

Abstract
Nearly all research on camouflage has investigated its effectiveness for concealing stationary objects. However, animals have to move, and patterns that only work when the subject is static will heavily constrain behaviour. We investigated the effects of different camouflages on the three stages of predation—detection, identification and capture—in a computer-based task with humans. An initial experiment tested seven camouflage strategies on static stimuli. In line with previous literature, background-matching and disruptive patterns were found to be most successful. Experiment 2 showed that if stimuli move, an isolated moving object on a stationary background cannot avoid detection or capture regardless of the type of camouflage. Experiment 3 used an identification task and showed that while camouflage is unable to slow detection or capture, camouflaged targets are harder to identify than uncamouflaged targets when similar background objects are present. The specific details of the camouflage patterns have little impact on this effect. If one has to move, camouflage cannot impede detection; but if one is surrounded by similar targets (e.g. other animals in a herd, or moving background distractors), then camouflage can slow identification. Despite previous assumptions, motion does not entirely ‘break’ camouflage.
Keywords: motion camouflage, background matching, crypsis, disruptive patterns
1. Introduction
Predation is a major factor in selection. The process of predation can be broken down into three stages: detection, identification and capture. Initially, a predator must detect that there is a potential target present; then, it must identify whether that target really is a prey item; and finally, it can attempt to capture the target. This means that camouflage can act at any or all of these stages to reduce the chance of successful predation.
For an animal to conceal itself, there are a number of strategies it can use. It can blend with the background (background matching, BM), break up its shape (disruptive camouflage) or mimic irrelevant objects (masquerade) [1–3]. Here, we focus on BM and different sub-types of disruptive coloration that have been discussed in the recent literature.
BM is the most intuitive method for concealing a target. At its most basic, it involves coloration that represents a random sample of the background [1]. This type of camouflage functions to avoid the target ever being detected (the first stage of predation), and achieves this simply by increasing the similarity of the target and background [4] so that discrimination between the two is impeded [5].
Disruptive camouflage provides a useful addition to BM, when multiple habitats are involved. Thayer [6] and Cott [7] proposed that the two most important criteria for disruptive camouflage are differential blending and maximum disruptive contrast. Differential blending means that some patches of the pattern will blend into the background while others stand out. Empirical research supports this principle, but only that adjacent patches or pattern elements should have high contrast, not the greatest possible contrast [8,9]. Together, differential blending with the background and internal contrast between adjacent colour patches cause the appearance of false (non-object-bounding) edges. The patches that stand out appear as separate objects from the patches that appear to be background. The overall effect helps to break up the outline or continuity of the surface [10].
These disruptive markings can be placed near to the edge of the animal (external) or closer to the centre (internal). External markings help to break up the outline of the body, whereas internal markings may help to mask a potentially lower contrast real outline via lateral inhibition, or act to distract attention away from the edges [11]. In both cases, disruptive markings help to disguise the shape of an object [3,6–8,12], and so potentially provide effective camouflage in multiple habitats. A number of studies have shown disruptive patterns positioned around the edge of a shape to be more successful at preventing predation than disruptive patterns positioned in the centre [8,9,13]. Another study [11], however, found the opposite effect, showing that centrally placed markings can produce surface disruption effects and thus may be more effective for camouflage than those placed at the edge.
Nearly all research on camouflage has investigated its effectiveness for concealing stationary objects. However, both animals and military personnel require the ability to move, and a pattern that only works when the subject is static will heavily constrain behaviour. Attention is quickly drawn to motion. Moving objects ‘pop out’ and are easily observed independent of the number of other stationary objects that are present [14]. Another issue presented by motion, is that when the background contains any spatial pattern, movement of an object causes figure-ground segregation to become trivial [15]. The edges of the moving object become clearly defined as it moves across the background pattern.
Together, these effects mean that an individual target moving on its own is highly conspicuous [16]. Under stationary conditions camouflage patterns (BM and disruptive) have been found to be successful at preventing the capture of a target. However, the same camouflage patterns have been shown to be ineffective at reducing capture rates for moving targets [16]. Therefore, it seems camouflage patterns are effective when stationary and ineffective, but not necessarily costly, during motion.
How camouflage operates in moving groups of animals may be a different matter. The confusion effect is defined as the reduction of the attack-to-kill ratio, in predators, owing to an inability to single out and attack an individual when prey are aggregated [17]. When multiple similar targets are present, increased processing is required, resulting in decreased search efficiency [18]. To maximize the effect, multiple individuals should be of similar appearance [19]. Odd individuals, in a group that otherwise match each other, reduce the confusion effect and are much easier for the predator to target [20]. Therefore, the confusion effect may provide a useful mechanism to hinder identification of a moving target amongst other moving individuals, as long as all the objects are of similar appearance.
The visual search literature also provides evidence that under static conditions the presence of distractors can slow search efficiency. In order for distractors to affect identification of a target, they should be similar in appearance to the target [21]. However, targets are harder to detect when all the distractors are similar to the target, but not identical to each other [21,22]. Motion-defined visual search literature has centred around search for the conjunction of movement and form, which has been shown to be a parallel process [23]. Visual search appears to be remarkably robust against motion [24]. Most of the literature in this area involves synchronized directions of movement; however, there is evidence that search is less efficient when distractors move in multiple directions [25].
Over the years, it has been generally assumed that camouflage (such as BM and disruptive patterns) only works when the target is stationary. This assumption has resulted in a common phrase to sum up the situation, ‘motion breaks camouflage’. However, there have been very few empirical studies that specifically investigate this effect and while it is intuitive that camouflage is unlikely to prevent the detection of a moving target, there are other methods by which camouflage could affect predation of a moving object, such as impeding target recognition or enhancing the confusion effect.
We, therefore, created seven camouflage patterns based on the strategies of BM and disruption, and compared them with a no pattern control. Three of the patterns bore elements of previously tested strategies (BM; disruptive edge (DE); and disruptive centre (DC); [8,26]). The other four were simple variations on disruptive patterning (step edge (SE); step centre (SC); graduating edge (GE); graduating centre (GC)), where the boundary between the high- and low-contrast regions was either sharp (SE and SC) or graduated (GE and GC). These variants were chosen because of their differences in the location and sharpness of their internal patterns. Sharp discontinuities might be more detectable because they are characteristic of the boundaries that our visual system exploits for object recognition [27] but, placed appropriately, sharp discontinuities can create false boundaries that may either disrupt the true outline or, placed internally, mask or distract attention from the true object edge [2,3,8,9,11,26].
We then used three computer-based experiments to investigate how camouflage can affect the different stages of predation. In the first experiment, we tested the camouflage patterns on stationary targets in order to verify previous demonstrations of their effectiveness for static viewing [8,9,11] when applied to our model system. All of the camouflage patterns were predicted to perform better than a no-pattern control (NP). BM and disruptive patterns were expected to be the most successful strategies, and because this is the most commonly reported finding in the literature, we predicted that external disruptive patterns would be more effective than internal ones. When motion was added in the second experiment, it was predicted that all effects of camouflage would disappear and all targets would be captured equally effectively. Finally, multiple distractors were added in the third experiment, and it was predicted that identification rate would be slowed in the presence of distractors that matched the target.
2. Material and methods
All participants were naive, had normal or corrected-to-normal vision, and gave informed consent in accordance with the Declaration of Helsinki. All trials were presented on a gamma-corrected, 22″, 1024 × 768 pixel laCie Electron 22Blue monitor with a refresh rate of 100 Hz and a mean luminance of 21.7 cd m−2. Data have been deposited in the Dryad digital repository (doi:10.5061/dryad.vc60q).
(a). Experiment 1
Backgrounds were static in all experiments. A new background and target were generated for each trial. For the background, a first-order autoregressive spatial process with normal error distribution [28] was used to create two different, high-contrast patterns. These were combined and manipulated (see the electronic supplementary material for details) in order to produce a grey scale background pattern (size: 13.2° × 13.2°) with a coarse, local and spatial structure. This meant that randomly placed, static, BM targets had small discontinuities at their boundaries; without such spatial structure, static versions of these targets would have been completely undetectable.
Irregular shapes (targets) were generated from similar background patterns in three sizes (1.0°, 1.4° and 2.0° in mean diameter) and eight different patterns: a NP (mean luminance of background) and seven different camouflage patterns (figure 1). These camouflage patterns were chosen to instantiate elements of three strategies (BM; edge disruptive with high-contrast patches at the target edge, DE; and surface-disruptive with central high-contrast patches, DC). These have been previously proposed to be effective camouflage for static targets [4–6,12]. In addition, four simple variations on these targets were created where the boundary between the high and low-contrast regions within a target was either sharp (SE and SC) or graduated (GE and GC).
Figure 1.

Sample stimuli patterns. Artificial patterns were generated and used for stimuli: (a) no pattern (NP), (b) background matching (BM), (c) step edge (SE), (d) step centre (SC), (e) graduating edge (GE), (f) graduating centre (GC), (g) disruptive edge (DE), and (h) disruptive centre (DC). Each pattern implements a camouflage strategy previously proposed in the literature.
Each trial consisted of a stationary target displayed on the monitor, and participants detected and clicked on the target as quickly as possible, using the mouse to move a cursor. Response times (RTs) were recorded. Ten repetitions of each pattern at each size were presented in random order, giving a total of 240 trials per subject. Ten naive participants (five males and five females) were tested and were recruited from the postgraduate population at the University of Bristol and reimbursed for their time.
(b). Experiment 2
Conditions were the same as experiment 1, but in this case, the targets moved at 4 degrees per second in a straight line until encountering a boundary, at which point they rebounded at an angle equal to the angle of incidence. RTs, position of the target and accuracy of the mouse-click with respect to target location were recorded. The same 10 participants as in the previous experiment were tested.
(c). Experiment 3
The background and target patterns from the previous two experiments were used, but the target was now an ellipse (minor axis = 0.7 × major axis: major axis 1.63°; minor axis 1.13°) with its major axis oriented either vertically or horizontally. The orientation was assigned randomly from trial to trial, with the caveat that there was an equal number of each orientation within each condition. Circular distractors (1.4° diameter with the same area as ellipses) were displayed along with the target. Each distractor was generated individually to ensure that while they displayed the same type of pattern they were not identical. There were 0, 5 or 10 distractors in one of two combinations with the targets. Either the target and distractors contained the same pattern, or the target contained a pattern and the distractors were plain. The condition with NP targets and plain distractors falls into both of these categories but was only presented once. The trials were blocked by distractor pattern (plain or patterned), and counterbalanced across participants. The trials within a block were presented in random order. The target and distractors moved at 4 degrees per second in any direction with no occlusion; instead they rebounded off each other as well as the boundary at an angle equal to the angle of incidence. Participants were asked to identify the elliptical target's orientation as quickly and accurately as possible with a key press. RT and correctness of response were recorded for each trial. Sixteen participants were recruited from the undergraduate population at the University of Bristol, completing the experiment for course credit. Only participants who scored a minimum of 90 per cent correct on both experimental blocks were included in the analysis, as this allowed the assumption that incorrect trials were due to non-systematic error rather than a differential speed-accuracy trade-off across blocks. Analysis was carried out on all correct trials of the 12 participants (11 females and one male) meeting this criterion. Within this group, there were no systematic differences in errors across conditions. Median RTs for each participant in each condition were calculated, and different conditions were then compared using repeated-measures ANOVA with Greenhouse–Geisser adjustment of the degrees of freedom to account for within-subject correlations [29]. Values are quoted as mean ± s.e.m.
3. Results
(a). Experiment 1: detection and capture of static targets
In this experiment stationary targets were displayed on-screen and participants were asked to capture the target as quickly as possible using a mouse-click. Performance was assessed via capture accuracy and RTs, and camouflage patterns were compared with an unpatterned control of the same mean luminance as the background (NP, figure 1). Three different target sizes were tested.
Capture accuracy and RTs were affected by target size, pattern and an interaction between the two (capture accuracy: size, F2,18 = 43.81, p < 0.001; pattern, F7,63 = 31.21, p < 0.001; interaction, F14,126 = 2.83, p = 0.001; RTs: size, F1.10,9.87 = 22.38, p < 0.001; pattern, F1.58,14.23 = 14.32, p < 0.001; interaction, F3.62,32.55 = 2.40, p = 0.005). For the smallest target size (1.0 deg, figure 2a), four camouflage patterns (BM, SC, GE and GC) clearly increased RTs compared with the NP control (F2.45,22.01 = 8.72, p < 0.001; pairwise comparisons: NP versus BM, F1,9 = 13.22, p = 0.005; NP versus SC, F1,9 = 16.84, p = 0.003; NP versus DE, F1,9 = 16.14, p = 0.003; NP versus DC, F1,9 = 25.30, p = 0.001).
Figure 2.
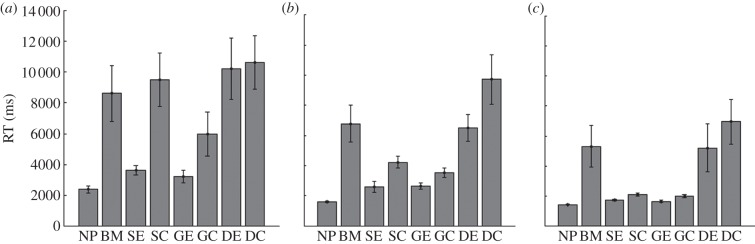
Mean median response times (RT) (10 participants, ±s.e.m.) for detection and capture of three different static target sizes: (a) 1.0°, (b) 1.4°, and (c) 2.0°. As target size increases the success of the camouflage types decreases, leaving background matching, DE and DC as the most successful patterns for the largest target size. See figure 1 for patterns used.
As the targets increased in size and the task became easier (figure 2b,c), SC became less effective, leaving BM, DE and DC as the most successful camouflage patterns for the largest targets (F1.89,17 = 6.81, p = 0.007; pairwise comparisons: NP versus BM, F1,9 = 7.64, p = 0.022; NP versus DE, F1,9 = 5.62, p = 0.042; NP versus DC, F1,9 = 13.64, p = 0.005). As can be seen, at least three of the camouflage strategies (BM, DE and DC), produce a significant lengthening of capture times in all conditions. The capture accuracy results mimicked the RT results, showing that when targets were hard to find, the trial took longer and the participant was less likely to be correct. This shows that the BM, DE and DC patterns (which contain disruptive elements of varying types) as well as matching the background, function as proposed when targets are static, a precondition of our subsequent investigations.
(b). Experiment 2: detection and capture of moving targets
The second experiment was similar to the first, but with the important difference that the objects moved. The addition of motion caused all RTs to fall dramatically compared with experiment 1. The target patterns all produced very similar RTs, with no difference between the different camouflage patterns, or between the camouflage and no-pattern targets (figure 3). There was an effect of size, with larger targets being captured more quickly than smaller targets (size, F1.15,10.36 = 60.01, p < 0.001; pattern, F7,63 = 1.621, p = 0.146; interaction, F14,126 = 0.78, p = 0.690).
Figure 3.
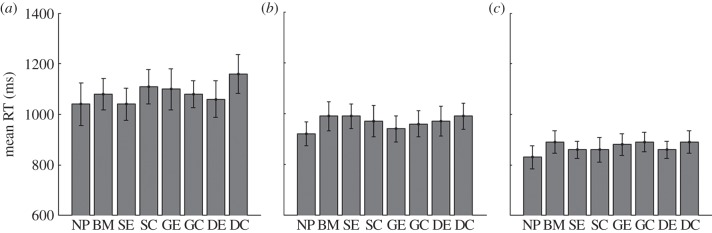
Mean median response times (RT) (10 participants, ±s.e.m.) for detection and capture of moving targets of sizes (a) 1.0°, (b) 1.4°, and (c) 2.0°. There were no differences between the target patterns (figure 1 for patterns used). RTs fell with increased target size for all patterns. Note that the RTs in this experiment were approximately a factor of 10 faster than those in the previous experiment (illustrated in figure 2).
The position of the mouse-click was recorded relative to the position of the target. There was no difference between capture accuracy (hit or miss) for the different patterns. However, there was an effect of size with large targets being captured more accurately (size, F2,18 = 49.62, p < 0.001; pattern, F7,63 = 1.32, p = 0.257; interaction, F14,126 = 0.72, p = 0.751). The distances of the misses (measured from the outer edge of the target), however, did not differ between different target sizes or camouflage patterns (size, F1.38,12.39 = 1.449, p = 0.263; pattern, F1.58,14.18 = 1.001, p = 0.373; interaction, F1.48,13.28 = 1.103, p = 0.341). The main result of this experiment is a simple one: as can be seen from figure 3, once a target moves, detection is relatively rapid, and camouflage has no significant effect on capture times. Motion breaks camouflage.
(c). Experiment 3: identification of moving targets in the presence of distractors
The same target patterns as in the previous experiments were displayed on elliptical targets amongst circular distractors. Each trial involved a moving target amongst 0, 5 or 10 moving distractors. Participants were required to identify the target and indicate its orientation (horizontal or vertical) as quickly as possible. Owing to the conditions involved (we did not include a condition with plain targets and patterned distractors), this experiment could not be a fully factorial design, and so separate analyses for each subset of conditions are presented. In trials with no distractors, there was no significant effect of any of the target patterns on the speed of identification (F3.2,35.2 = 2.447, p = 0.076).
When the distractors were of type NP, and did not match the target, identification did not differ between target patterns and was just as fast as in the absence of distractors. This was true when there were either five or 10 distractors (five distractors, F6,66 = 1.0, p = 0.433; 10 distractors, F6,66 = 0.745, p = 0.615), and RTs were similar to those for the no distractor condition. The speed at which targets were identified under this condition suggests that search was parallel for a conjunction of movement, form and texture. This is consistent with McLeod et al. [23] who reported that search for a conjunction of movement and form is a parallel process.
When instead the distractors matched the targets, there were significant differences between RTs for the different target patterns (five distractors, F7,77 = 4.74, p < 0.001; 10 distractors, F7,77 = 6.06, p < 0.001). This effect is owing to shortened RTs for the no-pattern stimuli in comparison with the camouflage stimuli (figure 4): any camouflage pattern is better than none. For five distractors, the no-pattern condition was significantly different from all the camouflage patterns (pairwise comparisons: NP versus BM, F1,11 = 14.96, p = 0.003; NP versus SE, F1,11 = 11.80, p = 0.006; NP versus SC, F1,11 = 11.99, p = 0.005; NP versus GE, F1,11 = 15.40, p = 0.002; NP versus GC, F1,11 = 22.33, p = 0.001; NP versus DE, F1,11 = 13.51, p = 0.004; NP versus DC, F1,11 = 15.53, p = 0.002; all remain significant after Bonferroni control for multiple testing). Excluding the no-pattern targets from the analysis leaves no remaining differences between the camouflage patterns (F6,66 = 0.614, p = 0.719).
Figure 4.
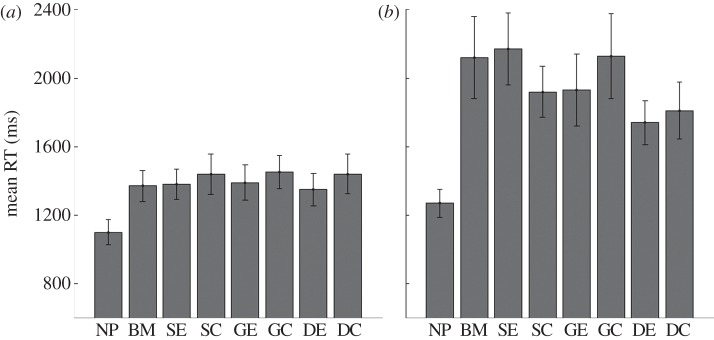
Mean median response times (RT) (12 participants, ±s.e.m.) for target identification with (a) five distractors and (b) 10 distractors. In both cases RTs are longer for the camouflage patterns than the no-pattern condition. RTs are also generally longer when there are more distractors present. See figure 1 for patterns used.
For 10 distractors, the no-pattern condition was also different from all the camouflage patterns (pairwise comparisons: NP versus BM, F1,11 = 17.99, p = 0.001; NP versus SE, F1,11 = 36.71, p < 0.001; NP versus SC, F1,11 = 27.05, p < 0.001; NP versus GE, F1,11 = 23.18, p = 0.001; NP versus GC, F1,11 = 15.21, p = 0.002; NP versus DE, F1,11 = 31.2, p < 0.001; NP versus DC, F1,11 = 19.68, p = 0.001; all significant after Bonferroni correction). Again, exclusion of the no-pattern targets from the analysis revealed no remaining differences between the camouflage patterns (F6,66 = 1.89, p = 0.096).
The number of distractors also affected RTs. When distractors matched the targets, the RTs in trials involving 10 distractors (1886±64 ms) were much longer than for those involving five distractors (1365±32 ms) (paired samples t-test, t95 = 10.24, p < 0.001). This effect was also apparent for the condition where the distractors did not match the target, although the difference was much smaller (10 distractors 790±12 ms; five distractors 754±10 ms; paired samples t-test, t83 = 4.12, p < 0.001). This pattern of results for the identification of moving targets was very different from that for the capture of a single target: camouflage inhibits identification, and does so more when there are more distractors, but the particular nature of the camouflage patterns has no detectable influence of the size of this effect.
4. Discussion
All camouflage patterns tested increased RTs for stationary targets when compared with an unpatterned target, and so all the camouflage patterns were effective. Furthermore, of all the strategies, BM and disruptive (both edge and centre) patterns were the most successful types of camouflage, neatly coinciding with the fact that these are the pattern strategies that occur most commonly in the natural world [6,7,10].
Several authors have emphasized how disruptive and BM camouflages act together. Cott [7] clarified Thayer's [6] proposal that the most effective disruptive camouflage should involve patches of BM coloration alongside areas of high-contrast patterning (a process he called differential blending). The areas of high contrast will create false bounding contours [10] and can be positioned either near the edge of the animal (external) or near to the centre (internal). The results from experiment 1 support the idea that disruptive camouflage requires both differential blending and disruptive contrast, because the step and graduating patterns, which did not include both factors, were less successful at hiding the target. Other studies have found that edge disruptive patterns provide additive benefits on top of BM [8,9], or that central disruptive patterns are more effective than ones located on the target's edge [11]. We found no evidence to support either of these previous results. This could be attributable to any or all of the differences in background structure, target design (e.g. our BM targets, by virtue of being random background samples, also had edge-disrupting elements), protocol and species. This issue is not central, given that our interest is in the effect of movement.
Attention is quickly drawn to motion [14]. Motion relative to a background allows figure-ground segregation [15] as long as the target, background or both contain some spatial pattern. This makes the large difference we observed in RTs between the stationary and dynamic conditions unsurprising: the camouflage patterns that were found to be successful at hiding the position of stationary targets for detection and capture were ineffective once the target moved. Consistent with Stevens et al. [16], it is clear from our data that camouflage strategies such as BM and disruptive patterns are not useful for reducing predation at either the detection or capture stages, if the target is in motion. Interestingly, feedback from regular soldiers about ineffectiveness of camouflage when moving is ostensibly one reason why the United States Army dispensed with camouflage for all but snipers and reverted to monochrome olive drab at the end of second world war [30].
The third experiment did, however, provide evidence that camouflage patterns can slow down the process of being identified while moving. The camouflage patterns did not prevent identification, but RTs were slower for targets with camouflage compared with those without as long as other camouflaged shapes were also present and in motion. There was no difference between the different camouflage patterns tested. The results, therefore, suggest that the details of the pattern are unimportant in hindering identification while moving: any pattern which displayed some similarity to the background and to other moving objects had a similar effect. While the effect of the camouflage is not large enough to prevent identification completely, it can slow the process by up to 70 per cent (for the most successful camouflage pattern, with 10 matching distractors). This could provide a long enough delay to increase a prey item's chance of escaping.
When the target was the only moving object, the camouflage patterns had no effect. The same result was seen when there were distractors that did not match the target. It, therefore, appears that it is the presence of other similarly camouflaged moving objects that helps to conceal the identity of a target. This effect grows as the number of distractors increases.
What is the relevance to the real world? Natural backgrounds often have moving elements (leaves, twigs, etc.), and BM animals sample from these elements or, in masquerade [31], mimic the elements. As long as the motion of the animal and background elements are similar [32], this would constitute a target–distractor discrimination task analogous to that modelled in experiment 3. This pattern of results can also be applied to herding/shoaling animals that may take advantage of such an effect to confuse predators. With the same pattern evident on many individuals, a predator's task of identifying and isolating a weaker, younger or more profitable individual is hindered. This may increase the ‘confusion effect’, as reported in fish [33], reptilian and primate [34] predators, where predator attacks on groups are delayed, compared with those on solitary individuals. Although targets and distractors in experiment 3 differed only along a single dimension (a minor–major axis difference), addition of camouflage patterns, although irrelevant to the discrimination, seems to have decreased search efficiency, an effect proposed to be central to the confusion effect [18].
5. Conclusions
Camouflaging moving objects is a difficult problem because the movement ‘pops out’ [14]. Camouflage, such as BM and disruptive coloration, is unsuccessful at preventing the detection or capture of single moving targets. However, a pattern with similarities to the background may be effective in slowing identification of a target in the presence of other moving objects displaying the same pattern. The ‘distractors’ could be moving elements of the background or conspecifics. In short, when you have to move, it is better to be camouflaged and among companions than plain and isolated. We conclude, therefore, that motion does not entirely ‘break’ camouflage.
Acknowledgements
The study was approved by the Ethics Committee of the Faculty of Science of the University of Bristol.
This research was supported by a CASE Studentship to J.R.H. funded by the EPSRC and QinetiQ. We are grateful to three anonymous referees for their insightful comments on a previous version of this manuscript and to Jens Krause and Ute Leonards for discussion of the work upon which it is based.
References
- 1.Ruxton GD, Sherratt TN, Speed MP. 2004. Avoiding attack: the evolutionary ecology of crypsis, warning signals and mimicry. Oxford, UK: Oxford University Press [Google Scholar]
- 2.Stevens M. 2007. Predator perception and the interrelation between different forms of protective coloration. Proc. R. Soc. B 272, 1457–1464 10.1098/rspb.2007.0220 (doi:10.1098/rspb.2007.0220) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Stevens M, Merilaita S. 2009. Defining disruptive coloration and distinguishing its functions. Phil. Trans. R. Soc. B 364, 481–488 10.1098/rstb.2008.0216 (doi:10.1098/rstb.2008.0216) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Webster RJ, Callahan A, Godin JJ, Sherratt TN. 2009. Behaviourally mediated crypsis in two nocturnal moths with contrasting appearance. Phil. Trans. R. Soc. B 364, 503–510 10.1098/rstb.2008.0215 (doi:10.1098/rstb.2008.0215) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cuthill IC, Stevens M, Windsor AMM, Walker HJ. 2006. The effects of pattern symmetry on detection of disruptive and background matching coloration. Behav. Ecol. 17, 828–832 10.1093/beheco/arl015 (doi:10.1093/beheco/arl015) [DOI] [Google Scholar]
- 6.Thayer GH. 1909. Concealing-coloration in the animal kingdom: an exposition of the laws of disguise through color and pattern: being a summary of Abbott H. Thayer's discoveries. New York, NY: Macmillan [Google Scholar]
- 7.Cott HB. 1940. Adaptive coloration in animals. London, UK: Methuen & Co. Ltd [Google Scholar]
- 8.Cuthill IC, Stevens M, Sheppard J, Maddocks T, Párraga CA, Troscianko TS. 2005. Disruptive coloration and background pattern matching. Nature 434, 72–74 10.1038/nature03312 (doi:10.1038/nature03312) [DOI] [PubMed] [Google Scholar]
- 9.Fraser S, Callahan A, Klassen D, Sherratt TN. 2007. Empirical tests of the role of disruptive coloration in reducing detectability. Proc. R. Soc. B 274, 1325–1331 10.1098/rspb.2007.0153 (doi:10.1098/rspb.2007.0153) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Stevens M, Cuthill IC. 2006. Disruptive coloration, crypsis and edge detection in early visual processing. Proc. R. Soc. B 273, 2141–2147 10.1098/rspb.2006.3556 (doi:10.1098/rspb.2006.3556) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Stevens M, Winney IS, Cantor A, Graham J. 2009. Outline and surface disruption in animal camouflage. Proc. R. Soc. B 276, 781–786 10.1098/rspb.2008.1450 (doi:10.1098/rspb.2008.1450) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dimitrova M, Merilaita S. 2009. Prey concealment: visual background complexity and prey contrast distribution. Behav. Ecol. 21, 176–181 10.1093/beheco/arp174 (doi:10.1093/beheco/arp174) [DOI] [Google Scholar]
- 13.Todd PA. 2009. Testing for camouflage using virtual prey and human ‘predators’. J. Biol. Educ. 43, 81–84 10.1080/00219266.2009.9656156 (doi:10.1080/00219266.2009.9656156) [DOI] [Google Scholar]
- 14.Rushton SK, Bradshaw MF, Warren PA. 2007. The pop out of scene-relative object movement against retinal motion due to self-movement. Cognition 105, 237–245 10.1016/j.cognition.2006.09.004 (doi:10.1016/j.cognition.2006.09.004) [DOI] [PubMed] [Google Scholar]
- 15.Regan D, Beverly KI. 1984. Figure-ground segregation by motion contrast and by luminance contrast. J. Opt. Soc. Am. A 1, 434–442 10.1364/JOSAA.1.000433 (doi:10.1364/JOSAA.1.000433) [DOI] [PubMed] [Google Scholar]
- 16.Stevens M, Searle WT, Seymour JE, Marshall KL, Ruxton GD. 2011. Motion dazzle and camouflage as distinct anti-predator defenses. BMC Biol. 9, 81. 10.1186/1741-7007-9-81 (doi:10.1186/1741-7007-9-81) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Krause J, Ruxton GD. 2002. Living in groups. New York, NY: Oxford University Press [Google Scholar]
- 18.Tosh CR, Krause J, Ruxton GD. 2009. Basic features, conjunctive searches, and the confusion effect in predator–prey interactions. Behav. Ecol. Sociobiol. 63, 473–475 10.1007/s00265-008-0667-4 (doi:10.1007/s00265-008-0667-4) [DOI] [Google Scholar]
- 19.Krakauer DC. 1995. Groups confuse predators by exploiting perceptual bottlenecks: a connectionist model of the confusion effect. Behav. Ecol. Sociobiol. 36, 421–429 10.1007/BF00177338 (doi:10.1007/BF00177338) [DOI] [Google Scholar]
- 20.Ruxton GD, Jackson AL, Tosh CR. 2007. Confusion of predators does not rely on specialist coordinated behaviour. Behav. Ecol. 18, 590–596 10.1093/beheco/arm009 (doi:10.1093/beheco/arm009) [DOI] [Google Scholar]
- 21.Duncan J, Humphreys G. 1989. Visual search and stimulus similarity. Psychol. Rev. 96, 433–458 10.1037/0033-295X.96.3.433 (doi:10.1037/0033-295X.96.3.433) [DOI] [PubMed] [Google Scholar]
- 22.Prinzmetal W, Banks W. 1977. Good continuation affects visual detection. Percept. Psychophys. 21, 389–395 10.3758/BF03199491 (doi:10.3758/BF03199491) [DOI] [Google Scholar]
- 23.McLeod P, Driver J, Crisp J. 1988. Visual search for a conjunction of movement and form is parallel. Nature 332, 154–155 10.1038/332154a0 (doi:10.1038/332154a0) [DOI] [PubMed] [Google Scholar]
- 24.Hulleman J. 2010. Inhibitory tagging in visual search: only in difficult search are items tagged individually. Vis. Res. 50, 2069–2079 10.1016/j.visres.2010.07.017 (doi:10.1016/j.visres.2010.07.017) [DOI] [PubMed] [Google Scholar]
- 25.Von Mühlenen A, Müller HJ. 1999. Visual search for motion-form conjunctions: selective attention to movement direction. J. Gen. Psychol. 126, 289–317 10.1080/00221309909595368 (doi:10.1080/00221309909595368) [DOI] [PubMed] [Google Scholar]
- 26.Merilaita S, Lind J. 2005. Background-matching and disruptive coloration, and the evolution of cryptic coloration. Proc. R. Soc. B 272, 665–670 10.1098/rspb.2004.3000 (doi:10.1098/rspb.2004.3000) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Marr D, Hildreth E. 1980. Theory of edge detection. Proc. R. Soc. Lond. B 207, 187–217 10.1098/rspb.1980.0020 (doi:10.1098/rspb.1980.0020) [DOI] [PubMed] [Google Scholar]
- 28.Yearsley J. 2004. Generate AR1 spatial data. See http://www.mathworks.com/matlabcentral/fileexchange/5099-generate-ar1-spatial-data
- 29.SPSS Inc 2010. PASW Advanced Statistics 18. Chicago, IL: SPSS Inc [Google Scholar]
- 30.Newark T, Miller J. 2007. Camouflage. London, UK: Thames & Hudson [Google Scholar]
- 31.Skelhorn J, Rowland HM, Speed MP, Ruxton GD. 2010. Masquerade: camouflage without crypsis. Science 327, 51–51 10.1126/science.1181931 (doi:10.1126/science.1181931) [DOI] [PubMed] [Google Scholar]
- 32.Fleishman LJ. 1985. Cryptic movement in the vine snake Oxybelis aeneus. Copeia 1, 242–245 10.2307/1444822 (doi:10.2307/1444822) [DOI] [Google Scholar]
- 33.Landeau L, Terbourgh J. 1986. Oddity and the confusion effect in predation. Anim. Behav. 34, 1372–1380 10.1016/S0003-3472(86)80208-1 (doi:10.1016/S0003-3472(86)80208-1) [DOI] [Google Scholar]
- 34.Schradin C. 2000. Confusion effect in a reptilian and a primate predator. Ethology 106, 691–700 10.1046/j.1439-0310.2000.00582 (doi:10.1046/j.1439-0310.2000.00582) [DOI] [Google Scholar]