

Abstract
Recent emergence and growing use of terahertz (THz) radiation for medical imaging and public security screening raise questions on reasonable levels of exposure and health consequences of this form of electromagnetic radiation. In particular, picosecond-duration THz pulses have shown promise for novel diagnostic imaging techniques. However, the effects of THz pulses on human cells and tissues thus far remain largely unknown. We report on the investigation of the biological effects of pulsed THz radiation on artificial human skin tissues. We observe that exposure to intense THz pulses for ten minutes leads to a significant induction of H2AX phosphorylation, indicating that THz pulse irradiation may cause DNA damage in exposed skin tissue. At the same time, we find a THz-pulse-induced increase in the levels of several proteins responsible for cell-cycle regulation and tumor suppression, suggesting that DNA damage repair mechanisms are quickly activated. Furthermore, we find that the cellular response to pulsed THz radiation is significantly different from that induced by exposure to UVA (400 nm).
OCIS codes: (170.0170) Medical optics and biotechnology, (170.1420) Biology, (170.6930) Tissue, (170.7160) Ultrafast technology
1. Introduction
Recent technological breakthroughs have led to numerous applications of THz radiation [1–6]. In the medical field, sensitivity of THz radiation to the molecular properties of biological tissue, from cell hydration to conformation states of important biomolecules [7–14], has spurred development of new THz-based approaches to cancer diagnosis. Application of THz pulsed imaging to detection of breast cancer [15], including in situ breast cancer [16], as well as melanoma [4] and basal cell skin carcinoma [17] has been demonstrated. The first in vivo trials of THz imaging as an intra-operative tool during cancer surgery are currently underway [18,19]. In security applications, THz radiation is now extensively applied for identification of concealed explosives and weapons [20,21]. While these new THz radiation-based applications have generated excitement in the research community, concerns have been raised regarding the possible health risks associated with THz exposure [22–28].
The current understanding of biological effects of THz radiation, i.e., its potential to induce DNA damage and impact cell activity, is still limited. Until recently, it was assumed that exposure risks are thermal in nature due to absorption of THz radiation by water in biological tissue [4,6,29,30]. However, several theoretical studies have suggested that while the energy of THz photons is too low to break chemical bonds, resonance-type linear and nonlinear interactions of THz electromagnetic fields with DNA may, under certain conditions, significantly alter DNA dynamics and even induce localized openings (bubbles) in the DNA strands [22,31,32]. This is of particular concern for applications of intense THz pulses of picosecond duration. A few experimental studies have explored the cellular and molecular responses to continuous-wave, monochromatic THz radiation and found evidence of THz-induced genotoxicity in human lymphocytes [23], spindle disturbances in human-hamster hybrid cells [24], as well as changes in gene expression and activation of apoptotic and necrotic processes in human dermal fibroblasts and Jurkat cells [25,26]. A recent study exposing mouse stem cells to broadband THz pulses reported the first experimental confirmation of THz-pulse-induced gene expression changes in mammalian cells, possibly correlated to THz-induced breathing vibrational modes in the corresponding promoter DNA [27,28]. However, the effects of intense broadband (picosecond) THz pulses on human cells and tissues are not known. In particular, since the penetration depth of THz radiation into the human body is limited to a fraction of a millimeter [5], knowledge of the biological effects of intense THz pulses used in novel cancer detection modalities on human skin is important.
In this work, we show evidence strongly indicative of double strand breaks (DSBs) in DNA induced by intense, picosecond THz pulses in exposed artificial human skin tissue models. We use the presence of phosphorylated H2AX (γH2AX), which is one of the earliest and most characterized cellular responses to DSBs [33,34], as a surrogate marker for DNA damage. At the same time, we observe THz-pulse-induced increases in the levels of multiple tumor suppressor and cell-cycle regulatory proteins that facilitate DNA repair. This may suggest that DNA damage in human skin arising from broadband THz pulse exposure could be quickly and efficiently repaired, therefore minimizing the risk of point mutation, prelude to carcinogenesis. We note that the energy of the intense THz pulses used in this study of 0.1 – 1.0 μJ is many orders of magnitude higher than pulse energies of approximately 10−8 μJ used in current medical imaging applications that use traditional photoconductive THz pulse emitters [30]. However, the observed capability of intense THz pulses to affect DNA and cellular functions warrants exploration into potential therapeutic applications of intense THz pulses.
2. Materials and methods
2.1 THz pulse source, tissue models, and exposure conditions
In our experiments, we exposed artificial human 3D skin tissues to broadband THz pulses with ~1.7 ps duration, 1 kHz repetition rate, and pulse energy variable up to 1 μJ. The THz pulses were generated by optical rectification of tilted-pulse-front 800 nm pulses from an amplified Ti:sapphire laser source in LiNbO3 [35,36], as shown in Fig. 1(a) . A pyroelectric detector (Spectrum Detector) was used to measure the THz pulse energies. Free-space electro-optic sampling was used to record the THz pulse waveform, as shown in Fig. 1(b). The corresponding amplitude spectrum of the THz pulse, which is shown in Fig. 1(c), is peaked at 0.5 THz and has a bandwidth of 0.1 - 2 THz. The full thickness human skin tissue model (EpiDermFT, MatTek) reconstructs normal tissue structure and consists of normal, human-derived epidermal keratinocytes and dermal fibroblasts that form a multilayered, highly differentiated model of human dermis and epidermis, as shown in Fig. 1(d). It is mitotically and metabolically active, and preserves the arrangement and communication of cells in skin tissue in vivo [37,38], and thus provides an appropriate platform for assessing the effects of exposure to high intensity THz pulses on human skin. The tissues were cultured according to the manufacturer’s protocol, using a conventional air-liquid interface tissue culture technique. The top surface of the tissue was exposed to air, and the bottom surface of the tissue was fed by a culture medium through pores in a membrane. Tissue samples in single well plate inserts were removed from the medium and placed at the focus of the 1 kHz pulsed THz beam with pulse energies of either 1 μJ (referred to here as “high” THz energy) or 0.1 μJ (“low” THz energy) for 10 minutes. The THz radiation was incident on the top surface of the tissue sample. The THz spot size at the focus was 1.5 mm diameter, as imaged by a pyroelectric infrared camera (Electrophysics PV320) and shown in Fig. 1(e). This produced peak incident THz electric fields at the sample of approximately 220 kV/cm and 70 kV/cm at high and low THz pulse energies, respectively. Since the penetration depth of THz radiation in human skin is less than 150 μm [5], most of the THz pulse energy (> 93%) is absorbed within the 400 μm thick tissue samples.Exposure to UVA, a known genotoxic agent, was performed using 400 nm, 50 fs, 0.080 μJ pulses with 1 kHz repetition rate for 2 minutes and was used as a positive control for induction of DNA damage [39–42]. The UVA beam spot size at the tissue surface was 2.7 mm diameter. Unexposed tissue samples served as controls (CT). Four tissue samples were used for each of the four experimental conditions (CT, Low, High, and UVA). The tissues were then stored in a multiwell dish filled with fresh medium and incubated at 37 °C for 30 minutes, followed by a snap freeze on dry ice. Control tissues underwent the same procedures other than being irradiated. THz exposures were carried out at biologically-low ambient temperature (21 °C). The time-averaged THz power density was 57 mW/cm2 at the highest THz pulse energy. Applying the theoretical formalism developed in [30], we estimate that temperature increase due to THz exposure was less than 0.7 °C.
Fig. 1.

Exposure of artificial human skin tissues equivalents to intense THz pulses. (a) Schematic of the tilted-pulse-front THz pulse source and EpiDermFT tissue in a single-well insert placed at the focus of THz beam. (b) Waveform of a 1.0 μJ THz pulse and (c) corresponding amplitude spectrum. (d) Histology of the EpiDermFT tissue sample (400X image by MatTek Corporation, used with permission). The stained paraffin section reveals epidermis containing basal, spinous, granular keratinocytes and stratum corneum. The dermis contains numerous viable fibroblasts. (e) Image of the THz beam spot size at the sample location. The 1/e2 diameter of the THz beam is 1.5 mm. The dashed square shows the size of the area (2x2 mm2) that was cut out to separate exposed from unexposed tissue.
2.2 Protein extraction and western immunoblotting
The central square region (2 mm x 2 mm, Fig. 1(e)) containing the exposed portion of the frozen tissues was cut out and used for western immunoblotting analysis for determining the levels of various cell-cycle regulatory and tumor suppressant proteins, as described in [43]. Briefly, tissue samples were sonicated in 0.4 ml of ice-chilled 1% sodium dodecyl sulphate (SDS) and boiled for 10 min. Small aliquots (10 µl) of homogenate were reserved for protein determination using assay reagents from BioRad (Hercules, CA, USA). Since the amount of starting tissue was very small, upon quantification protein extracts of three independent biological replicates were pooled together with equal representation of each sample. Pooled protein extracts were separated by SDS–polyacrylamide electrophoresis (PAGE) in slab gels of 12% polyacrylamide, made in duplicates, and transferred to PVDF membranes (Amersham, Baie d’Urfe, QC, Canada). Membranes were incubated with antibodies against γH2AX, p21, p16, p15, p27, EGR1 (1:1000, Cell Signaling, Danvers, MA, USA), p53 (1:500, Santa Cruz Biotechnology, Santa Cruz, CA), p-p53 (Ser15) (1:500, Cell Signaling) KU70 (1:1000, Santa Cruz Biotechnology), actin (1:2000, Abcam, Cambridge, MA). Antibody binding was revealed by incubation with horseradish peroxidase-conjugated secondary antibodies (Amersham) and ECL Plus immunoblotting detection (Amersham). Chemiluminescence was detected by a FluorChem HD2 System (Cell Biosciences/ProteinSimple, Santa Clara, CA, USA) and Biomax MR films (Eastman Kodak, New Haven, CT, USA). Unaltered PVDF membranes were stained with Coomassie Blue (BioRad).
2.3 γ-H2AX immunochemistry
For each distinctive exposure type condition (high, low, UVA, CT), one frozen tissue was sectioned into 5-10-μm-thick slices for immunochemistry analysis, as described in [38]. Briefly, for the γ-H2AX assay, frozen sections were dried, fixed in 2% paraformaldehyde, permeabilized with 1% Triton X-100, and processed for immunostaining with an anti-γH2AX rabbit antibody. The nuclei were counterstained with 4,6-diamidino-2-phenylindole (DAPI).
3. Results and discussion
Earlier theoretical studies have suggested that exposure to high intensity THz radiation with frequencies resonant with the local breathing dynamics of double-stranded DNA may lead to the formation of localized openings in the double strands [22]. Picosecond duration, intense THz pulses have instantaneous THz fields of hundreds of kV/cm, which might be sufficient to not only temporarily disrupt DNA dynamics but also cause DSBs to occur. In DSB, two complementary strands of the DNA double helix are damaged simultaneously. It is potentially the most dangerous type of DNA damage, which, if unrepaired, can lead to cell death or cancer [34]. DSBs can occur in cells due to many different endogenous factors, such as oxidative damage by reactive oxygen species, as well as exogenous factors. Of the known exogenous factors, ionizing radiation, such as X-rays [44] or UV radiation [39–41] are the most prominent causes of DSBs. Regardless of the exact mechanism of DSB formation, they can be experimentally detected by assaying for the presence of the phosphorylated histone H2AX (γH2AX). Hundreds of H2AX molecules in the chromatin surrounding the break site become phosphorylated at the Serine 139 position within minutes of DSB formation [34,45]. The resulting γH2AX foci attract repair proteins to the DSB site, thus playing a critical role in DSB recognition and initiation of repair processes [34]. Analysis of γH2AX expression is widely used as a surrogate marker for DNA damage and a reliable DSB detection tool, since it is orders of magnitude more sensitive compared to other DSB visualization techniques [34,46].
To assess the occurrence of DSB in THz-exposed and UV-exposed tissues compared to unexposed controls, we assayed for the presence of γH2AX using western blot analysis. We observe that exposure to intense THz pulses for just ten minutes leads to a significant induction of H2AX phosphorylation compared to control samples, as seen in Western blot images for both high and low THz pulse energy exposures in Fig. 2 . This is an indication that intense THz pulses cause DNA DSBs to occur in human skin cells. UVA (400 nm), which creates reactive oxygen species and causes oxidative DNA damage [40,41,47], also leads to significant increases in γH2AX levels in exposed tissues (Fig. 2).
Fig. 2.

Induction of γH2AX in artificial human skin tissues equivalents following 10 minute exposure to either high (1.0 μJ) or low (0.1 μJ) energy THz pulses, or 2 minute exposure to UVA (400 nm) pulses (0.080 μJ), as compared to control (CT) samples. The pulse repetition rate was 1 kHz. Actin was used as a loading control. Each experiment included pooled lysates from three tissues for each exposure condition with equal representation for each tissue.
The presence and localization of DSB sites in exposed tissue samples can be also studied by visualizing γ-H2AX-expressing cells by in situ immunofluorescence with γH2AX antibodies. Figure 3 shows representative images of CT, high, low, and UVA sample cross-sections. In these images, γH2AX foci appear as bright green, and DNA as bright blue spots. Co-localization of green (γH2AX) and blue (DNA) staining indicates sites within the cells where DNA DSBs likely occurred. In UVA-exposed tissue, we observed DNA damage in the uppermost epidermis, as expected [39–41]. In tissue exposed to high energy THz pulses, multiple γH2AX expressing cells appear in the top layer of epidermis. In low THz pulse energy-exposed tissue, γH2AX expressing cells are seen in the top epidermal layer, as well as in the dermis. As only one tissue sample per each exposure condition was dedicated to immunochemical analysis, we are unable to carry out reliable statistical analysis of the incidence of γH2AX-expressng cells within the tissues. While more systematic immunochemical analysis of THz-exposed skin tissues is necessary to get a quantitative measure of γH2AX expressing cells within the tissues, their depth distribution and exposure parameter dependence, this data set serves as an independent verification of THz-pulse induced DNA damage in artificial human skin tissues.
Fig. 3.
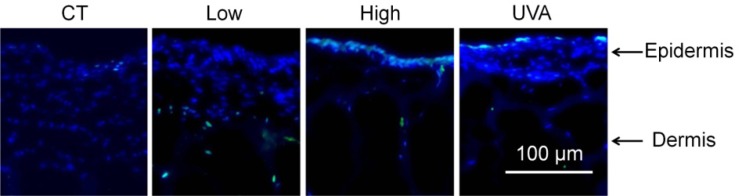
Confocal fluorescence images of tissue sample cross-sections showing the incidence of γH2AX foci (localized green spots). Blue spots indicate cell nuclei. γH2AX-harboring cells (markers for double strand breaks in DNA) inside cell nuclei appear as blue-green spots. Panels from left to right show representative images for control (CT), low THz pulse energy (0.1 μJ), high THz pulse energy (1.0 μJ), and UVA-exposed tissues. Scale bar is 100 μm.
Having seen the THz-induced increase of γH2AX levels, which are indicative of DNA damage, we next proceeded to analyze possible impacts on cellular function by assaying the levels of proteins involved in cell-cycle control and DNA repair. Numerous mechanisms exist by which cells can repair DSBs. Once DNA damage occurs, checkpoints that pause the cell cycle and allow time for DNA repair are activated [46,48]. Two pathways by which DSBs can be repaired are homologous recombination and nonhomologous end joining, as reviewed in [44,49,50]. The latter mechanism is faster and dominates DSB repair, but is susceptible to error as it links broken DNA ends together without reference to accurate base pairing [44]. In order to infer how efficient repair of DNA damage caused by intense THz pulse exposure may be, we analyzed how THz exposure affects the expression level of various proteins that take part in cell cycle control and DSB repair processes by western immunoblotting (Fig. 4 ).
Fig. 4.

Expression of tumor suppressor and cell cycle regulatory proteins in artificial human 3D skin tissues equivalents following 10 minute exposure to either high (1.0 μJ) or low (0.1 μJ) energy THz pulses, or two minute exposure to 0.080 μJ UVA (400 nm) pulses, as compared to unexposed control (CT) samples. Actin was used as a loading control.
One of our most striking findings is an increase in the levels of protein p53, which is significantly more pronounced in THz-exposed tissues compared to UVA-exposed tissues (Fig. 4). An important tumor suppressor and cell-cycle regulator, p53 has been termed ‘the guardian of the genome’ [51,52]. It regulates the cell growth arrest and apoptosis, and also takes part in DSB repair [51,52]. Previous studies have reported that repetitive exposures to continuous-wave UVA induced a strong increase in the levels of p53-positive cells in the human epidermis after a total UVA dose of over 12.5 J/cm2 [53], 100 fold larger than our two minute exposure to pulsed UVA light, which we observed was insufficient to cause significant upregulation of p53. On the other hand, the level of the phosphorylated variant of the p53 protein, p-p53, is barely affected in THz-exposed tissues compared to controls, as shown in Fig. 4, and significantly increases with UV exposure. Here, we measured the levels of p53 phosphorylation of Serine 15 (p-p53 Ser15). This modification is usually catalysed by damage–activated ATM kinase and further transactivates the kinase inhibitor p21, leading to cell-cycle arrest at the G1/S checkpoint. Moreover, p53 may influence the G2/M checkpoint via transactivation of p21. In our experiment, the tissues were frozen 30 minutes after THz exposure, which might not be sufficient time for p53 phosphorylation at Ser15 by ATM. Alternatively, THz-induced DNA damage may not activate ATM, or may cause its delayed activation. Another potent in vivo tumor suppressor and p53 target, early growth response 1 (EGR1) [54], is also slightly upregulated by THz exposure, albeit weaker than by UVA. Recent studies have shown that p53 and EGR1 collaborate in controlling cellular growth and cell cycle, thus allowing time for DNA repair [55]. In mammals, most DSBs are repaired by nonhomologous end joining [44,50,56]. Protein KU70 is essential for this process and is induced in mammalian cells exposed to X-rays [57], gamma-rays [58], as well as UV radiation [42]. We note a significant upregulation of KU70 in THz and UVA-exposed tissues, as shown in Fig. 4, indicative of activation of nonhomologous end joining.
We have also measured the levels of four key cell cycle regulator proteins, p21, p16, p15 and p27, in THz and UVA exposed tissues. These all belong to the cyclin-dependent kinase inhibitor protein family, and act as tumor suppressors [59]. Their upregulation provides cells crucial time for repairing DNA. We note an increase in the levels of p21, p16, and p27 in THz-exposed tissues but not in UVA-exposed tissues. The levels of another cell cycle regulatory protein, p15, were unaltered by either THz pulses or UVA. The observed concerted upregulation of p53 (discussed earlier), p16 and p21 is a hallmark of DNA damage response in cells [60]. Thus, the observed simultaneous increase in expression of multiple known tumor suppressor and damage repair proteins (p53, p21, p16, p27 and KU70) by intense THz pulse exposure suggests that DNA damage was induced, and that damage repair mechanisms were promptly activated. However, while further study is required to elucidate the specific roles of the aforementioned proteins in cellular response to THz pulse exposure, the observed ability of intense THz pulses to induce DNA damage and affect cellular repair functions naturally leads to the question: Can THz pulses be used to treat of diseases such as cancer? Given that the penetration depth of THz waves into the human body is sub-mm, exploring the possibility of the application of THz pulses to treat skin cancers holds the most promise for having significant impact on patient care.
Furthermore, it is known that UVA radiation damages DNA via photosensitized reactions that form reactive oxygen species [40–42]. While this damage is repaired by base excision, it often yields abasic sites and single-strand breaks, which may later be converted to DSBs during DNA replication [42]. THz photons have energies three orders of magnitude smaller than UVA photons, which is insufficient to directly break covalent bonds and form free radicals. However, recent studies have shown that DNA, proteins, as well as other important biomolecules have collective structural vibrational modes at THz frequencies [7,8,10–14]. Resonant coupling of intense THz pulses to those intermolecular vibrational modes may therefore induce new conformation states of various cellular biomolecules, or destabilize the double-stranded DNA structure via nonlinear resonant effects as suggested in [22], thereby affecting cellular functions. More theoretical and experimental studies are clearly needed to elucidate the mechanisms by which THz pulses interact with living cells and tissues.
4. Summary
In summary, we have observed that exposure to intense THz pulses induces phosphorylation of H2AX, indicative of the formation of DNA double strand breaks, and at the same time profoundly activates DNA damage response in artificial human skin tissues. Simultaneous upregulation of multiple important tumor suppressor proteins in the exposed skin strongly suggests that THz-pulse-induced DNA damage may be effectively repaired. Future studies are needed to understand the precise molecular mechanisms for the induction of DNA damage and changes in cellular signaling by intense picosecond THz pulses, as well as the cellular repercussions of such changes. This study may therefore serve as an important roadmap for future analysis of the mechanisms and cellular effects of pulsed THz exposure in human tissue.
Acknowledgments
We wish to acknowledge financial support from the Alberta Cancer Foundation, Alberta Innovates – Health Solutions (AIHS), Natural Sciences and Engineering Research Council of Canada (NSERC), Canada Foundation for Innovation (CFI), and the Alberta Science and Research Investments Program (ASRIP). We are also grateful for technical assistance from D. Mullin, G. Popowich and A. Kovalchuk. We thank M. Reid for use of the pyroelectric infrared camera (Electrophysics PV320) for THz spot size imaging.
References and links
- 1.D. M. Mittleman, Sensing with Terahertz Radiation (Springer, 2010). [Google Scholar]
- 2.Tonouchi M., “Cutting-edge terahertz technology,” Nat. Photonics 1(2), 97–105 (2007). 10.1038/nphoton.2007.3 [DOI] [Google Scholar]
- 3.Jepsen P. U., Cooke D. G., Koch M., “Terahertz spectroscopy and imaging — Modern techniques and applications,” Laser Photonics Rev. 5(1), 124–166 (2011). 10.1002/lpor.201000011 [DOI] [Google Scholar]
- 4.Fitzgerald A. J., Berry E., Zinovev N. N., Walker G. C., Smith M. A., Chamberlain J. M., “An introduction to medical imaging with coherent terahertz frequency radiation,” Phys. Med. Biol. 47(7), R67–R84 (2002). 10.1088/0031-9155/47/7/201 [DOI] [PubMed] [Google Scholar]
- 5.Siegel P. H., “Terahertz Technology in Biology and medicine,” IEEE Trans. Microw. Theory Tech. 52(10), 2438–2447 (2004). 10.1109/TMTT.2004.835916 [DOI] [Google Scholar]
- 6.Wilmink G. J., Grundt J. E., “Current State of Research on Biological Effects of Terahertz Radiation,” J. Infrared Millim. Terahz Waves 32(10), 1074–1122 (2011). 10.1007/s10762-011-9794-5 [DOI] [Google Scholar]
- 7.Markelz A. G., Whitmire S., Hillebrecht J., Birge R., “THz time domain spectroscopy of biomolecular conformational modes,” Phys. Med. Biol. 47(21), 3797–3805 (2002). 10.1088/0031-9155/47/21/318 [DOI] [PubMed] [Google Scholar]
- 8.Fischer B. M., Walther M., Jepsen P. U., “Far-infrared vibrational modes of DNA components studied by terahertz time-domain spectroscopy,” Phys. Med. Biol. 47(21), 3807–3814 (2002). 10.1088/0031-9155/47/21/319 [DOI] [PubMed] [Google Scholar]
- 9.Brucherseifer M., Nagel M., Haring Bolivar P., Kurz H., Bosserhoff A., Büttner R., “Label-free probing of the binding state of DNA by time-domain terahertz sensing,” Appl. Phys. Lett. 77(24), 4049–4051 (2000). 10.1063/1.1332415 [DOI] [PubMed] [Google Scholar]
- 10.Smye S. W., Chamberlain J. M., Fitzgerald A. J., Berry E., “The interaction between Terahertz radiation and biological tissue,” Phys. Med. Biol. 46(9), R101–R112 (2001). 10.1088/0031-9155/46/9/201 [DOI] [PubMed] [Google Scholar]
- 11.Nagel M., Haring Bolivar P., Brucherseifer M., Kurz H., Bosserhoff A., Büttner R., “Integrated THz technology for label-free genetic diagnostics,” Appl. Phys. Lett. 80(1), 154–156 (2002). 10.1063/1.1428619 [DOI] [Google Scholar]
- 12.Vasudev M., Yang J., Jung H.-S., Stroscio M. A., Dutta M., “Integrated nanostructure–semiconductor molecular complexes as tools for THz spectral studies of DNA,” IEEE Sens. J. 10(3), 524–530 (2010). 10.1109/JSEN.2009.2037804 [DOI] [Google Scholar]
- 13.Kim S. J., Born B., Havenith M., Gruebele M., “Real-time detection of protein-water dynamics upon protein folding by terahertz absorption spectroscopy,” Angew. Chem. Int. Ed. Engl. 47(34), 6486–6489 (2008). 10.1002/anie.200802281 [DOI] [PubMed] [Google Scholar]
- 14.Markelz A. G., “Terahertz Dielectric Sensitivity to Biomolecular Structure and Function,” IEEE J. Sel. Top. Quantum Electron. 14(1), 180–190 (2008). 10.1109/JSTQE.2007.913424 [DOI] [Google Scholar]
- 15.Ashworth P. C., Pickwell-MacPherson E., Provenzano E., Pinder S. E., Purushotham A. D., Pepper M., Wallace V. P., “Terahertz pulsed spectroscopy of freshly excised human breast cancer,” Opt. Express 17(15), 12444–12454 (2009). 10.1364/OE.17.012444 [DOI] [PubMed] [Google Scholar]
- 16.Fitzgerald A. J., Wallace V. P., Jimenez-Linan M., Bobrow L., Pye R. J., Purushotham A. D., Arnone D. D., “Terahertz pulsed imaging of human breast tumors,” Radiology 239(2), 533–540 (2006). 10.1148/radiol.2392041315 [DOI] [PubMed] [Google Scholar]
- 17.Woodward R. M., Wallace V. P., Arnone D. D., Linfield E. H., Pepper M., “Terahertz pulsed imaging of skin cancer in the time and frequency domain,” J. Biol. Phys. 29(2/3), 257–259 (2003). 10.1023/A:1024409329416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.P. C. Ashworth, P. O’Kelly, A. D. Purushotham, S. E. Pinder, M. Kontos, M. Pepper, and V. P. Wallace, “An intra-operative THz probe for use during the surgical removal of breast tumors,” in 33rd International Conference on Infrared, Millimeter and Terahertz Waves, 2008. IRMMW-THz 2008 (2008), pp. 1–3. doi: 10.1109/ICIMW.2008.4665810. 10.1109/ICIMW.2008.4665810 [DOI] [Google Scholar]
- 19.“TeraView receives MHRA approval to conduct in-vivo cancer imaging trials,” http://www.teraview.com/news/terahertz-news/TeraView-receives-in-vivo-trials-approval.html
- 20.Federici J. F., Schulkin B., Huang F., Gary D., Barat R., Oliveira F., Zimdars D., “THz imaging and sensing for security applications: explosives, weapons and drugs,” Semicond. Sci. Technol. 20(7), S266–S280 (2005). 10.1088/0268-1242/20/7/018 [DOI] [Google Scholar]
- 21.Zimdars D., White J. S., “Terahertz reflection imaging for package and personnel inspection,” Proc. SPIE 5411, 78–83 (2004). 10.1117/12.562216 [DOI] [Google Scholar]
- 22.Alexandrov B. S., Gelev V., Bishop A. R., Usheva A., Rasmussen K. O., “DNA breathing dynamics in the presence of a terahertz field,” Phys. Lett. A 374(10), 1214–1217 (2010). 10.1016/j.physleta.2009.12.077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Korenstein-Ilan A., Barbul A., Hasin P., Eliran A., Gover A., Korenstein R., “Terahertz radiation increases genomic instability in human lymphocytes,” Radiat. Res. 170(2), 224–234 (2008). 10.1667/RR0944.1 [DOI] [PubMed] [Google Scholar]
- 24.Hintzsche H., Jastrow C., Kleine-Ostmann T., Stopper H., Schmid E., Schrader T., “Terahertz radiation induces spindle disturbances in human-hamster hybrid cells,” Radiat. Res. 175(5), 569–574 (2011). 10.1667/RR2406.1 [DOI] [PubMed] [Google Scholar]
- 25.Wilmink G. J., Ibey B. L., Roth C. L., Vincelette R. L., Rivest B. D., Horn C. B., Bernhard J., Roberson D., Roach W. P., “Determination of death thresholds and identification of terahertz (THz) – specific gene expression signatures,” Proc. SPIE 7562, 75620K–, 75620K-8. (2010). 10.1117/12.844917 [DOI] [Google Scholar]
- 26.Wilmink G. J., Rivest B. D., Ibey B. L., Roth C. L., Bernhard J., Roach W. P., “Quantitative investigation of bioeffects associated with terahertz radiation,” Proc. SPIE 7562, 75620L–, 75620L-10. (2010). 10.1117/12.844916 [DOI] [Google Scholar]
- 27.Bock J., Fukuyo Y., Kang S., Phipps M. L., Alexandrov L. B., Rasmussen K. Ø., Bishop A. R., Rosen E. D., Martinez J. S., Chen H.-T., Rodriguez G., Alexandrov B. S., Usheva A., “Mammalian stem cells reprogramming in response to terahertz radiation,” PLoS ONE 5(12), e15806 (2010). 10.1371/journal.pone.0015806 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Alexandrov B. S., Rasmussen K. Ø., Bishop A. R., Usheva A., Alexandrov L. B., Chong S., Dagon Y., Booshehri L. G., Mielke C. H., Phipps M. L., Martinez J. S., Chen H.-T., Rodriguez G., “Non-thermal effects of terahertz radiation on gene expression in mouse stem cells,” Biomed. Opt. Express 2(9), 2679–2689 (2011). 10.1364/BOE.2.002679 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Berry E., Walker G. C., Fitzgerald A. J., Zinov’ev N. N., Chamberlain M., Smye S. W., Miles R. E., Smith M. A., “Do in vivo terahertz imaging systems comply with safety guidelines?” J. Laser Appl. 15(3), 192–198 (2003). 10.2351/1.1585079 [DOI] [Google Scholar]
- 30.Kristensen T. T. L., Withayachumnankul W., Jepsen P. U., Abbott D., “Modeling terahertz heating effects on water,” Opt. Express 18(5), 4727–4739 (2010). 10.1364/OE.18.004727 [DOI] [PubMed] [Google Scholar]
- 31.Chitanvis S. M., “Can low-power electromagnetic radiation disrupt hydrogen bonds in dsDNA?” J. Polym. Sci., B, Polym. Phys. 44(18), 2740–2747 (2006). 10.1002/polb.20910 [DOI] [Google Scholar]
- 32.Zhuang W., Feng Y., Prohofsky E. W., “Self-consistent calculation of localized DNA vibrational properties at a double-helix-single-strand junction with anharmonic potential,” Phys. Rev. A 41(12), 7033–7042 (1990). 10.1103/PhysRevA.41.7033 [DOI] [PubMed] [Google Scholar]
- 33.Rogakou E. P., Pilch D. R., Orr A. H., Ivanova V. S., Bonner W. M., “DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139,” J. Biol. Chem. 273(10), 5858–5868 (1998). 10.1074/jbc.273.10.5858 [DOI] [PubMed] [Google Scholar]
- 34.Sedelnikova O. A., Pilch D. R., Redon C., Bonner W. M., “Histone H2AX in DNA damage and repair,” Cancer Biol. Ther. 2(3), 233–235 (2003). 10.4161/cbt.2.3.373 [DOI] [PubMed] [Google Scholar]
- 35.Blanchard F., Sharma G., Razzari L., Ropagnol X., Bandulet H.-C., Vidal F., Morandotti R., Kieffer J.-C., Ozaki T., Tiedje H., Haugen H., Reid M., Hegmann F. A., “Generation of intense terahertz radiation via optical methods,” IEEE J. Sel. Top. Quantum Electron. 17(1), 5–16 (2011). 10.1109/JSTQE.2010.2047715 [DOI] [Google Scholar]
- 36.Hoffmann M. C., Fülöp J. A., “Intense ultrashort terahertz pulses: generation and applications,” J. Phys. D Appl. Phys. 44(8), 083001 (2011). 10.1088/0022-3727/44/8/083001 [DOI] [Google Scholar]
- 37.Boelsma E., Gibbs S., Faller C., Ponec M., “Characterization and comparison of reconstructed skin models: morphological and immunohistochemical evaluation,” Acta Derm. Venereol. 80(2), 82–88 (2000). [PubMed] [Google Scholar]
- 38.Sedelnikova O. A., Nakamura A., Kovalchuk O., Koturbash I., Mitchell S. A., Marino S. A., Brenner D. J., Bonner W. M., “DNA double-strand breaks form in bystander cells after microbeam irradiation of three-dimensional human tissue models,” Cancer Res. 67(9), 4295–4302 (2007). 10.1158/0008-5472.CAN-06-4442 [DOI] [PubMed] [Google Scholar]
- 39.Agar N. S., Halliday G. M., Barnetson R. S., Ananthaswamy H. N., Wheeler M., Jones A. M., “The basal layer in human squamous tumors harbors more UVA than UVB fingerprint mutations: a role for UVA in human skin carcinogenesis,” Proc. Natl. Acad. Sci. U.S.A. 101(14), 4954–4959 (2004). 10.1073/pnas.0401141101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wang S. Q., Setlow R., Berwick M., Polsky D., Marghoob A. A., Kopf A. W., Bart R. S., “Ultraviolet A and melanoma: a review,” J. Am. Acad. Dermatol. 44(5), 837–846 (2001). 10.1067/mjd.2001.114594 [DOI] [PubMed] [Google Scholar]
- 41.Cadet J., Sage E., Douki T., “Ultraviolet radiation-mediated damage to cellular DNA,” Mutat. Res. 571(1-2), 3–17 (2005). 10.1016/j.mrfmmm.2004.09.012 [DOI] [PubMed] [Google Scholar]
- 42.Rastogi R. P., Richa A., Kumar A., Tyagi M. B., Sinha R. P., “Molecular mechanisms of ultraviolet radiation-induced DNA damage and repair,” J. Nucleic Acids 2010, 592980 (2010). 10.4061/2010/592980 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kovalchuk O., Zemp F. J., Filkowski J. N., Altamirano A. M., Dickey J. S., Jenkins-Baker G., Marino S. A., Brenner D. J., Bonner W. M., Sedelnikova O. A., “microRNAome changes in bystander three-dimensional human tissue models suggest priming of apoptotic pathways,” Carcinogenesis 31(10), 1882–1888 (2010). 10.1093/carcin/bgq119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hoeijmakers J. H. J., “Genome maintenance mechanisms for preventing cancer,” Nature 411(6835), 366–374 (2001). 10.1038/35077232 [DOI] [PubMed] [Google Scholar]
- 45.Dickey J. S., Zemp F. J., Altamirano A., Sedelnikova O. A., Bonner W. M., Kovalchuk O., “H2AX phosphorylation in response to DNA double-strand break formation during bystander signalling: effect of microRNA knockdown,” Radiat. Prot. Dosimetry 143(2-4), 264–269 (2011). 10.1093/rpd/ncq470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Bonner W. M., Redon C. E., Dickey J. S., Nakamura A. J., Sedelnikova O. A., Solier S., Pommier Y., “γH2AX and cancer,” Nat. Rev. Cancer 8(12), 957–967 (2008). 10.1038/nrc2523 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wang H. T., Choi B., Tang M. S., “Melanocytes are deficient in repair of oxidative DNA damage and UV-induced photoproducts,” Proc. Natl. Acad. Sci. U.S.A. 107(27), 12180–12185 (2010). 10.1073/pnas.1005244107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Fillingham J., Keogh M.-C., Krogan N. J., “γH2AX and its role in DNA double-strand break repair,” Biochem. Cell Biol. 84(4), 568–577 (2006). 10.1139/o06-072 [DOI] [PubMed] [Google Scholar]
- 49.McGlynn P., Lloyd R. G., “Recombinational repair and restart of damaged replication forks,” Nat. Rev. Mol. Cell Biol. 3(11), 859–870 (2002). 10.1038/nrm951 [DOI] [PubMed] [Google Scholar]
- 50.Khanna K. K., Jackson S. P., “DNA double-strand breaks: signaling, repair and the cancer connection,” Nat. Genet. 27(3), 247–254 (2001). 10.1038/85798 [DOI] [PubMed] [Google Scholar]
- 51.Menendez D., Inga A., Resnick M. A., “The expanding universe of p53 targets,” Nat. Rev. Cancer 9(10), 724–737 (2009). 10.1038/nrc2730 [DOI] [PubMed] [Google Scholar]
- 52.Bolderson E., Richard D. J., Zhou B. B., Khanna K. K., “Recent advances in cancer therapy targeting proteins involved in DNA double-strand break repair,” Clin. Cancer Res. 15(20), 6314–6320 (2009). 10.1158/1078-0432.CCR-09-0096 [DOI] [PubMed] [Google Scholar]
- 53.Seité S., Moyal D., Verdier M. P., Hourseau C., Fourtanier A., “Accumulated p53 protein and UVA protection level of sunscreens,” Photodermatol. Photoimmunol. Photomed. 16(1), 3–9 (2000). 10.1034/j.1600-0781.2000.160103.x [DOI] [PubMed] [Google Scholar]
- 54.Krones-Herzig A., Mittal S., Yule K., Liang H., English C., Urcis R., Soni T., Adamson E. D., Mercola D., “Early growth response 1 acts as a tumor suppressor in vivo and in vitro via regulation of p53,” Cancer Res. 65(12), 5133–5143 (2005). 10.1158/0008-5472.CAN-04-3742 [DOI] [PubMed] [Google Scholar]
- 55.Zwang Y., Sas-Chen A., Drier Y., Shay T., Avraham R., Lauriola M., Shema E., Lidor-Nili E., Jacob-Hirsch J., Amariglio N., Lu Y., Mills G. B., Rechavi G., Oren M., Domany E., Yarden Y., “Two phases of mitogenic signaling unveil roles for p53 and EGR1 in elimination of inconsistent growth signals,” Mol. Cell 42(4), 524–535 (2011). 10.1016/j.molcel.2011.04.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mahaney B. L., Meek K., Lees-Miller S. P., “Repair of ionizing radiation-induced DNA double-strand breaks by non-homologous end-joining,” Biochem. J. 417(3), 639–650 (2009). 10.1042/BJ20080413 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Reynolds P., Anderson J. A., Harper J. V., Hill M. A., Botchway S. W., Parker A. W., O’Neill P., “The dynamics of Ku70/80 and DNA-PKcs at DSBs induced by ionizing radiation is dependent on the complexity of damage,” Nucleic Acids Res. 40(21), 10821–10831 (2012). 10.1093/nar/gks879 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Chen H., Bao Y., Yu L., Jia R., Cheng W., Shao C., “Comparison of cellular damage response to low-dose-rate 125I seed irradiation and high-dose-rate gamma irradiation in human lung cancer cells,” Brachytherapy 11(2), 149–156 (2012). 10.1016/j.brachy.2011.05.002 [DOI] [PubMed] [Google Scholar]
- 59.Sherr C. J., Roberts J. M., “Inhibitors of mammalian G1 cyclin-dependent kinases,” Genes Dev. 9(10), 1149–1163 (1995). 10.1101/gad.9.10.1149 [DOI] [PubMed] [Google Scholar]
- 60.Sperka T., Wang J., Rudolph K. L., “DNA damage checkpoints in stem cells, ageing and cancer,” Nat. Rev. Mol. Cell Biol. 13(9), 579–590 (2012). 10.1038/nrm3420 [DOI] [PubMed] [Google Scholar]