

Abstract
It has been widely accepted that the growth-related phytohormone auxin is the endogenous signal that initiates bending movements of plant organs. In 1875, Charles Darwin described how the bending movement of leaves in carnivorous sundew species formed an ‘outer stomach’ that allowed the plants to enclose and digest captured insect prey. About 100 years later, auxin was suggested to be the factor responsible for this movement. We report that prey capture induces both leaf bending and the accumulation of defence-related jasmonate phytohormones. In Drosera capensis fed with fruitflies, within 3 h after prey capture and simultaneous with leaf movement, we detected an increase in jasmonic acid and its isoleucine conjugate. This accumulation was spatially restricted to the bending segment of the leaves. The application of jasmonates alone was sufficient to trigger leaf bending. Only living fruitflies or the body fluids of crushed fruitflies induced leaf curvature; neither dead flies nor mechanical treatment had any effect. Our findings strongly suggest that the formation of the ‘outer stomach’ in Drosera is a chemonastic movement that is triggered by accumulation of endogenous jasmonates. These results suggest that in carnivorous sundew plants the jasmonate cascade might have been adapted to facilitate carnivory rather than to defend against herbivores.
Keywords: carnivorous plants, Drosera, leaf movement, auxin, jasmonate, chemonasty
1. Introduction
Carnivorous plants of the worldwide genus Drosera (sundew) catch their prey by using adhesive traps baited with sticky polysaccharide mucilage [1,2]. The glue is produced by stalked glands (the so-called tentacles) on the leaves' surface. The glands also secrete enzymes to digest captured prey and to absorb the resulting nutrients. Once the prey (mainly small insects) comes in contact with the sticky glands, the lateral glandular tentacles bend towards the prey, which can no longer escape, and move it to the centre of the leaf [1,2]. Mechanical and chemical stimuli from the prey induce electric potentials in the glands that move towards the base of the tentacles in the epidermal layer of the leaf [3]. After this first, fast tentacle movement, a second, slower movement of the whole leaf starts to enclose the caught prey [1]. As a consequence, the prey comes in contact with numerous digestive glands. In Drosera capensis, the cape sundew that is indigenous to South Africa, this curvature of the adaxial leaf surface is highly pronounced: because the leaf almost completely encloses the prey, an ‘outer stomach’ is formed [4] (figure 1). Although Darwin [1] described the leaf-bending phenomenon, real prey-induced endogenous plant signals that direct this movement have so far not been identified.
Figure 1.

Drosera capensis leaf 12 h after insect prey capture. Scale bar, 1 cm.
Early studies of the endogenous plant signals in Drosera leaf movement suggested a role for the phytohormone auxin, indole-3-acetic acid (IAA) [4,5]. Based on studies with the auxin transport inhibitor 2,3,5-triiodobenzoic acid (TIBA), the authors concluded that captured prey induced an IAA flux from the leaf tip downstream to the bending point. However, radio-labelled auxin, 3-indolyl-(2–14C)-acetic acid, provided externally to the tip of a D. capensis leaf, was not transported basipetally to the site of the prey [4]. In addition, the external application of IAA induced only weak bending [5]. In a different experiment, an anti-IAA antiserum-based enzyme immunoassay suggested the presence of increased auxin at the site of the prey [4]. Unfortunately, a control for the presence of cross-reacting substances in the provided ‘prey’ (a piece of cheese) was missing. Moreover, false-positive results by the anti-IAA antiserum owing to cross-reactivity with structurally non-related compounds could not be ruled out [6]. Thus, whether auxin represents the signal responsible for outer stomach formation remained unknown.
In this study, we provided Drosophila melanogaster as prey and subsequently analysed the induced phytohormone level in the Drosera leaves upon prey capture. Leaf bending was a chemonastic movement initiated by a chemical signal derived from the insect, not by simple mechanical treatments. Exogenously applied phytohormones were used to trigger leaf bending without prey. We correlate the leaf bending with the presence of jasmonate, and suggest that the jasmonate signalling pathway was involved when plants developed carnivory.
2. Results and discussion
Living fruitflies (D. melanogaster) were provided as prey in order to observe the kinetics of the bending reaction in D. capensis. Within a short time after prey capture, the leaves started to bend at the exact site where the flies were placed; after 3 h, bending was highly pronounced (figure 2a,b). Analysis of the treated leaves showed that IAA levels did not change, in contrast to the significant increase in oxylipin phytohormones, namely jasmonic acid (JA) and its physiologically most active form, the isoleucine conjugate (JA-Ile; figure 2c). Jasmonates are phytohormones that regulate plant developmental processes as well as defences [7–9]. To further investigate whether the accumulation of jasmonates was restricted to the bending segment only, leaves challenged by D. melanogaster were cut into three parts (non-curled upper part, curled middle part and non-curled lower part; figure 3a) and analysed. Compared with the control, after 3 h, only the curled tissue of the leaves showed the accumulation of JA and JA-Ile (1.67 versus 241.5 ng gFW−1 and 0.1 versus 42.0 ng gFW−1, respectively). Moreover, the increase in jasmonate levels was drastic, whereas IAA levels (6.13 versus 12.6 ng gFW−1) increased only twofold (figure 3b). The results strongly suggested that jasmonates at the very least contributed to the leaves' movement. Therefore, an external application of these phytohormones should be sufficient to induce leaf bending without the presence of prey. Indeed, as expected, JA-Ile induced bending at the site of application (figure 4). Moreover, the biosynthetic precursors of jasmonates—12-oxo-phytodienoic acid (OPDA), JA and its synthetic mimic, coronalon [10]—were able to induce the same reaction (see the electronic supplementary material, figure S1a). The leaf curvature reaction was concentration-dependent (see the electronic supplementary material, figure S2). Occasionally, the leaf became completely rolled, which sometimes can also be seen upon prey capture. Interestingly, the bending reaction was observed only in the adaxial part of the leaf, regardless of whether the stimulus was applied adaxially or abaxially, indicating that leaf curvature in D. capensis is a nastic movement.
Figure 2.

Kinetics of leaf bending in Drosera capensis. (a) Control leaves and (b) leaves after capture of two living Drosophila melanogaster. Arrows indicate the position of the flies. Scale bars, 1 cm. (c) Phytohormone concentrations (ng per g fresh weight; 3–5 leaves were collected for one single measurement) in D. capensis leaves treated with (white circle) and without (black circle) D. melanogaster after 45, 90 and 180 min (mean ± s.d.; n = 5). Asterisks indicate significant differences between treatments; n.s., not significant (ANOVA, SNK-test, p < 0.001). Five leaves were collected for each single time point. All phytohormones were analysed in parallel from the same samples. IAA, indole-3-acetic acid; JA, jasmonic acid; JA-Ile, jasmonic acid–isoleucine conjugate. (Online version in colour.)
Figure 3.
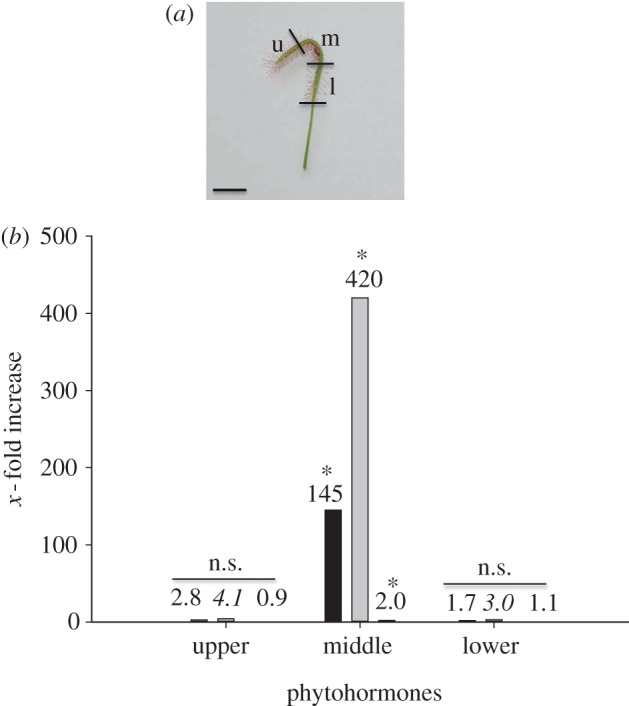
(a) Definition of Drosera capensis leaf segments observed 3 h after fruitfly capture; u, upper; m, middle; l, lower. (b) Phytohormone analyses of the various segments (9–12 leaves were collected for each single measurement); indicated is the increase in the respective phytohormone 3 h after capture of two Drosophila melanogaster compared with the control. From left to right: jasmonic acid (black bar), jasmonic acid–isoleucine conjugate (grey bar), indole-3-acetic acid (white bar); exact values are given above the bars (n = 3). Asterisks indicate significant differences between treatments; n.s., not significant (ANOVA, SNK-test, p < 0.001 for JA and JA-Ile, p < 0.003 for IAA).
Figure 4.

Three Drosera capensis leaves treated with three 10 µl-drops JA-Ile (jasmonic acid–isoleucine conjugate, 100 µM) at the same time in the centre of the adaxial site of the intact leaf. The leaves shown represent three different plants. Photograph taken after 12 h, immediately after cutting off the leaves. Scale bar, 1 cm.
In contrast to jasmonates, IAA did not induce such leaf-bending reaction either at low (1 µM) or high concentrations (1 mM; electronic supplementary material, figure S1b), supporting our results that auxin was not the only determining factor for the leaf movement. However, the role of auxin in the orchestration of plant movements is well established [11]. Thus, we speculated that not a locally increased IAA concentration but a local IAA gradient along the adaxial/abaxial axis was involved. Such gradient might have been induced by jasmonates. Because it was impossible to determine phytohormone concentrations in both the upper and lower epidermis independently, TIBA was used to inhibit auxin transport. Therefore, the curvature of the leaves was determined in degrees [5]. In control plants, coronalon induced leaf bending (119.3°±72.9; n = 14), whereas in the presence of TIBA the coronalon-initiated bending (83.6°±66.7; n = 11) was as often induced as inhibited, resulting in non-significant differences (p = 0.22; Mann–Whitney rank sum U-test). This inconsistent result can be explained by the fact that TIBA needs to interact directly with its target proteins localized in leaf lamina cells to execute its action; but in contrast to the tentacles, some exogenously applied compounds hardly penetrate the cuticle and enter the epidermal or mesophyll cells, as shown for the fluorescent dyes Calcofluor White and DAPI (see the electronic supplementary material, figure S3), an observation that was also mentioned recently [12].
Jasmonates are induced by various stress factors such as insect herbivory or tissue wounding [9,13]. In D. capensis, there is no tissue wounding involved because the mouthparts of D. melanogaster are spongy and unable to injure the plant. Thus, the question remained: what was responsible for the induction of jasmonate biosynthesis and the subsequent leaf bending—contact with the tentacles and the leaf surface, or chemical stimuli? This issue has never been investigated systematically. When provided as prey, living D. melanogaster induced movement in leaves, unlike dead flies or small stones (figure 5). These results suggested that the movement of flies struggling to escape the adhesive trap elicited the response. However, the stimulation of the tentacles and the leaves with a small brush could not induce leaf bending, which argues against the response being a thigmonastic reaction. When we observed that the presence of dead, crushed flies caused a strong leaf-bending reaction (figure 5), we concluded that certain insect-derived compounds, recognized as chemical signals by the plant, were inducing the response.
Figure 5.

Drosera capensis leaves treated with different modes of prey: two living individuals of Drosophila melanogaster (D. m.); two dead individuals of D. melanogaster (D. m. †); a small stone with the size of two fruitflies; challenge with a brush for 15 min; freshly crushed D. melanogaster (D. m., crushed). The leaves shown represent three different plants. Each single photo was taken after 12 h, immediately after cutting off the leaves. Arrows indicate the position of the prey. Scale bars, 1 cm. (Online version in colour.)
However, such chemical cues might be released as well during the effort by captured, living flies to escape but not by dead, intact flies. Darwin [1] observed the leaf edge of Drosera rotundifolia bending 24 h after challenge with dead prey, and dead fruitflies induced leaf curvature in D. capensis after 1 day [14]. In fact, this does not contradict our hypothesis because once the digestion of dead insect prey has started, chemical signals are released and available, although delayed by several hours. Thus, we argue that prey-derived material serves as a signal for the plant to generate jasmonates and, eventually, to induce the chemonastic movement of D. capensis leaves to form the ‘outer stomach’. Up to now, only one other example is known where jasmonates are involved in plant movements, namely tendril coiling in Bryonia dioica [6,15]; but in contrast to Drosera, tendril coiling is a thigmonastic reaction induced only by touch as external stimulus. However, in B. dioica also exogenously applied auxin can induce tendril coiling [15].
The exact nature of the chemical signals in Drosera–insect–prey interaction remains to be elucidated. The signals could simply be any kind of nutrient that is valuable for the plant. However, other types of stress besides herbivory and wounding can induce jasmonate accumulation in plants; for example, pathogen-derived elicitors that eventually induce plant defences in cell suspension cultures [16,17]. Similarly, in some plant–insect interactions, fatty acid–glutamine conjugates from feeding herbivores have been shown to actively induce jasmonates and, subsequently, direct and indirect defences [18,19]. Conceivably, the molecule that is recognized by D. capensis and initiates the formation of the ‘stomach’ could be any defence-activating signalling compound as defined by herbivore-associated molecular patterns [20]. Specifically, in carnivorous sundew, the jasmonate cascade, which is typically involved in plant defence against insect herbivores, might be used to facilitate prey capture.
Recently, in Dionaea muscipula, the venus flytrap, it was shown that application of the Pseudomonas syringae-derived phytotoxin coronatine, a structural mimic of jasmonates, induced leaf lobes to close, thereby forming the ‘outer stomach’ of this carnivorous plant [21]. However, this movement took several hours, whereas the natural closing induced by prey is accomplished within a second [22,23]. Why in those experiments only the phytotoxin was active but jasmonates failed remained an unsolved question. Moreover, even upon prey capture, only after several hours could an increase in OPDA be measured, and then less than twofold, but no increase in JA and JA-Ile was detected. These results prevent us from concluding that jasmonates have a role in the prey capture mechanism in Dionaea as well. In carnivorous plants, jasmonates might be involved in chemonastic reactions rather than in thigmonastic movements.
As already known, jasmonates are part of a network of different phytohormones, including auxin, that work together in order to exploit, coordinate and fine-tune the defensive capacity of plants upon challenges in various biotic interactions [24,25]. Thus, it is tempting to speculate that also in the leaf-bending reaction of Drosera a network of phytohormones, consisting of at least auxin and jasmonate, is involved. Further studies will analyse how phytohormones cross-talk to initiate leaf movement in Droseraceae and other plants. Insights into the evolution of carnivory may even shift our understanding of jasmonates from a role in defence to a role in plant nutrition. In addition, our findings suggest that jasmonates, in addition to auxin, might be involved in growth-dependent plant movements such as in phototropism or gravitropism [11]. The availability of highly sensitive analytical methods for phytohormone analyses will allow a deeper understanding of the roles and networking of various phytohormones.
3. Material and methods
(a). Plant material and treatment
Drosera capensis L. plants were grown in the greenhouse at 22–24°C (day) and 20–22°C (night), 60–70 per cent humidity, and light from 6.00 to 22.00. Plants were kept wet the entire time with collected rain water. All treatments were performed with the intact leaves of the plants for the time indicated in the particular experiments. Applications were done in the centre of adult leaves.
For treatment with Drosophila melanogaster (reared on conventional cornmeal agar), either two living or two dead flies (frozen at −20°C) were placed on the same leaf site with a distance of approximately 5 mm between them. Living flies were cooled on ice for 30 min before being used. In the signal compound treatment, three 10 µl drops (100 µM, if not indicated otherwise) or water for the control were applied side by side. For inhibitor (250 µM) experiments, 10 µl drops were applied 2 h before jasmonates (100 µM) were added at the same site for 6 h. Fluorescent dye (Calcofluor White M2R 1 µg ml−1 for cell wall staining; DAPI 10 µg ml−1 for DNA staining) uptake was documented by an Olympus BX 41 reflecting fluorescence microscope equipped with a colour view soft imaging system.
JA was synthesized from commercially available methyl-JA (Sigma) by saponification; JA-Ile, coronalon and OPDA were synthesized according to [26–28]; IAA, TIBA and fluorescent dyes were purchased (Calcofluor White: Fluka, DAPI: Sigma), respectively. All experiments were performed independently at least three times.
(b). Quantification of jasmonic acid, JA-Ile, OPDA and indole acetic acid in sundew leaves
Five leaves were collected for each single measurement of phytohormones. Plant material of individually treated leaves was harvested, collected, weighed and frozen with liquid nitrogen, and samples were kept at −80°C until use. For phytohormone analysis, finely ground material (100 mg) was extracted with 1.5 ml of methanol containing 60 ng of 9,10-D2–9,10-dihydrojasmonic acid, 60 ng of D6-abscisic acid (Santa Cruz Biotechnology) and 15 ng of JA-[13C6]isoleucine conjugate as internal standards. JA-[13C6]isoleucine conjugate was synthesized as described [28] using [13C6]Ile (Sigma). The homogenate was mixed for 30 min and centrifuged at 13 000 r.p.m. for 20 min at 4°C. After the supernatant was collected, the homogenate was re-extracted with 500 µl methanol, mixed and centrifuged, and supernatants were pooled. The combined extracts were evaporated in a speed-vac at 30°C and re-dissolved in 500 µl methanol. Chromatography was performed on an Agilent 1200 HPLC system (Agilent Technologies). Separation was achieved on a Zorbax Eclipse XDB-C18 column (50 × 4.6 mm, 1.8 µm, Agilent). Formic acid (0.05%) in water and acetonitrile was used as mobile phases A and B, respectively. The elution profile was as follows: 0–0.5 min, 5 per cent B; 0.5–9.5 min, 5–42 per cent B; 9.5–9.51 min, 42–100 per cent B; 9.51–12 min, 100 per cent B; and 12.1–15 min, 5 per cent B. The mobile phase flow rate was 1.1 ml min−1. The column temperature was maintained at 25°C, injection volume was 2 µl. An API 5000 tandem mass spectrometer (Applied Biosystems) equipped with a Turbospray ion source was operated in the negative ionization mode. The instrument parameters were optimized by infusion experiments with pure standards, where available. The ion spray voltage was maintained at −4500 eV. The turbo gas temperature was set at 700°C. Nebulizing gas was set at 60 psi, curtain gas at 25 psi, heating gas at 60 psi and collision gas at 7 psi. Multiple reaction monitoring was used to monitor the analyte parent ion; m/z 209.1 → 59.0 (collision energy (CE) −24 V; declustering potential DP −35 V) for JA; m/z 213.1 → 56.0 (CE −24 V; DP −35 V) for 9,10-D2–9,10-dihydrojasmonic acid; m/z 322.2 → 130.1 (CE −30 V; DP −50 V) for the JA-isoleucine conjugate; m/z 328.2 → 136.1 (CE −30 V; DP −50 V) for the JA-[13C6]isoleucine conjugate; m/z 269 → 159.2 (CE −22 V; DP −35 V) for D6-abscisic acid; and m/z 173.85 → 129.9 (CE −14 V; DP −25 V) for IAA. Both Q1 and Q3 quadrupoles were maintained at unit resolution. Analyst v. 1.5 software (Applied Biosystems) was used for data acquisition and processing. Linearity in ionization efficiencies was verified by analysing dilution series of standard mixtures. Phytohormones were quantified relative to the signal of their corresponding internal standard. The peak of the endogenous bioactive form of JA-Ile, (+)-7-iso-jasmonoyl-l-isoleucine [29], was used for JA-Ile quantification. For quantification of IAA, D6-abscisic acid was used as the internal standard (because it elutes in a 0.4 min retention time window from IAA) applying an experimentally determined response factor of 3.4.
Acknowledgements
The work was supported by the Max Planck Society. Special thanks to W. Boland and J. Gershenzon for continuous support, I. Lichtscheidl for valuable advice with the fluorescent staining, and A. Lehr, C. Lembke, B. Arnold, T. Krügel, S. Trautheim, J. Vadassery and A. Bellaire for help in the laboratory, growing plants, providing fruitflies and stimulating discussions. We thank E. Wheeler, Boston, for editorial assistance.
References
- 1.Darwin CR. 1875. Insectivorous plants. London, UK: John Murray [Google Scholar]
- 2.Adlassnig W, Lendl T, Peroutka M, Lang I. 2010. Deadly glue-adhesive traps of carnivorous plants. In Biological adhesive systems (eds von Byern J, Grunwald I.), pp. 15–28 Heidelberg, Germany: Springer [Google Scholar]
- 3.Williams SE, Pickard BG. 1972. Properties of action potentials in Drosera tentacles. Planta 103, 222–240 10.1007/BF00386845 (doi:10.1007/BF00386845) [DOI] [PubMed] [Google Scholar]
- 4.Bopp M, Weiler EW. 1985. Leaf blade movement of Drosera and auxin distribution. Naturwissenschaften 72, 434. 10.1007/BF00404889 (doi:10.1007/BF00404889) [DOI] [Google Scholar]
- 5.Bopp M, Weber I. 1981. Hormonal regulation of leaf blade movement of Drosera capensis. Physiol. Plant 53, 491–496 10.1111/j.1399-3054.1981.tb02738.x (doi:10.1111/j.1399-3054.1981.tb02738.x) [DOI] [Google Scholar]
- 6.Falkenstein E, Groth B, Mithöfer A, Weiler EW. 1991. Methyljasmonate and α-linolenic acid are potent inducers of tendril coiling. Planta 185, 316–322 10.1007/BF00201050 (doi:10.1007/BF00201050) [DOI] [PubMed] [Google Scholar]
- 7.Sembdner G, Parthier B. 1993. The biochemistry and the physiological and molecular actions of jasmonates. Annu. Rev. Plant Physiol. Plant Mol. Biol. 44, 569–589 10.1146/annurev.pp.44.060193.003033 (doi:10.1146/annurev.pp.44.060193.003033) [DOI] [Google Scholar]
- 8.Creelman RA, Mullet JE. 1997. Biosynthesis and action of jasmonates in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 48, 355–381 10.1146/annurev.arplant.48.1.355 (doi:10.1146/annurev.arplant.48.1.355) [DOI] [PubMed] [Google Scholar]
- 9.Wasternack C. 2007. Jasmonates: an update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 100, 681–697 10.1093/aob/mcm079 (doi:10.1093/aob/mcm079) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mithöfer A, Maitrejean M, Boland W. 2005. Structural and biological diversity of cyclic octadecanoids, jasmonates, and mimetics. J. Plant Growth Regul. 23, 170–178 [Google Scholar]
- 11.Santner A, Calderon-Villalobos LIA, Estelle M. 2009. Plant hormones are versatile chemical regulators of plant growth. Nat. Chem. Biol. 5, 301–307 10.1038/nchembio.165 (doi:10.1038/nchembio.165) [DOI] [PubMed] [Google Scholar]
- 12.Adlassnig W, Koller-Peroutka M, Bauer S, Koshkin E, Lendl T, Lichtscheidl I. 2012. Endocytotic uptake of nutrients in carnivorous plants. Plant J. 71, 303–313 10.1111/j.1365-313X.2012.04997.x (doi:10.1111/j.1365-313X.2012.04997.x) [DOI] [PubMed] [Google Scholar]
- 13.Glauser G, Grata E, Dubugnon L, Rudaz S, Farmer EE, Wolfender JL. 2008. Spatial and temporal dynamics of jasmonate synthesis and accumulation in Arabidopsis in response to wounding. J. Biol. Chem. 283, 16 400–16 407 10.1074/jbc.M801760200 (doi:10.1074/jbc.M801760200) [DOI] [PubMed] [Google Scholar]
- 14.Adamec L. 2002. Leaf absorption of mineral nutrients in carnivorous plants stimulates root nutrient uptake. New Phytol. 155, 89–100 10.1046/j.1469-8137.2002.00441.x (doi:10.1046/j.1469-8137.2002.00441.x) [DOI] [PubMed] [Google Scholar]
- 15.Weiler EW, Albrecht T, Groth B, Zia ZQ, Luxem S, Liss H, Andert L, Spengler P. 1993. Evidence for the involvement of jasmonates and their octadecanoid precursors in the tendril coiling response of Bryonia dioica. Phytochemistry 32, 591–600 10.1016/S0031-9422(00)95142-2 (doi:10.1016/S0031-9422(00)95142-2) [DOI] [Google Scholar]
- 16.Gundlach H, Müller MJ, Kutchan TM, Zenk MH. 1992. Jasmonic acid is a signal transducer in elicitor-induced plant cell cultures. Proc. Natl Acad. Sci. USA 89, 2389–2393 10.1073/pnas.89.6.2389 (doi:10.1073/pnas.89.6.2389) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nojiri H, Sugimori M, Yamane H, Nishimura Y, Yamada A, Shibuya N, Kodama O, Murofushi N, Omori T. 1996. Involvement of jasmonic acid in elicitor-induced phytoalexin production in suspension-cultured rice cells. Plant Physiol. 110, 387–392 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Halitschke R, Schittko U, Pohnert G, Boland W, Baldwin IT. 2001. Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. III. Fatty acid-amino acid conjugates in herbivore oral secretions are necessary and sufficient for herbivore-specific plant responses. Plant Physiol. 125, 711–717 10.1104/pp.125.2.711 (doi:10.1104/pp.125.2.711) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Schmelz EA, Alborn HT, Tumlinson JH. 2003. Synergistic interactions between volicitin, jasmonic acid and ethylene mediate insect-induced volatile emission in Zea mays. Physiol. Plant 117, 403–412 10.1034/j.1399-3054.2003.00054.x (doi:10.1034/j.1399-3054.2003.00054.x) [DOI] [PubMed] [Google Scholar]
- 20.Mithöfer A, Boland W. 2008. Recognition of herbivory-associated molecular patterns. Plant Physiol. 146, 825–831 10.1104/pp.107.113118 (doi:10.1104/pp.107.113118) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Escalante-Perez M, Krol E, Stange A, Geiger D, Al-Rasheid KAS, Hause B, Neher E, Hedrich R. 2011. A special pair of phytohormones controls excitability, slow closure, and external stomach formation in the Venus flytrap. Proc. Natl Acad. Sci. USA 108, 15 492–15 497 10.1073/pnas.1112535108 (doi:10.1073/pnas.1112535108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Burdon-Sanderson J. 1874. Venus fly-trap (Dionaea muscipula). Nature 10, 105–107 10.1038/010105c0 (doi:10.1038/010105c0) [DOI] [Google Scholar]
- 23.Forterre Y, Skotheim JM, Dumais J, Mahadevan L. 2005. How the Venus flytrap snaps? Nature 433, 421–425 10.1038/nature03185 (doi:10.1038/nature03185) [DOI] [PubMed] [Google Scholar]
- 24.Pieterse CMJ, Leon-Reyes A, Van der Ent S, Van Wees SCM. 2009. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 5, 308–316 10.1038/nchembio.164 (doi:10.1038/nchembio.164) [DOI] [PubMed] [Google Scholar]
- 25.Ballaré CL. 2011. Jasmonate-induced defenses: a tale of intelligence, collaborators and rascals. Trends Plant Sci. 16, 249–257 10.1016/j.tplants.2010.12.001 (doi:10.1016/j.tplants.2010.12.001) [DOI] [PubMed] [Google Scholar]
- 26.Krumm T, Bandemer K, Boland W. 1995. Induction of volatile biosynthesis in the Lima bean (Phaseolus lunatus) by leucine and isoleucine conjugates of 1-oxo- and 1-hydroxyindan-4-carboxylic acid: evidence for amino acid conjugates of jasmonic acid as intermediates in the octadecanoid signalling pathway. FEBS Lett. 377, 523–529 10.1016/0014-5793(95)01398-9 (doi:10.1016/0014-5793(95)01398-9) [DOI] [PubMed] [Google Scholar]
- 27.Schüler G, Görls H, Boland W. 2001. 6-Substituted indanoyl isoleucine conjugates mimic the biological activity of coronatine: elicitors of plant secondary metabolism V. Eur. J. Org. Chem. 9, 1663–1668 (doi:10.1002/1099-0690(200105)2001:9<1663::AID-EJOC1663>3.0.CO;2-I) [DOI] [Google Scholar]
- 28.Kramell R, Schmidt J, Schneider G, Sembdner G, Schreiber K. 1988. Synthesis of N-(jasmonoyl)amino acid conjugates. Tetrahedron 44, 5791–5807 10.1016/S0040-4020(01)81437-X (doi:10.1016/S0040-4020(01)81437-X) [DOI] [Google Scholar]
- 29.Fonseca S, Chini A, Hamberg M, Adie B, Porzel A, Kramell R, Miersch O, Wasternack C, Solano R. 2009. (+)-7-iso-Jasmonoyl-l-isoleucine is the endogenous bioactive jasmonate. Nat. Chem. Biol. 5, 344–350 10.1038/nchembio.161 (doi:10.1038/nchembio.161) [DOI] [PubMed] [Google Scholar]