

Abstract
Background
Cerebral blood flow velocity (CBFV) and sleep physiology in healthy children exposed to hypoxia and hypocarbia are under-researched.
Aim
To investigate associations between sleep variables, daytime end-tidal carbon dioxide (EtCO2) and CBFV in children during high-altitude ascent.
Methods
Vital signs, overnight cardiorespiratory sleep studies and transcranial Doppler were undertaken in nine children (aged 6–13 years) at low altitude (130 m), and then at moderate (1300 m) and high (3500 m) altitude during a 5-day ascent.
Results
Daytime (130 m: 98%; 3500 m: 90%, p=0.004) and mean (130 m: 97%, 1300 m: 94%, 3500: 87%, p=0.0005) and minimum (130 m: 92%, 1300 m: 84%, 3500 m: 79%, p=0.0005) overnight pulse oximetry oxyhaemoglobin saturation decreased, and the number of central apnoeas increased at altitude (130 m: 0.2/h, 1300 m: 1.2/h, 3500 m: 3.5/h, p=0.2), correlating inversely with EtCO2 (R2 130 m: 0.78; 3500 m: 0.45). Periodic breathing occurred for median (IQR) 0.0 (0; 0.3)% (130 m) and 0.2 (0; 1.2)% (3500 m) of total sleep time. At 3500 m compared with 130 m, there were increases in middle (MCA) (mean (SD) left 29.2 (42.3)%, p=0.053; right 9.9 (12)%, p=0.037) and anterior cerebral (ACA) (left 65.2 (69)%, p=0.024; right 109 (179)%; p=0.025) but not posterior or basilar CBFV. The right MCA CBFV increase at 3500 m was predicted by baseline CBFV and change in daytime SpO2 and EtCO2 at 3500 m (R2 0.92); these associations were not seen on the left.
Conclusions
This preliminary report suggests that sleep physiology is disturbed in children even with slow ascent to altitude. The regional variations in CBFV and their association with hypoxia and hypocapnia require further investigation.
Keywords: Transcranial Doppler, hypoxia, acute mountain sickness, altitude, Sleep
What is already known on this topic.
Children have less periodic breathing in sleep than adults, after rapid ascent to 3500 m, despite similar mean and minimum overnight oxygen saturations (SpO2) and pCO2.
At sea level, cerebral blood flow velocity (CBFV) is directly dependent on pCO2, but in normocarbic adults, CBFV increases at SpO2<90%.
Increased CBFV is seen in young children with sleep-disordered breathing, and may reverse in association with improved overnight oxygen saturation after adenotonsillectomy.
What this study adds.
Low oxygen saturation (SpO2) is confirmed in sleeping children at altitude, and periodic breathing is related to pCO2.
SpO2 and pCO2 reduction and cerebral blood flow velocity (CBFV) increase at altitude are lower if sea level right middle cerebral artery CBFV is higher, suggesting interaction with hypoxic hyperventilatory response.
In acute hypoxia, cerebral oxygen delivery is maintained by an increase in CBFV anteriorly, but may fall posteriorly.
Introduction
Sleep disordered breathing (SDB) is reported in acclimatised and non-acclimatised adults sleeping at altitude.1 2 Despite frequent exposure and concerns over developmental effects,3 there are limited data from children,2 particularly during slow acclimatisation. Nocturnal breathing is influenced by the delicate chemoreceptor balance affected by sleep stage.4 The central medullary chemoreceptors respond to hydrogen ion concentration [H+], altered by partial pressure of carbon dioxide (pCO2) via the cerebrospinal fluid (CSF) bicarbonate buffering system, and by changing cerebral blood flow (CBF).5 6 The carotid body peripheral chemoreceptors respond to pO2 as well as pCO2 and pH.6 There is a hypoxic ventilatory response (HVR) at pO2 <60 mm Hg, but in place of the normoxic hypercapnic drive, the hyperventilation leads to hypocapnia, reducing the change in pCO2 required to reach the apnoea threshold,7 around which oscillations of pulse oximetry oxyhaemoglobin saturation (SpO2) and pCO2 are associated with periodic breathing.1 8
Transcranial Doppler (TCD) measurement of change in CBF velocities (CBFV) can assess regional perfusion change.9 At sea level, with SpO2>97%, there is a linear relationship between middle cerebral (MCA) and basilar (BA) CBFV and pCO2 between 20 and 60 mm Hg,6 9 10 also mediated via local [H+]. However, although CBFV typically increases when SpO2 is <90%,11 the response to hypoxia is variable between subjects12 13 and within subjects, both regionally12 14 15 and diurnally.16 Technical advances now allow informative portable physiological recording at altitude, overcoming many prior difficulties in studying healthy children.
The purpose of this Smiths Medical Young Everest Study17 was to document natural sleep and daytime CBFV after gradual ascent to 3500 m in comparison with data collected at 130 m, to explore the effects of change in daytime end-tidal (Et) pCO2 and SpO2 and overnight SpO2 on sleep physiology and daytime regional cerebral circulation, and to determine if altitude-induced sleep difficulties would impact on episodic memory recall.18
Methods
Children were recruited from families undertaking a trekking holiday in Nepal17; none had recently travelled to altitudes >1000 m. The study was approved by the UCL research ethics committee (ID:0894/001), and informed written assent/consent obtained from children and their parents. Baseline measurements were undertaken by five specialists in paediatric respiratory and sleep physiology/medicine at ≤130 m in England, and during slow graded ascent (5 days from sea level to 3500 m), planned to maximise safety and minimise acute mountain sickness risk.17 19 Intermediate altitude studies were performed at Kathmandu (1300 m). After flying to Lukla (2670 m), the children trekked for 2 days, sleeping one night at 2650 m and the next at 2835 m, before ascending to Namche Bazaar (3500 m) on the third trek day (day 5 from sea level) where the high-altitude studies were undertaken. With the exception of the sleep studies, recordings were made in a research laboratory setting between 10:00 and 14:00 the day after arrival.17
Sleep studies
Cardiorespiratory sleep studies (LS-100 LifeShirts, Vivometrics, USA) recorded overnight SpO2, ECG, body position, and abdominal and thoracic respiratory effort via inductance plethysmography, but not nasal airflow. Overnight studies were recorded over the first two nights following arrival. Data from the first 6 h of sleep were analysed using paediatric industry standards (eg, 3% oxygen desaturations).
Daytime recordings of vital signs
Daytime SpO2, EtCO2, heart rate (HR), Respiratory Rate (RR) (Tidal Wave 710 Monitor, Respironics, Pennsylvania, USA) and blood pressure were recorded.17
Cerebral blood flow velocity
CBFV was measured by JCG and/or FJK, using TCD (EZ-Dop, DWL, Germany) in the BA and right and left MCA and anterior (ACA) and posterior (PCA) cerebral arteries, each located according to a previously described protocol for which the coefficient of variation was 7%.20 Highest time-averaged maximum mean CBFV (cm/s) was reported over at least six cardiac cycles, analysed blind to subject and altitude. Assuming stable haemoglobin and vessel diameter, proportional change in cerebral oxygen delivery (COD) from baseline at altitude (Alt) compared with sea level (Sea) was calculated using the formula21:
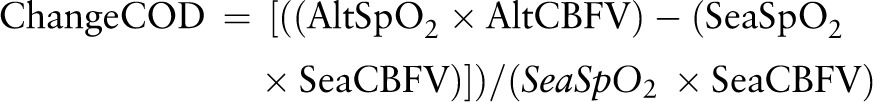 |
Memory
Rivermead Memory Test story recall was used at all three altitudes to determine the ability to recall a short story immediately, and after a delay.22
Statistical analysis
Descriptive statistics are reported at all three altitudes. Variables were examined for normality and non-parametric descriptives and statistical tests selected. Between altitude (130 m, 1300 m, 3500 m) and hemisphere (left, right) comparisons were conducted using paired samples t tests, or Wilcoxon's for two measures, and repeated measures analysis of variance (ANOVA), or Friedman's ANOVA for three. Paired t tests or Wilcoxon signed-rank test and an appropriate Bonferroni correction were used for posthoc testing. Linear regression was used to examine the degree of association for continuous variables. Analysis was conducted using SPSS software for Windows (V.17, SPSS Inc, Illinois, USA).
Results
Participants
Nine children (5 boys) aged 6–13 years (mean age 8.8, SD 2.4), all right-handed and remaining well, were studied.17 Lung function remained within 7% of sea level baseline in all but two.17 All sleep studies at 130 m and 3500 m, but only seven at 1300 m, recorded at least 6 h of high-quality data (table 1).
Table 1.
Vital signs and sleep variables
| Sea level (130 m) | Moderate altitude (1300 m) | High altitude (3500 m) | High altitude %Δ from sea | p Value | |
|---|---|---|---|---|---|
| Mean daytime oxygen saturation (SpO2; %)* | 98 (98, 99) | NA | 90 (87, 91)† | −9.17 (2.1) | 0.004 |
| Mean overnight oxygen saturation (SpO2; %)* ‡ § | 97 (96, 98) | 94 (92, 97)† | 87(85, 88)† ¶ | −11.1 (2.9) | <0.0005 |
| Minimum oxygen saturation (SpO2; %)* ‡ | 92 (89, 93) | 84 (78, 94)* | 79 (76, 80)† ¶ | −14.4 (7.7) | <0.0005 |
| Number of oxygen desaturations per hour* ‡ | 0.2 (0, 1.2) | 4.9 (2, 12)* | 10.0 (4, 22)† ¶ | <0.0005 | |
| Number of central apnoeas per hour* ** | 0.2 (0, 1.2) | 1.2 (0.7, 3.2) | 3.5 (1.1, 5.7)† | 0.2 | |
| Periodic breathing (% TST)* ** | 0.0 (0, 0.3) | 0 (0, 1.2) | 0.2 (0, 1.15)† | 0.5 | |
| Daytime EtCO2 (mmHg) | 42.7 (40, 45) | NA | 36.0 (34, 38)† | −16 (5.1) | 0.004 |
| Daytime respiratory rate (breaths per min) | 21 (20, 24) | NA | 23 (22, 26)† | 8.2 (7.0) | 0.01 |
| Daytime heart rate (bpm) | 78 (72, 85) | NA | 97 (87, 109)† | 26.7 (16.3) | 0.004 |
*For the sleep and respiratory variables, transformation did not redress distributional problems, so data are presented as median and IQR (25th and 75th centiles), and were compared using Wilcoxon signed-rank test where there were no data at mid-altitude, or Friedman's ANOVA where data were available at all three altitudes, with posthoc testing using the Wilcoxon signed-rank test. p Values refer to significant change across all three altitudes.
†Significantly different from 130 m.
‡(η2) for mean overnight oxygen saturation=16.2, for minimum oxygen saturation=13.8 and for number of oxygen desaturations per hour=16.2.
§n=7 at moderate altitude.
¶Significantly different from 1300 m.
**n=3 at moderate altitude, n=8 at high altitude.
EtCO2, end-tidal pCO2; TST, total sleep time; NA, equipment failure.
Respiratory and sleep measures
Mean SpO2 was higher during daytime than overnight at 130 m and 3500 m (table 1). With increasing altitude, daytime and overnight mean SpO2 and EtCO2 decreased (table 1). Daytime RR and HR increased significantly (table 1). Compared with baseline, overnight SpO2 decreased significantly, both in terms of overall mean, and in the minimum reached overnight, while the number of oxygen desaturations increased (table 1). Despite no overall difference in the number of central apnoeas or percentage of total sleep time spent in periodic breathing across the three altitudes, on posthoc testing, there were significant differences between 130 m and 3500 m (table 1). Daytime and mean overnight SpO2 were inversely correlated with daytime EtCO2 at 3500 m but not at 130 m (figure 1A,B; table 2), while minimum overnight SpO2 was weakly positively and inversely correlated with daytime EtCO2 at 150 m and 3500 m, respectively (figure 1C). The number of desaturations/hour and percentage of sleep spent in periodic breathing correlated with daytime EtCO2 at 130 m but not at 3500 m (figures 1D,F; table 2). The number of central apnoeas/hour was higher in those with lower daytime EtCO2 both at 130 and at 3500 m (figure 1E). Mean overnight SpO2 (range 80–89%) was significantly correlated (R2 0.51, p=0.03) with daytime values (range 85–92%) at 3500 m, whereas, there was no correlation (R2 0.04, p=0.6) at 130 m.
Figure 1.

Effect of daytime end-tidal carbon dioxide on other respiratory measurements. (a) Daytime haemoglobin oxygen saturation (SpO2, %); (b) mean overnight SpO2; (d) number of desaturations per hour; (e) number of central apnoeas per hour; (f) percentage of total sleep time in periodic breathing each plotted against daytime end-tidal partial pressure of carbon dioxide (pCO2) for the nine children studied at 130 m (crosses) and 3500 m (open triangles); (c) relationship between mean overnight and daytime SpO2. Mean overnight SpO2 was significantly correlated (R2 0.49, p=0.035) with daytime values at 3500 m (Δ), whereas there was no correlation (R2 0.04, p=0.626) at 130 m (x).
Table 2.
Relationship between daytime EtCO2, daytime SpO2 and sleep variables at baseline (130 m) and altitude (3500 m)
| 130 m | 3500 m | |||||||
|---|---|---|---|---|---|---|---|---|
| EtCO2=38–47 mm Hg | Daytime EtCO2=31–40 mm Hg | |||||||
| R2 | p Value | β | Range | R2 | p Value | β | Range | |
| Daytime SpO2 (%) | 0.32 | 0.1 | −0.561 | 97–100 | 0.65 | 0.009 | −0.806 | 85–92 |
| Mean overnight SpO2 (%) | 0.12 | 0.4 | 0.353 | 96–98 | 0.40 | 0.066 | −0.635 | 80–89 |
| Minimum overnight SpO2 (%) | 0.46 | 0.046 | 0.675 | 84–94 | 0.38 | 0.078 | −0.615 | 60–82 |
| Number of desaturations/hour overnight | 0.75 | 0.003 | −0.865 | 0–1.7 | 0.16 | 0.3 | 0.398 | 0.7–49.3 |
| Number of central apnoeas/hour overnight | 0.78 | 0.002 | −0.882 | 0–1.7 | 0.45 | 0.047 | −0.673 | 0.3–9.5 |
| % total sleep time spent in periodic breathing | 0.76 | 0.017 | −0.763 | 0–0.4 | 0.27 | 0.15 | −0.159 | 0–17.5 |
EtCO2, end-tidal partial pressure of carbon dioxide; SpO2, haemoglobin oxygen saturation from pulse oximeter.
Transcranial Doppler
There were significant increases in CBFV in the left MCA and both ACAs at 3500 m compared with 130 m (table 3, figure 2). Left MCA velocities were lower than right at sea level, and increased more at altitude (table 3).23 There were no significant velocity changes in the PCA or BA. With the exception of the BA, calculated COD increased in all arteries. There was no correlation between change in BA velocity and change in central apnoeas or percentage periodic breathing between 130 and 3500 m.
Table 3.
Maximum cerebral blood flow velocities (CBFV) and cerebral oxygen delivery (COD) by hemisphere and altitude
| CBFV | COD | |||||
|---|---|---|---|---|---|---|
| Sea level (130 m) | Moderate altitude (1300 m) (Kathmandu) | High altitude (3500 m) (Namche Bazaar) | Cerebral oxygen delivery | |||
| Mean (SD) Velocity (cm/sec) |
Mean (SD) % Change from low altitude |
Mean (SD) % Change from low altitude |
Mean (SD) % Change from moderate altitude |
Mean (SD) % Change in from low to high altitude | ||
| MCA* (n=9) | Left | 69.1 (12.5) | 35.2 (33.4)1 | 29.2 (42.3)2 | 4.2 (19.3) | 18.7 (44.0) |
| Right | 84.0 (8.4) | 7.2 (11.4) | 9.9 (12.0) | 3.0 (10.9) | 1.9 (7.5) | |
| ACA* (n=7) | Left | 45.3 (13.2) | 43.4 (53.4) | 65.2 (69.0)2 | 28.3 (42.7) | 47.8 (60.4) |
| Right | 51.1 (22.4) | 94.1 (159.7) | 109.1 (179.2)2 | 13.5 (34.4) | 41.6 (68.7) | |
| PCA*$ (n=7) | Left | 37.4 (11.2) | 40.5 (71.0) | 49.9 (75.0) | 15.1 (35.4) | 34.9 (65.6) |
| Right | 38.7 (6.2) | 21.1 (43.9) | 17.5 (33.5) | 6.4 (29.4) | 12.6 (25.7) | |
| BA* (n=9) | 53.2 (12.6) | 4.5 (16.4) | 9.5 (21.8) | 4.9 (14.0) | −1.3 (19.0) | |
Percentage change in COD was calculated using the following formula: ((AltSPo2×AltCBFV)−(SeaSpo2×SeaCBFV))/(SeaSpo2×SeaCBFV) where: AltSpo2 is the SpO2 value at altitude; AltCBFV is the CBFV at altitude; SeaSpO2 is the SpO2 at sea level; and sea CBFV is the CBFV at sea level, and haemoglobin is assumed to be constant.
There were significant effects of hemisphere, F(1,8)=10.66, p=0.011, η2=0.57 and of altitude, F(2,7)=5.08, p=0.043, η2=0.59 on maximum MCA velocities, and a significant interaction, F (2,7)=5.33, p=0.039, η2=0.60. Inspection of table 3 reveals that CBFV in the MCA significantly increased at moderate and high altitude, compared with sea level, to a greater degree in the left than in the right hemisphere.
Although inspection of table 3 shows that CBFV appeared to increase within the ACA, both left and right, this just failed to reach significance despite a large effect, F(2,5)=5.28, p=0.059, η2=0.68, likely due to the small N, a finding confirmed by posthoc, pairwise comparisons which revealed significant increases in ACA CBFV in both hemispheres from low to high altitude. There were no significant effects of hemisphere or altitude on PCA, and no effect of altitude in the BA, all F<2.3.
Data are presented as mean or geometric mean (SD), velocities are reported as cm/s; 1=significant between groups difference between low and moderate altitude; 2=between low and high altitude.
$PCA values were transformed for the analysis using the natural logarithm; however, the mean (SD) values are provided for ease of interpretation.
*Middle cerebral artery (MCA), anterior cerebral artery (ACA), posterior cerebral artery (PCA), basilar artery (BA).
Figure 2.

Velocities in the left and right middle, anterior, posterior and basilar arteries at 130 m, 1350 m and 3500 m in nine children resident at sea level.
Daytime SpO2 and MCA velocities
An exploratory multiple regression analysis was built to predict right MCA CBFV at 3500 m, entering baseline right MCA CBFV at 130 m (step 1), daytime SpO2 at 3500 m as a percentage of daytime SpO2 at 130 m (step 2) and EtCO2 at 3500 m as a percentage of EtCO2 at 130 m (step 3). Right MCA CBFV at 130 m was not a significant predictor of right MCA CBFV at 3500 m (R2 0.2, p=0.225). However, a model with right MCA CBFV at 130 m and daytime SpO2 at 3500 m as a percentage of SpO2 at 130 m was significant, with both right MCA CBFV at 130 m (β 0.91 (95% CI 0.46 to 1.59), p=0.004), and change in daytime SpO2 (β −0.91 B −4 (95% CI −6.7 to −1.9), p=0.007), as significant predictors (R2 change 0.60; p=0.006, F[2,8]=1.79; p=0.008). EtCO2 at 3500 m as a percentage of EtCO2 at 130 m also contributed significant variance to right MCA CBFV at 3500 m (R2 change=0.13; p=0.034; F[3,8]=20.21; p=0.003). Together, these three variables explained 92% of the variance in right MCA CBFV at 3500 m. Change in HR was not a significant predictor. Inspection of the β weights revealed that increase in right MCA CBFV and greater reduction in SpO2 and EtCO2 between 130 m and 3500 m significantly predicted right MCA CBFV at 3500 m. Sea level right MCA CBFV was not predictive until SpO2 changes were entered into the model, consistent with an interaction between individual differences in right MCA CBFV and the degree to which SpO2 and EtCO2 were compromised at altitude. The residuals were normally distributed, consistent with good model fit to the data. For left MCA and BA velocities, none of the models were significant.
Memory
There were no differences between Rivermead scores at the three altitudes.
Discussion
Respiratory and sleep physiology
Prior to this expedition, data on the effects of altitude on daytime and/or sleep physiology in children sojourning at altitude were lacking. This study appears to show that children have similar physiological responses to altitude as those recorded previously in adults,1 2 but have less central apnoeas24 and periodic breathing.2 The number of central apnoeas, and percentage of periodic breathing in sleep, increased at high altitude, with concurrent increases in the number and severity of oxygen desaturations. EtCO2 was associated with the number of central apnoeas at 130 m, suggesting that at normoxia in children, the baseline degree of hypocapnia may determine whether the apnoea threshold is reached. At 3500 m, as in adults,24 the relationship was still significant, but without overnight EtCO2 measurement, we could not determine the EtCO2 at which apnoeas were triggered.2 Daytime HR increased significantly (table 1), consistent with a 24% increase in cardiac output assuming no change in stroke volume.
Our results support previous findings of low nocturnal SpO2 at altitude in both native25 26 and non-acclimatised children,2 27 and illuminate the associated SDB patterns. The severity of hypoxia, hypoventilation and periodic breathing experienced at high altitude exacerbates sleep fragmentation, with recent evidence that hypoxia is the most important factor,28 but our study is in line with previous literature showing no effect on episodic memory.29 Mean daytime and overnight SpO2 decreased at high altitude, despite increases in RR and decreases in EtCO2 compatible with an intact HVR.
Change in CBFV and COD in anterior and posterior circulations
To our knowledge, this study is the first to examine regional TCD velocities in children unaccustomed to high altitudes in relation to daytime respiratory variables and sleep. Exposure to hypoxia and hypocapnia has conflicting effects on cerebral perfusion. In adults residing at altitude, the cerebral circulation appears less sensitive to hypoxia than to hypocapnia,30 but CBFV increases with decreasing pO2 at altitude in adults resident at sea level and in hypoxic adults at sea level, with no evidence for an effect of EtCO2.31 32 In our data, right MCA CBFV was generally higher than left, consistent with increased CBF to a larger hemisphere.33 Although there was considerable variability,12 left MCA and both ACA velocities increased, with increased calculated COD even assuming no increase in haemoglobin. However, BA and PCA velocities did not increase significantly.14 In elite climbers, basilar CBFV reduces at altitude,13 perhaps allowing build up of CO2 and, therefore, unstable breathing.6 34 Although in our data the change in basilar velocity was not associated with change in central apnoeas or periodic breathing, this could be further explored in overnight studies. Calculated COD appears to fall in the basilar distribution on exposure to high altitude,35 which may be related to reduced metabolic rate, but might increase the risk of compromise to the parieto-occiptal cerebellar brain tissues associated with the common complaints of nausea, dizziness, ataxia and visual symptoms.36
Asymmetry in haemodynamic response
In humans native to high altitude, CBFV changes in response to isocapnic hypoxia may show regional brain differences,12 37 38 but there are few data in children resident at sea level exposed to hypocapnic hypoxia. Although we lacked direct measurements of blood gases, the increase in RR and decrease in pCO2 suggests a response to hypoxia in this study, and there appears to be an interaction between baseline right MCA CBFV and change in day (and night-time) SpO2 and daytime EtCO2 in predicting 92% of the variance in high-altitude right MCA CBFV. This is consistent with the interaction between right MCA CBFV and the hypercapnic ventilatory response previously documented at sea level in healthy adults.5 In turn, this means that the degree of desaturation and hypocapnia at altitude is less and, therefore, the change in right MCA CBFV is less if right MCA CBFV is higher at sea level. Most adult studies have only reported right MCA CBFV, but Feddersen, who examined the left MCA, also found asymmetry.39 There is evidence that the right hemisphere controls neural sympathetic outflow, with a possible interaction with pCO2,40 but the direction and lateralisation of the association we have demonstrated requires further investigation, including direct measurement of blood gases.
Control of change in CBFV on exposure to hypocapnic hypoxia at altitude
In line with previous data in adults, our data suggest the importance of oxygen content in the regulation of the ACA circulation, so that at altitude, in children resident at sea level, CBF, for which CBFV is a proxy, increases in response to hypoxaemia,32 even when there is hypocapnia.30
Limitations
MCA diameter may increase on acute exposure to 12% hypoxia or at extreme altitude (>5300).41 In our study, the children made a slow ascent to 3500 m, and cerebral artery diameters were assumed to be unchanged, as is also the case with SpO2 as low as 70% at sea level.38 A change of <10% in arterial diameter was recorded with EtCO2 as low as 20 mm Hg in adults.38 Future studies should check the assumption that vascular diameter does not change over these ranges, and should look at regional cerebral perfusion variations and diurnal variation of CBF in relation to EtCO2 and breathing instability in various sleep stages, determined using EEG, to determine whether clinically relevant differences occur. In addition, recording pH, for instance, in urine, would be important, provided validation compared with blood measurements, which are difficult to justify on ethical grounds, is achieved. In combination with serial CBFV measurements, this would allow exploration of the metabolic compensation to hypocapnia, which occurs in the first 2–3 days after arrival at altitude, involving renal loss of serum bicarbonate, as well as reduction in CSF bicarbonate production, thereby minimising the deviation of arterial and CSF pH.
In summary, our preliminary data confirm that healthy children resident at sea level, and exposed to high altitudes during a slow ascent to 3500 m, experience periodic breathing and further haemoglobin oxygen desaturation during sleep. SpO2 and EtCO2 appear to be important determinants of the increase in right-sided MCA CBFV that occurs on exposure to altitude. Despite hypocapnia, this study demonstrates for the first time in children that there is an increase in CBFV at altitude, adequate to preserve COD in the distribution of all vessels but the BA.
Footnotes
Contributors: JCG collected and analysed the data, and wrote the first draft of the paper. JS designed the study, collected and analysed the data, and edited the draft of the paper. AL collected and analysed the data, and edited the draft of the paper. EF collected and analysed the data, and edited the draft of the paper. RB undertook the final statistical analysis of the data, and edited the draft of the paper. SS collected and analysed the data, and edited the draft of the paper. JC analysed the memory data, and edited the draft of the paper. MPG designed the study, collected and analysed the data, and edited the draft of the paper. DZL designed the study, collected and analysed the data, and edited the draft of the paper. DSM designed the study, collected and analysed the data, and edited the draft of the paper. CHI designed the study, collected and analysed the data, and edited the draft of the paper. FJK designed the analysis and analysed the data, edited the draft of the paper, and acts as guarantor for the data.
Funding: The Young Everest Study was funded by Smiths Medical.
Competing interests: None.
Ethics approval: UCL Research Ethics Committee.
Provenance and peer review: Not commissioned; externally peer reviewed.
Open Access: This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 3.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/3.0/
References
- 1.Berssenbrugge A, Dempsey J, Iber C, et al. Mechanisms of hypoxia-induced periodic breathing during sleep in humans. J Physiol 1983;343:507–26 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kohler M, Kriemler S, Wilhelm EM, et al. Children at high altitude have less nocturnal periodic breathing than adults. Eur Respir J 2008;32:189–97 [DOI] [PubMed] [Google Scholar]
- 3.Virues-Ortega J, Bucks R, Kirkham FJ, et al. Changing patterns of neuropsychological functioning in children living at high altitude above and below 4000 m: a report from the Bolivian Children Living at Altitude (BoCLA) study. Dev Sci 2011;14:1185–93 [DOI] [PubMed] [Google Scholar]
- 4.Dempsey JA, Veasey SC, Morgan BJ, et al. Pathophysiology of sleep apnea. Physiol Rev 2010;90:47–112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Xie A, Skatrud JB, Morgan B, et al. Influence of cerebrovascular function on the hypercapnic ventilatory response in healthy humans. J Physiol 2006; 577(Pt 1):319–29 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ainslie PN, Duffin J. Integration of cerebrovascular CO2 reactivity and chemoreflex control of breathing: mechanisms of regulation, measurement, and interpretation. Am J Physiol Regul Integr Comp Physiol 2009;296:R1473–95 [DOI] [PubMed] [Google Scholar]
- 7.Dempsey JA, Smith CA, Przybylowski T, et al. The ventilatory responsiveness to CO(2) below eupnoea as a determinant of ventilatory stability in sleep. J Physiol 2004;560(Pt 1):1–11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dempsey JA. Crossing the apnoeic threshold: causes and consequences. Exp Physiol 2005;90:13–24 [DOI] [PubMed] [Google Scholar]
- 9.Kirkham FJ, Padayachee TS, Parsons S, et al. Transcranial measurement of blood velocities in the basal cerebral arteries using pulsed Doppler ultrasound: velocity as an index of flow. Ultrasound Med Biol 1986;12:15–21 [DOI] [PubMed] [Google Scholar]
- 10.Park CW, Sturzenegger M, Douville CM, et al. Autoregulatory response and CO2 reactivity of the basilar artery. Stroke 2003;34:34–9 [DOI] [PubMed] [Google Scholar]
- 11.Gupta AK, Menon DK, Czosnyka M, et al. Thresholds for hypoxic cerebral vasodilation in volunteers. Anesth Analg 1997;85:817–20 [DOI] [PubMed] [Google Scholar]
- 12.Noth U, Kotajima F, Deichmann R, et al. Mapping of the cerebral vascular response to hypoxia and hypercapnia using quantitative perfusion MRI at 3 T. NMR Biomed 2008;21:464–72 [DOI] [PubMed] [Google Scholar]
- 13.Jansen GF, Kagenaar DA, Basnyat B, et al. Basilar artery blood flow velocity and the ventilatory response to acute hypoxia in mountaineers. Respir Physiol Neurobiol 2002;133:65–74 [DOI] [PubMed] [Google Scholar]
- 14.Garbin L, Habetswallner F, Clivati A. Vascular reactivity in middle cerebral artery and basilar artery by transcranial Doppler in normals subjects during hypoxia. Ital J Neurol Sci 1997;18:135–7 [DOI] [PubMed] [Google Scholar]
- 15.Smith KJ, Wong LE, Eves ND, et al. Regionalcerebral blood flow distribution during exercise: Influence of oxygen. Respir Physiol Neurobiol 2012; 184:97–105 [DOI] [PubMed] [Google Scholar]
- 16.Meadows GE, O'Driscoll DM, Simonds AK, et al. Cerebral blood flow response to isocapnic hypoxia during slow-wave sleep and wakefulness. J Appl Physiol 2004;97:1343–8 [DOI] [PubMed] [Google Scholar]
- 17.Scrase E, Laverty A, Gavlak JC, et al. The Young Everest Study: effects of hypoxia at high altitude on cardiorespiratory function and general well-being in healthy children. Arch Dis Child 2009;94:621–6 [DOI] [PubMed] [Google Scholar]
- 18.Beebe DW. Neurobehavioral effects of obstructive sleep apnea: an overview and heuristic model. Curr Opin Pulm Med 2005;11:494–500 [DOI] [PubMed] [Google Scholar]
- 19.Levett DZ, Martin DS, Wilson MH, et al. Design and conduct of Caudwell Xtreme Everest: an observational cohort study of variation in human adaptation to progressive environmental hypoxia. BMC Med Res Methodol 2010;10:98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Padayachee TS, Kirkham FJ, Lewis RR, et al. Transcranial measurement of blood velocities in the basal cerebral arteries using pulsed Doppler ultrasound: a method of assessing the Circle of Willis. Ultrasound Med Biol 1986;12:5–14 [DOI] [PubMed] [Google Scholar]
- 21.Imray CH, Myers SD, Pattinson KT, et al. Effect of exercise on cerebral perfusion in humans at high altitude. J Appl Physiol 2005;99:699–706 [DOI] [PubMed] [Google Scholar]
- 22.Aldrich FK, Wilson B. Rivermead Behavioural Memory Test for Children (RBMT-C): a preliminary evaluation. Br J Clin Psychol 1991;30(Pt 2):161–8 [DOI] [PubMed] [Google Scholar]
- 23.Vavilala MS, Kincaid MS, Muangman SL, et al. Gender differences in cerebral blood flow velocity and autoregulation between the anterior and posterior circulations in healthy children. Pediatr Res 2005;58:574–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Burgess KR, Johnson PL, Edwards N. Central and obstructive sleep apnoea during ascent to high altitude. Respirology 2004;9:222–9 [DOI] [PubMed] [Google Scholar]
- 25.Gamponia MJ, Babaali H, Yugar F, et al. Reference values for pulse oximetry at high altitude. Arch Dis Child 1998;78:461–5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Vazquez JC, Montes FM, Rivera CA, et al. Clinical predictors of sleep disordered breathing in children at moderate altitude. Arch Med Res 2004;35:525–31 [DOI] [PubMed] [Google Scholar]
- 27.Yaron M, Waldman N, Niermeyer S, et al. The diagnosis of acute mountain sickness in preverbal children. Arch Pediatr Adolesc Med 1998; 152:683–7 [DOI] [PubMed] [Google Scholar]
- 28.Nussbaumer-Ochsner Y, Ursprung J, Siebenmann C, et al. Effect of short-term acclimatization to high altitude on sleep and nocturnal breathing. Sleep 2012;35:419–23 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Thomas RJ, Tamisier R, Boucher J, et al. Nocturnal hypoxia exposure with simulated altitude for 14 days does not significantly alter working memory or vigilance in humans. Sleep 2007;30:1195–203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Norcliffe LJ, Rivera-Ch M, Claydon VE, et al. Cerebrovascular responses to hypoxia and hypocapnia in high-altitude dwellers. J Physiol 2005;566(Pt 1):287–94 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ainslie PN, Burgess K, Subedi P, et al. Alterations in cerebral dynamics at high altitude following partial acclimatization in humans: wakefulness and sleep. J Appl Physiol 2007;102:658–64 [DOI] [PubMed] [Google Scholar]
- 32.Baumgartner RW, Bartsch P, Maggiorini M, et al. Enhanced cerebral blood flow in acute mountain sickness. Aviat Space Environ Med 1994; 65:726–9 [PubMed] [Google Scholar]
- 33.Carne RP, Vogrin S, Litewka L, et al. Cerebral cortex: an MRI-based study of volume and variance with age and sex. J Clin Neurosci 2006;13:60–72 [DOI] [PubMed] [Google Scholar]
- 34.Xie A, Skatrud JB, Barczi SR, et al. Influence of cerebral blood flow on breathing stability. J Appl Physiol 2009;106:850–6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Brothers MD, Wilber RL, Byrnes WC. Physical fitness and hematological changes during acclimatization to moderate altitude: a retrospective study. High Alt Med Biol 2007;8:213–24 [DOI] [PubMed] [Google Scholar]
- 36.Wilson MH, Newman S, Imray CH. The cerebral effects of ascent to high altitudes. Lancet Neurol 2009;8:175–91 [DOI] [PubMed] [Google Scholar]
- 37.Binks AP, Cunningham VJ, Adams L, et al. Gray matter blood flow change is unevenly distributed during moderate isocapnic hypoxia in humans. J Appl Physiol 2008;104:212–17 [DOI] [PubMed] [Google Scholar]
- 38.Willie CK, Macleod DB, Shaw AD, et al. Regional brain blood flow in man during acute changes in arterial blood gases. J Physiol 2012;590(Pt 14):3261–75 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Feddersen B, Ausserer H, Neupane P, et al. Right temporal cerebral dysfunction heralds symptoms of acute mountain sickness. J Neurol 2007;254:359–63 [DOI] [PubMed] [Google Scholar]
- 40.Diedrich A, Porta A, Barbic F, et al. Lateralization of expression of neural sympathetic activity to the vessels and effects of carotid baroreceptor stimulation. Am J Physiol Heart Circ Physiol 2009;296:H1758–65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wilson MH, Edsell ME, Davagnanam I, et al. Cerebral artery dilatation maintains cerebral oxygenation at extreme altitude and in acute hypoxia–an ultrasound and MRI study. J Cereb Blood Flow Metab 2011;31:2019–29 [DOI] [PMC free article] [PubMed] [Google Scholar]