

Abstract
During Drosophila embryogenesis the process of dorsal closure (DC) results in continuity of the embryonic epidermis, and DC is well recognized as a model system for the analysis of epithelial morphogenesis as well as wound healing. During DC the flanking lateral epidermal sheets stretch, align, and fuse along the dorsal midline, thereby sealing a hole in the epidermis occupied by an extra-embryonic tissue known as the amnioserosa (AS). Successful DC requires the regulation of cell shape change via actomyosin contractility in both the epidermis and the AS, and this involves bidirectional communication between these two tissues. We previously demonstrated that transcriptional regulation of myosin from the zipper (zip) locus in both the epidermis and the AS involves the expression of Ack family tyrosine kinases in the AS in conjunction with Dpp secreted from the epidermis. A major function of Ack in other species, however, involves the negative regulation of Egfr. We have, therefore, asked what role Egfr might play in the regulation of DC. Our studies demonstrate that Egfr is required to negatively regulate epidermal expression of dpp during DC. Interestingly, we also find that Egfr signaling in the AS is required to repress zip expression in both the AS and the epidermis, and this may be generally restrictive to the progression of morphogenesis in these tissues. Consistent with this theme of restricting morphogenesis, it has previously been shown that programmed cell death of the AS is essential for proper DC, and we show that Egfr signaling also functions to inhibit or delay AS programmed cell death. Finally, we present evidence that Ack regulates zip expression by promoting the endocytosis of Egfr in the AS. We propose that the general role of Egfr signaling during DC is that of a braking mechanism on the overall progression of DC.
Introduction
Dorsal closure (DC) is a developmental event occurring in the Drosophila embryo between stages 11 and 16, commencing immediately after germband retraction (reviewed in [1]). Upon the completion of germband retraction, a large dorsal opening is evident in the epidermis. The amnioserosa (AS), an extra-embryonic tissue composed of a single layer of large flat epithelial cells, spans the opening. The two opposing lateral epidermal flanks elongate in the dorsal-ventral (D-V) axis and move dorsal ward to seal the dorsal hole. The dorsal-most epidermal (DME) cells from one side of the embryo meet their counterpart DME cells at the dorsal midline. The epidermal sealing process occurs in a zipper-like manner, progressing simultaneously from both the anterior and posterior ends of the dorsal opening and finishing at the center of the dorsal midline. During DC, the AS contracts and its cells become more cuboidal in shape; the AS also actively extrudes approximately 10% of its cells with the effect of increasing the rate of DC [2]–[4]. Upon the completion of DC, the entire AS degenerates by programmed cell death [2].
The DME cells experience a range of morphogenetic events during DC. These include elongation in the D-V axis, formation of actin-based membrane extensions, and adhesion with their partners from the other side of the embryo. DC is a popular model system to study cell shape change in epithelial morphogenesis and multiple signaling proteins have been characterized in this context. In addition to signaling pathways and proteins, there are a number of mechanical forces driving DC [4](reviewed in [5]). These include a supracellular actomyosin cable that is assembled at the leading edge (LE) of the DME cells to form a contractile “purse string”. This contractile apparatus constricts the DME cells in the anterior-posterior axis and thus contributes to their stretching in the D–V axis and movement towards the dorsal midline. Actin-based filopodia and lamellipodia also project from the leading edge of the DME cells, and these are thought to contribute to the alignment and adhesion of opposing DME cells as DC concludes [6], [7]. Finally, as was demonstrated by elegant laser micro-dissection experiments, AS constriction not only removes this tissue as an impediment to movement of the epidermis, but also pulls the DME cells dorsal ward [4].
Among the numerous signaling proteins known to regulate DC is Dpp, a member of the transforming growth factor-β superfamily of cytokines. Dpp expression in DME cells is required for morphogenesis of both the amnioserosa and the epidermis during DC. This requirement for Dpp expression in the DME cells is associated, at least in part, with the regulation of the expression of zipper (zip), which encodes non-muscle myosin II heavy chain [8]–[26]. We previously demonstrated that two members of the Ack family of nonreceptor tyrosine kinases, Ack and PR2, co-operate with Dpp to regulate myosin levels in the AS and epidermis during DC. This work led us to propose the existence of a diffusible signal that is generated by the AS and is regulated by Ack and PR2 in conjunction with Dpp [26]. The zip product is required for cell shape change in both the AS and the epidermis; zip expression, which is regulated by the Ack/PR2/Dpp signaling network, may ultimately coordinate the overall progression of DC [27].
There is considerable evidence that a major function of Ack is the negative regulation of Egfr, and this is thought to occur through the regulation of Egfr by endocytosis and/or ubiquitination [28]–[32]. Egfr may, therefore, play a key role in the Ack/PR2/Dpp regulatory pathway during DC. The Egfr pathway is used repeatedly throughout Drosophila development and appears to regulate a myriad of processes including cell proliferation, cell differentiation, apoptosis, cell motility and adhesion (reviewed in [33], [34]). While it has long been recognized that Egfr has multiple roles in regulating morphogenesis, including germband retraction, its role in DC has not been specifically addressed [35], [36].
The results presented here demonstrate that Egfr is required in both the AS and epidermis for proper DC. In addition, we demonstrate that the function of Egfr in the AS involves the transcriptional repression of zip in both the AS and the DME cells, and we suggest that this repression involves the regulation of the same diffusible signal previously proposed to be regulated through Ack and PR2. Consistent with this, we present results suggesting that Ack negatively regulates Egfr in the AS by targeting it for endocytosis. Similar to Ack, we suggest that Egfr regulates zip expression in parallel to Dpp signaling, but we find that Egfr also has a strong negative effect on epidermal Dpp transcription. Finally, we confirm that Egfr signaling has an additional role in the AS as an inhibitor of apoptosis. The various roles identified for Egfr signaling during DC are consistent with negative regulation of morphogenesis, and we propose that Egfr acts as a brake to adjust the rate of closure in response to endocytic regulation.
Results
Egfr is Required for Normal DC
To address the potential function of Egfr in DC we chose three alleles previously shown to disrupt embryogenesis: Egfrf2, a severe loss-of-function allele, Egfr2C82, a moderate loss-of-function allele, and Egfr1F26, a conditional allele [35]–[37]. Two previously uncharacterized embryonic lethal alleles, Egfr1a15 and EgfrH25, were also used in the course of this study [38]. Phenotypic analysis of the latter two alleles indicated that Egfr1a15 is a severe loss-of-function allele (equivalent to Egfrf2) and that EgfrH25is also a strong loss-of-function allele, but is slightly less severe than either Egfrf2or Egfr1a15 (data not shown). Cuticle preparations of embryos heteroallelic for Egfrf2 and Egfr2C82 showed severe defects in morphogenesis. Greater than 95% of embryos exhibited the previously described “faint little ball” phenotype [36], [37], being “curled” up, with the posterior end of the embryo in close proximity to the head, indicating a defect in germband retraction (Fig. 1B). In less severely curled embryos it was possible to observe holes in the dorsal surface that typically extended anteriorly into the head (Fig. 1C). In general, severe Egfr mutants were associated with a terminal phenotype that included severe defects in head development as well as a complete failure in germ band retraction – both of which effectively precluded any analysis of DC. We, therefore, sought approaches that would allow us to observe DC defects in individuals with impaired Egfr function. The first of the two approaches involved temperature shift experiments using the conditional allele Egfr1F26 [35]. Embryos collected at the permissive temperature of 18°C were aged for various periods of time before being shifted to the restrictive temperature of 29°C. Embryos transferred to 29°C at any stage prior to stage 10 showed severe cuticle defects similar to those seen with other strong loss-of-function Egfr alleles (data not shown). Shifting slightly older (approximately stage 11) Egfr1F26 mutant embryos to 29°C, however, resulted in a less severe mutant phenotype comprising a smaller head hole, distinct from other defects in the dorsal surface, and a less severe defect in germband retraction (Fig. 1D). In addition, these embryos consistently displayed creases or “puckers” in the dorsal surface, which together with the mild germband retraction defect, gave them a bowed appearance. Approximately 52% of Egfr1F26 embryos were scored as having this bowed phenotype in a temperature shift experiment where a 2.5-hour collection of embryos from the Egfr1F26 stock was aged at 18°C for 12 hours and then shifted to 29°C (corresponding to late stage 10/stage 11 at the time of the temperature shift). Of these bowed embryos, half exhibited an additional phenotype of a hole or scab in the dorsal surface (Fig. 1D). Few defects were seen in the dorsal surface of Egfr1F26 embryos when they were shifted to 29°C at stage 12 or later (data not shown).
Figure 1. Embryos with either global or local loss of Egfr signaling have defects in epithelial morphogenesis.

(A–F, I, J) Cuticle preparations. Black arrows, posterior spiracles; black arrowheads, puckers in cuticle; white arrowheads, dorsal hole or scab. (G, H, K, K´) Confocal micrographs of embryos during dorsal closure (DC). (A) Dorsolateral view of wild-type embryo showing intact dorsal surface. (B) Egfrf2/Egfr2C82 embryo, selected by absence of GFP balancer chromosome, showing “curled up” phenotype. (C) Egfrf2/Egfr2C82 embryo showing large dorsal hole. (D) Embryo from temperature-sensitive Egfr1F26 stock that had been allowed to develop at 18°C before shifting to restrictive temperature of 29°C at about stage 10/11. Embryo has a bowed appearance characterized by pulling in of tail (marked by posterior spiracles) and puckering of cuticle. There is a small dorsal hole in the cuticle. (E) Embryo in which EgfrDN had been expressed in the epidermis using the 69B-Gal4 driver showing dorsal scab and mild bowing. (F) Embryo in which EgfrDN had been expressed in the epidermis using the ptc-Gal4 driver showing bowed appearance and dorsal hole, similar to the embryo in panel D. (G, H) Still images from Movie S1 showing restricted expression pattern of LE-Gal4 driver, revealed using a UAS-GFP-NLS reporter. Cell outlines were revealed through expression of a Ubi-DEcadherin-GFP transgene. (G) LE-Gal4 is not expressed at beginning of DC. (H) Midway through DC, GFP-NLS is expressed in the first two rows of cells flanking the amnioserosa (AS), visualized as GFP signal in nuclei. (I) Embryo in which EgfrDN had been expressed using the LE-Gal4 driver showing mild bowing and dorsal scab. (J) Embryo in which EgfrDN had been expressed in the AS using the Gal4c381 driver showing small dorsal hole. (K) Anti-phospho-MAPK staining of a wild-type embryo showing strong immunoreactivity in the center of the amnioserosa and lateral epidermis but little staining in dorsal epidermis and cells at periphery of the AS. (K´) Same embryo as in K with phospho-MAPK in red and cell outlines revealed with anti-phosphotyrosine (PY, green). Scale bars: 50 µm (A–J)(K, K´).
A second approach for facilitating the analysis of DC defects in Egfr-deficient embryos, which also permitted characterization of the tissue specificity of Egfr function, involved the inducible expression of a dominant negative version of Egfr, EgfrDN. EgfrDN competes with endogenous Egfr for ligand binding, but lacks the cytoplasmic domain, which contains the tyrosine kinase domain necessary for trans-phosphorylation and receptor activation [39]. As a result, EgfrDN attenuates activation of the Egfr cascade in a cell autonomous manner. We expressed a UAS-EgfrDN transgene in various spatial patterns, starting with general epidermal expression and epidermal stripes using the 69B-Gal4 and ptc-Gal4 drivers, respectively [40], [41]. Both patterns of EgfrDN expression resulted in bowed embryos with occasional dorsal holes or scabs, similar to the temperature shift phenotype (Fig. 1E, F). 100% of embryos in which EgfrDN was expressed with ptc-Gal4 were bowed and 10% had a dorsal hole or scab. The dorsal epidermis plays an important role in DC and we impaired Egfr function in this tissue by expressing EgfrDN using the LE-Gal4 driver, which is active only during DC primarily within a subset of cells in the first two rows of dorsal epidermal cells flanking the amnioserosa (Fig. 1G, H, and Movie S1) [13]. This resulted in a bowed embryo phenotype in about a quarter of EgfrDN-expressing embryos (Fig. 1I). Bowed embryos and dorsal holes were also seen when Egfr activity was blocked in the AS using the AS-specific driver Gal4c381 [42](Fig. 1J). We conclude that the normal progression of DC requires Egfr signaling in both the epidermis and the AS.
A major route for signaling by Egfr is the Raf-MAPK pathway, the activation of which can be detected using anti-phospho-MAPK antibodies [43], [44]. We observed strong phospho-MAPK immunoreactivity in the central AS cells of wild-type embryos, but little staining in cells of the AS periphery or in the dorsal epidermis (Fig. 1 K, K’). This result suggests that either Egfr is not using the MAPK pathway in these cells or that the pathway is under tight negative control. Consistent with Egfr signaling generating phospho-MAPK in the AS, the anti-phospho-MAPK immunoreactivity in the AS was absent in embryos in which EgfrDN was expressed with Gal4c381 (data not shown).
We extended our analysis of Egfr function in DC by live imaging embryos homozygous or heteroallelic for the alleles Egfrf2 [37], Egfr1a15 and EgfrH25 [38] and carrying a Ubi-DEcadherin-GFP transgene to visualize cell outlines [45]. At least four movies were taken for each genotype and very consistent phenotypes were observed. Prior to the initiation of germband retraction, degradation of the AS commenced in Egfr mutant embryos, which in the most severe cases led to a complete and dramatic loss of the tissue (Compare Movie S2 to Movies S3 and S4, and Fig. 2A, B to Fig. 2 C, D and Fig. 2 E, F). In some embryos the AS persisted throughout germband retraction and DC, but had noticeably fewer cells than wild-type (Movie S5 and Fig. 2G, H, M–P). In such embryos germband retraction proceeded to a point and then appeared to reverse, with the posterior end of the embryo moving anteriorly. Accompanying this was a bunching of the epidermis characterized by inappropriate adhesion between the dorsal end of non-adjacent segments on the same side of the embryo producing (arrowhead in Fig. 2H). In addition, AS morphogenesis was abnormal, with the tissue constricting perpendicular to the normal anterior-posterior direction (Compare Movie S5 to Movie S6 and Fig. 2I–L to Fig. 2M–P). Finally, heads of Egfr mutant embryos exhibited a precipitous loss of epidermal integrity during embryogenesis with the brain becoming exposed during DC and pushing toward the posterior end of the embryo (Fig. 2G, H, Movie S5).
Figure 2. Impairment of Egfr signaling affects morphogenesis of the AS and dorsal epidermis.

(A, B) Still images from Movie S2 showing unfolding of the AS as germband retraction proceeds in a Ubi-DEcadherin-GFP-expressing embryo. (C, D) Still images from Movie S3 showing delayed germband retraction and disintegration of AS in Egfrf2 mutant embryo expressing Ubi-DEcadherin-GFP. (E, F) Still images from Movie S4 showing delayed germband retraction and disintegration of AS in Egfrf2/EgfrH25 mutant embryo expressing Ubi-DEcadherin-GFP. (G, H) Still images from Movie S5 showing bowing of Egfr1a15/EgfrH25 mutant, Ubi-DEcadherin-GFP-expressing embryo. Note bunching of segments (arrowhead in panel H). (I–L) Still images from Movie S6 showing morphogenesis of the AS in a Ubi-DEcadherin-GFP-expressing embryo. (M–P) Close-ups of dorsal surface in still images from Movie S5 showing defective AS morphogenesis in Egfr mutant embryo. Note that AS has fewer cells than wild-type and constricts perpendicular to the normal anterior-posterior direction. Note that the posterior end of the embryo moves anteriorly in progression from panel N to panel P as the embryo undergoes bowing. (Q-S) Confocal micrographs of dorsal epidermis of embryos stained with anti-phosphotyrosine. (Q) Wild-type embryo showing uniform shape of DME cells and fairly smooth leading edge. (R) Egfr1a15/EgfrH25 embryo showing considerable variation in shape of DME cells and jagged leading edge. Arrow marks a cluster of very constricted DME cells and arrowhead a cluster of cuboidal DME cells. (S) Embryo in which EgfrDN had been expressed using the LE-Gal4 driver showing bunching of segments. Scale bars: 50 µm (A–H)(I–P)(Q–S).
We also assessed morphogenesis in fixed embryos by staining with an antibody against phosphotyrosine to reveal cell outlines. Heteroallelic Egfr mutant embryos showed highly variable cell shape change of the DME cells compared to wild-type, suggestive of misregulated actomyosin contractility (Fig. 2Q, R). We suspect that this uneven contractility in the dorsal epidermis underlies the bunching of the segments in Egfr mutant embryos, and consistent with this expression of EgfrDN with LE-Gal4 led to segmental bunching (Fig. 2S).
Egfr Negatively Regulates dpp Expression in the Epidermis During DC
The bowed embryo phenotype associated with reductions in Egfr function is similar to the cuticle phenotype of embryos in which Dpp is ectopically expressed in the dorsal epidermis [17], [46]–[48]. Furthermore, crosstalk between Egfr and Dpp/TGFβ signaling in the form of antagonistic or cooperative interactions has been reported for a number of developmental events [49]–[82]. We, therefore, next examined dpp expression in embryos having altered Egfr function. We confirmed a previously published observation that ectopic dpp expression extends ventrally along the segmental grooves of Egfr mutant embryos [35], and saw a similar pattern of dpp expression in embryos in which Egfr signaling was attenuated in the epidermis via UAS-EgfrDN expression using 69B-Gal4 or LE-Gal4 drivers (Fig. 3C–F, arrowheads). The phenocopy of Egfr loss-of-function mutants by UAS-EgfrDN expression was underscored by the fact that widespread epidermal expression of this transgene resulted in a reduction in the separation, from one side of the embryo to the other, between the ventral stripes of dpp expression as previously reported for Egfr and D-raf mutant embryos [35], [83]. This decrease in separation allowed the ventral stripe on the other side of the embryo to be seen in a lateral view (arrows in Fig. 3C, E). Given the many studies indicating communication between the AS and epidermis during DC [10], [13], [25], [26], [84]–[89] and the requirement for Egfr in the AS during DC, we looked at dpp expression in embryos in which EgfrDN had been expressed in the AS using Gal4c381, but found no effect (data not shown).
Figure 3. dpp transcription is repressed by Egfr signaling during DC.

Panels A, C, E and F are digoxigenin in situ hybridizations and panels B, D and H–H´ ´ are FISH, with all embryos at beginning of DC. (A, B) Wild-type embryos showing horizontal dorsal and ventrolateral stripes of dpp expression. The dorsal stripe is dpp expression in the DME cells. (C, D) Egfrf2 embryo (C) and Egfrf2/Egfr2C82 embryo (D) showing ectopic dpp expression ventral to the DME cells (arrowheads). Arrow in (C) shows ventrolateral stripe visible on other side of embryo due to decreased distance between stripes compared to wild-type. (E) Embryo in which EgfrDN had been expressed in the epidermis using the 69B-Gal4 driver showing ectopic dpp expression (arrowhead). Arrow shows ventrolateral stripe visible on other side of embryo. (F) Embryo in which EgfrDN had been expressed using the LE-Gal4 driver showing elevated dpp expression in the dorsal epidermis (arrowhead). (G, H–H´ ´) Increasing EGFR signaling by expression of sSpi (G) or Egfr-EGFP (H–H´ ´) in vertical stripes using the ptc-Gal4 driver causes breaks in the dorsal and ventrolateral dpp stripes. Anti-GFP staining (H´, H´ ´) reveals the expression pattern of Egfr-EGFP. Note that remnants of dpp expression (arrowheads in H–H´ ´) are seen where Egfr-EGFP was not expressed. Scale bar: 50 µm.
To examine the effects of excessive Egfr signaling on dpp expression we used two transgenes, UAS-sSpi and UAS-Egfr-EGFP. UAS-sSpi encodes a secreted, active version of the Egfr ligand, Spitz, which can directly bind to Egfr to activate the Egfr pathway, whereas UAS-Egfr-EGFP encodes a biologically active Egfr tagged with enhanced green fluorescent protein (EGFP) [90], [91]. These transgenes were expressed in stripes in the embryo using ptc-Gal4 and effects on dpp expression assessed by in situ hybridization. For both transgenes, the dpp expression stripes in the dorsal and ventrolateral epidermis became fragmented (Fig. 3G, H). Staining with anti-GFP antibodies revealed that remaining patches of dpp expression were in areas where Egfr-EGFP had not been expressed (Fig. 3H ´, H´ ´). We conclude from our loss- and gain-of-function studies that Egfr signaling negatively regulates dpp expression in the epidermis during DC.
Egfr Negatively Regulates zip Expression in the Epidermis and AS During DC
An important target of Dpp regulation during DC is zip, and we evaluated zip expression in Egfr mutant embryos and embryos with tissue-specific attenuation of Egfr signaling. zip shows two major events of transcriptional upregulation that are relevant to the morphogenetic events during DC: first, a burst of expression occurs in the AS during germband retraction and terminates around the beginning of DC; and, second, upregulation occurs in the DME cells beginning during the germband retraction stage and persists throughout DC [12], [24], [26](Fig. 4A). Egfr mutant embryos showed excessive accumulation of zip transcripts in the DME cells, in addition to some ectopic zip transcription in the epidermis (Fig. 4B). We occasionally found less severely disrupted Egfr mutant embryos where the AS was intact; in these zip transcripts persisted in the AS during DC, in contrast to wild-type embryos where the AS was devoid of transcripts by this stage (compare Fig. 4C with Fig. 4A). Reduction of Egfr function in the epidermis through expression of UAS-EgfrDN using the LE- and 69B-Gal4 drivers also caused excessive epidermal zip expression (data not shown). We previously demonstrated that the tyrosine kinase Ack, a putative negative regulator of Egfr, controls zip levels in the AS [26]; we, therefore, also tested the effect of disrupting Egfr specifically in this tissue by expressing UAS-EgfrDN using the AS-specific Gal4c381 driver. In Gal4c381>UAS-EgfrDN embryos we observed ectopic zip expression throughout the AS during DC as well as elevated zip levels in the head, the latter indicating some degree of cell non-autonomous control of zip by Egfr (Fig. 4D).
Figure 4. zip transcription is repressed by Egfr signaling during DC.

zip FISH on embryos at beginning of DC. (A) Wild-type embryo showing high levels of zip transcription in DME cells and absence of zip expression in the AS. Prior to completion of germband retraction there are high levels of zip in the AS of wild-type embryos (see Fig. 6A). (B) Egfrf2 embryo showing intense zip signal in DME cells and ectopic zip expression (arrowheads). (C) Mildly affected Egfrf2 embryo showing modest retention of zip in AS. (D, E) Embryos in which Egfr signaling had been impaired in the AS by expression of either EgfrDN (D) or RasN17 (E) showing significant retention of zip in AS, modest elevation of zip expression in the DME cells and ectopic zip transcripts in the head. (F) Elevation of Egfr signaling in the AS through expression of Egfr-EGFP causes down-regulation of zip expression in DME cells. Scale bar: 50 µm.
A major downstream effector for Egfr is Ras and expression of a dominant negative Ras transgene, RasN17 [92], in the AS was found to be associated with a similar increase in zip transcript levels (Fig. 4E). Given the robust levels of zip transcripts normally seen in the DME cells, we found it difficult to ascertain if knock down of Egfr in the AS affected zip expression in the DME cells, but we suspect that it caused a modest elevation (Fig. 4D). To determine if excessive Egfr signaling in the AS would have the opposite effect on zip expression, we expressed the Egfr-EGFP transgene with Gal4c381 (Fig. 4F). zip levels in the DME cells were decreased in association with excessive Egfr signaling in the AS and we conclude that Egfr represses zip transcription in a cell non-autonomous manner during DC.
Egfr Inhibits Apoptosis in the AS
The similarity of Egfr mutant embryonic phenotypes to those of the U-shaped group of genes, which are required for maintenance of the AS, has been noted and studies on fixed preparations suggest premature apoptosis in Egfr mutant embryos [35], [93], [94]. Apoptosis of the AS cells contributes to the forces driving DC, and negative regulation of cell death in the AS could be an important component of the participation of Egfr in DC [2], [3]. A negative regulatory role for the Egfr pathway is further supported by the observation that expression of a constitutively active version of Ras, RasV12, in the AS causes the tissue to persist longer than wild-type [95]. Furthermore, Ras appears to negatively regulate apoptosis throughout the embryo, as revealed by acridine orange staining of embryos with global gains or losses of Ras signaling [96]. As described above, AS cells are lost prematurely in Egfr mutant embryos (Fig. 2D, F) suggesting an early onset of apoptosis. To visualize the effects of losses or gains of Egfr signaling on AS apoptosis in live embryos, we used the caspase sensor Apoliner [97], [98]. Apoliner consists of a monomeric red fluorescent protein (RFP) tethered to EGFP by a caspase-sensitive linker [97]. Furthermore, the design of the Apoliner construct includes a transmembrane domain that precedes the RFP component while the EGFP component includes a nuclear localization signal (NLS). As a consequence, the two fluorophores co-localize to membranes in live cells lacking caspase activity, but caspase activation in live cells results in separation of the fluorophores, with Apoliner-RFP remaining at membranes while Apoliner-EGFP is translocated to the nucleus due to its NLS. At the beginning of germband retraction in wild-type embryos there was little nuclear EGFP in the AS, indicating minimal caspase activity (Fig. 5 A–A´ ´), but there was strong nuclear EGFP in the AS of Egfr mutant embryos at the same stage (Fig. 5 B–B´ ´). As DC proceeded nuclear EGFP accumulated in the AS of wild-type embryos (Fig. 5C–C ´´) and this accumulation could be blocked by expression of the baculovirus caspase inhibitor p35 [99] (Fig. 5D–D ´´). To promote Egfr signaling in the AS, we expressed either sSpi or RasV12 and found that, in both cases, cells showed little nuclear EGFP even late in DC, similar to what was seen with p35 expression (Fig. 5E–F ´´). We conclude that Egfr signaling inhibits caspase activation in the AS. If Egfr impedes apoptosis in the AS, then excessive Egfr signaling might be expected to affect AS morphogenesis. A robust increase in Egfr levels in the AS through expression of Egfr-EGFP using the double driver combination Gal4NP3312+ GAL4NP5328 resulted in a failure of the AS to properly complete morphogenesis (compare Fig. 5G to Fig. 5H, I and Movie S7 to Movies S8 and S9). In addition, the AS of these embryos persisted beyond the normal time of AS programmed cell death.
Figure 5. Egfr inhibits apoptosis and morphogenesis in the AS.

(A–F´ ´) Apoliner signals in the AS. Apoliner reporter had been expressed either globally with the tub-Gal4 driver or in the AS using the LP1-Gal4 driver. For each embryo RFP, EGFP signals and merge are shown. On the right side of each panel is a higher power view of AS cells. In the absence of caspase activity, RFP and EGFP co-localize at various membranes and there is little EGFP signal in the nucleus. In the presence of caspase activity, EGFP is cleaved away from RFP and moves into the nucleus. (A–A´ ´) AS of wild-type embryo prior to germband retraction showing co-localization of RFP and EGFP signals and weak EGFP signals in the nucleus. (B–B´ ´) AS of Egfr mutant embryo prior to germband retraction showing strong EGFP signals in the nucleus. (C–C´ ´) AS of wild-type embryo during DC showing strong EGFP signals in the nucleus. (D–D´ ´) AS of p35-expressing embryo during DC showing weak EGFP signals in the nucleus. (E–E´ ´) AS of sSpi-expressing-expressing embryo during DC showing weak EGFP signals in the nucleus. (F–F´ ´) AS of RasV12-expressing embryo during DC showing weak EGFP signals in the nucleus. (G) Still from Movie S7 showing AS of stage 15 wild-type embryo in which GFP had been expressed with the Gal4NP3312 AS driver, showing narrow, tube-like AS. (H, I) Stills from Movies S8 (H) and S9 (I) showing AS of stage 15 embryos in which Egfr-EGFP and GFP-NLS had been expressed with the double driver combination Gal4NP3312+ Gal4NP5328 showing failure of AS morphogenesis. The AS in panel H has failed to narrow throughout while that in panel I has failed to narrow at the anterior end. Scale bars: 50 µm (A–B´ ´); 10 µm (C–I).
Evidence that Ack and Endocytosis Negatively Regulate Egfr Levels in the AS
An important route through which Egfr signaling is down regulated is by clathrin-mediated endocytosis (reviewed in [100]). When imaging Egfr-EGFP in the AS for the apoptosis study, we noticed that in addition to localizing cortically in AS cells, much of the protein appeared to be accumulating in vesicles (Fig. 6F). Given the literature demonstrating that Ack family tyrosine kinases promote down regulation of Egfr by endocytosis and subsequent degradation [29]–[32], we looked for evidence that AS Ack was controlling zip expression through down regulation of Egfr in this tissue. Over-expression of Ack in the AS during germband retraction causes a dramatic increase in zip levels in this tissue [26](Compare Fig. 6A to Fig. 6B), but co-expression with Egfr-EGFP (but not a control lacZ transgene) restored wild-type zip levels, suggesting that Ack controls zip through down regulation of Egfr (Fig. 6C, D). We subsequently over-expressed Ack in prd stripes in the AS and examined the effect on Egfr distribution by comparison with adjacent amnioserosa cells not over-expressing Ack. AS cells with endogenous levels of Ack showed strong cortical Egfr immunostaining as well as staining in cytoplasmic puncta, some of which were Rab5 positive and therefore early endosomes (Fig. 6E–E ´´´). In Ack-over-expressing cells (identified by increased levels of phosphotyrosine [101]) there was a decrease in cortical Egfr staining and an increase in Egfr-positive cytoplasmic puncta, with some of these being Rab5-positive; these cells also showed a general increase in the levels of early endosomes (Fig. 6E–E ´´´). Many of the Egfr-positive puncta in these cells were Rab5-negative and we suspect that they may be multivesicular bodies, where endocytosed Egfr is known to accumulate (reviewed in [100]). The increase in early endosomes in Ack-over-expressing cells indicates that Ack over-expression leads to a general increase in endocytosis in AS cells. In support of this is an observation we made when trying to observe the effects of Ack on apoptosis using the Apoliner reporter. Here, with the expression of kinase-dead Ack, which is more effective than wild-type Ack at inducing zip expression [26], a highly punctate distribution of membrane-localized Apoliner-RFP signal was observed while control embryos at the same stage of DC displayed a typical homogeneous distribution (Fig. 6G, H). We interpret this difference as reflecting a general increase in intracellular vesicular traffic, consistent with the effect of Ack over-expression in promoting Egfr endocytosis.
Figure 6. Evidence that Egfr signaling is negatively regulated by endocytosis in the AS.

(A–D) zip FISH on embryos late in germband retraction. (A) Wild-type embryo showing zip expression in AS. (B) Expression of Ack in the AS using the Gal4c381 driver causes an increase in zip levels in this tissue relative to wild-type. (C) Ack fails to elevate zip levels when co-expressed with Egfr–EGFP. (D) zip levels are elevated when Ack is co-expressed with control lacZ gene. (E–É ´´) AS in which Ack had been over-expressed in prd stripes, triple-stained with anti-phosphotyrosine (anti-PY) (E), anti-Egfr (É) and anti-Rab5 (É ´). (E) Cells over-expressing Ack are marked by high levels of anti-PY (outlined with dotted lines). (É) Egfr shows strong cortical localization in wild-type AS cells but a more cytoplasmic distribution in Ack-over-expressing cells. (É ´) There is an increase in Rab5-positive early endosomes in Ack-over-expressing cells. (É ´´) Merge of panels É and É ´. Arrowheads and arrows mark Egfr-positive early endosomes in wild-type cells and Ack-over-expressing cells, respectively. (F) Egfr-EGFP expressed in the AS using the Gal4NP3312 driver shows vesicular accumulation in addition to being at the plasma membrane. (G) AS cells in embryo in which Apoliner has been expressed with LP1-Gal4 driver showing localization of Apoliner-RFP signal to membranes. (H) AS cells in embryo in which Apoliner and kinase-dead Ack have been co-expressed with LP1-Gal4 driver showing punctate localization of Apoliner-RFP signal. Scale bars: 50 µm in A-D; 5 µm in E-H.
Discussion
We have determined that Egfr is required in both the AS and epidermis for DC to proceed normally and our results suggest that Egfr signaling has a least three distinct roles in DC, all of which act to repress morphogenesis (see model in Fig. 7). Egfr is a negative regulator of dpp expression in the epidermis as loss of Egfr function in either Egfr mutant embryos or as a result of EgfrDN expression leads to ectopic dpp expression. dpp is expressed in two stripes during DC, one composed of the DME cells and the other running along the ventrolateral epidermis, where dpp expression in the DME cells, but not the ventrolateral stripe, is dependent on a JNK MAPK cascade [13]–[18]. Consistent with the notion that Egfr functions as a negative regulator of dpp expression, activation of the Egfr pathway can repress dpp expression in either stripe. The down regulation of dpp expression in both stripes, however, supports the view that Egfr does not reduce dpp transcription by impacting the JNK pathway, in which case we would expect to observe down regulation of dpp expression only in the DME stripe. How might Egfr signaling be regulating dpp expression? Wingless (Wg) is a diffusible signal required for proper dpp expression in both the dorsal and ventrolateral stripes during DC [102], [103]. Egfr negatively regulates Wg levels in the eye imaginal disc by transcriptionally regulating phyllopod [104]; we looked for evidence that Egfr might be controlling dpp expression through Wg, but altering Egfr signaling had no discernable effect on phyllopod transcription or Wg distribution in the embryo (X. C., unpublished results).
Figure 7. Model for Egfr acting as a brake on DC.
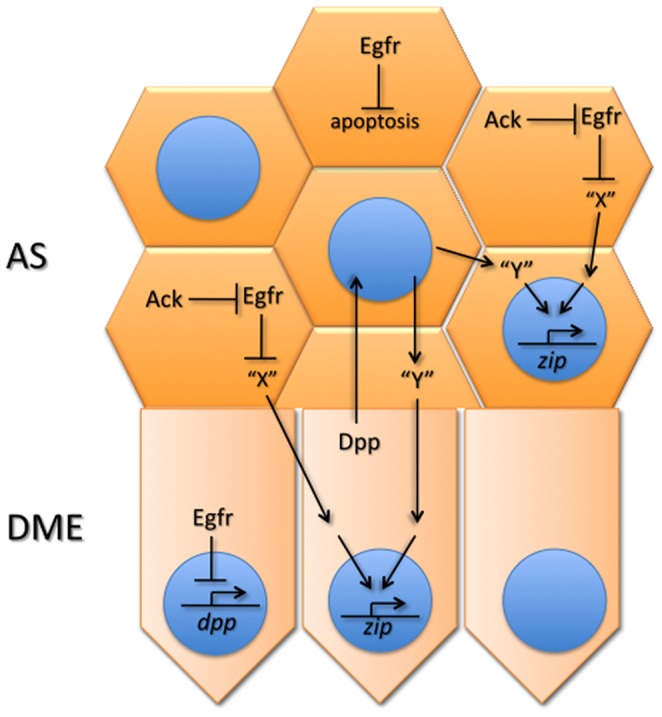
Egfr negatively regulates the production and/or secretion of a diffusible signal “X” in the AS (AS) and is itself negatively regulated by Ack through endocytosis. “X” signals into both the AS and the DME cells where it activates a pathway promoting transcription of myosin from the zip locus. Previous work from our group and others, and unpublished results from our group, suggest that Dpp from the DME cells diffuses to the AS where it regulates production of a second diffusible signal “Y” providing a parallel input into zip transcription. Myosin produced through the cooperation of the two pathways then drives morphogenesis of the AS and DME cells. Egfr additionally regulates this signaling network by negatively regulating dpp transcription in the epidermis, including the DME cells. Egfr further regulates AS morphogenesis by inhibiting apoptosis in this tissue.
The defects in morphogenesis seen in embryos with impaired Egfr signaling are likely at least in part due to misregulated actomyosin contractility. A recurring theme associated with various circumstances of Egfr impairment is the bowed embryo phenotype, where segments are bunched together at the leading edge of the epidermis during DC. We suggest this is due to uneven actomyosin contractility in the dorsal epidermis associated with excessive zip expression. The loss of epithelial integrity or “pulling apart” of the head seen in live imaging of Egfr mutant embryos may similarly be due to misregulated actomyosin contractility and/or excessive apoptosis. Previous work has indicated that a major function for Egfr in the head is inhibition of apoptosis, similar to its role in the AS [74], [105], [106]. Parallels between the regulation of head involution and DC have been noted and Egfr may function to modulate these two morphogenetic events through similar mechanisms [107].
While it is likely that increased levels of Dpp in Egfr mutant embryos contribute to the elevated zip levels, our results of manipulating Egfr signaling support the interpretation of a separate route for zip regulation that involves signaling from the AS to both the AS and the epidermis. This signaling is not operating through the regulation of dpp expression as impairment of Egfr signaling in the AS does not affect Dpp levels. Thus, we consider this zip regulation a second distinct role for Egfr in DC and we believe this signaling is the same as that regulated by Ack in its control of zip expression. Consistent with this, gains or losses of Ack do not affect the Dpp pathway, supporting the view that Ack operates in parallel to Dpp signaling [26], [101]. The Ack/Egfr-regulated signal could be a diffusible ligand (“X” in Fig. 7) produced in the AS cells that activates a pathway in the AS and DME cells, thereby driving zip expression [26]. Alternatively, Egfr could promote the production of a signal that negatively regulates the pathway required for zip expression. Moreover, the upregulation of zip expression in Egfr signaling deficient embryos does not appear to be due to premature apoptosis of the AS as elevated zip can be seen in Egfr signaling deficient embryos that still have an intact AS. It is possible, however, that the signaling events regulating zip expression occur at a stage prior to the stage at which the AS is lost in Egfr mutants, i.e. before initiation of germband retraction. Preliminary data leads us to propose that Dpp acts in parallel to produce a second diffusible ligand (“Y” in Fig. 7) that activates a second pathway contributing to zip expression (W. S and X. C., unpublished observations). Thus, the interplay between Egfr and Dpp during DC is complex, involving multiple pathways and bidirectional communication between two tissues, and this complex signaling arrangement may function to ensure the coordinated morphogenesis of the AS and epidermis.
A third major role for Egfr in DC is as a negative regulator of apoptosis in the AS. Enhancement of apoptosis accelerates DC whereas suppression of apoptosis slows it, indicating that apoptosis, similar to actomyosin contractility, provides a force for morphogenesis [3]. Thus, down regulation of Egfr in the AS during DC provides two means to accelerate the process: increased myosin expression and increased cell death. The “tweaking” of Egfr function in the AS could constitute an important regulatory mechanism for controlling the rate of closure. We have provided evidence that endocytosis, promoted by Ack, is a route by which Egfr signaling is controlled in the AS cells. Our results suggest that Ack would have a pro-apoptotic role in the AS through promotion of Egfr endocytosis. This in contrast to the Drosophila eye in which Ack has an anti-apoptotic function that is independent of Egfr [108].
A recent study has demonstrated that endocytosis in the AS is required for its correct morphogenesis during DC, but this work focused on the role of endocytosis in removing membrane to promote cell shape change [109]. Our results indicate that another route of action for endocytosis in the AS is in regulation of Egfr signaling. It has been suggested that endocytosis could act as a rheostat in which membrane area is adjusted in response to actomyosin contractility [109]; such a rheostat could also be used to adjust Egfr signaling throughout DC.
Additional avenues for Egfr regulation during DC could be control of ligands binding to Egfr and feedback inhibition [110], [111], but we have yet to address these. In summary, we have identified Egfr signaling as an inhibitor of morphogenesis during DC that acts at several distinct levels. Having a single pathway control multiple aspects of this complex process may simplify feedback regulation, ensuring that morphogenesis occurs in a coordinated fashion. In essence, Egfr signaling acts as a brake that can be applied when required to ensure that closure proceeds smoothly and without loss of epidermal integrity. DC shows striking parallels to the healing of induced wounds in the Drosophila embryo, with the two processes using similar cytoskeletal and signaling machineries [112]–[114]. Egfr has recently been shown to be required for healing of induced wounds in the embryo and it will be of interest to determine if it uses similar routes of action in this as we have shown in DC [115].
Materials and Methods
Fly STOCKS
Egfr2C82 and Egfr1F26 were gifts from T. Schüpbach, UAS-Egfr-EGFP from J. Duffy, UAS-sSpi from B. Shilo, UAS-RasN17 from T. Lee, LE-Gal4 from S. Noselli, Ubi-DEcadherin-GFP from H. Oda, LP1-Gal4 from G. Morata, Gal4NP5328 and Gal4NP3312 from the Kyoto Drosophila Resource Center and UAS-Apoliner and tub-Apoliner from P.L. Bardet. Egfr1a15 and EgfrH25 were isolated from a collection of EMS-mutagenized second chromosomes [38]. cn1 Egfrf2 bw1 sp1/CyO flies and all other stocks were obtained from the Bloomington Drosophila Stock Center. Crosses were performed at 25°C.
Cuticle Preparations
Cuticles were prepared as described but with the fixation step removed [116]. At least 100 embryos were examined in each experiment.
Immunohistochemistry and RNA in Situ Hybridization
Fixing and antibody staining of embryos were done as previously described [117], [118]. The following primary antibodies were used: mouse anti-phosphotyrosine (Cell Signaling)(1∶1000), rabbit anti-GFP (1∶500)(Sigma), mouse anti-GFP (1∶500)(Sigma), goat anti-Egfr (Santa Cruz)(1∶5), rabbit anti-phospho-MAPK (Cell Signaling)(1∶50) and rabbit anti-Rab5 (Abcam)(1∶1000). All secondary antibodies were from Vector Laboratories and used at a 1∶200 dilution. In situ mRNA hybridizations using digoxigenin-labeled RNA probes and FISH were performed as described [119], [120]. cDNAs for in situ hybridization probes were obtained from the Canadian Drosophila Microarray Centre. Fluorescently-stained embryos were examined on either a Zeiss LSM 410 laser-scanning confocal microscope or a Quorum spinning disk confocal microscope, and digoxigenin-labeled embryos imaged using a Zeiss Axioplan 2 microscope. Images were processed in Adobe Photoshop. The genotypes of all fluorescently-stained embryos were established by tracking balancer chromosomes bearing GFP reporters.
Live Imaging of Embryos
Chromosomes carrying Ubi-DEcadherin-GFP in combination with the alleles Egfrf2, Egfr1a15, and EgfrH25 were recovered by meiotic recombination. Similarly, the tub-Apoliner insertion, which expresses Apoliner under the control of the tubulin 1 α promoter (described in [97]), was recombined with Egfrf2. All recombinant chromosomes were maintained over so-called “GFP-balancer” chromosomes obtained from the Bloomington Drosophila Stock Center (either CyO, P{w[+mC] = GAL4-Kr.C}DC3, P{w[+mC] = UAS-GFP.S65T}DC7 or CyO, P{w[+mC] = GAL4-twi.G}2.2, P{UAS-2xEGFP}AH2.2). For live imaging experiments Egfr mutant embryos were unambiguously identified as those lacking GFP expression derived from the GFP-balancer. Since the onset of the Egfr mutant phenotype preceded the timing of GFP expression associated with either GFP-balancer stock, the selection of embryos for live imaging was random and identification of mutant embryos was achieved subsequent to image acquisition. The early onset of the Egfr mutant phenotype also preceded the time at which all available amnioserosa specific GAL4 drivers could induce reporter gene expression, and for this reason our analysis of caspase activity in Egfr mutant embryos required using tub-Apoliner carried by the maternal parent.
Embryos were prepared for live imaging using the hanging drop protocol, which eliminates effects of compression on the mounted embryo [121]. Time-lapse confocal microscopy was performed using a 20X Plan Apo VC objective on a Nikon Eclipse 90 i microscope with a Nikon D-eclipse C1 scan head. Images were saved as animated projections using the Nikon EZ-C1 3.70 software and further processed using ImageJ (NIH).
Supporting Information
Time-lapse showing restricted expression pattern of LE-Gal4 driver, revealed using a UAS-GFP-NLS reporter. Cell outlines were revealed through expression of a Ubi-DEcadherin-GFP transgene.
(MOV)
Time-lapse showing unfolding of the AS as germband retraction proceeds in a Ubi-DEcadherin-GFP- expressing embryo.
(MOV)
Time-lapse showing delayed germband retraction and disintegration of AS in Egfrf2 mutant embryo expressing Ubi-DEcadherin-GFP .
(MOV)
Time-lapse showing delayed germband retraction and disintegration of AS in Egfrf2 / EgfrH25 mutant embryo expressing Ubi-DEcadherin-GFP .
(MOV)
Time-lapse showing bowing of Egfr1a15 / EgfrH25 mutant, Ubi-DEcadherin-GFP- expressing embryo.
(MOV)
Time-lapse showing morphogenesis of the AS in a Ubi-DEcadherin-GFP- expressing embryo.
(MOV)
Time-lapse showing AS of stage 15 wild-type embryo in which GFP had been expressed with the Gal4NP3312 AS driver, showing morphogenesis of the AS into a narrow, tube-like structure.
(MOV)
Time-lapse showing AS of stage 15 embryo in which Egfr-EGFP and GFP-NLS had been expressed with the double driver combination Gal4NP3312 + Gal4NP5328 showing failure of AS morphogenesis.
(MOV)
Time-lapse showing AS of stage 15 embryo in which Egfr-EGFP and GFP-NLS had been expressed with the double driver combination Gal4NP3312 + Gal4NP5328 showing failure of AS morphogenesis.
(MOV)
Acknowledgments
We thank Baharak Zahedi and Barton Xu for preliminary observations suggesting that Ack negatively regulates Egfr signaling during development. We thank P. L. Bardet, J. Duffy, G. Morata, T. Lee, S. Noselli, H. Oda, T. Schüpbach, and B. Shilo for fly stocks, and T. Orr-Weaver, I. Royzman and A. Whittaker for the unpublished Egfr1a15 and EgfrH25 alleles. We are grateful to Esther Verheyen for discussions and comments on the manuscript.
Funding Statement
This work was funded by Canadian Institutes of Health Research grant no. MOP 74513 to NH. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Harden N (2002) Signaling pathways directing the movement and fusion of epithelial sheets: lessons from dorsal closure in Drosophila . Differentiation 70: 181–203. [DOI] [PubMed] [Google Scholar]
- 2. Reed BH, Wilk R, Schock F, Lipshitz HD (2004) Integrin-dependent apposition of Drosophila extraembryonic membranes promotes morphogenesis and prevents anoikis. Curr Biol 14: 372–380. [DOI] [PubMed] [Google Scholar]
- 3. Toyama Y, Peralta XG, Wells AR, Kiehart DP, Edwards GS (2008) Apoptotic force and tissue dynamics during Drosophila embryogenesis. Science 321: 1683–1686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Kiehart DP, Galbraith CG, Edwards KA, Rickoll WL, Montague RA (2000) Multiple forces contribute to cell sheet morphogenesis for dorsal closure in Drosophila . J Cell Biol 149: 471–490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Heisenberg CP (2009) Dorsal closure in Drosophila: cells cannot get out of the tight spot. Bioessays 31: 1284–1287. [DOI] [PubMed] [Google Scholar]
- 6. Jacinto A, Wood W, Balayo T, Turmaine M, Martinez-Arias A, et al. (2000) Dynamic actin-based epithelial adhesion and cell matching during Drosophila dorsal closure. Curr Biol 10: 1420–1426. [DOI] [PubMed] [Google Scholar]
- 7. Jacinto A, Wood W, Woolner S, Hiley C, Turner L, et al. (2002) Dynamic analysis of actin cable function during Drosophila dorsal closure. Curr Biol 12: 1245–1250. [DOI] [PubMed] [Google Scholar]
- 8. Riesgo-Escovar JR, Hafen E (1997) Common and distinct roles of DFos and DJun during Drosophila development. Science 278: 669–672. [DOI] [PubMed] [Google Scholar]
- 9. Zeitlinger J, Kockel L, Peverali FA, Jackson DB, Mlodzik M, et al. (1997) Defective dorsal closure and loss of epidermal decapentaplegic expression in Drosophila fos mutants. EMBO J 16: 7393–7401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Wada A, Kato K, Uwo MF, Yonemura S, Hayashi S (2007) Specialized extraembryonic cells connect embryonic and extraembryonic epidermis in response to Dpp during dorsal closure in Drosophila . Dev Biol 301: 340–349. [DOI] [PubMed] [Google Scholar]
- 11. Ricos MG, Harden N, Sem KP, Lim L, Chia W (1999) Dcdc42 acts in TGF-β signaling during Drosophila morphogenesis: distinct roles for the Drac1/JNK and Dcdc42/TGF-β cascades in cytoskeletal regulation. J Cell Sci 112: 1225–1235. [DOI] [PubMed] [Google Scholar]
- 12. Young PE, Richman AM, Ketchum AS, Kiehart DP (1993) Morphogenesis in Drosophila requires nonmuscle myosin heavy chain function. Genes Dev 7: 29–41. [DOI] [PubMed] [Google Scholar]
- 13. Glise B, Noselli S (1997) Coupling of Jun amino-terminal kinase and Decapentaplegic signaling pathways in Drosophila morphogenesis. Genes Dev 11: 1738–1747. [DOI] [PubMed] [Google Scholar]
- 14. Jackson PD, Hoffmann FM (1994) Embryonic expression patterns of the Drosophila decapentaplegic gene: separate regulatory elements control blastoderm expression and lateral ectodermal expression. Dev Dyn 199: 28–44. [DOI] [PubMed] [Google Scholar]
- 15. St Johnston RD, Gelbart WM (1987) Decapentaplegic transcripts are localized along the dorsal-ventral axis of the Drosophila embryo. EMBO J 6: 2785–2791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Sluss HK, Davis RJ (1997) Embryonic morphogenesis signaling pathway mediated by JNK targets the transcription factor JUN and the TGF-β homologue decapentaplegic . J Cell Biochem 67: 1–12. [PubMed] [Google Scholar]
- 17. Riesgo-Escovar JR, Hafen E (1997) Drosophila Jun kinase regulates expression of decapentaplegic via the ETS-domain protein Aop and the AP-1 transcription factor DJun during dorsal closure. Genes Dev 11: 1717–1727. [DOI] [PubMed] [Google Scholar]
- 18. Hou XS, Goldstein ES, Perrimon N (1997) Drosophila Jun relays the Jun amino-terminal kinase signal transduction pathway to the Decapentaplegic signal transduction pathway in regulating epithelial cell sheet movement. Genes Dev 11: 1728–1737. [DOI] [PubMed] [Google Scholar]
- 19. Childs SR, Wrana JL, Arora K, Attisano L, O'Connor MB, et al. (1993) Identification of a Drosophila activin receptor. Proc Natl Acad Sci U S A 90: 9475–9479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Affolter M, Nellen D, Nussbaumer U, Basler K (1994) Multiple requirements for the receptor serine/threonine kinase thick veins reveal novel functions of TGF β homologs during Drosophila embryogenesis. Development 120: 3105–3117. [DOI] [PubMed] [Google Scholar]
- 21. Penton A, Chen Y, Staehling-Hampton K, Wrana JL, Attisano L, et al. (1994) Identification of two bone morphogenetic protein type I receptors in Drosophila and evidence that Brk25D is a Decapentaplegic receptor. Cell 78: 239–250. [DOI] [PubMed] [Google Scholar]
- 22. Letsou A, Arora K, Wrana JL, Simin K, Twombly V, et al. (1995) Drosophila Dpp signaling is mediated by the punt gene product: a dual ligand-binding type II receptor of the TGF β receptor family. Cell 80: 899–908. [DOI] [PubMed] [Google Scholar]
- 23. Ruberte E, Marty T, Nellen D, Affolter M, Basler K (1995) An absolute requirement for both the type II and type I receptors, Punt and Thick veins, for Dpp signaling in vivo. Cell 80: 889–897. [DOI] [PubMed] [Google Scholar]
- 24. Arquier N, Perrin L, Manfruelli P, Semeriva M (2001) The Drosophila tumor suppressor gene lethal(2)giant larvae is required for the emission of the Decapentaplegic signal. Development 128: 2209–2220. [DOI] [PubMed] [Google Scholar]
- 25. Fernandez BG, Arias AM, Jacinto A (2007) Dpp signalling orchestrates dorsal closure by regulating cell shape changes both in the amnioserosa and in the epidermis. Mech Dev 124: 884–897. [DOI] [PubMed] [Google Scholar]
- 26. Zahedi B, Shen W, Xu X, Chen X, Mahey M, et al. (2008) Leading edge-secreted Dpp cooperates with ACK-dependent signaling from the amnioserosa to regulate myosin levels during dorsal closure. Dev Dyn 237: 2936–2946. [DOI] [PubMed] [Google Scholar]
- 27. Franke JD, Montague RA, Kiehart DP (2005) Nonmuscle myosin II generates forces that transmit tension and drive contraction in multiple tissues during dorsal closure. Curr Biol 15: 2208–2221. [DOI] [PubMed] [Google Scholar]
- 28. Hopper NA, Lee J, Sternberg PW (2000) ARK-1 inhibits EGFR signaling in C. elegans. Mol Cell 6: 65–75. [PubMed] [Google Scholar]
- 29.Lin Q, Lo CG, Cerione RA, Yang W (2002) The Cdc42 target ACK2 interacts with sorting nexin 9 (SH3PX1) to regulate epidermal growth factor receptor degradation. J Biol Chem 277: 10134–10138. Epub 12002 Jan 10117. [DOI] [PubMed]
- 30. Shen F, Lin Q, Gu Y, Childress C, Yang W (2007) Activated Cdc42-associated kinase 1 is a component of EGF receptor signaling complex and regulates EGF receptor degradation. Mol Biol Cell 18: 732–742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Lin Q, Wang J, Childress C, Sudol M, Carey DJ, et al. (2010) HECT E3 ubiquitin ligase Nedd4–1 ubiquitinates ACK and regulates epidermal growth factor (EGF)-induced degradation of EGF receptor and ACK. Mol Cell Biol 30: 1541–1554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Grovdal LM, Johannessen LE, Rodland MS, Madshus IH, Stang E (2008) Dysregulation of Ack1 inhibits down-regulation of the EGF receptor. Exp Cell Res 314: 1292–1300. [DOI] [PubMed] [Google Scholar]
- 33. Schweitzer R, Shilo BZ (1997) A thousand and one roles for the Drosophila EGF receptor. Trends Genet 13: 191–196. [DOI] [PubMed] [Google Scholar]
- 34. Shilo BZ (2003) Signaling by the Drosophila epidermal growth factor receptor pathway during development. Exp Cell Res 284: 140–149. [DOI] [PubMed] [Google Scholar]
- 35. Clifford R, Schupbach T (1992) The torpedo (DER) receptor tyrosine kinase is required at multiple times during Drosophila embryogenesis. Development 115: 853–872. [DOI] [PubMed] [Google Scholar]
- 36. Clifford RJ, Schupbach T (1989) Coordinately and differentially mutable activities of torpedo, the Drosophila melanogaster homolog of the vertebrate EGF receptor gene. Genetics 123: 771–787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Nusslein-Volhard C, Wieschaus E, Kluding H (1984) Mutations affecting the pattern of the larval cuticle in Drosophila melanogaster I. Zygotic loci on the second chromosome. Roux's Arch Dev Biol 193: 267–282. [DOI] [PubMed] [Google Scholar]
- 38. Royzman I, Whittaker AJ, Orr-Weaver TL (1997) Mutations in Drosophila DP and E2F distinguish G1-S progression from an associated transcriptional program. Genes Dev 11: 1999–2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Perkins LA, Johnson MR, Melnick MB, Perrimon N (1996) The nonreceptor protein tyrosine phosphatase corkscrew functions in multiple receptor tyrosine kinase pathways in Drosophila . Dev Biol 180: 63–81. [DOI] [PubMed] [Google Scholar]
- 40. Brand AH, Perrimon N (1993) Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118: 401–415. [DOI] [PubMed] [Google Scholar]
- 41. Hinz U, Giebel B, Campos-Ortega JA (1994) The basic-helix-loop-helix domain of Drosophila lethal of scute protein is sufficient for proneural function and activates neurogenic genes. Cell 76: 77–87. [DOI] [PubMed] [Google Scholar]
- 42. Manseau L, Baradaran A, Brower D, Budhu A, Elefant F, et al. (1997) GAL4 enhancer traps expressed in the embryo, larval brain, imaginal discs, and ovary of Drosophila . Dev Dyn 209: 310–322. [DOI] [PubMed] [Google Scholar]
- 43. Gabay L, Seger R, Shilo BZ (1997) MAP kinase in situ activation atlas during Drosophila embryogenesis. Development 124: 3535–3541. [DOI] [PubMed] [Google Scholar]
- 44. Gabay L, Seger R, Shilo BZ (1997) In situ activation pattern of Drosophila EGF receptor pathway during development. Science 277: 1103–1106. [DOI] [PubMed] [Google Scholar]
- 45. Oda H, Tsukita S (2001) Real-time imaging of cell-cell adherens junctions reveals that Drosophila mesoderm invagination begins with two phases of apical constriction of cells. J Cell Sci 114: 493–501. [DOI] [PubMed] [Google Scholar]
- 46. Byars CL, Bates KL, Letsou A (1999) The dorsal-open group gene raw is required for restricted DJNK signaling during closure. Development 126: 4913–4923. [DOI] [PubMed] [Google Scholar]
- 47. Wei HC, Sanny J, Shu H, Baillie DL, Brill JA, et al. (2003) The Sac1 lipid phosphatase regulates cell shape change and the JNK cascade during dorsal closure in Drosophila . Curr Biol 13: 1882–1887. [DOI] [PubMed] [Google Scholar]
- 48. Martin-Blanco E, Gampel A, Ring J, Virdee K, Kirov N, et al. (1998) puckered encodes a phosphatase that mediates a feedback loop regulating JNK activity during dorsal closure in Drosophila . Genes Dev 12: 557–570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Yagi Y, Hayashi S (1997) Role of the Drosophila EGF receptor in determination of the dorsoventral domains of escargot expression during primary neurogenesis. Genes Cells 2: 41–53. [DOI] [PubMed] [Google Scholar]
- 50. Chen CK, Kuhnlein RP, Eulenberg KG, Vincent S, Affolter M, et al. (1998) The transcription factors KNIRPS and KNIRPS RELATED control cell migration and branch morphogenesis during Drosophila tracheal development. Development 125: 4959–4968. [DOI] [PubMed] [Google Scholar]
- 51. Dobens LL, Peterson JS, Treisman J, Raftery LA (2000) Drosophila bunched integrates opposing DPP and EGF signals to set the operculum boundary. Development 127: 745–754. [DOI] [PubMed] [Google Scholar]
- 52. Kumar JP, Moses K (2001) The EGF receptor and notch signaling pathways control the initiation of the morphogenetic furrow during Drosophila eye development. Development 128: 2689–2697. [DOI] [PubMed] [Google Scholar]
- 53. Chen Y, Schupbach T (2006) The role of brinker in eggshell patterning. Mech Dev 123: 395–406. [DOI] [PubMed] [Google Scholar]
- 54. Carneiro K, Fontenele M, Negreiros E, Lopes E, Bier E, et al. (2006) Graded maternal short gastrulation protein contributes to embryonic dorsal-ventral patterning by delayed induction. Dev Biol 296: 203–218. [DOI] [PubMed] [Google Scholar]
- 55. Shravage BV, Altmann G, Technau M, Roth S (2007) The role of Dpp and its inhibitors during eggshell patterning in Drosophila . Development 134: 2261–2271. [DOI] [PubMed] [Google Scholar]
- 56. Yan SJ, Zartman JJ, Zhang M, Scott A, Shvartsman SY, et al. (2009) Bistability coordinates activation of the EGFR and DPP pathways in Drosophila vein differentiation. Mol Syst Biol 5: 278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Liu M, Lim TM, Cai Y (2010) The Drosophila female germline stem cell lineage acts to spatially restrict DPP function within the niche. Sci Signal 3: ra57. [DOI] [PubMed] [Google Scholar]
- 58. Szuts D, Bienz M (2000) An autoregulatory function of Dfos during Drosophila endoderm induction. Mech Dev 98: 71–76. [DOI] [PubMed] [Google Scholar]
- 59. Kopp A, Blackman RK, Duncan I (1999) Wingless, decapentaplegic and EGF receptor signaling pathways interact to specify dorso-ventral pattern in the adult abdomen of Drosophila . Development 126: 3495–3507. [DOI] [PubMed] [Google Scholar]
- 60. Wappner P, Gabay L, Shilo BZ (1997) Interactions between the EGF receptor and DPP pathways establish distinct cell fates in the tracheal placodes. Development 124: 4707–4716. [DOI] [PubMed] [Google Scholar]
- 61. Peri F, Roth S (2000) Combined activities of Gurken and decapentaplegic specify dorsal chorion structures of the Drosophila egg. Development 127: 841–850. [DOI] [PubMed] [Google Scholar]
- 62. Deng WM, Bownes M (1997) Two signalling pathways specify localised expression of the Broad-Complex in Drosophila eggshell patterning and morphogenesis. Development 124: 4639–4647. [DOI] [PubMed] [Google Scholar]
- 63. de Celis JF (1997) Expression and function of decapentaplegic and thick veins during the differentiation of the veins in the Drosophila wing. Development 124: 1007–1018. [DOI] [PubMed] [Google Scholar]
- 64. Carmena A, Gisselbrecht S, Harrison J, Jimenez F, Michelson AM (1998) Combinatorial signaling codes for the progressive determination of cell fates in the Drosophila embryonic mesoderm. Genes Dev 12: 3910–3922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Martin-Blanco E, Roch F, Noll E, Baonza A, Duffy JB, et al. (1999) A temporal switch in DER signaling controls the specification and differentiation of veins and interveins in the Drosophila wing. Development 126: 5739–5747. [DOI] [PubMed] [Google Scholar]
- 66. Jordan KC, Clegg NJ, Blasi JA, Morimoto AM, Sen J, et al. (2000) The homeobox gene mirror links EGF signalling to embryonic dorso-ventral axis formation through notch activation. Nat Genet 24: 429–433. [DOI] [PubMed] [Google Scholar]
- 67. Dequier E, Souid S, Pal M, Maroy P, Lepesant JA, et al. (2001) Top-DER- and Dpp-dependent requirements for the Drosophila fos/kayak gene in follicular epithelium morphogenesis. Mech Dev 106: 47–60. [DOI] [PubMed] [Google Scholar]
- 68. Shirai T, Maehara A, Kiritooshi N, Matsuzaki F, Handa H, et al. (2003) Differential requirement of EGFR signaling for the expression of defective proventriculus gene in the Drosophila endoderm and ectoderm. Biochem Biophys Res Commun 311: 473–477. [DOI] [PubMed] [Google Scholar]
- 69. Motola S, Neuman-Silberberg FS (2004) spoonbill, a new Drosophila female-sterile mutation, interferes with chromosome organization and dorsal-ventral patterning of the egg. Dev Dyn 230: 535–545. [DOI] [PubMed] [Google Scholar]
- 71. O'Keefe DD, Prober DA, Moyle PS, Rickoll WL, Edgar BA (2007) Egfr/Ras signaling regulates DE-cadherin/Shotgun localization to control vein morphogenesis in the Drosophila wing. Dev Biol 311: 25–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Yakoby N, Lembong J, Schupbach T, Shvartsman SY (2008) Drosophila eggshell is patterned by sequential action of feedforward and feedback loops. Development 135: 343–351. [DOI] [PubMed] [Google Scholar]
- 73. Kim SY, Jung KI, Kim SH, Jeon SH (2008) Dpp represses eagle expression at short-range, but can repress its expression at a long-range via EGFR signal repression. Mol Cells 26: 576–582. [PubMed] [Google Scholar]
- 74. Chang T, Shy D, Hartenstein V (2003) Antagonistic relationship between Dpp and EGFR signaling in Drosophila head patterning. Dev Biol 263: 103–113. [DOI] [PubMed] [Google Scholar]
- 75. Crozatier M, Glise B, Vincent A (2002) Connecting Hh, Dpp and EGF signalling in patterning of the Drosophila wing; the pivotal role of collier/knot in the AP organiser. Development 129: 4261–4269. [DOI] [PubMed] [Google Scholar]
- 76. Kubota K, Goto S, Eto K, Hayashi S (2000) EGF receptor attenuates Dpp signaling and helps to distinguish the wing and leg cell fates in Drosophila . Development 127: 3769–3776. [DOI] [PubMed] [Google Scholar]
- 77. Letizia A, Barrio R, Campuzano S (2007) Antagonistic and cooperative actions of the EGFR and Dpp pathways on the iroquois genes regulate Drosophila mesothorax specification and patterning. Development 134: 1337–1346. [DOI] [PubMed] [Google Scholar]
- 78. Sotillos S, De Celis JF (2005) Interactions between the Notch, EGFR, and decapentaplegic signaling pathways regulate vein differentiation during Drosophila pupal wing development. Dev Dyn 232: 738–752. [DOI] [PubMed] [Google Scholar]
- 79. Szuts D, Eresh S, Bienz M (1998) Functional intertwining of Dpp and EGFR signaling during Drosophila endoderm induction. Genes Dev 12: 2022–2035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. von Ohlen T, Doe CQ (2000) Convergence of Dorsal, Dpp, and Egfr signaling pathways subdivides the Drosophila neuroectoderm into three dorsal-ventral columns. Dev Biol 224: 362–372. [DOI] [PubMed] [Google Scholar]
- 81. Wahlstrom G, Norokorpi HL, Heino TI (2006) Drosophila alpha-actinin in ovarian follicle cells is regulated by EGFR and Dpp signalling and required for cytoskeletal remodelling. Mech Dev 123: 801–818. [DOI] [PubMed] [Google Scholar]
- 82. Kretzschmar M, Doody J, Massague J (1997) Opposing BMP and EGF signalling pathways converge on the TGF-β family mediator Smad1. Nature 389: 618–622. [DOI] [PubMed] [Google Scholar]
- 83. Radke K, Johnson K, Guo R, Davidson A, Ambrosio L (2001) Drosophila-raf acts to elaborate dorsoventral pattern in the ectoderm of developing embryos. Genetics 159: 1031–1044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Lamka ML, Lipshitz HD (1999) Role of the amnioserosa in germ band retraction of the Drosophila melanogaster embryo. Dev Biol 214: 102–112. [DOI] [PubMed] [Google Scholar]
- 85. Reed BH, Wilk R, Lipshitz HD (2001) Downregulation of Jun kinase signaling in the amnioserosa is essential for dorsal closure of the Drosophila embryo. Curr Biol 11: 1098–1108. [DOI] [PubMed] [Google Scholar]
- 86. Stronach BE, Perrimon N (2001) Investigation of leading edge formation at the interface of amnioserosa and dorsal ectoderm in the Drosophila embryo. Development 128: 2905–2913. [DOI] [PubMed] [Google Scholar]
- 87. Conder R, Yu H, Ricos M, Hing H, Chia W, et al. (2004) dPak is required for integrity of the leading edge cytoskeleton during Drosophila dorsal closure but does not signal through the JNK cascade. Dev Biol 276: 378–390. [DOI] [PubMed] [Google Scholar]
- 88. Scuderi A, Letsou A (2005) Amnioserosa is required for dorsal closure in Drosophila . Dev Dyn 232: 791–800. [DOI] [PubMed] [Google Scholar]
- 89. Lada K, Gorfinkiel N, Martinez Arias A (2012) Interactions between the amnioserosa and the epidermis revealed by the function of the u-shaped gene. Biol Open 1: 353–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Schweitzer R, Shaharabany M, Seger R, Shilo BZ (1995) Secreted Spitz triggers the DER signaling pathway and is a limiting component in embryonic ventral ectoderm determination. Genes Dev 9: 1518–1529. [DOI] [PubMed] [Google Scholar]
- 91. Goentoro LA, Reeves GT, Kowal CP, Martinelli L, Schupbach T, et al. (2006) Quantifying the Gurken morphogen gradient in Drosophila oogenesis. Dev Cell 11: 263–272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Lee T, Feig L, Montell DJ (1996) Two distinct roles for Ras in a developmentally regulated cell migration. Development 122: 409–418. [DOI] [PubMed] [Google Scholar]
- 93. Frank LH, Rushlow C (1996) A group of genes required for maintenance of the amnioserosa tissue in Drosophila . Development 122: 1343–1352. [DOI] [PubMed] [Google Scholar]
- 94. Goldman-Levi R, Miller C, Greenberg G, Gabai E, Zak NB (1996) Cellular pathways acting along the germband and in the amnioserosa may participate in germband retraction of the Drosophila melanogaster embryo. Int J Dev Biol 40: 1043–1051. [PubMed] [Google Scholar]
- 95. Mohseni N, McMillan SC, Chaudhary R, Mok J, Reed BH (2009) Autophagy promotes caspase-dependent cell death during Drosophila development. Autophagy 5: 329–338. [DOI] [PubMed] [Google Scholar]
- 96. Kurada P, White K (1998) Ras promotes cell survival in Drosophila by downregulating hid expression. Cell 95: 319–329. [DOI] [PubMed] [Google Scholar]
- 97. Bardet PL, Kolahgar G, Mynett A, Miguel-Aliaga I, Briscoe J, et al. (2008) A fluorescent reporter of caspase activity for live imaging. Proc Natl Acad Sci U S A 105: 13901–13905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Cormier O, Mohseni N, Voytyuk I, Reed BH (2012) Autophagy can promote but is not required for epithelial cell extrusion in the amnioserosa of the Drosophila embryo. Autophagy 8: 252–264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Clem RJ, Fechheimer M, Miller LK (1991) Prevention of apoptosis by a baculovirus gene during infection of insect cells. Science 254: 1388–1390. [DOI] [PubMed] [Google Scholar]
- 100. Eden ER, White IJ, Futter CE (2009) Down-regulation of epidermal growth factor receptor signalling within multivesicular bodies. Biochem Soc Trans 37: 173–177. [DOI] [PubMed] [Google Scholar]
- 101. Sem KP, Zahedi B, Tan I, Deak M, Lim L, et al. (2002) ACK family tyrosine kinase activity is a component of Dcdc42 Signaling during dorsal closure in Drosophila melanogaster . Mol Cell Biol 22: 3685–3697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Morel V, Arias AM (2004) Armadillo/beta-catenin-dependent Wnt signalling is required for the polarisation of epidermal cells during dorsal closure in Drosophila . Development 131: 3273–3283. [DOI] [PubMed] [Google Scholar]
- 103. McEwen DG, Cox RT, Peifer M (2000) The canonical Wg and JNK signaling cascades collaborate to promote both dorsal closure and ventral patterning. Development 127: 3607–3617. [DOI] [PubMed] [Google Scholar]
- 104. Nagaraj R, Banerjee U (2009) Regulation of Notch and Wingless signalling by phyllopod, a transcriptional target of the EGFR pathway. EMBO J 28: 337–346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Dumstrei K, Nassif C, Abboud G, Aryai A, Hartenstein V (1998) EGFR signaling is required for the differentiation and maintenance of neural progenitors along the dorsal midline of the Drosophila embryonic head. Development 125: 3417–3426. [DOI] [PubMed] [Google Scholar]
- 106. Dumstrei K, Wang F, Shy D, Tepass U, Hartenstein V (2002) Interaction between EGFR signaling and DE-cadherin during nervous system morphogenesis. Development 129: 3983–3994. [DOI] [PubMed] [Google Scholar]
- 107. VanHook A, Letsou A (2008) Head involution in Drosophila: genetic and morphogenetic connections to dorsal closure. Dev Dyn 237: 28–38. [DOI] [PubMed] [Google Scholar]
- 108. Schoenherr JA, Drennan JM, Martinez JS, Chikka MR, Hall MC, et al. (2012) Drosophila activated Cdc42 kinase has an anti-apoptotic function. PLoS Genet 8: e1002725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Mateus AM, Gorfinkiel N, Schamberg S, Martinez Arias A (2011) Endocytic and recycling endosomes modulate cell shape changes and tissue behaviour during morphogenesis in Drosophila . PLoS One 6: e18729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110. Freeman M (2000) Feedback control of intercellular signalling in development. Nature 408: 313–319. [DOI] [PubMed] [Google Scholar]
- 111. Segatto S, Anastasi S, Alema S (2011) Regulation of epidermal growth factor signalling by inducible feedback inhibitors. J Cell Sci 124: 1785–1793. [DOI] [PubMed] [Google Scholar]
- 112. Wood W, Jacinto A, Grose R, Woolner S, Gale J, et al. (2002) Wound healing recapitulates morphogenesis in Drosophila embryos. Nat Cell Biol 4: 907–912. [DOI] [PubMed] [Google Scholar]
- 113. Martin P, Parkhurst SM (2004) Parallels between tissue repair and embryo morphogenesis. Development 131: 3021–3034. [DOI] [PubMed] [Google Scholar]
- 114. Campos I, Geiger JA, Santos AC, Carlos V, Jacinto A (2010) Genetic screen in Drosophila melanogaster uncovers a novel set of genes required for embryonic epithelial repair. Genetics 184: 129–140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Geiger JA, Carvalho L, Campos I, Santos AC, Jacinto A (2011) Hole-in-one mutant phenotypes link EGFR/ERK signaling to epithelial tissue repair in Drosophila . PLoS One 6: e28349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Ashburner M (1989) Drosophila: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. [Google Scholar]
- 117. Harden N, Lee J, Loh HY, Ong YM, Tan I, et al. (1996) A Drosophila homolog of the Rac- and Cdc42-activated serine/threonine kinase PAK is a potential focal adhesion and focal complex protein that colocalizes with dynamic actin structures. Mol Cell Biol 16: 1896–1908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. Harden N, Ricos M, Ong YM, Chia W, Lim L (1999) Participation of small GTPases in dorsal closure of the Drosophila embryo: distinct roles for Rho subfamily proteins in epithelial morphogenesis. J Cell Sci 112: 273–284. [DOI] [PubMed] [Google Scholar]
- 119.Van Vactor D, Kopczynski C (1999) Anatomical Techniques for Analysis of Nervous System Development in the Drosophila Embryo. In: Richter J, editor. A Comparative Methods Approach to the Study of Oocytes and Embryos. New York: Oxford University Press. 490–513. [Google Scholar]
- 120.Lecuyer E, Parthasarathy N, Krause HM (2007) Fluorescent in situ Hybridization Protocols in Drosophila Embryos and Tissues. In: Dahmann C, editor. Drosophila: Methods and Protocols. Totowa, NJ: Humana Press. [DOI] [PubMed] [Google Scholar]
- 121.Reed BH, McMillan SC, Chaudhary R (2009) The preparation of Drosophila embryos for live-imaging using the hanging drop protocol. J Vis Exp. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Time-lapse showing restricted expression pattern of LE-Gal4 driver, revealed using a UAS-GFP-NLS reporter. Cell outlines were revealed through expression of a Ubi-DEcadherin-GFP transgene.
(MOV)
Time-lapse showing unfolding of the AS as germband retraction proceeds in a Ubi-DEcadherin-GFP- expressing embryo.
(MOV)
Time-lapse showing delayed germband retraction and disintegration of AS in Egfrf2 mutant embryo expressing Ubi-DEcadherin-GFP .
(MOV)
Time-lapse showing delayed germband retraction and disintegration of AS in Egfrf2 / EgfrH25 mutant embryo expressing Ubi-DEcadherin-GFP .
(MOV)
Time-lapse showing bowing of Egfr1a15 / EgfrH25 mutant, Ubi-DEcadherin-GFP- expressing embryo.
(MOV)
Time-lapse showing morphogenesis of the AS in a Ubi-DEcadherin-GFP- expressing embryo.
(MOV)
Time-lapse showing AS of stage 15 wild-type embryo in which GFP had been expressed with the Gal4NP3312 AS driver, showing morphogenesis of the AS into a narrow, tube-like structure.
(MOV)
Time-lapse showing AS of stage 15 embryo in which Egfr-EGFP and GFP-NLS had been expressed with the double driver combination Gal4NP3312 + Gal4NP5328 showing failure of AS morphogenesis.
(MOV)
Time-lapse showing AS of stage 15 embryo in which Egfr-EGFP and GFP-NLS had been expressed with the double driver combination Gal4NP3312 + Gal4NP5328 showing failure of AS morphogenesis.
(MOV)