

Abstract
Leishmania spp. are protozoa responsible for leishmaniasis, a neglected disease that kills up to 50,000 people every year. Current therapies mainly rely on antimonial drugs that are inadequate because of their poor efficacy and safety and increased drug resistance. An urgent need exists to find new and more affordable drugs. Our previous study demonstrated the antileishmanial activity of eupomatenoid-5, a neolignan obtained from leaves of Piper regnellii var. pallescens. The aim of the present study was to clarify the mode of action of eupomatenoid-5 against L. amazonensis. We used biochemical and morphological techniques and demonstrated that eupomatenoid-5 induced cell death in L. amazonensis promastigotes, sharing some phenotypic features observed in metazoan apoptosis, including increased reactive oxygen species production, hypopolarization of mitochondrial potential, phosphatidylserine exposure, decreased cell volume, and G0/G1 phase cell cycle arrest.
1. Introduction
Leishmaniasis remains a significant neglected tropical disease that puts 350 million people in 88 countries on four continents at risk for infection, with approximately 50,000 deaths per year [1]. Approximately 21 species have been found to cause three different clinical manifestations of leishmaniasis: cutaneous, in which the lesions are confined to the site of inoculation by the sandfly, and mucocutaneous, which affects mucosal tissues, and visceral leishmaniasis affecting the inner organs [2].
Current chemotherapy for leishmaniasis is still based on the use of pentavalent antimonials as first-line drugs and pentamidine, amphotericin B (free or liposomal forms), paromomycin, and miltefosine as second-line drugs. Although these drugs are usually effective, they have limitations, such as toxicity in the host and long-term treatment [3, 4]. These drawbacks reveal the urgent need to develop new therapeutic agents for the treatment of leishmaniasis.
Natural compounds, known to be valuable sources of new medicinal agents, have been exhaustively evaluated against Leishmania [5–7]. One example is eupomatenoid-5, a neolignan obtained from the leaves of Piper regnellii var. pallescens, representing a subclass with various biological activities, such as antifungal, antibacterial, insecticidal, and trypanocidal effects [8–12]. Our previous study reported the antileishmanial effect of eupomatenoid-5 [13]. Considering such antileishmanial activity, the aim of the present study was to better characterize the biochemical alterations induced by this compound against promastigote forms of L. amazonensis and elucidate the mechanism of action of eupomatenoid-5 involved in the cell death of this protozoan parasite.
2. Materials and Methods
2.1. Chemicals
Actinomycin D, antimycin A, bovine serum albumin, carbonyl cyanide m-chlorophenylhydrazone (CCCP), digitonin, dimethylsulfoxide (DMSO), rhodamine 123 (Rh123), 2′,7′-dichlorofluorescin diacetate (H2DCFDA), and 5-5′-dithiobis(2-nitrobenzoic acid) (DTNB) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Fetal bovine serum (FBS) was obtained from Invitrogen (Grand Island, NY, USA). Annexin-V FITC, 3,8-phenanthridine diamine-5-(6-triphenylphosphonium hexyl)-5,6-dihydro-6-phenyl (MitoSOX) and propidium iodide (PI) were obtained from Invitrogen (Eugene, OR, USA). The protein assay kit was obtained from Bio-Rad (Hercules, CA, USA). All of the other reagents were of analytical grade.
2.2. Isolation of Eupomatenoid-5 from Leaves of Piper regnellii Var. pallescens
Eupomatenoid-5 (Figure 1) was isolated from the leaves of P. regnellii, which was collected in the Professor Irenice Silva Garden of Medicinal Plants on the campus of the State University of Maringa (UEM) in Parana, Brazil. A voucher specimen (no. HUM 8392) was deposited at the UEM Herbarium. Briefly, the dry plant material was extracted by exhaustive maceration at room temperature in the dark in an ethanol : water ratio of 90 : 10. Fractionation was performed from the ethyl acetate extract to obtain the hexane fraction, and a dihydrobenzofuran neolignan, eupomatenoid-5, was isolated from this fraction as described previously [9]. The compound was purified using absorption-chromatographic methods and identified by analyzing the ultraviolet, infrared, 1H nuclear magnetic resonance (NMR), 13C NMR, distortionless enhancement polarization transfer (DEPT), correlated spectroscopy (COSY), heteronuclear correlation (HETCOR), nuclear overhauser effect spectroscopy (NOESY), heteronuclear multiple bond correlation (HMBC), and gas chromatography/mass spectrometry (GC/MS) spectra. The data were compared with the literature [14].
Figure 1.
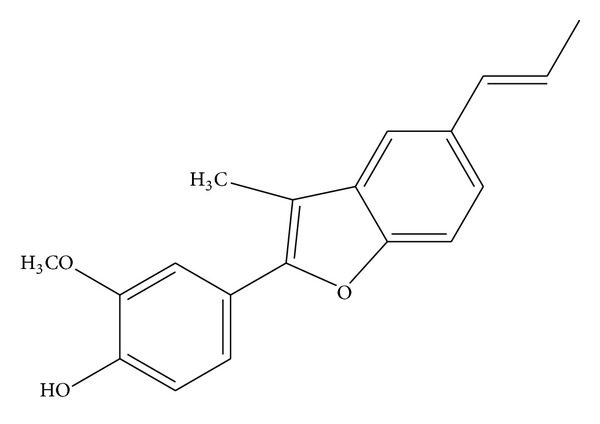
Structure of eupomatenoid-5 isolated from the leaves of Piper regnellii var. pallescens.
Stock solutions of eupomatenoid-5 were prepared aseptically in DMSO and diluted in culture medium so that the DMSO concentration did not exceed 1% in the experiments. The concentrations of eupomatenoid-5 used in the assays were 30.0, 85.0, and 170.0 μM, representing the IC50, IC90, and twofold IC90, respectively [13].
2.3. Parasites
Leishmania amazonensis promastigotes (MHOM/BR/Josefa) were maintained at 25°C in Warren's medium (brain-heart infusion plus hemin and folic acid; pH 7.2) supplemented with 10% heat-inactivated FBS.
2.4. Determination of Mitochondrial Transmembrane Potential (ΔΨm)
Promastigotes (1 × 107 cells/mL) were treated or untreated with 30.0, 85.0, and 170.0 μM eupomatenoid-5 for 24 h at 37°C, harvested, and washed with phosphate-buffered saline (PBS). The parasites were incubated with 1 mL (5 mg/mL in ethanol) of Rh123, a fluorescent probe that accumulates in mitochondria, for 15 min, resuspended in 0.5 mL PBS, and incubated for an additional 30 min. The assay was conducted according to the manufacturer's instructions. The parasites were analyzed using a BD FACSCalibur flow cytometer and CellQuest Pro software (Becton Dickinson and Company, USA, 1997). A total of 10,000 events were acquired in the region that corresponded to the parasites. CCCP at 100 μM was used as a positive control [15].
2.5. Measurement of Reactive Oxygen Species
Promastigotes (1 × 107 cells/mL) were treated or untreated with 30.0, 85.0, and 170.0 μM for 3 and 24 h, centrifuged, washed, and resuspended in PBS (pH 7.4). Afterward, these parasites were loaded with 10 μM of a permeant probe, H2DCFDA, in the dark for 45 min [16]. Reactive oxygen species (ROS) were measured as an increase in fluorescence caused by the conversion of nonfluorescent dye to highly florescent 20,70-dichlorofluorescein, with an excitation wavelength of 488 nm and emission wavelength of 530 nm in a fluorescence microplate reader (Victor X3, PerkinElmer, Finland).
2.6. Fluorimetric Detection of Mitochondrial-Derived O2 •−
Promastigotes (2 × 107 cells/mL) were loaded with 5 μM of a fluorescent O2 •−-sensitive, mitochondrial-targeted probe, MitoSOX, for 10 min at 25°C and then washed with Krebs-Henseleit (KH) buffer (pH 7.3) that contained 15 mM NaHCO3, 5 mM KCl, 120 mM NaCl, 0.7 mM Na2HPO4, and 1.5 mM NaH2PO4 [17]. After the parasites were treated or untreated with 30.0, 85.0, and 170.0 μM eupomatenoid-5, the fluorescence intensity was detected after 1, 2, 3, and 4 h of treatment using a fluorescence microplate reader (Victor X3, PerkinElmer, Finland), with an excitation wavelength of 510 nm and emission wavelength of 580 nm. Oxidized MitoSOX (oxMitoSOX) becomes highly fluorescent upon binding to nucleic acids. Cells were exposed to 10 μM antimycin A, a stimulus known to induce mitochondrial O2 •− production.
2.7. Estimation of Decrease in Reduced Thiol Level
Thiol levels were determined using DTNB. Promastigotes (1 × 107 cells/mL) were treated or untreated with 30.0, 85.0, and 170.0 μM eupomatenoid-5 for 24, 48, and 72 h at 25°C. Afterward, the parasites were centrifuged, dissolved in 10 mM Tris-HCl buffer (pH 2.5), and sonicated. Acidic pH was used during sonication to prevent oxidation of the free thiol groups. Cellular debris was removed by centrifugation, and 100 μL of the supernatant and 100 μL of 500 mM phosphate buffer (pH 7.5) were taken in each microtiter well, followed by the addition of 20 μL of 1 mM DTNB to each well. Absorbance was measured at 412 nm [16].
2.8. Phosphatidylserine Exposure
Promastigotes (1 × 107 cells/mL) were treated or untreated with 30.0, 85.0, and 170.0 μM eupomatenoid-5 for 24 h at 25°C. Afterward, the parasites were washed and resuspended in 100 µL of binding buffer (140 mM NaCl, 5 mM CaCl2, and 10 mM HEPES-Na, pH 7.4), followed by the addition of 5 μL of a calcium-dependent phospholipid binding protein, annexin-V FITC, for 15 min at room temperature. Binding buffer (400 μL) and 50 μL PI were then added. Antimycin A (125.0 μM) was used as a positive control. Data acquisition and analysis were performed using a BD FACSCalibur flow cytometer equipped with CellQuest software. A total of 10,000 events were acquired in the region that was previously established as the one that corresponded to the parasites. Cells that were stained with annexin-V (PI-positive or -negative) were considered apoptotic, and cells that were only PI-positive were considered necrotic [18].
2.9. Determination of Cell Volume of Parasites
Promastigotes (1 × 107 cells/mL) were treated or untreated with 30.0, 85.0, and 170.0 μM eupomatenoid-5 for 24 h at 25°C, harvested, and washed with PBS. Subsequently, the parasites were analyzed using a BD FACSCalibur flow cytometer and CellQuest Pro software. Histograms were generated, and FSC-H represented the cell volume. A total of 10,000 events were acquired in the region that corresponded to the parasites. Actinomycin D (20.0 mM) was used as a positive control [19].
2.10. Determination of Cellular Membrane Integrity
Promastigotes (1 × 107 cells/mL) were treated or untreated with 30.0, 85.0, and 170.0 μM eupomatenoid-5 for 24 h at 32°C, harvested, and washed with PBS. The parasites were incubated with 50 mL of 2 mg/mL PI for 5 min according to the instructions provided by the manufacturer. Immediately thereafter, the parasites were analyzed using a BD FACSCalibur flow cytometer equipped with CellQuest software. A total of 10,000 events were acquired in the region that corresponded to the parasites. Digitonin (40.0 μM) was used as a positive control [19].
2.11. Scanning Electron Microscopy
Promastigotes (1 × 106 cells/mL) were treated or untreated with 30.0 and 85.0 μM eupomatenoid-5 for 48 h at 25°C and fixed in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer for 1–3 h. Subsequently, the parasites were adhered on poly-L-lysine-coated coverslips and dehydrated in increasing concentrations of ethanol. The samples were critical point dried in CO2, coated with gold, and observed in a Shimadzu SS-550 (Japan) scanning electron microscope [20].
2.12. Cell Cycle
Promastigotes (1 × 107 cells/mL) were treated or untreated with 30.0, 85.0, and 170.0 μM eupomatenoid-5 for 24 h at 25°C. After incubation, the parasites were centrifuged and washed twice in PBS (pH 7.4). The resultant pellet was resuspended in 500 μL of a cold methanol/PBS (70% v/v) mixture and maintained at 4°C for 1 h. Afterward, the pellet was centrifuged, resuspended in PBS with 10 μg/mL PI and 20 μg/mL of DNase-free RNase (200 mg), and incubated for 45 min at 37°C. Data were acquired using a BD FACSCalibur flow cytometer and analyzed using CellQuest Pro software [21].
2.13. Statistical Analysis
The data shown in the graphs are expressed as the means ± standard deviation (SD) of the mean of at least three independent experiments. The data were analyzed using one- and two-way analysis of variance (ANOVA). Significant differences among means were identified using the Tukey post hoc test. Values of P ≤ 0.05 were considered statistically significant. Statistical analyses were performed using Prism 5 (GraphPad Software, San Diego, CA, USA, 2007).
3. Results
3.1. Mitochondrial Membrane Potential
Our previous study used transmission electron microscopy and found that eupomatenoid-5 caused damage and significant changes in promastigote mitochondria [13]. Based on this, we decided to evaluate the ΔΨm in eupomatenoid-5-treated parasites using flow cytometry and Rh123, a fluorescent marker that indicates mitochondrial membrane potential. The histograms showed that eupomatenoid-5 decreased total Rh123 fluorescence intensity at all of the concentrations tested compared with the control group, indicating mitochondrial depolarization (Figures 2(a)–2(c)). This loss of ΔΨm was higher at the IC90 (95.9%) than that at the IC50 (76.8%), and at the twofold IC90, the loss of ΔΨm remained the same as the IC90 (94.9%). The positive control, CCCP, induced a decrease of 62.5% in mitochondrial membrane potential.
Figure 2.
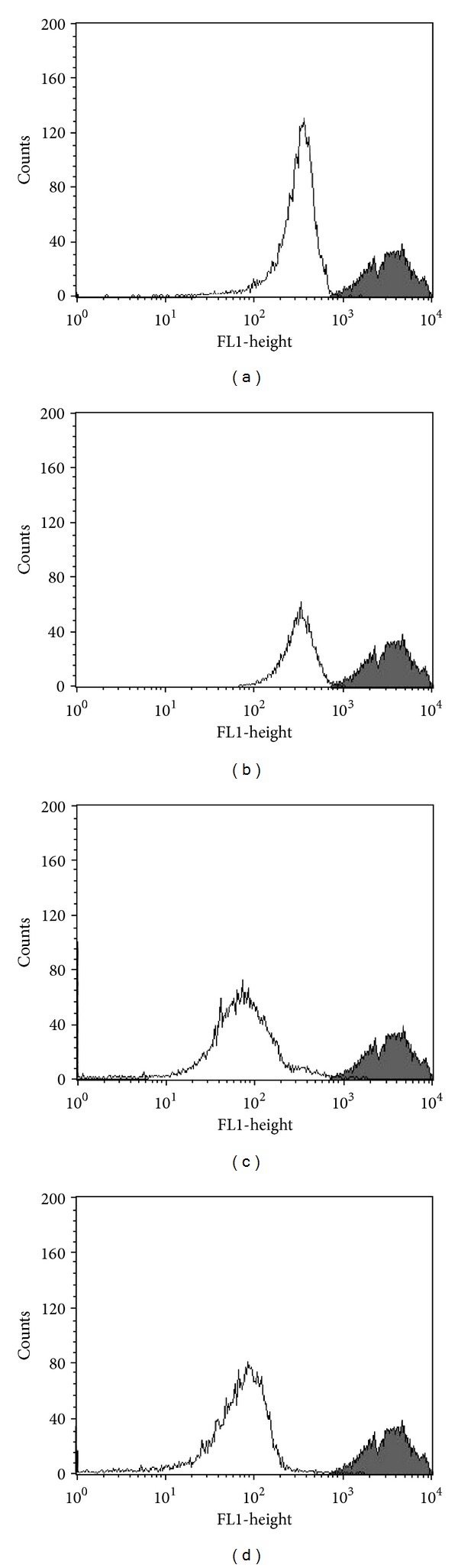
Mitochondrial membrane potential assay in promastigote forms of L. amazonensis treated with eupomatenoid-5 at (b) 30.0 μM, (c) 85.0 μM, and (d) 170.0 μM for 24 h and stained with Rh123, which accumulates in mitochondria. (a) Positive control (CCCP). The gray area corresponds to the control group (i.e., untreated parasites). Typical histograms of at least three independent experiments are shown.
3.2. Mitochondrial-Derived O2 •− Production
Based on our ΔΨm results, we evaluated O2 •− production in eupomatenoid-5-treated parasites. Mitochondrial-derived O2 •− production was evaluated using MitoSOX reagent, which measures the mitochondrial accumulation of superoxide based on its hydrophobic nature and positively charged triphenylphosphonium moiety [20]. Figure 3 shows that eupomatenoid-5 significantly increased the production of mitochondrial O2 •− at most of the concentrations and times tested compared with the control group. After 1 and 2 h of treatment, eupomatenoid-5 increased O2 •− production by approximately 40% only at higher concentrations. In contrast, after 3 and 4 h of treatment, eupomatenoid-5 increased O2 •− production by more than 50% at all of the concentrations tested. The positive control, antimycin A, induced a two-fold increase in mitochondrial O2 •− production.
Figure 3.
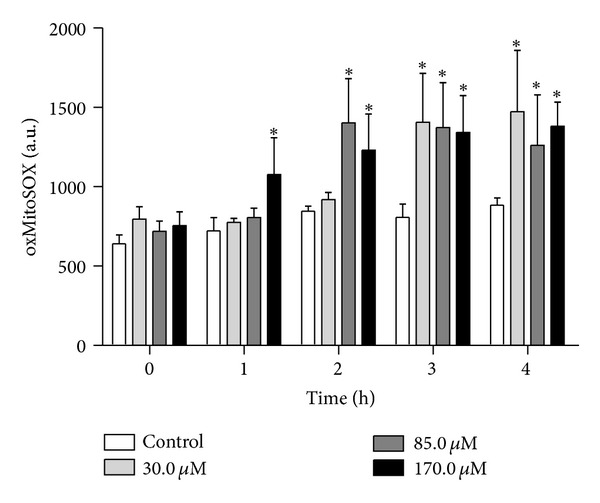
Mitochondrial O2 •− production in promastigote forms of L. amazonensis treated with eupomatenoid-5 at 30.0, 85.0, and 170.0 μM for up to 4 h using the fluorescence probe MitoSOX. At the indicated times, parasites were used to fluorometrically measure oxidized MitoSOX (oxMitoSOX). The data are expressed as the mean fluorescence (in arbitrary units) ± SD of at least three independent experiments. *P ≤ 0.05, significant difference compared with the control group (i.e., untreated parasites).
3.3. Reactive Oxygen Species Level
Based on the MitoSOX data, we evaluated the effects of total ROS production in eupomatenoid-5-treated parasites using a fluorescent probe, H2DCFDA. This probe primarily detects H2O2 and hydroxyl radicals and fluoresces after forming dichlorofluorescein [22]. Our results showed that eupomatenoid-5 increased total ROS production at all of the concentrations and times tested compared with the control group (Figure 4). However, a significant increase in total ROS production of approximately 25% was observed after 3 h of treatment at 85.0 and 170.0 μM. After 24 h treatment, total ROS production increased by 16.3% even at the lower concentration.
Figure 4.
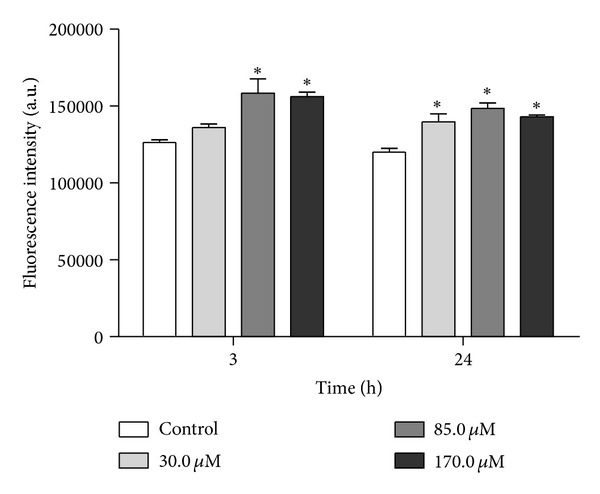
Total ROS production in promastigote forms of L. amazonensis treated with eupomatenoid-5 at 30.0, 85.0, and 170.0 μM for 3 and 24 h using the fluorescence probe H2DCFDA. The data are expressed as the mean fluorescence (in arbitrary units) ± SD of at least three independent experiments. *P ≤ 0.05, significant difference compared with the control group (i.e., untreated parasites).
3.4. Reduced Thiol Levels
Our data suggest that eupomatenoid-5 induces oxidative imbalance, attributable to enhanced ROS production. However, oxidative imbalance conditions depend on both increased oxidant species and decreased antioxidant effectiveness [23]. Therefore our next step was to assess the effect of eupomatenoid-5 on the level of reduced thiols, which might be decreased, for example, by a reduction in trypanothione reductase (TR) activity. Eupomatenoid-5 dose-dependently decreased total reduced thiol levels at all of the concentrations and times tested compared with the control group (Figure 5). This decrease might also be considered time-dependent, with reductions of thiol levels of approximately 10, 15, and 20% after 24, 48, and 72 h treatment, respectively.
Figure 5.
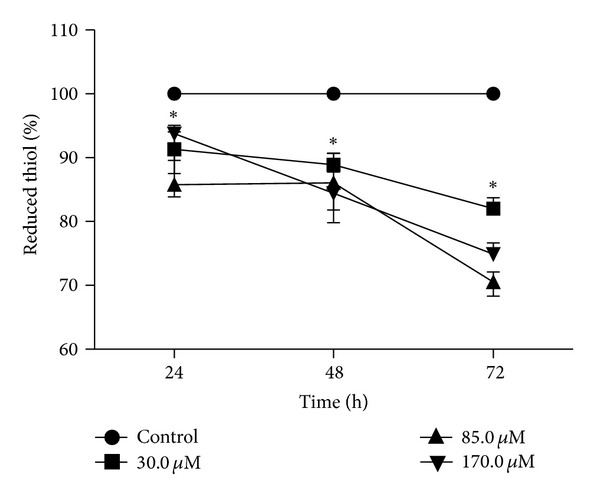
Thiol levels in promastigote forms of L. amazonensis treated with eupomatenoid-5 at 30.0, 85.0, and 170.0 μM for 24, 48, and 72 h using DTNB. The data are expressed as the mean ± SD of at least three independent experiments. *P ≤ 0.05, significant difference compared with the control group (i.e., untreated parasites).
3.5. Phosphatidylserine Exposure
To determine whether the mechanism of cell death triggered by eupomatenoid-5 involves apoptosis, we evaluated the externalization of phosphatidylserine, an apoptotic marker that is present in the outer leaflet of plasmalemma [24] in promastigotes treated with eupomatenoid-5 for 24 h and double stained with FITC-conjugated annexin-V and PI. As shown in Figure 6, eupomatenoid-5 increased annexin-V fluorescence intensity more than 30% at higher concentrations (85.0 and 170.0 μM) compared with the control group, indicating phosphatidylserine exposure (Figures 6(c) and 6(d)).
Figure 6.
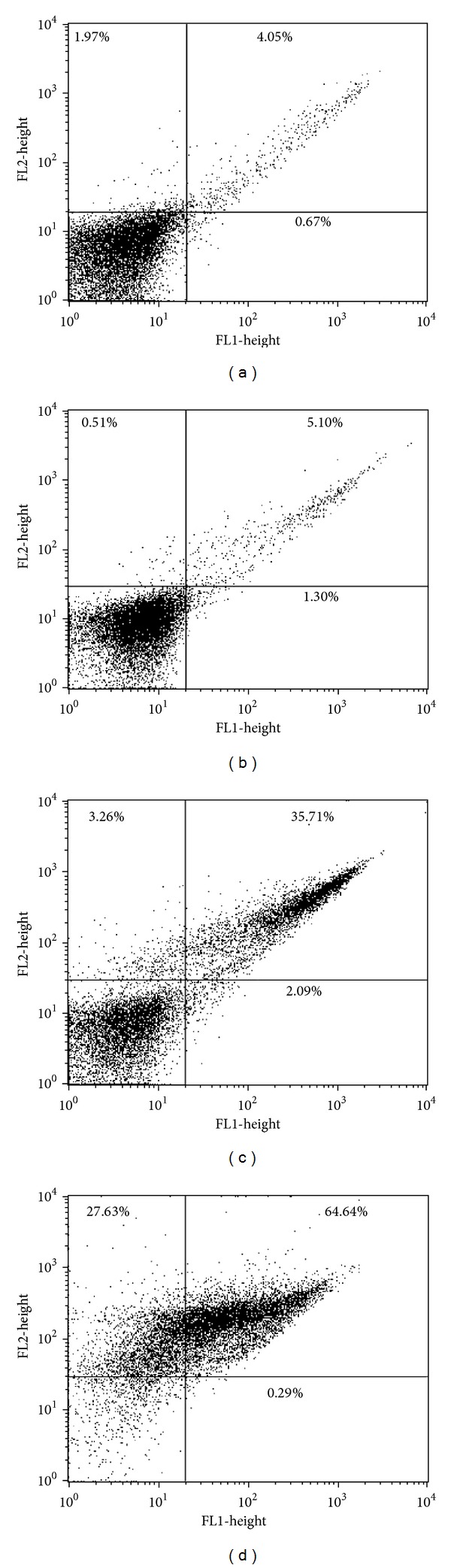
Phosphatidylserine exposure in promastigote forms of L. amazonensis treated with eupomatenoid-5 at (b) 30.0 μM, (c) 85.0 μM, and (d) 170.0 μM for 24 h using annexin-V FITC and PI. (a) corresponds to the control group (i.e., untreated parasites). The results are presented in a dot plot together with the negative control. Typical histograms of at least three independent experiments are shown.
3.6. Cell Volume
In addition to biochemical alterations, apoptosis also induces morphological alterations. Based on this, we evaluated cell shrinkage, a hallmark of apoptotic death, in eupomatenoid-5-treated parasites [25]. As shown in Figure 7, a dose-dependent decrease in cell volume (30.4, 63.1, and 84.3%, resp.) was observed at all of the concentrations tested compared with the control group.
Figure 7.
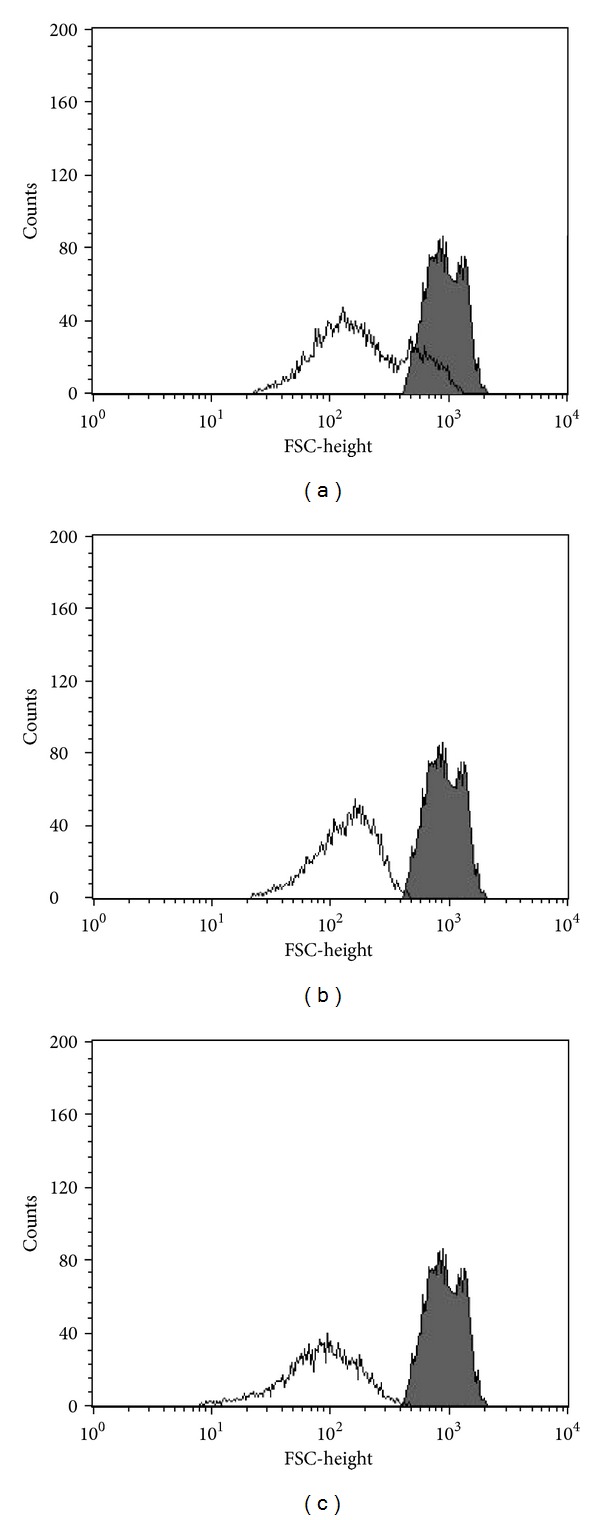
Cell volume in promastigote forms of L. amazonensis treated with eupomatenoid-5 at (a) 30.0 μM, (b) 85.0 μM, and (c) 170.0 μM. FSC-H was considered a function of cell size. The gray area corresponds to the control group (i.e., untreated parasites). Typical histograms of at least three independent experiments are shown.
3.7. Cell Membrane Integrity
To determine whether the mechanism of cell death triggered by eupomatenoid-5 also involves the necrotic death pathway, we evaluated plasma membrane integrity in eupomatenoid-5-treated promastigotes stained with PI, which diffuse across permeable membranes and bind to nucleic acids. As shown in Figure 8, eupomatenoid-5 at 30.0, 85.0, and 170.0 μM dose-dependently increased PI-stained parasites from 1.9% (untreated promastigotes; Figure 8(a), upper quadrants) to 24.3%, 64.6%, and 86.3%, respectively (Figures 8(b)–8(d), upper quadrants), indicating permeabilization of the plasma membrane. The positive control, digitonin, showed a 47.7% increase in the gated percentage of PI-stained cells.
Figure 8.
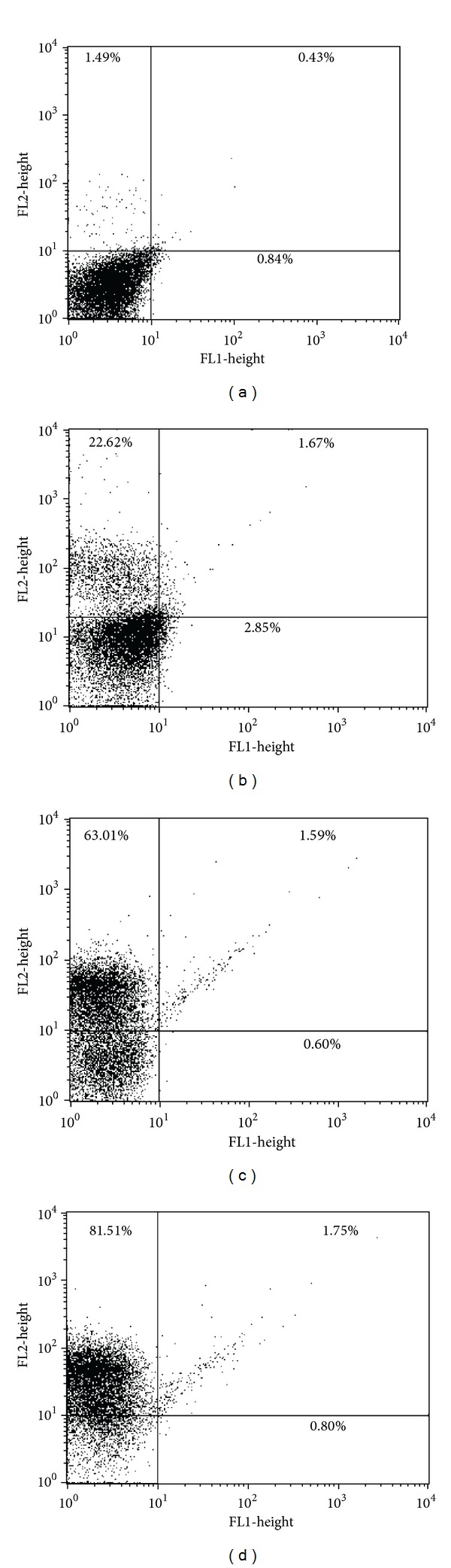
Cell membrane integrity assay in promastigote forms of L. amazonensis treated with eupomatenoid-5 at (b) 30.0 μM, (c) 85.0 μM, and (d) 170.0 μM for 24 h and stained with PI. The numbers show the percentage of PI-positive parasites in (a) and (b). (a) corresponds to the control group (i.e., untreated parasites). Typical histograms of at least three independent experiments are shown.
3.8. Scanning Electron Microscopy
To confirm that eupomatenoid-5 induced morphological alterations in promastigotes, we further evaluated morphological alterations using scanning electron microscopy. Photomicrographs revealed that untreated protozoa had typical characteristics, with an elongated shape and terminal flagellum. In contrast, eupomatenoid-5 dose-dependently altered the size and shape of the treated parasites, including a reduction and rounding of the cellular body (Figure 9).
Figure 9.
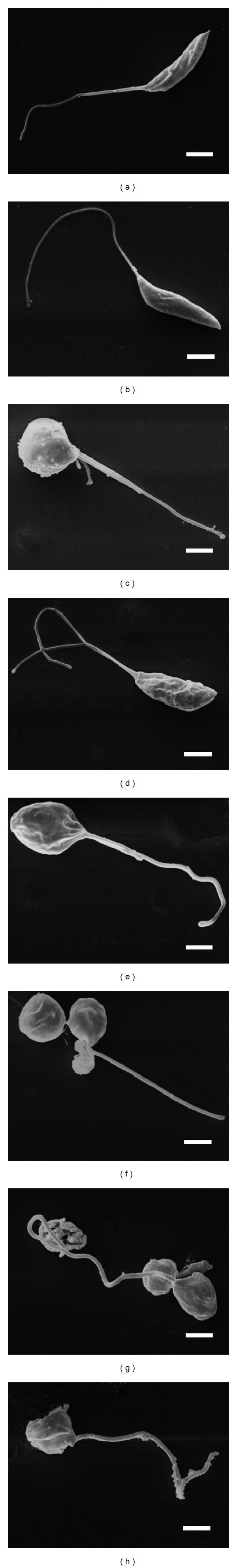
Scanning electron microscopy images of promastigote forms of L. amazonensis incubated in the ((a) and (b)) absence or presence of eupomatenoid-5 at ((c)–(e)) 30.0 μM and ((f)–(h)) 85.0 μM for 48 h. Scale bar = 2 μm.
3.9. Cell Cycle
To evaluate the ratio of pseudohypodiploid cells, flow cytometry after cell permeabilization and PI labeling were used. In a given cell, the amount of bound dye correlates with the DNA content, and thus DNA fragmentation in apoptotic cells is reflected by fluorescence intensity that is lower than that of G0/G1 cells (i.e., a sub-G0/G1 peak) [26]. The incubation of promastigotes with eupomatenoid-5 for 3 and 24 h resulted in a 16% increase in the proportion of cells in the sub-G0/G1 phase at the lower concentration (30.0 μM) and a 28% increase at the higher concentrations (85.0 and 170.0 μM) compared with the control group (Table 1). In contrast, an increase in the number of cells in the sub-G0/G1 phase led to a decrease in the number of cells in the G2/M phase compared with untreated cells.
Table 1.
Effect of eupomatenoid-5 on the cell cycle of L. amazonensis promastigotes (1 × 107 cells/mL) that were treated with 30.0, 85.0, and 170.0 μM eupomatenoid-5 for 24 h and processed for cell cycle analysis as described in Section 2. The data are expressed as percentages.
| Group | Sub-G0/G1 (M1) |
G0/G1 (M2) |
S and G2/M (M3) |
|---|---|---|---|
| Control | 4.16 | 57.31 | 37.16 |
| 30.0 μM | 16.28 | 55.29 | 21.20 |
| 85.0 μM | 28.11* | 47.03 | 16.96 |
| 170.0 μM | 28.41* | 45.44 | 15.84 |
4. Discussion
Despite recent advances, the treatment of leishmaniasis continues to be unsatisfactory. Pentavalent antimonials remain the first-line treatment for this infection in most endemic areas, despite their limitations, such as high toxicity and increased drug resistance [27, 28]. Thus, an urgent need exists to develop new drugs and therapeutic strategies. We have been extensively exploring plant resources to find effective antileishmanial agents. Our previous studies indicated that eupomatenoid-5 contained in the crude extracts and chloroform fractions of P. regnellii leaves are responsible for the antileishmanial activity of this plant [13]. Furthermore, eupomatenoid-5 showed more selective against protozoan than macrophage cells [13]. Additionally, transmission electron microscopy revealed many ultrastructural alterations, especially in the mitochondria of treated parasites, indicating damage and significant changes in this organelle [13]. The present study sought to further elucidate the mechanism of action of eupomatenoid-5 in the cell death of this protozoan.
We initially focused on investigating mitochondrial alterations and their consequences, especially with regard to parasite death. We found that parasites treated with eupomatenoid-5 exhibited a decrease in ΔΨm and increase in mitochondrial ROS production. Leishmania is known to have a single mitochondrion, and so the maintenance of mitochondrial transmembrane potential is essential for parasite survival [29]. In fact, a number of studies have been published describing compounds that target at Trypanosomatids mitochondria [16, 30]. Eupomatenoid-5 also induced a time-dependent decrease in reduced thiol levels of treated parasites. The trypanothione system is unique in trypanosomatid parasites and plays an important role in the homeostasis of parasite redox metabolism [31, 32]. In this system, trypanothione is reduced to a dithiol T(SH)2 by trypanothione/trypanothione reductase (TR). The inhibition of TR decreases total reduced thiol [16]. Our data suggest that eupomatenoid-5 induces oxidative imbalance in promastigote forms through two pathways, increasing ROS production by disrupted mitochondria and decreasing hydroperoxide detoxification by reducing TR activity.
Additionally, eupomatenoid-5 induced the externalization of phosphatidylserine and a reduction of parasite volume, indicated by flow cytometry and scanning electron microscopy. Altogether, our data showed that eupomatenoid-5-treated parasites exhibited apoptotic-like events. An increase in the generation of ROS in the cytosol, an established event in most apoptotic cells, might direct the cell toward this death pathway [33]. Interestingly, programmed cell death in protists appears to share some morphological features with apoptosis in multicellular organisms, including cell shrinkage, the loss of mitochondrial membrane potential, and the externalization of phosphatidylserine [34, 35]. Following phosphatidylserine flip, apoptotic cells lose their plasma membrane integrity. This signal was also induced by eupomatenoid-5 and revealed by the addition of PI, a cellimpermeable nuclear dye [36]. Interestingly, similar results were found in parasitic forms of Trypanosoma cruzi after treatment with eupomatenoid-5 [19].
Finally, we demonstrated that eupomatenoid-5 induced G0/G1 phase cell cycle arrest using flow cytometry and PI labeling, in which the amount of bound dye correlated with the DNA content, and thus DNA fragmentation in apoptotic cells is translated into fluorescence intensity that was lower than that of G0/G1 cells (i.e., a sub-G0/G1 peak) [26, 37].
5. Conclusion
In parasites, apoptosis appears to be the predominant form of cell death [38], which has been observed in kinetoplastids in response to chemotherapeutic agents, such as amphotericin B and plant extracts, such as Aloe Vera leaf exudates [39, 40]. We used biochemical and morphological techniques and demonstrated that eupomatenoid-5 induced cell death, sharing some phenotypic features observed in metazoan apoptosis, including increased ROS, hypopolarization of mitochondrial potential, phosphatidylserine exposure, a reduction of cell volume, and G0/G1 phase cell cycle arrest. The study of the major pathways involved in Leishmania apoptosis-like death will provide insights into the future design of newer chemotherapeutic strategies.
Conflict of Interests
The authors have no conflict of interests.
Acknowledgments
This work was supported through Grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Capacitação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Financiadora de Estudos e Projetos (FINEP), Programa de Pós-Graduação em Ciências Biológicas da Universidade Estadual de Maringá and Complexo de Centrais de Apoio a Pesquisa (COMCAP).
References
- 1.WHO. Control of the Leishmaniasis: Report of a Meeting of the WHO. Geneva, Switzerland: Expert Committee on the Control of Leishmaniasis; 2010. (WHO Technical Report Series no 949). [Google Scholar]
- 2.Macedo-Silva ST, de Oliveira Silva TLA, Urbina JA, de Souza W, Rodrigues JCF. Antiproliferative, ultrastructural, and physiological effects of amiodarone on promastigote and amastigote forms of Leishmania amazonensis . Molecular Biology International. 2011;2011:12 pages. doi: 10.4061/2011/876021.876021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Neto RLM, Sousa LMA, Dias CS, Filho JMB, Oliveira MR, Figueiredo RCBQ. Morphological and physiological changes in Leishmania promastigotes induced by yangambin, a lignan obtained from Ocotea duckei . Experimental Parasitology. 2011;127(1):215–221. doi: 10.1016/j.exppara.2010.07.020. [DOI] [PubMed] [Google Scholar]
- 4.Seifert K, Croft SL. In vitro and in vivo interactions between miltefosine and other antileishmanial drugs. Antimicrobial Agents and Chemotherapy. 2006;50(1):73–79. doi: 10.1128/AAC.50.1.73-79.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Tiuman TS, Ueda-Nakamura T, Garcia Cortez DA, et al. Antileishmanial activity of parthenolide, a sesquiterpene lactone isolated from Tanacetum parthenium . Antimicrobial Agents and Chemotherapy. 2005;49(1):176–182. doi: 10.1128/AAC.49.11.176-182.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ueda-Nakamura T, Mendonça-Filho RR, Morgado-Dıaz JA, et al. Antileishmanial activity of Eugenol-rich essential oil from Ocimum gratissimum . Parasitology International. 2006;55(2):99–105. doi: 10.1016/j.parint.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 7.Santos AO, Ueda-Nakamura T, Dias-Filho BP, Veiga Junior VF, Nakamura CV. Copaiba oil: an alternative to development of new drugs against Leishmaniasis. Evidence Based Complementary and Alternative Medicine. 2012;2012:7 pages. doi: 10.1155/2012/898419.898419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pessini GL, Dias Filho BP, Nakamura CV, Cortez DAG. Antifungal activity of the extracts and neolignans from Piper regnellii (Miq.) C. DC. var. pallescens (C. DC.) Yunck. Journal of the Brazilian Chemical Society. 2005;16(6 A):1130–1133. [Google Scholar]
- 9.Luize PS, Ueda-Nakamura T, Dias Filho BP, Cortez DAG, Nakamura CV. Activity of neolignans isolated from Piper regnellii (Miq.) C. DC. var. pallescens (C. DC.) Yunck against Trypanosoma cruzi . Biological and Pharmaceutical Bulletin. 2006;29(10):2126–2130. doi: 10.1248/bpb.29.2126. [DOI] [PubMed] [Google Scholar]
- 10.Koroishi AM, Foss SR, Cortez DAG, Ueda-Nakamura T, Nakamura CV, Dias Filho BP. In vitro antifungal activity of extracts and neolignans from Piper regnellii against dermatophytes. Journal of Ethnopharmacology. 2008;117(2):270–277. doi: 10.1016/j.jep.2008.01.039. [DOI] [PubMed] [Google Scholar]
- 11.Holetz FB, Pessini GL, Sanches NR, Cortez DAG, Nakamura CV, Dias Filho BP. Screening of some plants used in the Brazilian folk medicine for the treatment of infectious diseases. Memorias do Instituto Oswaldo Cruz. 2002;97(7):1027–1031. doi: 10.1590/s0074-02762002000700017. [DOI] [PubMed] [Google Scholar]
- 12.Sartorelli P, Benevides PJC, Ellensohn RM, Rocha MVAF, Moreno PRH, Kato MJ. Enantioselective conversion of p-hydroxypropenylbenzene to (+)-conocarpan in Piper regnellii . Plant Science. 2001;161(6):1083–1088. [Google Scholar]
- 13.Vendrametto MC, Santos AOD, Nakamura CV, Filho BPD, Cortez DAG, Ueda-Nakamura T. Evaluation of antileishmanial activity of eupomatenoid-5, a compound isolated from leaves of Piper regnellii var. pallescens. Parasitology International. 2010;59(2):154–158. doi: 10.1016/j.parint.2009.12.009. [DOI] [PubMed] [Google Scholar]
- 14.Chauret DC, Bernard CB, Arnason JT, et al. Insecticidal neolignans from Piper decurrens . Journal of Natural Products. 1996;59(2):152–155. doi: 10.1021/np960036y. [DOI] [PubMed] [Google Scholar]
- 15.Menna-Barreto RFS, Goncalves RLS, Costa EM, et al. The effects on Trypanosoma cruzi of novel synthetic naphthoquinones are mediated by mitochondrial dysfunction. Free Radical Biology and Medicine. 2009;47(5):644–653. doi: 10.1016/j.freeradbiomed.2009.06.004. [DOI] [PubMed] [Google Scholar]
- 16.Shukla AK, Patra S, Dubey VK. Iridoid glucosides from Nyctanthes arbortristis result in increased reactive oxygen species and cellular redox homeostasis imbalance in Leishmania parasite. European Journal of Medicinal Chemistry. 2012;54:49–58. doi: 10.1016/j.ejmech.2012.04.034. [DOI] [PubMed] [Google Scholar]
- 17.Piacenza L, Irigoín F, Alvarez MN, et al. Mitochondrial superoxide radicals mediate programmed cell death in Trypanosoma cruzi: cytoprotective action of mitochondrial iron superoxide dismutase overexpression. Biochemical Journal. 2007;403(2):323–334. doi: 10.1042/BJ20061281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jimenez V, Paredes R, Sosa MA, Galanti N. Natural programmed cell death in T. cruzi epimastigotes maintained in axenic cultures. Journal of Cellular Biochemistry. 2008;105(3):688–698. doi: 10.1002/jcb.21864. [DOI] [PubMed] [Google Scholar]
- 19.Lazarin-Bidóia D, Desoti VC, Ueda-Nakamura T, Dias-Filho BP, Nakamura CV, Silva SO. Further evidence of the trypanocidal action of eupomatenoid-5: confirmation of involvement of reactive oxygen species and mitochondria owing to a reduction in trypanothione reductase activity. Free Radical Biology and Medicine. 2013 doi: 10.1016/j.freeradbiomed.2013.01.008. [DOI] [PubMed] [Google Scholar]
- 20.Britta EA, Barbosa Silva AP, Ueda-Nakamura T, et al. Benzaldehyde thiosemicarbazone derived from limonene complexed with copper induced mitochondrial dysfunction in Leishmania amazonensis . PLoS ONE. 2012;7(8):414–440. doi: 10.1371/journal.pone.0041440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ferreira C, Soares DC, Barreto-Junior CB, et al. Leishmanicidal effects of piperine, its derivatives, and analogues on Leishmania amazonensis . Phytochemistry. 2011;72(17):2155–2164. doi: 10.1016/j.phytochem.2011.08.006. [DOI] [PubMed] [Google Scholar]
- 22.Wan CP, Myung E, Lau BHS. An automated micro-fluorometric assay for monitoring oxidative burst activity of phagocytes. Journal of Immunological Methods. 1993;159(1-2):131–138. doi: 10.1016/0022-1759(93)90150-6. [DOI] [PubMed] [Google Scholar]
- 23.Leal CA, Shetinger MR, Leal DB, et al. Oxidative stress and antioxidant defenses in pregnant women. Redox Report. 2011;16:230–236. doi: 10.1179/1351000211Y.0000000013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Debrabant A, Nakhasi H. Programmed cell death in trypanosomatids: is it an altruistic mechanism for survival of the fittest? Kinetoplastid Biology and Disease. 2003;2, article 7 doi: 10.1186/1475-9292-2-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kroemer G, Galluzzi L, Vandenabeele P, et al. Classification of cell death: recommendations of the Nomenclature Committee on Cell Death 2009. Cell Death and Differentiation. 2009;16(1):3–11. doi: 10.1038/cdd.2008.150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Khademvatan S, Gharavi MJ, Saki J. Miltefosine induces metacaspase and PARP genes expression in Leishmania infantum . Brazilian Journal of Infectious Disease. 2011;15(5):442–448. doi: 10.1016/s1413-8670(11)70225-2. [DOI] [PubMed] [Google Scholar]
- 27.Croft SL, Sundar S, Fairlambm AH. Drug resistance in leishmaniasis. Clinical Microbiology Review. 2006;19:111–126. doi: 10.1128/CMR.19.1.111-126.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Natera S, Machuca C, Padrón-Nieves M, Romero A, Díaz E, Ponte-Sucre A. Leishmania spp.: proficiency of drug-resistant parasites. International Journal of Antimicrobial Agents. 2007;29(6):637–642. doi: 10.1016/j.ijantimicag.2007.01.004. [DOI] [PubMed] [Google Scholar]
- 29.Kowaltowski AJ, Vercesi AE. Mitochondrial damage induced by conditions of oxidative stress. Free Radical Biology and Medicine. 1999;26(3-4):463–471. doi: 10.1016/s0891-5849(98)00216-0. [DOI] [PubMed] [Google Scholar]
- 30.Fernandes MP, Inada NM, Chiaratti MR, et al. Mechanism of Trypanosoma cruzi death induced by Cratylia mollis seed lectin. Journal of Bioenergetics and Biomembranes. 2010;42(1):69–78. doi: 10.1007/s10863-010-9268-9. [DOI] [PubMed] [Google Scholar]
- 31.Fairlamb AH, Blackburn P, Ulrich P. Trypanothione: a novel bis(glutathionyl)spermidine cofactor for glutathione reductase in trypanosomatids. Science. 1985;227(4693):1485–1487. doi: 10.1126/science.3883489. [DOI] [PubMed] [Google Scholar]
- 32.Fairlamb AH, Cerami A. Metabolism and functions of trypanothione in the kinetoplastida. Annual Review of Microbiology. 1992;46:695–729. doi: 10.1146/annurev.mi.46.100192.003403. [DOI] [PubMed] [Google Scholar]
- 33.Chipuk JE, Green DR. Do inducers of apoptosis trigger caspase-independent cell death? Nature Reviews Molecular Cell Biology. 2005;6(3):268–275. doi: 10.1038/nrm1573. [DOI] [PubMed] [Google Scholar]
- 34.Arambage SC, Grant KM, Pardo I, Ranford-Cartwright L, Hurd H. Malaria ookinetes exhibit multiple markers for apoptosis-like programmed cell death in vitro . Parasites and Vectors. 2009;2(1, article 32) doi: 10.1186/1756-3305-2-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Duszenko M, Figarella K, Macleod ET, Welburn SC. Death of a trypanosome: a selfish altruism. Trends in Parasitology. 2006;22(11):536–542. doi: 10.1016/j.pt.2006.08.010. [DOI] [PubMed] [Google Scholar]
- 36.Bouchier-Hayes L, Muñoz-Pinedo C, Connell S, Green DR. Measuring apoptosis at the single cell level. Methods. 2008;44(3):222–228. doi: 10.1016/j.ymeth.2007.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Sen R, Bandyopadhyay S, Dutta A, et al. Artemisinin triggers induction of cell-cycle arrest and apoptosis in Leishmania donovani promastigotes. Journal of Medical Microbiology. 2007;56(9):1213–1218. doi: 10.1099/jmm.0.47364-0. [DOI] [PubMed] [Google Scholar]
- 38.Arnoult D, Akarid K, Grodet A, Petit PX, Estaquier J, Ameisen JC. On the evolution of programmed cell death: apoptosis of the unicellular eukaryote Leishmania major involves cysteine proteinase activation and mitochondrion permeabilization. Cell Death and Differentiation. 2002;9(1):65–81. doi: 10.1038/sj.cdd.4400951. [DOI] [PubMed] [Google Scholar]
- 39.Dutta A, Mandal G, Mandal C, Chatterjee M. In vitro antileishmanial activity of Aloe vera leaf exudate: a potential herbal therapy in leishmaniasis. Glycoconjugate Journal. 2007;24(1):81–86. doi: 10.1007/s10719-006-9014-z. [DOI] [PubMed] [Google Scholar]
- 40.Dutta A, Mandal G, Mandal C, Chatterjee M. Racemoside A, an antileishmanial, water-soluble, natural steroidal saponin, induces programmed cell death in Leishmania donovani. Journal of Medical Microbiology. 2007;56:1196–1204. doi: 10.1099/jmm.0.47114-0. [DOI] [PubMed] [Google Scholar]