

Abstract
A novel non-invasive technique was applied to measure velocity within slow conducting axons in the distal extreme of the sciatic nerve (i.e., digital nerve) in a rat model. The technique is based on the extraction of rectified multiple unit activity (MUA) from in vivo whole nerve compound responses. This method reliably identifies compound action potentials in thinly myelinated fibers conducting at a range of 9-18 m/s (Aδ axons), as well as in a subgroup of unmylinated C fibers conducting at approximately 1-2 m/s. The sensitivity of the method to C-fiber conduction was confirmed by the progressive decrement of the responses in the 1-2 m/s range over a 20-day period following the topical application of capsaicin (ANOVA p<0.03). Increasing the frequency of applied repetitive stimulation over a range of 0.75 Hz to 6.0 Hz produced slowing of conduction and a significant decrease in the magnitude of the compound C-fiber response (ANOVA p<0.01). This technique offers a unique opportunity for the non-invasive, repeatable, and quantitative assessment of velocity in the subsets of Aδ and C fibers in parallel with evaluation of fast nerve conduction.
Keywords: C fibers, Aδ fibers, small fiber neuropathy, nerve conduction velocity, digital nerve, capsaicin
1. INTRODUCTION
The evaluation of peripheral nerve conduction velocity (NCV)1 is a well-established and highly-utilized procedure for the assessment of the pattern, symmetry and extent of neuropathy in both pre-clinical and clinical studies [1-4]. When properly performed, NCV provides a sensitive, objective, and specific index of the onset and progression of multiple forms of nerve damage. C fibers make up the majority of fibers in some nerve segments, however, standard NCV measures are only sensitive to activity in fast conducting, heavily myelinated axons. The in vivo assessment of conduction in small diameter axons is a formidable task due to the requirement of high threshold stimulation, asynchronous, slow conduction and phase cancellation with distance [5]. The inability to assess conduction in thinly myelinated Aδ axons or in unmyelinated C fibers is a severe limitation of NCV procedures. There is a growing recognition that many forms of idiopathic small fiber neuropathies (SFN) may have been poorly diagnosed and their prevalence underestimated due, in part, to a lack of sensitive, non-invasive measures of this condition [5-10]. SFN can result in pain, paresthesias, abnormal thermal sensitivity, and various autonomic dysfunctions. Emerging data suggests that SFNs have unique etiologies, distinct genetic contributions and they potentially differ in their response to therapeutic approaches [11-24].
The clinical diagnosis of SFN is generally based on symptoms or the presence of specific sensory deficits revealed by questionnaires and/or physical examination (e.g., diminished perception of “sharp”). These measures are subjective and often highly variable. The addition of epidermal fiber density has added sensitivity to the assessment of SFNs, however, the interpretation of these findings is often limited by wide ranges of “normal values” and differences in fiber counts as a function of location [25-37].
The situation is even more difficult in preclinical models of SFN, which generally are forced to rely on behavioral assessment as a measure of small fiber dysfunction [38-45]. The behaviors in question (e.g., thermal withdrawal reaction) are often complex, they involve both CNS and PNS functions, and they can be influenced by learning and motivation. Although neuropathologic evaluation can be applied to animal models, the accurate evaluation of structural deficits in C fibers requires invasive, time consuming, teased fiber techniques [11, 14, 46, 47]. Epidermal fiber density measures have also been applied to preclinical studies, but the sensitivity of the procedures has often been limited by the same high variability issues seen in the clinic [48-52]. Single unit recording provides the most complete and specific evaluation of the function of slow conducting fibers. These studies are critical for defining cellular and molecular mechanisms of SFN, but they require complicated, invasive procedures often restricted to the proximal segments of the nerves, and have limited applications in assessment of the disease progression and/or in screening of potential therapeutics [53-63]. Microneurographic studies have reported that painful SFNs are often associated with an increase in spontaneous activity and changes in excitability of C fibers [64-67]. The introduction of microneurography to animal modeling confirmed similar patterns of C-fiber activity in rats and humans in both normal and pathological conditions; however, the accurate tracing of C-fiber activity in animal models required exposing the sciatic nerve, precluding longitudinal assessments [68,69].
The present study was designed to define a reliable experimental technique for the assessment of slowly conducting axons. The approach differs from the well-established use of single axon recording in several important ways: 1) it assesses whole nerve compound responses, allowing quantitative evaluation of the effects of experimental treatment or disease on both fast and slow conducting axons in parallel; 2) it is non-invasive, permitting repeat measurement of the slow nerve conduction as deficits progress and possibly recover; 3) it is a relatively simple procedure that could be implemented, in most cases, with existing electrophysiologic equipment. Application of this technique could augment the specificity of microneurography with longitudinal and quantitative assessment of activity in the whole intact nerve.
2. MATERIALS AND METHODS
2.1. Animals
Electrophysiological studies were performed on 15 Sprague-Dawley female rats (Charles River Lab., Wilmington, MA) that were 5-6 months old at the start of the study. All animals were given free access to standard laboratory chow and water throughout the study. Five rats were treated with the topical application of capsaicin (CAP). The magnitude of C-fiber conduction in these rats were compared to that determined in 5 age-matched control rats at baseline (prior to the treatment),10, 20, and 30-45 days after the administration of CAP. The remaining rats (N=5) were used to help optimize the recording procedures and analysis. The use of all animals and procedures were in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and approved by the Animal Institute Care and Use Committee at Albert Einstein College of Medicine.
2.2. Electrophysiological procedures
All experiments were done under general anesthesia (Isoflurane 1.5-2.5%/O2), with the rats positioned on a heating pad and monitored for respiration and rectal temperature. Evoked activity was recorded in the digital nerve using subcutaneous platinum needle electrodes (Grass Astro-Med, Inc. Warnick, RI) for both recording and stimulation. The digital nerve was stimulated at the dorsal-superficial surface of 3rd cuneiform/proximal end of metatarsal (Fig. 1, S1). A distal set of recording electrodes was positioned on the lateral side of 3rd toe (Fig. 1, R1) 20 mm distal from the stimulation point. Another set of recording electrodes was positioned at the sciatic notch (75-85 mm proximally to the stimulation point) to monitor nerve conduction over the proximal segment of the nerve (Fig. 1, R2). After evaluation of the proximal nerve conduction, percutaneous injection of 0.1-0.3 mL of 2% Lidocaine (SPARHAWK, Laboratories, Inc., Lenexa, KS) was applied between the recording sites to block nerve conduction [70] and prevent the masking of slow conducting responses by central reflexes (e.g., dorsal root reflexes, 71). The effective blocking of conduction in fast fibers was verified by the absence of neural response at the proximal site and monitored throughout the experiment. Responses were evoked by constant current electrical stimulation (Grass stimulator -11, isolation unit SIU-7, Warnick, RI), which was systematically increased in intensity (0.1-15 mA and duration 0.01-3.0 ms) until the magnitude of the C-fiber signal (i.e., velocities < 2.0 m/s) was maximal.
Figure 1.
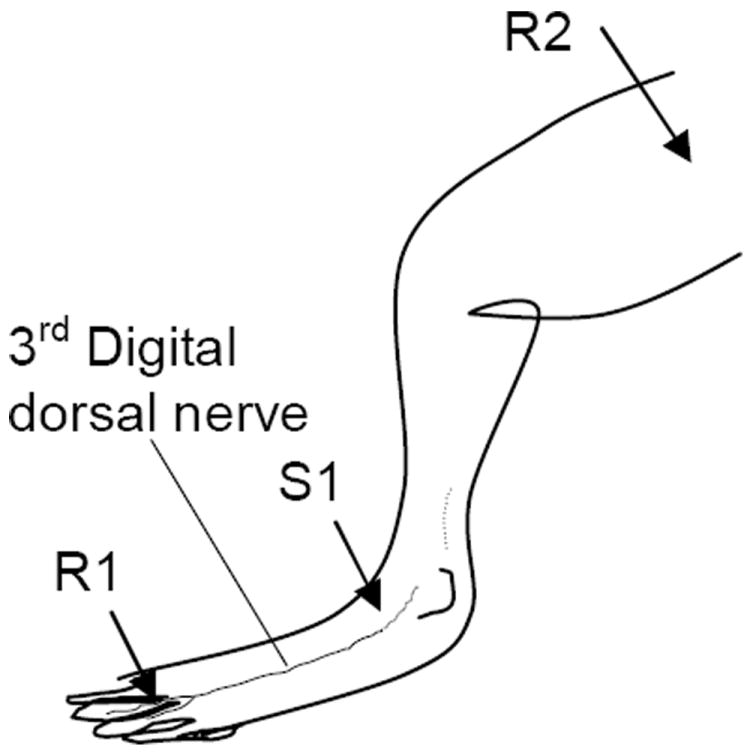
The schematic of electrode positions for electrophysiological recording over distal (R1) and proximal (R2) segments of the digital nerve with respect to stimulation (S1).
The effects of high frequency repetitive stimulation (HFRS) on C-fiber activity were explored using stimulation rates of 0.75, 1.5, 3.0 and 6.0 Hz. The use of this procedure has been previously described for the evaluation of fast conducting fibers [72]. HFRS was conducted in step-like order from the lowest (0.75 Hz) to the highest (6.0 Hz) frequency. Prior to each escalation in frequency, a 0.75 Hz signal was evaluated to test the return of the slow fiber neural activity to the baseline level. Neuroelectric signals were impedance-matched and differentially amplified with a gain of 20,000 and a frequency band of 20 Hz-3 kHz using a BIOPAC MP100 and AcqKnowledge software version 3.7.2 (BIOPAC Systems Inc., Goleta, CA). Data were scanned for artifacts, digitized at the rate of 20 kHz and averaged across 50-100 stimuli for slow nerve conduction, and 5-20 stimuli for the fast nerve conduction. Single sweeps of slow nerve conduction were stored in a digital format (MDI DT-1600) for further off-line analysis.
2.3. Multiple Unit Activity (MUA)
Measures of MUA were extracted from single sweep recording by first high-pass filtering the response above 450 Hz, then full-wave rectifying the signal prior to averaging [73-75]. Rectification minimizes phase cancellation of the asynchronous neural signals of slow conduction during averaging. MUA data represent composite voltages of the averaged whole nerve responses digitized at a constant sampling rate (i.e., voltage per time bin). At the specific latencies over a fixed distance from the site of stimulation, these responses were integrated across a specific time period to reflect the signal strength in a specific range of velocities (i.e., 1 to 2 m/s). To balance the difference between the time periods of faster and slower velocities (e.g., 3 ms for 1.5-2 m/s and 20 ms for 0.5-1 m/s), the response integrated over each range of NCV was divided by the number of time bins (voltage samplings) within this range. The integrated MUA signal was expressed as a magnitude of response over the range of 0.5-2 m/s with respect to the level of spontaneous activity in the pre-stimulation epoch [76-77].
2.4. Application of Capsaicin
Local skin denervation was induced by topical application of 0.1% CAP crème (Capzasin-HP, Chattem, Inc., Chattanooga, TN) to the 3rd toe of rat hind paw. CAP was applied under continuous light isoflurane anesthesia (0.5-1.0 % in 0.3 L/min flow of O2) for 10 hours/day over a four-day period. This procedure minimized discomfort of the animal during CAP application and assured consistency of treatment across animals. After each daily application, the residual crème was cleaned from the skin. The first post-CAP electrophysiologic measures occurred 10 days after discontinuation of CAP treatment; at that time there was no evidence of inflammation or skin redness.
2.5. Statistics
Comparisons across conditions utilized standard inferential procedures (ie., ANOVA); alpha levels were set at 0.05. If warranted, a Tukey’s Multiple Comparison Test was used for cross-sectional evaluations. Experimental data are presented as mean ± SD, unless otherwise indicated.
3. RESULTS
3.1. Distal Digital Nerve Electrophysiology
The patterns of digital nerve activity varied as a function of stimulus intensity and distal-to-proximal recording site. The lowest intensity evaluated (5 mA - 0.01 ms) uniquely activated low threshold axons and resulted in a composite response dominated by velocities over 55 m/sec in the proximal nerve segments and between 26-42 m/s at the distal segments (Fig. 2A). Increasing the stimulation intensity (10-15 mA - 0.01-0.05 ms), added the activation of slower conduction fibers with velocities less than 20 m/sec (semi-late responses, Fig. 2A). In the distal segment of digital nerve, maximal NCV was 35.1± 6.1 m/s with an amplitude of 11.6± 4.4 μV, the NCV of semi-late responses was evaluated by peak latency and varied between 9.0-18.2 m/s (first peak mean NCV -16.3± 1.6 m/s). Simultaneously recorded neural response at the sciatic notch (proximal segment) demonstrated maximal NCV of 59.1± 4.2 m/s reflecting conduction in Aαβ fibers. These measurements are in agreement with previously published evaluations of maximal NCV and single unit recordings [62,78-80] as well as histological evaluations of the morphometric fiber composition of the digital nerve as a distal branch of superficial peroneal nerve [81].
Figure 2.
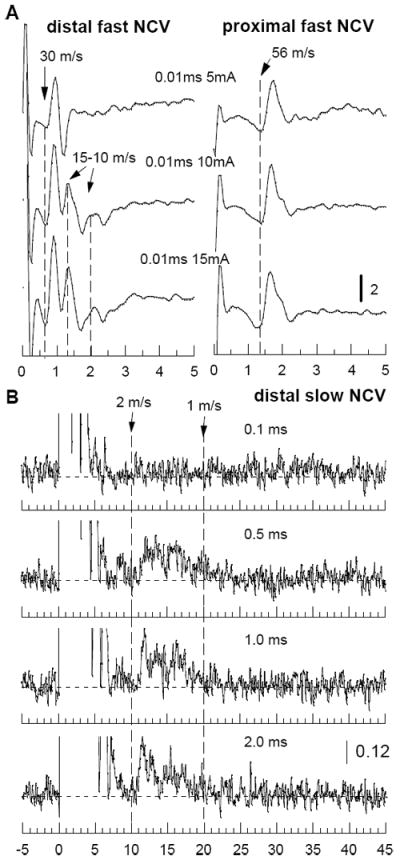
Representative traces of evoked potentials (A) and MUA (B) of a digital nerve are displayed as a function of intensity of stimulation. Numbers in the middle represent duration of the stimulus (ms) and intensity of constant current (mA). In B, all recordings were done at constant current of 15 mA, response of fast conduction is left out of scale (initial 5 ms after start of stimulation). Here and on the following figures, timescale is given in milliseconds and calibration is given in microvolts. Vertical dash lines indicate latencies for the marked NCV (m/s). Horizontal dash lines denote level of activity in prestimuli interval.
The activation of C fibers with associated conduction velocities less than 2.0 m/s required the use of high intensity stimulation (15 mA - 0.5 -2 ms, Fig. 2B). These stimulation currents and durations produced artifactual contamination of the earliest responses and therefore the analysis of C-fiber activity was limited to the later MUA signals. The maximum magnitude of the slow NCV was observed in the range of 1-2 m/s, with the peak of activity in a range of 1.5-1.7 m/s (Fig. 3). As expected, C-fiber responses extracted from whole nerve recordings were small and variable across animals (Fig. 3A). Data recorded at a fixed distance from the stimulating cathode were averaged across subjects to improve the signal/noise ratio for the low amplitude, slow conducting responses. MUA was integrated across fixed time windows corresponding to velocities in the range of 2.0 to 1.6 m/s, 1.5 to 1.1 m/s and 1.0 to 0.5 m/s and compared to the strength of the signal prior to stimulation. As a result, each compound C-fiber response was evaluated for a total magnitude of activity and its distribution in the range of velocities from 0.5 to 2 m/s (Fig. 3B and C).
Figure 3.
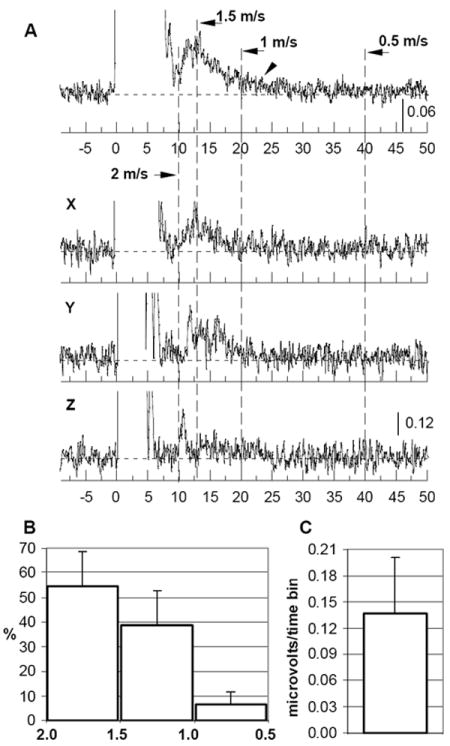
Compound C-fiber response is presented as MUA in digital nerve averaged across 15 animals (A, top trace) and compared to the samples from three rats (X, Y, Z). Response in the range of fast conduction is left out of scale (initial 5-7 ms after start of stimulation). Bar graphs illustrate distribution of activity within three ranges of slow NCV (horizontal axis, B) as a percent of the total response, and magnitude of the C-fiber compound response (C) within the range of NCV 0.5-2 m/s (mean, SD).
3.2. Effects of Topical CAP
The topical application of CAP resulted in a progressive decrease in the magnitude of the C-fiber compound response in the digital nerve over first 20 days after the end of treatment. Figure 4 illustrates the effects of CAP-induced denervation for responses with velocities in the C-fiber range. The effects of CAP are clearly seen in MUA averaged across 5 rats at four time points (Fig. 4A). Evaluation of total activity of C fibers in the range of slow NCV (0.5-2 m/s) across time points confirmed a statistically significant decrease in activity (Fig. 4B, ANOVA p<0.03). In addition to a decrease in total magnitude of the response, C fibers also demonstrated re-distribution of MUA with a shift towards a slower range of velocities at the early stage of denervation. Untreated age-matched controls demonstrated no change in the same measures of C-fiber activity. The recovery of slow NCV was observed in a period of 30-45 days after drug application.
Figure 4.
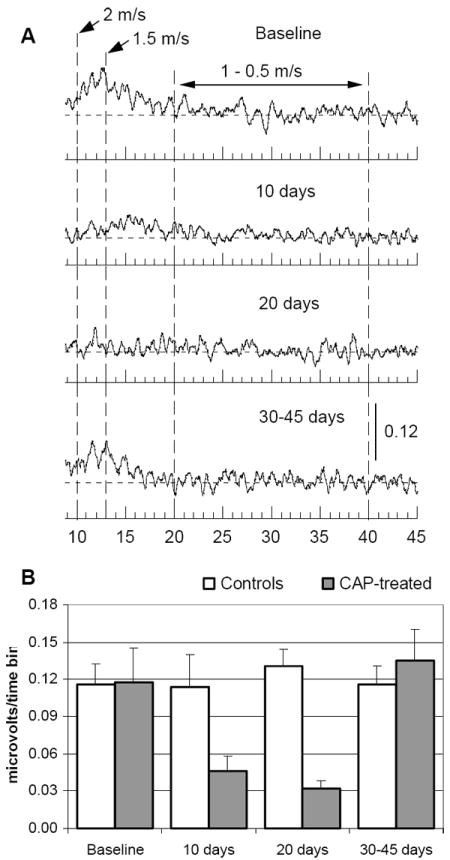
Effects of CAP application are reflected in the magnitude of compound C-fiber response in the digital nerve. A - MUA was recorded prior to application of CAP (Baseline), and 10, 20 and 30-45 days after application and averaged across 5 rats. B – a decrease in total magnitude of the C-fiber compound response within the range of NCV 0.5-2 m/s after treatment with CAP (ANOVA p< 0.03, closed bars, mean, SEM, N=5) is compared to no change of the same measure in age-matched controls (open bars, mean, SEM, N=5). For illustration purposes averaged MUA was smoothed.
3.3. Effects of HFRS
Conduction in C-fiber activity was clearly altered by increasing frequency rates. The initial decline in the magnitude of the C-fiber activity was evident with a HFRS of 1.5 Hz; at a rate of 6.0 Hz the C-fiber response had been reduced by approximately 50% (ANOVA p<0.01) compared to responses to low rates of stimulation (Fig. 5A). In parallel with changes in magnitude, the distribution of velocities in the C-fiber range was also shifted to slower response as a function of increase stimulation frequency (Fig. 5C). Surprisingly, C-fiber response recorded at the frequency of 0.75 Hz right after HFRS demonstrated minimal or no decline in total MUA (Fig. 5B, 0 min), while a recording after a 5-min rest period showed a significant reduction in magnitude (Fig. 5B, 5 min, ANOVA p<0.01). This paradox appears more prominent at the highest rates of stimulation (e.g., 3 and 6 Hz), and is most likely related to the slowing of Aδ fibers into the range of C-fiber activity under the pressure of the elevated rates of stimulation. Immediately after termination of HFRS, the responses of some Aδ fibers may still be delayed and cause an increase in compound response within the range of C-fiber conduction, while after 5 min rest, they recover and shift back to the shorter latencies, leaving the compound C-fiber response at the level of the magnitude comparable to one during HFRS. Figure 5D provides an example of changes in representative traces of the distal Aβ and Aδ responses under conditions of HFRS. The slowest subset of Aδ fibers (NCV range of 3-4 m/s) diminished the amplitude of responses during HFRS of 3-6 Hz and, therefore, may slow down into the range of conduction less than 2 m/s. As expected, population of faster Aδ fibers (e.g., NCV range of 7-12 m/s) demonstrated better resistance to the elevation of frequency and only slightly slowed conduction at the frequency of 6 Hz, while response in Aβ fibers (peak NCV – 17 m/s) did not change at any frequencies applied.
Figure 5.
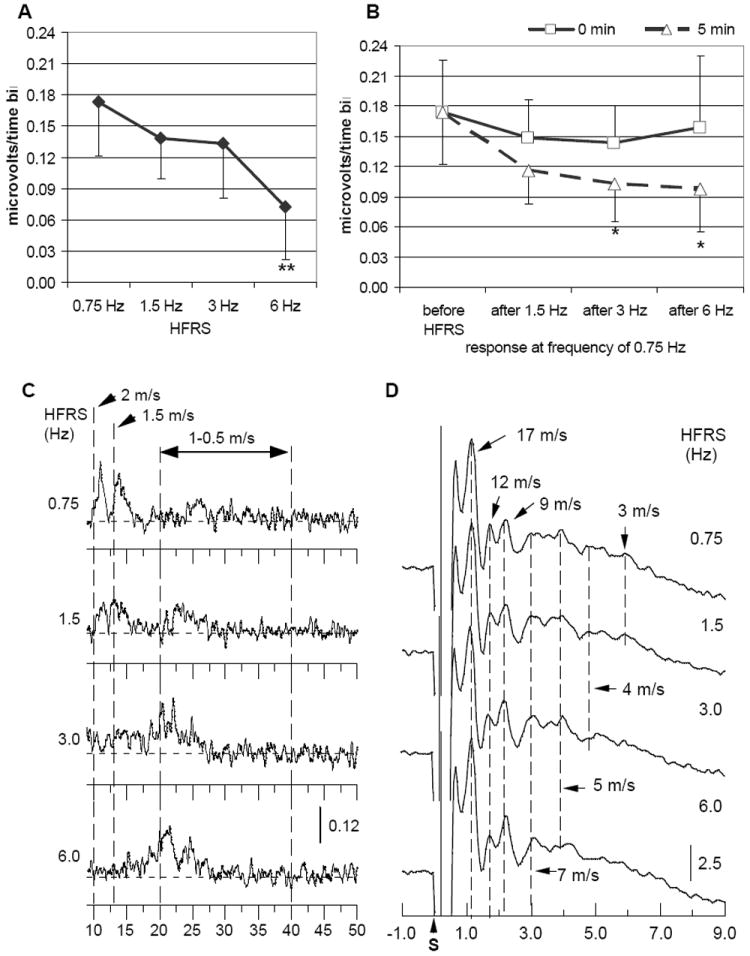
Changes in distal slow conduction under condition of HFRS: A and B - total magnitude of C-fiber compound response (mean, SD, N=8) during (A) and after HFRS (B, 0 min -solid line, 5 min -dashed line); C and D demonstrate changes in representative traces of C-fiber compound response (C, MUA) and in Aβ-Aδ evoked potential (D) under conditions of HFRS. “S” next to the time scale marks start of stimulation. Asterisks denote statistically significant difference (**-p<0.01, * - p<0.05 between the magnitude of the initial response to frequency 0.75 Hz and responses during and after HFRS.
4. DISCUSSION
The application of MUA techniques to in vivo electrophysiological measures provides an opportunity to measure the magnitude and range of NCV in C fibers in parallel with the assessment of conduction in the faster Aβ and Aδ fibers. This approach can identify velocities less than 2 m/s, and can be applied repeatedly to the vulnerable distal segments of specific nerves. In the present study, up to 85% of C-fiber activity was recorded in the range of NCV between 1 and 2 m/s. The results are comparable to those observed in microneurographic recordings in humans [66, 82-84]. However, single unit studies, especially with the use of teased fiber preparation, can identify C-fiber velocities less than 1 m/s [54, 56, 60, 62, 63, 78, 85]. In the non-invasive, whole nerve approach used in the present study, the temporal dispersion and phase cancellation of activity in the slowest subset of the C fibers may simply be too great for the summed response to significantly exceed prestimulus baseline levels. In addition, the use of stimulation of 0.75 Hz and above in the present study may be too rapid for activation of the slowest fibers, which in the single fiber studies are driven at the rate of 0.25-0.5 Hz [55, 63, 68, 83]. The increased sensitivity to activity in the subset of neurons conducting slower than 1 m/s in the single fiber techniques is gained at the expense of the sampling of a limited number of axons and/or the need for mechanical manipulation of the tissue (e.g., teased fiber preparation) which is generally performed at the more proximal nerve segments.
CAP selectively affects distal endings of Aδ and C fibers [86]. Prolonged topical application of CAP leads to local skin desensitization and depletion of peripheral endings of small diameter fibers evident by immunohistochemical markers [87-92]. We employed CAP to confirm that MUA in the range of slow NCV originates from the activity in C fibers and to explore the ability of new electrophysiological evaluation to register small fiber degeneration and regeneration. The compound C-fiber response was significantly reduced during the first 20 days after CAP application and recovered in 30-45 days. These functional measures were consistent with the results observed in clinic for the same population of axons evaluated by epidermal and sub-epidermal nerve fiber density (NFD) and quantitative sensory testing [91, 92]. In humans, a reduction of NFD in the skin was observed in 2-7 days after topical CAP application, while initial re-innervation of CAP-treated skin was marked between 20-40 days after application [92]. The longer time period needed for electrophysiological registration of denervations is most likely related to the greater extent of axonal degeneration which is required for elimination of nerve compound response compared to reduction of density of nerve endings in the skin and subcutaneous tissue. At the early stage of degeneration (10 days), C fibers demonstrated a re-distribution of MUA toward a slower range NCV, in addition to a decrease in total magnitude of the response. Similar signs of dysfunction were observed in distal diabetic autonomic neuropathy [77].
Assessment of activity-dependent changes is an important experimental tool for characterization of C-fiber function. While fast conducting fibers can easily follow frequencies up to 100 Hz, slow conducting fibers are known for significant limitations in ability to respond to rapid stimulation [53, 54, 57, 61, 63, 64, 66, 72, 82-85, 93-97]. Using repetitive stimulation, conduction of C fibers is altered at rates of 1 Hz and, in some circumstances, blocked at frequencies greater than 2 Hz [82, 93, 98]. In the present study, HFRS altered C-fiber compound response in two ways: higher stimulation rates reduced the overall response magnitude of the C-fiber response and shifted the distribution of NCV to slower values. The initial drop in magnitude of the compound response was observed at 1.5 Hz, while stimulation at 3 Hz increased the slowing of conduction. At a stimulation rate of 6 Hz, the C-fiber response was reduced by 60% compared to responses at the rate of 0.75 Hz, and the range of NCV has shifted from 1.5-2 m/s to less than 1.5 m/s.
The complex effects of stimulation rates in the present study support the contribution of a variety of unmyelinated fiber types to the compound C-fiber response. Single unit recordings confirm that high rates of repetitive stimulation are associated with progressive slowing, partial failure of conduction and an increase of temporal dispersion in C fibers [68, 82, 93, 98-100]. The nature and severity of these effects also depend on the class/modality of activated C fibers [54, 85, 100]. For instance, mechano-insensitive C fibers are very sensitive to stimulation rates, while sympathetic efferent fibers demonstrated minimal changes in conduction under the same conditions. When measured as part of a composite whole nerve response, effects on the various sub-types of C fibers can create complex patterns reflecting both slowing (e.g., re-distribution of NCV) and blocking of slow conduction (e.g., reduction in magnitude of response), as it was observed in the compound responses of the current study.
Another difficulty in the assessment of slowly conducting axons is the possible interaction of activity from overlapping populations of the slowest subset of Aδ fibers and the fastest of the C fibers. With greater distances, these velocities would be clearly segregated, but when the examining patterns are isolated to the distal extremes of whole nerve over a few centimeters, activity in these populations can be complex. Aδ fibers demonstrate slowing and partial failure of conduction at much lower frequencies than Aα and Aβ fibers; activity in these fibers come very close to the margins of C-fiber conduction in skin innervation [78, 93, 101, 102]. The sub-components of the late responses can be explored using different time periods after HFRS. Relatively fast conducting sub-populations demonstrate minimal suppression following HFRS, while activity in slower C fibers can continue to be diminished for a period of up to 5-min post-stimulation. Under the pressure of HFRS, some Aδ fibers may slow down to the C-fiber range of velocities [101], however, these larger diameter axons can be identified by their faster rate of recovery.
4.1. Advantages and limitations of the novel approach
The advantages of the use of non-invasive measures of C-fiber activity are obvious. The outlined technique can reliably quantify activity in axons conducting at 1-2 m/s, the measures are relatively rapid, they can be repeated in the same nerve segment in longitudinal studies and the procedure can measure conduction in fast and slow axons in the same experiment. As expected, the compound measure of slow conduction is sensitive to the effects of CAP and subcomponents of slow conducting fibers can be differentiated by their response to stimulation rate and their fatigue. In contrast to the single unit assessments, the obtained results reflect the activity of the population of C fibers within intact nerve. The measures are parametric and subject to statistical comparison across time and experimental conditions. The limitations of this technique are also clear. The activation of high-threshold C fibers requires intense stimulation which may limit this technique to anesthetized preparation. The composite nature of the C-fiber response confounds the identification of activity in specific sub-populations, although this can be facilitated by use of HFRS and recovery profiles. As measured in this study, the compound response is insensitive to conduction below 1 m/s, but this may be altered by the use of lower stimulation rates and a higher resolution averaging.
The relatively simple evaluation of C-fiber conduction outlined in the study offers a novel tool for examining unique electrophysiological aspects of SFN and may complement structural measures of skin biopsy with functional evaluation of deficits in transmembrane ion distribution, nodal integrity and energy utilization (mitochondrial function) acquired from the whole nerve response of small diameter nerve fibers. The longitudinal measure of conduction in Aδ and C fibers will bridge the gap between discoveries on the axonal level and their preclinical evaluation by gross behavior (e.g., withdrawal reaction).
Acknowledgments
The authors would like to thank Shirley Seto and Linda O’Donnell for their assistance in preparation of the manuscript. This work was supported by NIH grant R21 RR025896.
Footnotes
CAP-capsaicin, HFRS-high frequency repetitive stimulation, MUA-multiple unit activity, NCV-nerve conduction velocity, NFD-nerve fiber density, SFN-small fiber neuropathy.
Conflict of interest statement
The authors report no conflict of interest.
Contributor Information
Elena G. Zotova, Email: elena.zotova@einstein.yu.edu.
Joseph C. Arezzo, Email: joseph.arezzo@einstein.yu.edu.
References
- 1.Arezzo JC. New developments in the diagnosis of diabetic neuropathy. Am J Med. 1999;107:9S–16S. doi: 10.1016/s0002-9343(99)00008-x. [DOI] [PubMed] [Google Scholar]
- 2.Boyraz O, Saracoglu M. The effect of obesity on the assessment of diabetic peripheral neuropathy: a comparison of Michigan patient version test and Michigan physical assessment. Diabetes Res Clin Pract. 2010;90:256–60. doi: 10.1016/j.diabres.2010.09.014. [DOI] [PubMed] [Google Scholar]
- 3.Cavaletti G, Marmiroli P. Chemotherapy-induced peripheral neurotoxicity. Nat Rev Neurol. 2010;6:657–66. doi: 10.1038/nrneurol.2010.160. [DOI] [PubMed] [Google Scholar]
- 4.Dyck PJ, Albers JW, Andersen H, Arezzo JC, Biessels GJ, Bril V, Feldman EL, Litchy WJ, O’Brien PC, Russell JW on behalf of the Toronto Expert Panel on Diabetic Neuropathy. Diabetic Polyneuropathies: Update on Research Definition, Diagnostic Criteria and Estimation of Severity. Diabetes Metab Res Rev. 2011 Jun 21; doi: 10.1002/dmrr.1226. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 5.Arezzo JC, Zotova E. Electrophysiologic measures of diabetic neuropathy: mechanism and meaning. In: Tomlinson D, editor. Neurobiology of Diabetic Neuropathy. Academic Press; Amsterdam: 2002. pp. 230–55. [DOI] [PubMed] [Google Scholar]
- 6.Lacomis D. Small-fiber neuropathy. Muscle Nerve. 2002;26:173–88. doi: 10.1002/mus.10181. [DOI] [PubMed] [Google Scholar]
- 7.Fink E, Oaklander AL. Small-fiber neuropathy: answering the burning questions. Sci Aging Knowledge Environ. 2006;6:7. doi: 10.1126/sageke.2006.6.pe7. [DOI] [PubMed] [Google Scholar]
- 8.Feldman EL, Cornblath DR, Porter J, Dworkin R, Scherer S. National Institute of Neurological Disorders and Stroke (NINDS): advances in understanding and treating neuropathy. J Peripher Nerv Syst; Attendees of the NIH Peripheral Neuropathy Conference; 24-25 October 2006; Bethesda, Maryland. 2008. pp. 1–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Arezzo JC, Litwak MS, Zotova EG. Correlation and dissociation of electrophysiology and histopathology in the assessment of toxic neuropathy. Toxicol Pathol. 2011;39:46–51. doi: 10.1177/0192623310390231. [DOI] [PubMed] [Google Scholar]
- 10.So Y. New insights into small fiber neuropathy. Ann Neurol. 2012;71:3–4. doi: 10.1002/ana.22666. [DOI] [PubMed] [Google Scholar]
- 11.Yagihashi S, Kamijo M, Ido Y, Mirrlees DJ. Effects of long-term aldose reductase inhibition on development of experimental diabetic neuropathy. Ultrastructural and morphometric studies of sural nerve in streptozocin-induced diabetic rats. Diabetes. 1990;39:690–6. doi: 10.2337/diab.39.6.690. [DOI] [PubMed] [Google Scholar]
- 12.Boulton AJ, Vinik AI, Arezzo JC, Bril V, Feldman EL, Freeman R, Malik RA, Maser RE, Sosenko JM, Ziegler D American Diabetes Association. Diabetic neuropathies: a statement by the American Diabetes Association. Diabetes Care. 2005;28:956–62. doi: 10.2337/diacare.28.4.956. [DOI] [PubMed] [Google Scholar]
- 13.Flatters SJ, Bennett GJ. Studies of peripheral sensory nerves in paclitaxel-induced painful peripheral neuropathy: evidence for mitochondrial dysfunction. Pain. 2006;122:245–57. doi: 10.1016/j.pain.2006.01.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Keswani SC, Jack C, Zhou C, Höke A. Establishment of a rodent model of HIV-associated sensory neuropathy. J Neurosci. 2006;26:10299–304. doi: 10.1523/JNEUROSCI.3135-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lee JJ, Low JA, Croarkin E, Parks R, Berman AW, Mannan N, Steinberg SM, Swain SM. Changes in neurologic function tests may predict neurotoxicity caused by ixabepilone. J Clin Oncol. 2006;24:2084–91. doi: 10.1200/JCO.2005.04.2820. [DOI] [PubMed] [Google Scholar]
- 16.Herman RM, Brower JB, Stoddard DG, Casano AR, Targovnik JH, Herman JH, Tearse P. Prevalence of somatic small fiber neuropathy in obesity. Int J Obes (Lond) 2007;31:226–35. doi: 10.1038/sj.ijo.0803418. [DOI] [PubMed] [Google Scholar]
- 17.Vinik AI, Ziegler D. Diabetic cardiovascular autonomic neuropathy. Circulation. 2007;115:387–97. doi: 10.1161/CIRCULATIONAHA.106.634949. [DOI] [PubMed] [Google Scholar]
- 18.Gonzalez-Duarte A, Robinson-Papp J, Simpson DM. Diagnosis and management of HIV- associated neuropathy. Neurol Clin. 2008;26:821–32. doi: 10.1016/j.ncl.2008.04.001. [DOI] [PubMed] [Google Scholar]
- 19.Jin HW, Flatters SJ, Xiao WH, Mulhern HL, Bennett GJ. Prevention of paclitaxel-evoked painful peripheral neuropathy by acetyl-L-carnitine: effects on axonal mitochondria, sensory nerve fiber terminal arbors, and cutaneous Langerhans cells. Exp Neurol. 2008;210:229–37. doi: 10.1016/j.expneurol.2007.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Løseth S, Mellgren SI, Jorde R, Lindal S, Stålberg E. Polyneuropathy in type 1 and type 2 diabetes: comparison of nerve conduction studies, thermal perception thresholds and intraepidermal nerve fibre densities. Diabetes Metab Res Rev. 2010;26:100–6. doi: 10.1002/dmrr.1049. [DOI] [PubMed] [Google Scholar]
- 21.Vivithanaporn P, Heo G, Gamble J, Krentz HB, Hoke A, Gill MJ, Power C. Neurologic disease burden in treated HIV/AIDS predicts survival: a population-based study. Neurology. 2010;75:1150–8. doi: 10.1212/WNL.0b013e3181f4d5bb. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tavee J, Culver D. Sarcoidosis and small-fiber neuropathy. Curr Pain Headache Rep. 2011;15:201–6. doi: 10.1007/s11916-011-0180-8. [DOI] [PubMed] [Google Scholar]
- 23.Xiao WH, Zheng H, Zheng FY, Nuydens R, Meert TF, Bennett GJ. Mitochondrial abnormality in sensory, but not motor, axons in paclitaxel-evoked painful peripheral neuropathy in the rat. Neuroscience. 2011;199:461–9. doi: 10.1016/j.neuroscience.2011.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Xiao WH, Zheng H, Bennett GJ. Characterization of oxaliplatin-induced chronic painful peripheral neuropathy in the rat and comparison with the neuropathy induced by paclitaxel. Neuroscience. 2012;203:194–206. doi: 10.1016/j.neuroscience.2011.12.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Polydefkis M, Hauer P, Griffin JW, McArthur JC. Skin biopsy as a tool to assess distal small fiber innervation in diabetic neuropathy. Diabetes Technol Ther. 2001;3:23–8. doi: 10.1089/152091501750219994. [DOI] [PubMed] [Google Scholar]
- 26.Hoitsma E, Reulen JP, de Baets M, Drent M, Spaans F, Faber CG. Small fiber neuropathy: a common and important clinical disorder. J Neurol Sci. 2004;227:119–30. doi: 10.1016/j.jns.2004.08.012. [DOI] [PubMed] [Google Scholar]
- 27.Horowitz SH. The diagnostic workup of patients with neuropathic pain. Anesthesiol Clin. 2007;25:699–708. doi: 10.1016/j.anclin.2007.07.010. [DOI] [PubMed] [Google Scholar]
- 28.Devigili G, Tugnoli V, Penza P, Camozzi F, Lombardi R, Melli G, Broglio L, Granieri E, Lauria G. The diagnostic criteria for small fibre neuropathy: from symptoms to neuropathology. Brain. 2008;131:1912–25. doi: 10.1093/brain/awn093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Backonja MM, Walk D, Edwards RR, Sehgal N, Moeller-Bertram T, Wasan A, Irving G, Argoff C, Wallace M. Quantitative sensory testing in measurement of neuropathic pain phenomena and other sensory abnormalities. Clin J Pain. 2009;25:641–7. doi: 10.1097/AJP.0b013e3181a68c7e. [DOI] [PubMed] [Google Scholar]
- 30.Nebuchennykh M, Løseth S, Lindal S, Mellgren SI. The value of skin biopsy with recording of intraepidermal nerve fiber density and quantitative sensory testing in the assessment of small fiber involvement in patients with different causes of polyneuropathy. J Neurol. 2009;256:1067–75. doi: 10.1007/s00415-009-5065-y. [DOI] [PubMed] [Google Scholar]
- 31.Walk D, Sehgal N, Moeller-Bertram T, Edwards RR, Wasan A, Wallace M, Irving G, Argoff C, Backonja MM. Quantitative sensory testing and mapping: a review of nonautomated quantitative methods for examination of the patient with neuropathic pain. Clin J Pain. 2009;25:632–40. doi: 10.1097/AJP.0b013e3181a68c64. [DOI] [PubMed] [Google Scholar]
- 32.Cavaletti G, Frigeni B, Lanzani F, Mattavelli L, Susani E, Alberti P, Cortinovis D, Bidoli P. Chemotherapy-Induced Peripheral Neurotoxicity assessment: a critical revision of the currently available tools. Eur J Cancer. 2010;46:479–94. doi: 10.1016/j.ejca.2009.12.008. [DOI] [PubMed] [Google Scholar]
- 33.European Federation of Neurological Societies/Peripheral Nerve Society Guideline on the use of skin biopsy in the diagnosis of small fiber neuropathy. Report of a joint task force of the European Federation of Neurological Societies and the Peripheral Nerve Society. Joint Task Force of the EFNS and the PNS. J Peripher Nerv Syst. 2010;15:79–92. doi: 10.1111/j.1529-8027.2010.00269.x. [DOI] [PubMed] [Google Scholar]
- 34.Hlubocky A, Wellik K, Ross MA, Smith BE, Hoffman-Snyder C, Demaerschalk BM, Wingerchuk DM. Skin biopsy for diagnosis of small fiber neuropathy: a critically appraised topic. Neurologist. 2010;16:61–3. doi: 10.1097/NRL.0b013e3181c9c303. [DOI] [PubMed] [Google Scholar]
- 35.Lauria G, Bakkers M, Schmitz C, Lombardi R, Penza P, Devigili G, Smith AG, Hsieh ST, Mellgren SI, Umapathi T, Ziegler D, Faber CG, Merkies IS. Intraepidermal nerve fiber density at the distal leg: a worldwide normative reference study. J Peripher Nerv Syst. 2010;15:202–7. doi: 10.1111/j.1529-8027.2010.00271.x. [DOI] [PubMed] [Google Scholar]
- 36.Mellgren SI, Lindal S. Nerve biopsy--some comments on procedures and indications. Acta Neurol Scand. 2011;191:64–70. doi: 10.1111/j.1600-0404.2011.01546.x. [DOI] [PubMed] [Google Scholar]
- 37.Mackey S, Carroll I, Emir B, Murphy TK, Whalen E, Dumenci L. Sensory pain qualities in neuropathic pain. J Pain. 2012;13:58–63. doi: 10.1016/j.jpain.2011.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Authier N, Gillet JP, Fialip J, Eschalier A, Coudore F. Description of a short-term Taxol-induced nociceptive neuropathy in rats. Brain Res. 2000;887:239–49. doi: 10.1016/s0006-8993(00)02910-3. [DOI] [PubMed] [Google Scholar]
- 39.Baron R. Peripheral neuropathic pain: from mechanisms to symptoms. Clin J Pain. 2000;16:12–20. doi: 10.1097/00002508-200006001-00004. [DOI] [PubMed] [Google Scholar]
- 40.Gagliese L, Melzack R. Age differences in nociception and pain behaviours in the rat. Neurosci Biobehav Rev. 2000;24:843–54. doi: 10.1016/s0149-7634(00)00041-5. [DOI] [PubMed] [Google Scholar]
- 41.Rice AS, Cimino-Brown D, Eisenach JC, Kontinen VK, Lacroix-Fralish ML, Machin I . Preclinical Pain Consortium. Animal models and the prediction of efficacy in clinical trials of analgesic drugs: a critical appraisal and call for uniform reporting standards. In: Mogil JS, Stöhr T, editors. Pain. Vol. 139. 2008. pp. 243–7. [DOI] [PubMed] [Google Scholar]
- 42.Sullivan KA, Lentz SI, Roberts JL, Jr, Feldman EL. Criteria for creating and assessing mouse models of diabetic neuropathy. Curr Drug Targets. 2008;9:3–13. doi: 10.2174/138945008783431763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Authier N, Balayssac D, Marchand F, Ling B, Zangarelli A, Descoeur J, Coudore F, Bourinet E, Eschalier A. Animal models of chemotherapy-evoked painful peripheral neuropathies. Neurotherapeutics. 2009;6:620–9. doi: 10.1016/j.nurt.2009.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Obrosova IG. Diabetic painful and insensate neuropathy: pathogenesis and potential treatments. Neurotherapeutics. 2009;6:638–47. doi: 10.1016/j.nurt.2009.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sandkühler J. Models and mechanisms of hyperalgesia and allodynia. Physiol Rev. 2009;89:707–58. doi: 10.1152/physrev.00025.2008. [DOI] [PubMed] [Google Scholar]
- 46.Basbaum AI, Gautron M, Jazat F, Mayes M, Guilbaud G. The spectrum of fiber loss in a model of neuropathic pain in the rat: an electron microscopic study. Pain. 1991;47:359–67. doi: 10.1016/0304-3959(91)90229-Q. [DOI] [PubMed] [Google Scholar]
- 47.Sugimoto K, Yagihashi S. Peripheral nerve pathology in rats with streptozotocin-induced insulinoma. Acta Neuropathol. 1996;91:616–23. doi: 10.1007/s004010050475. [DOI] [PubMed] [Google Scholar]
- 48.Lauria G, Lombardi R, Borgna M, Penza P, Bianchi R, Savino C, Canta A, Nicolini G, Marmiroli P, Cavaletti G. Intraepidermal nerve fiber density in rat foot pad: neuropathologic-neurophysiologic correlation. J Peripher Nerv Syst. 2005;10:202–8. doi: 10.1111/j.1085-9489.2005.0010210.x. [DOI] [PubMed] [Google Scholar]
- 49.Siau C, Xiao W, Bennett GJ. Paclitaxel- and vincristine-evoked painful peripheral neuropathies: loss of epidermal innervation and activation of Langerhans cells. Exp Neurol. 2006;201:507–14. doi: 10.1016/j.expneurol.2006.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Beiswenger KK, Calcutt NA, Mizisin AP. Epidermal nerve fiber quantification in the assessment of diabetic neuropathy. Acta Histochem. 2008;110:351–62. doi: 10.1016/j.acthis.2007.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Cheng HT, Dauch JR, Hayes JM, Yanik BM, Feldman EL. Nerve growth factor/p38 signaling increases intraepidermal nerve fiber densities in painful neuropathy of type 2 diabetes. Neurobiol Dis. 2012;45:280–7. doi: 10.1016/j.nbd.2011.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Höke A. Animal Models of Peripheral Neuropathies. Neurotherapeutics. 2012 Mar 14; doi: 10.1007/s13311-012-0116-y. Epub ahead of print. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Shin HC, Lee YL, Kwon HY, Park HJ, Raymond SA. Activity-dependent variations in conduction velocity of C fibers of rat sciatic nerve. Neurosci Res. 1994;19:427–31. doi: 10.1016/0168-0102(94)90084-1. [DOI] [PubMed] [Google Scholar]
- 54.Gee MD, Lynn B, Cotsell B. Activity-dependent slowing of conduction velocity provides a method for identifying different functional classes of C-fibre in the rat saphenous nerve. Neuroscience. 1996;73:667–75. doi: 10.1016/0306-4522(96)00070-x. [DOI] [PubMed] [Google Scholar]
- 55.Bostock H, Campero M, Serra J, Ochoa J. Velocity recovery cycles of C fibres innervating human skin. J Physiol. 2003;553:649–63. doi: 10.1113/jphysiol.2003.046342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Chen X, Levine JD. Altered temporal pattern of mechanically evoked C-fiber activity in a model of diabetic neuropathy in the rat. Neuroscience. 2003;121:1007–15. doi: 10.1016/s0306-4522(03)00486-x. [DOI] [PubMed] [Google Scholar]
- 57.Peng YB, Ringkamp M, Meyer RA, Campbell JN. Fatigue and paradoxical enhancement of heat response in C-fiber nociceptors from cross-modal excitation. J Neurosci. 2003;23:4766–74. doi: 10.1523/JNEUROSCI.23-11-04766.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Tanner KD, Reichling DB, Gear RW, Paul SM, Levine JD. Altered temporal pattern of evoked afferent activity in a rat model of vincristine-induced painful peripheral neuropathy. Neuroscience. 2003;118:809–17. doi: 10.1016/s0306-4522(03)00023-x. [DOI] [PubMed] [Google Scholar]
- 59.Djouhri L, Koutsikou S, Fang X, McMullan S, Lawson SN. Spontaneous pain, both neuropathic and inflammatory, is related to frequency of spontaneous firing in intact C-fiber nociceptors. J Neurosci. 2006;26:1281–92. doi: 10.1523/JNEUROSCI.3388-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Chen X, Levine JD. Mechanically-evoked C-fiber activity in painful alcohol and AIDS therapy neuropathy in the rat. Mol Pain. 2007;3:5. doi: 10.1186/1744-8069-3-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Li BY, Feng B, Tsu HY, Schild JH. Unmyelinated visceral afferents exhibit frequency dependent action potential broadening while myelinated visceral afferents do not. Neurosci Lett. 2007;421:62–6. doi: 10.1016/j.neulet.2007.05.036. [DOI] [PubMed] [Google Scholar]
- 62.Xiao WH, Bennett GJ. Chemotherapy-evoked neuropathic pain: Abnormal spontaneous discharge in A-fiber and C-fiber primary afferent neurons and its suppression by acetyl-L-carnitine. Pain. 2008;135:262–70. doi: 10.1016/j.pain.2007.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Taguchi T, Ota H, Matsuda T, Murase S, Mizumura K. Cutaneous C-fiber nociceptor responses and nociceptive behaviors in aged Sprague-Dawley rats. Pain. 2010;151:771–82. doi: 10.1016/j.pain.2010.09.011. [DOI] [PubMed] [Google Scholar]
- 64.Schmelz M. Translating nociceptive processing into human pain models. Exp Brain Res. 2009;196:173–8. doi: 10.1007/s00221-009-1809-2. [DOI] [PubMed] [Google Scholar]
- 65.Serra J, Solà R, Quiles C, Casanova-Molla J, Pascual V, Bostock H, Valls-Solé J. C-nociceptors sensitized to cold in a patient with small-fiber neuropathy and cold allodynia. Pain. 2009;147:46–53. doi: 10.1016/j.pain.2009.07.028. [DOI] [PubMed] [Google Scholar]
- 66.Ørstavik K, Jørum E. Microneurographic findings of relevance to pain in patients with erythromelalgia and patients with diabetic neuropathy. Neurosci Lett. 2010;470:180–4. doi: 10.1016/j.neulet.2009.05.061. [DOI] [PubMed] [Google Scholar]
- 67.Schmidt R, Kleggetveit IP, Namer B, Helås T, Obreja O, Schmelz M, Jørum E. Double spikes to single electrical stimulation correlates to spontaneous activity of nociceptors in painful neuropathy patients. Pain. 2012;153:391–8. doi: 10.1016/j.pain.2011.10.041. [DOI] [PubMed] [Google Scholar]
- 68.Serra J, Bostock H, Navarro X. Microneurography in rats: a minimally invasive method to record single C-fiber action potentials from peripheral nerves in vivo. Neurosci Lett. 2010;470:168–74. doi: 10.1016/j.neulet.2009.09.061. [DOI] [PubMed] [Google Scholar]
- 69.Serra J, Bostock H, Solà R, Aleu J, García E, Cokic B, Navarro X, Quiles C. Microneurographic identification of spontaneous activity in C-nociceptors in neuropathic pain states in humans and rats. Pain. 2012;153:42–55. doi: 10.1016/j.pain.2011.08.015. [DOI] [PubMed] [Google Scholar]
- 70.Gokin AP, Philip B, Strichartz GR. Preferential block of small myelinated sensory and motor fibers by lidocaine: in vivo electrophysiology in the rat sciatic nerve. Anesthesiology. 2001;95:1441–54. doi: 10.1097/00000542-200112000-00025. [DOI] [PubMed] [Google Scholar]
- 71.Willis WD., Jr Dorsal root potentials and dorsal root reflexes: a double-edged sword. Exp Brain Res. 1999;124:395–421. doi: 10.1007/s002210050637. [DOI] [PubMed] [Google Scholar]
- 72.Zotova EG, Christ GJ, Zhao W, Tar M, Kuppam SD, Arezzo JC. Effects of fidarestat, an aldose reductase inhibitor, on nerve conduction velocity and bladder function in streptozotocin-treated female rats. J Diabetes Complications. 2007;21:187–95. doi: 10.1016/j.jdiacomp.2005.10.001. [DOI] [PubMed] [Google Scholar]
- 73.Legatt AD, Arezzo JC, Vaughan HG., Jr Averaged multiple unit activity as an estimate of phasic changes in local neuronal activity: effects of volume conducted potentials. J Neurosci Methods. 1980;2:203–17. doi: 10.1016/0165-0270(80)90061-8. [DOI] [PubMed] [Google Scholar]
- 74.Arezzo JC, Vaughan HG, Jr, Kraut MA, Steinschneider M, Legatt AD. Intracranial generators of event related potentials in the monkey. In: Cracco RQ, Bodis Wollner I, editors. Frontiers of Clinical Neuroscience Evoked Potentials. Vol. 3. Alan R Liss, Inc.; New York: 1986. pp. 174–89. [Google Scholar]
- 75.Steinschneider M, Fishman YI, Arezzo JC. Spectrotemporal analysis of evoked and induced electroencephalographic responses in primary auditory cortex (A1) of the awake monkey. Cereb Cortex. 2008;18:610–25. doi: 10.1093/cercor/bhm094. [DOI] [PubMed] [Google Scholar]
- 76.Schaumburg HH, Zotova E, Cannella B, Raine CS, Arezzo J, Tar M, Melman A. Structural and functional investigations of the murine cavernosal nerve: a model system for serial spatio-temporal study of autonomic neuropathy. BJU Int. 2007;99:916–24. doi: 10.1111/j.1464-410X.2006.06726.x. [DOI] [PubMed] [Google Scholar]
- 77.Zotova EG, Schaumburg HH, Raine CS, Cannella B, Tar M, Melman A, Arezzo JC. Effects of hyperglycemia on rat cavernous nerve axons: A functional and ultrastructural study. Exp Neurol. 2008;213:439–47. doi: 10.1016/j.expneurol.2008.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lewin GR, McMahon SB. Physiological properties of primary sensory neurons appropriately and inappropriately innervating skin in the adult rat. J Neurophysiol. 1991;66:1205–17. doi: 10.1152/jn.1991.66.4.1205. [DOI] [PubMed] [Google Scholar]
- 79.Li F, Abatan OI, Kim H, Burnett D, Larkin D, Obrosova IG, Stevens MJ. Taurine reverses neurological and neurovascular deficits in Zucker diabetic fatty rats. Neurobiol Dis. 2006;22:669–76. doi: 10.1016/j.nbd.2006.01.012. [DOI] [PubMed] [Google Scholar]
- 80.Schaumburg HH, Zotova E, Raine CS, Tar M, Arezzo J. The rat caudal nerves: a model for experimental neuropathies. J Peripher Nerv Syst. 2010;15:128–39. doi: 10.1111/j.1529-8027.2010.00262.x. [DOI] [PubMed] [Google Scholar]
- 81.Povlsen B, Stankovic N, Danielsson P, Hildebrand C. Fiber composition of the lateral plantar and superficial peroneal nerves in the rat foot. Anat Embryol (Berl) 1994;189:393–9. doi: 10.1007/BF00185434. [DOI] [PubMed] [Google Scholar]
- 82.Serra J, Campero M, Ochoa J, Bostock H. Activity-dependent slowing of conduction differentiates functional subtypes of C fibres innervating human skin. J Physiol. 1999;515:799–811. doi: 10.1111/j.1469-7793.1999.799ab.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Namer B, Barta B, Ørstavik K, Schmidt R, Carr R, Schmelz M, Handwerker HO. Microneurographic assessment of C-fibre function in aged healthy subjects. J Physiol. 2009;587:419–28. doi: 10.1113/jphysiol.2008.162941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Schmelz M, Schmidt R. Microneurographic single-unit recordings to assess receptive properties of afferent human C-fibers. Neurosci Lett. 2010;470:158–61. doi: 10.1016/j.neulet.2009.05.064. [DOI] [PubMed] [Google Scholar]
- 85.Ringkamp M, Johanek LM, Borzan J, Hartke TV, Wu G, Pogatzki-Zahn EM, Campbell JN, Shim B, Schepers RJ, Meyer RA. Conduction properties distinguish unmyelinated sympathetic efferent fibers and unmyelinated primary afferent fibers in the monkey. PLoS One. 2010;5:e9076. doi: 10.1371/journal.pone.0009076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Caterina MJ, Julius D. The vanilloid receptor: a molecular gateway to the pain pathway. Annu Rev Neurosci. 2001;24:487–517. doi: 10.1146/annurev.neuro.24.1.487. [DOI] [PubMed] [Google Scholar]
- 87.McMahon SB, Lewin G, Bloom SR. The consequences of long-term topical capsaicin application in the rat. Pain. 1991;44:301–10. doi: 10.1016/0304-3959(91)90101-3. [DOI] [PubMed] [Google Scholar]
- 88.Simone DA, Ochoa J. Early and late effects of prolonged topical capsaicin on cutaneous sensibility and neurogenic vasodilatation in humans. Pain. 1991;47:285–94. doi: 10.1016/0304-3959(91)90217-L. [DOI] [PubMed] [Google Scholar]
- 89.Nolano M, Simone DA, Wendelschafer-Crabb G, Johnson T, Hazen E, Kennedy WR. Topical capsaicin in humans: parallel loss of epidermal nerve fibers and pain sensation. Pain. 1999;81:135–45. doi: 10.1016/s0304-3959(99)00007-x. [DOI] [PubMed] [Google Scholar]
- 90.Khalili N, Wendelschafer-Crabb G, Kennedy WR, Simone DA. Influence of thermode size for detecting heat pain dysfunction in a capsaicin model of epidermal nerve fiber loss. Pain. 2001;91:241–50. doi: 10.1016/S0304-3959(00)00444-9. [DOI] [PubMed] [Google Scholar]
- 91.Malmberg AB, Mizisin AP, Calcutt NA, von Stein T, Robbins WR, Bley KR. Reduced heat sensitivity and epidermal nerve fiber immunostaining following single applications of a high-concentration capsaicin patch. Pain. 2004;111:360–7. doi: 10.1016/j.pain.2004.07.017. [DOI] [PubMed] [Google Scholar]
- 92.Polydefkis M, Hauer P, Sheth S, Sirdofsky M, Griffin JW, McArthur JC. The time course of epidermal nerve fibre regeneration: studies in normal controls and in people with diabetes, with and without neuropathy. Brain. 2004;127:1606–15. doi: 10.1093/brain/awh175. [DOI] [PubMed] [Google Scholar]
- 93.Torebjörk HE, Hallin RG. Responses in human A and C fibres to repeated electrical intradermal stimulation. J Neurol Neurosurg Psychiatry. 1974;37:653–64. doi: 10.1136/jnnp.37.6.653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Braune H-J. Testing of the refractory period in sensory nerve fibers is the most sensitive method to assess beginning polyneuropathy in diabetics. Electromyogr Clin Neurophysiol. 1999;39:355–9. [PubMed] [Google Scholar]
- 95.Ørstavik K, Namer B, Schmidt R, Schmelz M, Hilliges M, Weidner C, Carr RW, Handwerker H, Jørum E, Torebjörk HE. Abnormal function of C-fibers in patients with diabetic neuropathy. J Neurosci. 2006;26:11287–94. doi: 10.1523/JNEUROSCI.2659-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.De Col R, Messlinger K, Carr RW. Conduction velocity is regulated by sodium channel inactivation in unmyelinated axons innervating the rat cranial meninges. J Physiol. 2008;586:1089–103. doi: 10.1113/jphysiol.2007.145383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.De Col R, Messlinger K, Carr RW. Repetitive activity slows axonal conduction velocity and concomitantly increases mechanical activation threshold in single axons of the rat cranial dura. J Physiol. 2012;590:725–36. doi: 10.1113/jphysiol.2011.220624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Zhu ZR, Tang XW, Wang WT, Ren W, Xing JL, Zhang JR, Duan JH, Wang YY, Jiao X, Hu SJ. Conduction failures in rabbit saphenous nerve unmyelinated fibers. Neurosignals. 2009;17:181–95. doi: 10.1159/000209279. [DOI] [PubMed] [Google Scholar]
- 99.George A, Serra J, Navarro X, Bostock H. Velocity recovery cycles of single C fibres innervating rat skin. J Physiol. 2007;578:213–32. doi: 10.1113/jphysiol.2006.116129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Obreja O, Ringkamp M, Namer B, Forsch E, Klusch A, Rukwied R, Petersen M, Schmelz M. Patterns of activity-dependent conduction velocity changes differentiate classes of unmyelinated mechano-insensitive afferents including cold nociceptors, in pig and in human. Pain. 2010;148:59–69. doi: 10.1016/j.pain.2009.10.006. [DOI] [PubMed] [Google Scholar]
- 101.Lawson SN. Phenotype and function of somatic primary afferent nociceptive neurones with C-, Adelta- or Aalpha/beta-fibres. Exp Physiol. 2002;87:239–44. doi: 10.1113/eph8702350. [DOI] [PubMed] [Google Scholar]
- 102.Djouhri L, Lawson SN. Abeta-fiber nociceptive primary afferent neurons: a review of incidence and properties in relation to other afferent A-fiber neurons in mammals. Brain Res Brain Res Rev. 2004;46:131–45. doi: 10.1016/j.brainresrev.2004.07.015. [DOI] [PubMed] [Google Scholar]