developed more conspicuous and severe symptom and pathological changes. The kidney parenchyma was pale, swollen, and mottled. Furthermore, chickens in the pVAX1-16S1/M/N + inactivated vaccine group were efficiently protected following viral challenge and showed no clinical signs or death. To evaluate IBV levels after challenge, presence of the virus in the kidney and trachea was detected by RT-PCR. Results of this analysis showed that 10% of the pVAX1-16S1/M/N group and 20% of the birds receiving the inactive vaccine were positive. All chickens in the pVAX1 group were positive according to the RT-PCR analysis. However, none of the birds in the pVAX1-16S1/M/N + inactivated vaccine group had detectable levels of virus. These results indicated that chickens primed with the multivalent DNA vaccine and boosted with an inactivated vaccine are effectively protected from challenge with virulent IBVSX16 virus.
Discussion
In the present study, a prime-boost vaccination regimen that completely protected chickens against challenge with a virulent IBVSX16 strain was developed. Immune responses were evaluated in chickens immunized with the DNA vaccines either alone or with a mixture of the three vaccines. Findings from the present study agree with those of a previous investigation showing that all three IBV proteins selected for this study have their own unique and important roles in eliciting IBV immune responses [24]. The DNA vaccination with pVAX1-16M alone provoked the weakest immune response demonstrated by the lowest antibody titers (p < 0.01) in comparison with pVAX1-16S1 and pVAX1-16N. In contrast, pVAX1-16S1 induced the highest antibody titers among the three constructs. Chickens immunized with pVAX1-16N had significantly higher levels of IBV-specific cellular proliferation (p < 0.01) and T lymphocytes (p < 0.01) than birds immunized with pVAX1-16S1 and pVAX1-16M. The challenge assay also proved that the DNA vaccine targeting the N protein is more effective. Chickens that received a combination of the three DNA vaccines also mounted stronger immune responses than the birds immunized with each DNA vaccines alone, suggesting that a combined DNA vaccination with S1, M, and N genes may provide stronger protection against IBV.
CD4+ T cells may directly produce antiviral cytokines, which increasing B cell activity and increasing the proliferation, maturation, and functional activity of CD8+ CTLs. The CD8+ CTL plays a critical role in controlling IBV infection [18]. At 49 days of age, chickens immunized with plasmid constructs groups had percentages of CD4+CD3+ and CD8+CD3+ T lymphocytes subgroups higher than those of birds administered the control pVAX1 vector or inactivated vaccine (p < 0.01). Increased number of T lymphocytes indicates that administration of the DNA vaccine resulted in effective cellular immunization and promoted virus clearance [2].
As for the choice of a booster, the inactivated vaccine is prepared from the whole virus particle, so the immune responses induced by inactivated IBV vaccines could respond to the whole virus. The inactivated vaccine has a better capability than choosing DNA plasmid as the booster to evoke memory B cells [16]. The inactivated vaccine as a booster is also considered to be able to balance the Th1 and Th2 immune responses [10], which could enhance both specific cell-mediated immunity and neutralizing antibody inductions with increased T cell response. In the current study, the DNA vaccine stimulated a relatively strong IBV-specific cellular response but low antibody titers. When the DNA vaccine and inactivated vaccine were administered in combination, both higher cellular proliferation rates and antibody titers were observed among the DNA vaccine-primed chickens as seen in previous studies [10]. Chickens that received the DNA vaccine with the inactivated vaccine were fully protected from IBVSX16 challenge. None of these animals developed clinical signs or harbored detectable levels of virus in the trachea or kidney after challenge. These results clearly demonstrate that only the combined administration of DNA vaccine and inactivated vaccine, but not either alone, offered full protection from challenge with the IBVSX16 strain.
Our results directly demonstrated that the multivalent DNA vaccine targeting the S1, M, and N proteins of IBV developed humoral and cellular immune responses against IBV in chickens. We also showed that our prime-boost immunization strategy induced strong immune responses and provided effective protection from challenge with a high dose of virulent IBV. DNA vaccines are easy to prepare and inactivated vaccines are readily available. Administration of DNA vaccines with an inactivated vaccine is a safe and effective immunization technique. Given its superior efficacy, this combination represents a valuable strategy for protecting against IBV infection and can combat poor cross-protection among the numerous IBV serotypes along with the frequent emergence of new variants.
Acknowledgments
The study was funded by the Scientific and Technological Research Program of Shanxi Province (grant no. 20120311023-5), the Natural Science Foundation of Shanxi Province (grant no. 2007011083), Science and Technology Development Program for Colleges and Universities in Shanxi Province (grant no. 200611014), and a post-doctoral fellowship from Shanxi Agricultural University, China. The authors thank Dr. Kerong Shi for providing assistance in revising the manuscript.
References
- 1.Barta O, Barta V, Pierson FW. Optimum conditions for the chicken lymphocyte transformation test. Avian Dis. 1992;36:945–955. [PubMed] [Google Scholar]
- 2.Bijlenga G, Cook JKA, Gelb J, Jr, de Wit JJ. Development and use of the H strain of avian infectious bronchitis virus from the Netherlands as a vaccine: a review. Avian Pathol. 2004;33:550–557. doi: 10.1080/03079450400013154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Britton P, Cavanagh D. Avian coronavirus diseases and infectious bronchitis virus vaccine development. In: Thiel V, editor. Coronaviruses: Molecular and Cellular Biology. 1st ed. Norfolk: Caister Academic Press; 2007. pp. 161–181. [Google Scholar]
- 4.Cavanagh D. Severe acute respiratory syndrome vaccine development: experiences of vaccination against avian infectious bronchitis coronavirus. Avian Pathol. 2003;32:567–582. doi: 10.1080/03079450310001621198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cavanagh D, Casais R, Armesto M, Hodgson T, Izadkhasti S, Davies M, Lin F, Tarpey I, Britton P. Manipulation of the infectious bronchitis coronavirus genome for vaccine development and analysis of the accessory proteins. Vaccine. 2007;25:5558–5562. doi: 10.1016/j.vaccine.2007.02.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cavanagh D, Darbyshire JH, Davis P, Peters RW. Induction of humoral neutralising and haemagglutination-inhibiting antibody by the spike protein of avian infectious bronchitis virus. Avian Pathol. 1984;13:573–583. doi: 10.1080/03079458408418556. [DOI] [PubMed] [Google Scholar]
- 7.Farsang A, Ros C, Renström LHM, Baule C, Soós T, Belák S. Molecular epizootiology of infectious bronchitis virus in Sweden indicating the involvement of a vaccine strain. Avian Pathol. 2002;31:229–236. doi: 10.1080/03079450220136530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ignjatovic J, Sapats S. Identification of previously unknown antigenic epitopes on the S and N proteins of avian infectious bronchitis virus. Arch Virol. 2005;150:1813–1831. doi: 10.1007/s00705-005-0541-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kapczynski DR, Hilt DA, Shapiro D, Sellers HS, Jackwood MW. Protection of chickens from infectious bronchitis by in ovo and intramuscular vaccination with a DNA vaccine expressing the S1 glycoprotein. Avian Dis. 2003;47:272–285. doi: 10.1637/0005-2086(2003)047[0272:POCFIB]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 10.Larsen DL, Karasin A, Olsen CW. Immunization of pigs against influenza virus infection by DNA vaccine priming followed by killed-virus vaccine boosting. Vaccine. 2001;19:2842–2853. doi: 10.1016/s0264-410x(01)00014-7. [DOI] [PubMed] [Google Scholar]
- 11.Lee CW, Hilt DA, Jackwood MW. Typing of field isolates of infectious bronchitis virus based on the sequence of the hypervariable region in the S1 gene. J Vet Diagn Invest. 2003;15:344–348. doi: 10.1177/104063870301500407. [DOI] [PubMed] [Google Scholar]
- 12.Lee EK, Jeon WJ, Lee YJ, Jeong OM, Choi JG, Kwon JH, Choi KS. Genetic diversity of avian infectious bronchitis virus isolates in Korea between 2003 and 2006. Avian Dis. 2008;52:332–337. doi: 10.1637/8117-092707-ResNote.1. [DOI] [PubMed] [Google Scholar]
- 13.Liu S, Wang Y, Ma Y, Han Z, Zhang Q, Shao Y, Chen J, Kong X. Identification of a newly isolated avian infectious bronchitis coronavirus variant in China exhibiting affinity for the respiratory tract. Avian Dis. 2008;52:306–314. doi: 10.1637/8110-091307-ResNote.1. [DOI] [PubMed] [Google Scholar]
- 14.Mase M, Tsukamoto K, Imai K, Yamaguchi S. Phylogenetic analysis of avian infectious bronchitis virus strains isolated in Japan. Arch Virol. 2004;149:2069–2078. doi: 10.1007/s00705-004-0369-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.McKinley ET, Hilt DA, Jackwood MW. Avian coronavirus infectious bronchitis attenuated live vaccines undergo selection of subpopulations and mutations following vaccination. Vaccine. 2008;26:1274–1284. doi: 10.1016/j.vaccine.2008.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Pan Z, Zhang X, Geng S, Fang Q, You M, Zhang L, Jiao X, Liu X. Prime-boost immunization using a DNA vaccine delivered by attenuated Salmonella enterica serovar typhimurium and a killed vaccine completely protects chickens from H5N1 highly pathogenic avian influenza virus. Clin Vaccine Immunol. 2010;17:518–523. doi: 10.1128/CVI.00387-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pohuang T, Chansiripornchai N, Tawatsin A, Sasipreeyajan J. Detection and molecular characterization of infectious bronchitis virus isolated from recent outbreaks in broiler flocks in Thailand. J Vet Sci. 2009;10:219–223. doi: 10.4142/jvs.2009.10.3.219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Raj GD, Jones RC. Cross-reactive cellular immune responses in chickens vaccinated with live infectious bronchitis virus vaccine. Avian Pathol. 1997;26:641–649. doi: 10.1080/03079459708419240. [DOI] [PubMed] [Google Scholar]
- 19.Spaan W, Cavanagh D, Horzinek MC. Coronaviruses: structure and genome expression. J Gen Virol. 1988;69(Pt 12):2939–2952. doi: 10.1099/0022-1317-69-12-2939. [DOI] [PubMed] [Google Scholar]
- 20.Tang M, Wang H, Zhou S, Tian G. Enhancement of the immunogenicity of an infectious bronchitis virus DNA vaccine by a bicistronic plasmid encoding nucleocapsid protein and interleukin-2. J Virol Methods. 2008;149:42–48. doi: 10.1016/j.jviromet.2008.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yan F, Yue W, Liu J, Li XY, Zhao YJ. Isolation and biological properties of avian infectious bronchitis virus isolated from Shanxi province. Chin J Vet Sci. 2009;29:845–848. [Google Scholar]
- 22.Yan F, Zhao Y, Wu Q, Yue W, Li X, Ji W, Liu J, Liu F, Ren J, Hua L. Cloning and expression of the S1, N and M gene of IBV. Zhongguo Nong Xue Tong Bao. 2010;26:14–17. [Google Scholar]
- 23.Yan F, Zhao Y, Yue W, Yao J, Lihua L, Ji W, Li X, Liu F, Wu Q. Phylogenetic analysis of S1 gene of infectious bronchitis virus isolates from China. Avian Dis. 2011;55:451–458. doi: 10.1637/9446-070510-ResNote.1. [DOI] [PubMed] [Google Scholar]
- 24.Yang T, Wang HN, Wang X, Tang JN, Gao R, Li J, Guo ZC, Li YL. Multivalent DNA vaccine enhanced protection efficacy against infectious bronchitis virus in chickens. J Vet Med Sci. 2009;71:1585–1590. doi: 10.1292/jvms.001585. [DOI] [PubMed] [Google Scholar]
- 25.Yin Z, Liu J. Animal Virology. 2nd ed. Beijing: Science Press; 1997. pp. 329–331. [Google Scholar]
- 26.Zou NL, Zhao FF, Wang YP, Liu P, Cao SJ, Wen XT, Huang Y. Genetic analysis revealed LX4 genotype strains of avian infectious bronchitis virus became predominant in recent years in Sichuan area, China. Virus Genes. 2010;41:202–209. doi: 10.1007/s11262-010-0500-9. [DOI] [PMC free article] [PubMed] [Google Scholar]