

Abstract
I propose a re-conceptualization of key phenomena important in the study of emotion — those phenomena that reflect functions and circuits related to survival, and that are shared by humans and other animals. The approach shifts the focus from questions about whether emotions that humans consciously feel are also present in other animals, and towards questions about the extent to which circuits and corresponding functions that are present in other animals (survival circuits and functions) are also present in humans. Survival circuit functions are not causally related to emotional feelings, but obviously contribute to these, at least indirectly. The survival circuit concept integrates ideas about emotion, motivation, reinforcement, and arousal in the effort to understand how organisms survive and thrive by detecting and responding to challenges and opportunities in daily life.
Introduction
Emotion is a major research growth area in neuroscience and psychology today. A search of PubMed citations for the 1960s yields just over 100 papers with the word “emotion” in the title. With each subsequent decade, small increases resulted, until the last decade, when emotion titles grew exponentially — more than 2000 hits. Emotion has happened.
But what exactly is it that has happened? What is being studied in all these papers on emotion? Actually, the term “emotion” is not well-defined in most publications. Perhaps this is not surprising since there is little consensus about what emotion is, and how it differs from other aspects of mind and behavior, in spite of discussion and debate that dates back to the earliest days in modern biology and psychology (e.g. Darwin, 1872; James, 1884; Cannon, 1927, 1931; Duffy, 1934, 1941; Tomkins, 1962; Mandler, 1975; Schachter, 1975; Ekman, 1980, 1984, 1992; Izard, 2007; Frijda, 1986; Russell, 2003;; Ekman and Davidson, 1994; LeDoux, 1996; Panksepp, 1998, 2000, 2005; Rolls, 1999, 2005; Damasio, 1994, 1999; Leventhal and Scherer, 1987; Scherer, 2000; Ortony and Turner, 1990; Öhman, 1986, 2009; Johnson-Laird and Oatley, 1989; Ellsworth, 1994; Zajonc, 1980; Lazarus, 1981, 1991a, b; Barrett, 2006a, b; 2007; Kagan, 2007; Prinz, 2004; Scarantino, 2009; Griffiths, 2004; Ochsner and Gross, 2005; Lyons, 1980).
One point that many writers on this topic accept is that, while there are unique features of human emotion, at least some aspects of human emotion reflect our ancestral past. This conclusion is the basis of neurobiological approaches to emotion, since animal research is essential for identifying specific circuits and mechanisms in the brain that underlie emotional phenomena.
Progress in understanding emotional phenomena in the brains of laboratory animals has in fact helped elucidate emotional functions in the human brain, including pathological aspects of emotion. But what does this really mean? If we don’t have an agreed upon definition of emotion that allows us to say what emotion is, and how emotion differs from other psychological states, how can we study emotion in animals or humans, and how can we make comparisons between species?
The short answer is that we fake it. Introspections from personal subjective experiences tell us that some mental states have a certain “feeling” associated with them and others do not. Those states that humans associate with feelings are often called emotions. The terms “emotion” and “feeling” are, in fact, often used interchangeably. In English we have words like fear, anger, love, sadness, jealousy, and so on, for these feeling states, and when scientists study emotions in humans they typically use these “feeling words” as guideposts to explore the terrain of emotion.
The wisdom of using common language words that refer to feelings as a means of classifying and studying human emotions has been questioned by a number of authors over the years (e.g. Duffy, 1934, 1941; Mandler, 1975; Russell, 1991, 2003; Barrett, 2006a, 2006b; Kagan, 2007; Griffiths, 1997; Rorty, 1980; Dixon, 2001; Zachar, 2006). Whatever problems might arise from using feeling words to study human emotion, the complications are compounded many fold when such words are applied to other animals. While there are certainly emotional phenomena that are shared by humans and other animals, introspections from human subjective experience are not the best starting point for pursuing these. How, then, should the aspects of emotion relevant to animals and humans alike be pursued?
In answering this question it is important to separate the phenomena of interest from the overarching concept of emotion. One set of such phenomena includes responses that occur when an organism detects and responds to significant events in the course of surviving and/or maintaining well-being — for example, responses that occur when in danger or when in the presence of a potential mate or in the presence of food when hungry or drink when thirsty. These are fundamental phenomena that have always interested animal behavior scientists, and would be of interest even if the terms “emotion” and “feelings” never existed. The challenge for emotion researchers is to understand the relation of the phenomena to the field of emotion without redefining them as fundamentally emotional phenomena, and thus infusing the phenomena with confusing implications.
In this Perspectives I, therefore, describe a way of conceiving phenomena important to the study of emotion, but with minimal recourse to the terms emotion or feelings. The focus is instead on circuits that instantiate functions that allow organisms to survive and thrive by detecting and responding to challenges and opportunities. Included, at a minimum, are circuits involved in defense, maintenance of energy and nutritional supplies, fluid balance, thermoregulation, and reproduction. These survival circuits and their adaptive functions are conserved to a significant degree in across mammalian species, including humans. While there are species-specific aspects of these functions, there are also core components of these functions that are shared by all mammals.
By focusing on survival functions instantiated in conserved circuits, key phenomena relevant to emotions and feelings are discussed with the natural direction of brain evolution in mind (by asking to what extent are functions and circuits that are present in other mammals also present in humans) rather than by looking backwards, and anthropomorphically, into evolutionary history (by asking whether human emotions/feelings have counterparts in other animals).
Emotion, motivation, reinforcement and arousal are closely related topics and often appear together in proposals about emotion. Focusing on survival functions and circuits allows phenomena related to emotion, motivation, reinforcement and arousal to be treated as components of a unified process that unfolds when an organism faces a challenge or opportunity
What follows is not an attempt at explaining or defining emotion. Instead, the aim is to offer a framework for thinking about some key phenomena associated with emotion (phenomena related to survival functions) in a way that is not confounded by confusion over what emotion means. Stepping back from the over-arching concept of emotion and focusing instead on key phenomena that make emotion an interesting topic may be the best way out of the conceptual stalemate that results from endless debates about what emotion is.
Why Do We Need to Rethink the Relation of Emotion to Survival?
The relation of innate survival functions to emotions is hardly novel, and goes back at least to Darwin (1872). As a result, neuroscientists have long assumed that specific emotional/motivational circuits are innately wired into the brain by evolution, and that these mediate functions that contribute to survival and well-being of the organism (e.g. Cannon, 1929; MacLean, 1949, 1952; Hess, 1954; Stellar, 1954; von Holst and von Saint Paul, 1962; Flynn, 1967; Olds, 1977; Siegel and Edinger, 1981; Panksepp, 1982, 1998, 2005; Blanchard and Blanchard, 1972; Bolles and Fanselow, 1980; Damasio, 1994, 1999; Berridge, 1999; McNaughton, 1989; Swanson, 2000; Ferris et al, 2008; Choi et al, 2005; Motta et al, 2009; Lin et al, 2011; Öhman, 2009). That certain emotions are wired into the brain is also a major tenet of evolutionary psychology (e.g. Tooby and Cosmides, 1990; Pinker, 1997; Nesse, 1990). If many researchers in the field (past and present) believe this, why do we need to bother with another discussion of the topic?
A major controversy in the field of emotion research today is, in fact, about the issue of whether there are innate emotion circuits in the human brain. This debate is centered on the question of whether emotions are “natural kinds,” things that exist in nature as opposed to being inventions (constructions) of the human mind (e.g. Panksepp, 2000; Griffiths, 2004; Barrett, 2006a; Izard, 2007; Scarantino, 2009). Much of the discussion is focused the question of whether so-called “basic emotions” are natural kinds. Basic emotions are those that area said to be universally expressed and recognized in people around the world, conserved in our close animal ancestors, and supposedly hard-wired into brain circuits by evolution (Darwin, 1872; Tomkins, 1962; Ekman, 1972, 1980, 1984, 1992, 1999a, b; Izard, 1992, 2007; Damasio, 1994, 1999; Panksepp, 1998, 2000, 2005; Prinz, 2004). Contemporary theories recognize between 5 and 7 of these basic or primary emotions. Ekman’s (1972) list of six basic emotions is the canonical example, and includes fear, anger, happiness, sadness, disgust, and surprise. This list of putative hardwired basic emotions in fact serves as the foundation for much research on the neural basis of emotional functions in the human brain — a recent review uncovered 551 studies between 1990 and 2008 that used Ekman’s basic emotions faces or variants of these to study functional activity related to emotion in the human brain (see Fusar-Poli et al, 2009).
In spite of being well known and widely applied in research, the basic emotions point of view has been challenged on various grounds (e.g. Averill, 1980; Ortony and Turner, 1990; Russell, 1980, 2003; Barrett, 2006a; Barrett et al, 2007). For one thing, different theories have different numbers of basic emotions, and even different names for similar emotions. In addition, questions have been raised about the methods used to identify basic emotions (e.g. forced choice rather than free labeling of the emotion expressed in a face). Basic emotions theory has also been challenged on the basis of a lack of coherence of the phenomena that constitute individual emotions, and the diversity of states to which a given emotion label can refer. Others argue that emotions, even so-called basic emotions, are psychological/social constructions, things created by the mind when people interact with the physical or social environment, as opposed to biologically determined states. Also relevant is the fact that the main basic emotions theory based on brain research in animals (Panksepp, 1998, 2005) lists emotions that do not match up well with those listed by Ekman or otherS as human basic emotions.
Of particular relevance here is Barrett’s recent challenge to the natural kinds status of basic emotions, and particularly to the idea that the human brain has evolutionarily conserved neural circuits for basic emotions (Barrett, 2006a; Barrett et al, 2007). Her argument is centered on several points: that much of evidence in support of basic emotions in animals is based on older techniques that lack precision (electrical brain stimulation), that basic emotions identified in animals do not map onto the human categories, and that evidence from human imaging studies show that similar brain areas are activated in response to stimuli associated with different basic emotions. I disagree with Barrett’s conclusion that the similarity of functional activation in different emotions is an argument against basic emotions since imaging does not have the resolution necessary to conclude that the similarity of activation in different states means similar neural mechanisms. Yet, I concur with her conclusion that the foundation of support for the idea that basic emotions, as conventionally conceived, have dedicated neural circuits is weak. This does not mean that the mammalian brain lacks innate circuits that mediate fundamental phenomena relevant to emotion. It simply means that emotions, as defined in the context of human basic emotions theory, may not be the best way to conceive of the relevant innate circuits. Enter survival circuits.
Survival Circuits
It has long been known that the body is a highly integrated system consisting of multiple subsystems that work in concert to sustain life both on a moment to moment to basis and over long time scales (Bernard, 1878–1879; Cannon, 1929; Lashley, 1938; Morgan, 1943; Stellar, 1954; Selye, 1955; McEwen, 2009; Damasio, 1994, 1999; Pfaff, 1999; Schulkin, 2003). A major function of the brain is to coordinate the activity of these various body systems. An important category of life-sustaining brain functions are those that are achieved through behavioral interactions with the environment. As noted, these survival circuits include, at a minimum, circuits involved in defense, maintenance of energy and nutritional supplies, fluid balance, thermoregulation, and reproduction.
Survival circuits have their ultimate origins in primordial mechanisms that were present in early life forms. This is suggested by the fact that extant single cell organisms, such as bacteria, have the capacity to retract from harmful chemicals and to accept chemicals that have nutritional value (Macnab and Koshland, 1972). With the evolution of multicellular, and multisystem, eukaryotic organisms (Metazoa, or what we usually call animals), fundamental survival capacities increase in complexity and sophistication, in large part due to the presence of specialized sensory receptors and motor effectors, and a central nervous system that can coordinate bodily functions and interactions with the environment (Shepherd, 1988).
The brains of vertebrate organisms vary in size and complexity. Yet, in spite of these differences, there is a highly conserved organizational plan that is characteristic of all vertebrate brains (Nauta and Karten, 1970; Northcutt and Kass, 1995; Swanson, 2002; Butler and Hodos, 2005; Striedter, 2005). This conservation is most often discussed in terms of central sensory and motor systems. However, sensory motor systems do not exist in isolation, and in fact evolved to negotiate interactions with the environment for the purpose of sustaining life; for example, by maintaining energy and fluid supplies, regulating body temperature, defending against harm, and enabling reproduction.
The survival circuits listed do not align well with human basic emotions. However, my goal is not to align survival circuits with basic emotion categories. It is instead to break free from basic emotion categories based on human emotional feelings (introspectively labeled subjective states) and instead let conserved circuits do the heavy lifting. For example, there is no anger/aggression circuit in the present scheme. This might at first seem like a striking omission. However, it is important to note that aggression is not a unitary state with a single neural representation (Moyer, 1976; Chi and Flynn, 1971; Siegel and Edinger, 1981). The distinct forms of aggression (consepcific, defensive, and predatory aggression) might be more effectively segregated by the context in which the aggression occurs: defense circuitry (aggression in an attempt to protect one’s self from harm); reproductive circuitry (aggression related to competition for mates); feeding circuitry (predatory aggression towards prey species). Similarly, a joy/pleasure/happiness kind of circuit is not listed and might seem like a fatal flaw. However, behaviors used to index joy/pleasure/happiness are instead treated products of specific circuits involved in energy and nutrition, fluid balance, procreation, thermoregulation, etc. By focusing on the subjective state, joy/pleasure/happiness, emotion theories tend to gloss over the underlying details of emotional processing for the sake of converging on a single word that symbolizes diverse underlying states mediated by different kinds of circuits.
Each survival circuit may itself need to be refined. For example, it is unlikely that there is a single unified defense or reproductive circuit. The range of functions studied needs to be expanded to more effectively characterize these. Some variations on defense are described below, but still other refinements may be needed.
Another key difference between the survival circuit and basic emotions approaches is this. Basic emotion circuits are meant as an explanation of the feelings for which each circuit is said to be responsible. Survival circuits are not posited to have any direct relation (causal role) in feelings. They indirectly influence feelings, as described later, but their function is to negotiate behavioral interactions in situations in which challenges and opportunities exist, not to create feelings.
Survival circuits help organisms survive and thrive by organizing brain functions. When activated, specific kinds of responses rise in priority, other activities are inhibited, the brain and body are aroused, attention is focused on relevant environmental and internal stimuli, motivational systems are engaged, learning occurs and memories are formed (e.g. Morgan, 1943; Hebb, 1949; Bindra, 1969; Gallistel, 1980; Scherer, 1984, 2000; Maturana and Varela, 1987; LeDoux, 2002).
In sum, survival circuits are sensory-motor integrative devices that serve specific adaptive purposes. They are tuned to detect information relevant to particular kinds of environmental challenges and opportunities, and they use this information to control behavioral responses and internal physiological adjustment that help bring closure to the situation. All complex animals (invertebrates and vertebrates) have survival circuits. Core components of these circuits are highly conserved in vertebrates. I focus on vertebrates, especially mammals in this article, but consider the relation of invertebrate to vertebrate survival functions towards the end.
Nature and Nurture in Survival Circuits
Survival circuits detect key trigger stimuli on the basis of innate programming or past experience. By innate programming I mean genetically specified synaptic arrangements that are established in early development. Innate evaluative networks make possible species-wide stimulus-response connections that allow organisms to respond to specific stimulus patterns in tried and true ways (i.e. with hard-wired/innate reactions) that have been honed by natural selection.
By experience I mean conditions under which associations are formed between novel stimuli and biologically innately significant events, typically innate triggers. These experience-dependent associations allow meaningless stimuli that occur in conjunction with significant events to acquire the ability to activate the innate response patterns that are genetically wired to innate trigger stimuli. The fact that the response patterns are innately wired and initially expressed involuntarily does not mean that they are completely inflexible. Not only can they be coupled to novel stimuli through experience and learning, they can be regulated in terms of their time course and intensity, and perhaps in other ways.
Innate and experience-based evaluative mechanisms are, as noted, circuit-specific. Thus, defense, nutritional, reproductive, thermoregulatory and other survival systems are wired to detect unique innate triggers. By entering into associations with biologically significant stimuli, novel sensory events become learned triggers that activate survival circuits. We will consider innate and learned survival circuit triggers in the context of defense next. In the field of emotion, these are described as unconditioned and conditioned fear stimuli.
Defense as an Example
The evidence for conservation across mammals of mechanisms underlying survival functions such as defense (e.g. LeDoux, 1996, 2012; Phelps and LeDoux, 2005; Motta et al, 2009; Choi et al, 2005; Kalin et al, 2004; Amaral, 2003; Antoniadis et al, 2007) reproduction (e.g. Pfaff, 1999; Oomura et al, 1988; Blaustein, 2008), thermoregulation (Nakamura and Morrison, 2007), fluid balance (Johnson, 2007; Fitzsimons, 1979), and energy/nutritional regulation (Elmquist et al, 2005; Morton et al 2006; Saper et al, 2002) is strong. Space does not permit a detailed discussion of these circuits and their functions. Defense circuits in mammals will be used as an initial illustration.
Defense against harm is a fundamental requirement of life. As noted above, even single cell organisms can detect and respond to harmful environmental stimuli. In complex organisms (invertebrates and vertebrates), threat detection involves processing of innate and learned threats by the nervous system via transmission of information about the threat through sensory systems to specialized defense circuits.
Unconditioned threat stimuli are species-specific. The most common threat triggers are stimuli that signal other animals (predators and potentially harmful conspecifics), and these will obviously be different for different species. Examples of innately wired stimuli for rodents include predator odors (e.g. Motta et al, 2009; Pagani and Rosen, 2009; Blanchard et al, 1990), as well as high frequency predator warning sounds emitted by conspecifics (e.g. Litvin et al, 2007; Choi and Brown, 2003), high intensity auditory stimuli (e.g. Bordi and LeDoux, 1992) and bright open spaces (Thompson and LeDoux, 1974; Gray, 1987; Walker and Davis, 2002). In primates, the sight of snakes and spiders have an innate propensity to trigger defense (Amaral, 2003; Ohman, 1986; Mineka and Ohman, 2002). In spite of being genetically specified, innate stimulus processing is nevertheless subject to epigenetic modulation by various factors inside and outside the organism during development, and throughout life (Bendesky and Bargmann, 2011; Monsey et al, 2011; McEwen eta l, 2012; Brown and Hariri, 2006; Casey et al, 2011; Zhang et al, 2004). Indeed, some aspects of defense stimulus processing in primates, including humans, involves preferential rapid learning to certain classes of innately “prepared” stimuli (Seligman,1971; Ohman, 1986; Mineka and Ohman, 2002). Fearful and aggressive faces of conspecifics are also a potent innate defense trigger in humans and other primates (Adolphs, 2008; Davis et al, 2011).
Recent studies have revealed in some detail the circuits that allow rodents to respond to unconditioned threats, especially odors that signal predators or potentially dangerous conspecifics (Dielenberg et al, 2001; Canteras, 2002; Petrovich et al, 2001; Markham et al, 2004; Blanchard et al, 2003; Motta et al, 2009; Choi et al, 2005; Vyas et al, 2007; Pagani and Rosen, 2009) (Figure 1). The odors are detected by the vermonasal olfactory system and sent to the medial amygdala (MEA), which connects with the ventromedial hypothalamus (VMH). Outputs of the latter reach the premammillary nucleus (PMH) of the hypothalamus, which connects with dorsal periaqueductal gray (PAGd). But not all unconditioned threats are signaled by odors. Unconditioned threats processed by other (non-olfactory) modalities involve sensory transmission to the lateral amygdala (LA) and from there to the accessory basal amygdala (ABA), which connects with the VMH-PM-PAGv circuitry (Motta et al, 2009). Different subnuclei of the MEA, PMH and PAGd are involved in processing conspecific and predatory threats. Thus, in the case of both olfactory and non-olfactory unconditioned threat signals, the PAGd and its outputs to motor control areas direct the expression of behavioral responses that help promote successful resolution of the threatening event. The PAG is also involved in detection of internal physiological signals that trigger defensive behavior (Schimitel et al, 2011).
Figure 1. Circuits Underlying Defense Reactions Elicited by Unconditioned (Unlearned) and Conditioned (Learned) Threats.
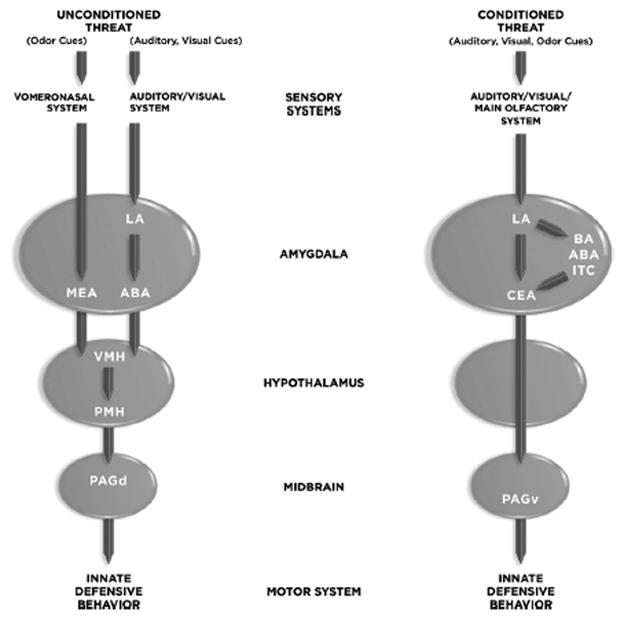
Abbreviations: ABA, accessory basal amygdala; BA, basal amygdala; CEA, central amygdala; LA, lateral amygdala; LH, lateral hypothalamus; MEA, medial amygdala; NAcc, nucleus accumbens; VMH, ventromedial hypothalamus; PAGd, dorsal periaqueductal gray region; PAGv, venral periaqueductal gray region; PMH, premammilary nucleus of the hypothalamus.
Biologically insignificant stimuli acquire status as threat signals results when they occur in conjunction with biologically significant threats. This is called Pavlovian defense conditioning, more commonly known as fear conditioning. Thus, a meaningless conditioned stimulus (CS) acquires threat status after occurring in conjunction with an aversive unconditioned stimulus (US). Most studies of Pavlovian defense conditioning involve the use of electric shock as the biologically significant US, though other modalities have been used as well. Typically, auditory, visual or olfactory stimuli as the insignificant CS. While a strong US can induce learning to most kinds of sensory stimuli, associability is not completely promiscuous — for example, taste stimuli associate more readily with gastric discomfort than with electric shock (Garcia et al, 1968). Once the association is formed, the CS itself has the ability to elicit innate defense responses.
The neural circuit by which a CS (auditory, visual, olfactory) elicits innate defense responses, such as freezing behavior, involves transmission of sensory inputs to the LA, intraamygdala connections (direct and indirect) linking the LA with the central nucleus of the amygdala (CEA), and connections from the medial CEA (CEm) to the ventrolateral PAG (PAGvl) (Johansen et al, 2011; LeDoux, 2000; Maren, 2001; Fanselow and Poulos, 2005; Davis et al, 1997; Rosenkranz and Grace, 2002; Cousens and Otto, 1998; Pare et al, 2004; Maren and Quirk, 2004; Quirk and Mueller, 2008). The indirect connections between LA and CEA include the basal (BA), AB, and intercalated (ITC) nuclei (Pitkanen et al, 1997; Pare et al, 2004). As with unconditioned threats, PAG outputs to motor control regions direct behavioral responses to the threat. While damage to the PAGvl disrupts defensive freezing behavior, lesions of the PAGdl enhance freezing (De Oca et al, 1998), suggesting interactions between these regions. Whether the CEA and PAG might also be linked via the VMH or other hypothalamic nuclei has not been carefully explored.
While most studies have focused on freezing, this behavior mainly occurs in confined spaces where escape is not possible (Fanselow, 1994; Blanchard et al, 1990; de Oca et al, 2007; Canteras et al, 2010). Little work has been done on the neural basis of defense responses other than freezing that are elicited by a conditioned cues (but see de Oca and Fanselow, 2004).
An important goal for future work is to examine the relation of circuits involved in innate and learned behavior. Electric shock simulates tissue damage produced by predator induced wounds. However, it is difficult to trace the unconditioned stimulus pathways with this kind of stimulus. Recent studies exploring interactions between circuits processing olfactory conditioned and unconditioned stimuli is an important new direction (Pavesi et al, 2011).
Another form of Pavlovian defense conditioning involves the association between a taste CS and a nausea-inducing US. The circuits underlying so called conditioned taste aversion also involve regions of the amygdala, such as CEA and the basoloateral complex (which includes the LA, BA, and ABA nuclei), as well as areas of taste cortex (Lamprecht and Dudai, 2000). However, the exact contribution of amygdala areas to learning and performance of the learned avoidance response is less clear than for the standard defense conditioning paradigms described above.
While much of the work on threat processing has been conducted in rodents, many of the findings apply to other species. For example, the amygdala nuclei involved in responding to conditioned threats in rodents appear to function similarly in rabbits (Kapp et al, 1992) and non-human primates (Kalin et al, 2001, 2004; Antoniadis et al, 2007). Evidence also exists for homologous amygdala circuitry in reptiles (Martinez-Garcia et al, 2002; Davies et al, 2002; Bruce and Neary, 1995) and birds (Cohen, 1974). In addition, functional imaging and lesion results from humans (e.g. Phelps, 2006; Damasio 1994, 1999; LaBar and Cabeza, 2006; Whalen and Phelps, 2009; Buchel and Dolan, 2000; Mobbs et al, 2009; Schiller and Delgado, 2010; Mobbs et al, 2009) show that the amygdala plays a key role in defense conditioning, and thus suggest that, at least to a first approximation, similar circuits are involved in humans as in other mammals. However, the level of detail that has been achieved in humans pales in comparison to the animal work. Methods available for studying humans are, and are likely to continue to be, limited to levels of anatomical resolution that obscure circuit details.
Because animal research is thus essential for relating detailed structure to function in the brain, it is extremely important that the phenomena of interest be conceptualized in a way that is most conducive to understanding the relation of findings from animal research to the human condition. Survival circuits provide such a conceptualization.
Interactions Between Survival Circuit Functions
Survival circuits interact to meet challenges and opportunities. Indeed, survival functions are closely intertwined (e.g. Saper, 2006).
In the presence of a threat to survival or well-being, the brain’s resources are monopolized by the task of coping with the threat. Other activities, such as eating, drinking, and sex, are actively suppressed (Gray, 1987; Lima and Dill, 1990; Blanchard et al, 1990; Fanselow, 1994; Choi et al, 2005). However, increased behavioral activity of any kind (fighting, fleeing, foraging for food or drink, sexual intercourse) expends energy, depleting metabolic resources. At some point, the need to replenish energy rises in priority and override defensive vigilance, which might otherwise keep the animal close to home. Foraging for food or liquids often requires exposure to threats and a balance has to be struck between seeking the needed resources and staying put. Metabolic activity during any active behavior (whether fighting, feeding, foraging, fornicating) produces heat that has to be counteracted by lowering body temperature. Thermoregulation is controlled directly by homeostatic alterations that include increased sweating or panting, and by various behavioral means, such as altering fluid intake or seeking shelter. We cannot consider all possible interactions between survival circuits here. Thus, interactions between the energy/nutritional regulation system and the defense system will be discussed in some detail for illustrative purposes.
Across mammalian species, circuits involving the arcuate, ventromeidal, dorsomedial, and lateral hypothalamus, and regulated by leptin, ghrelin, glucose and insulin, control feeding in relation to energy and nutritional demands (Elmquist et al, 2005; Morton et al 2006; Saper et al, 2002; Saper, 2006). In satisfying nutritional/energy demands, behavioral responses are guided by the sensory properties of potential food sources and by cues associated with food. For example, auditory or visual cues that occur in connection with food items can modulate the energy/nutritional circuitry (e.g. Petrovich, 2011). Specifically, areas of the basolateral amygdala (LA, BA, ABA) processes these learned cues associated with food and relay them to the LH. Such cues, if sufficiently potent, can stimulate eating in animals that are sated.
Feeding does not occur in a vacuum. As noted above, when threat levels rise, feeding is suppressed (Gray, 1987; Lima and Dill, 1990; Blanchard et al, 1990; Fanselow, 1994). For example, a tone previously paired with shock inhibits feeding (Petrovich, 2011) and food motivated instrumental behavior (e.g. Cardinal et al, 2002). Connections from the basolateral amygdala to the LH facilitate feeding by a CS associated with food, while the suppression of feeding by an aversive CS involves outputs of the CEA. The exact target remains to be determined but CEA connects with LH both directly and indirectly (Petrovich et al, 1996; Pitkanen et al, 1997). While threat processing normally trumps feeding, at some point the risk of encountering harm is balanced against the risk of starvation. A similar case can be made for the suppression of other behaviors by threat processing. For example, medial amygdala areas that process threat related odors suppress reproduction via connections to VHM reproductive circuits (Choi et al, 2005).
The fact that the amygdala contributes to appetitive states (e.g. Rolls, 1999, 2005; Everitt et al, 1999, 2003; Gallagher and Holland, 1994; Holland and Gallagher, 2004; Cardinal et al, 2002; Baxter and Murray, 2002; Moscarello et al, 2009) as well as defense (see above) does not mean that the amygdala processes food and threat related cues in the same way. Similarly, the fact that both appetitive and aversive stimuli activate the amgydala in fMRI studies (e.g. Canli et al, 2002; Hamann et al, 2002; Lane et al, 1999) does not mean that these stimuli are processed the same by the amygdala. Recent unit recording studies in primates show that appetitive and aversive signals are processed by distinct neuronal populations of cells in the lateral/basal amygdala (Paton et al, 2006; Belova et al, 2007; Belova et al, 2008; Morrison and Salzman, 2010; Ono and Nishijo, 1992; Rolls, 1992, 1999, 2005). Molecular imaging techniques with cellular resolution show that similarities in activation at the level of brain areas obscures differences at the microcircuit level (Lin et al, 2011).
Circuit Functions vs. Behavioral Responses
Because different groups of mammals faced different selective pressures, the behavioral responses controlled by conserved survival circuits can differ. As ethologists have long noted, many survival-related behaviors are expressed in species-specific ways (e.g. Tinbergen, 1951; Lorenz, 1981; Manning, 1967).
Consider escape from a threat. We’ve seen evidence for conserved defense circuits across mammals and even across vertebrates, but behavioral responses controlled by these circuits can differ dramatically. For example, while most mammals flee on all fours, some use only two legs (humans), others escape by flying (bats), and still others by swimming (whales, seals and walrus). Similarly, sexual and feeding behavior, while largely conserved at the neural system level, is also expressed behaviorally in diverse ways within mammals. For example, although androgen activity in the hypothalamus is important in all male mammals, the semen delivery process varies in males, in part because of different approaches required given the configuration of the male and female body (e.g. Pfaff, 1999). This is perhaps most dramatically illustrated by the lordosis posture of female rats. The male cannot insert his penis into the vaginal cavity of a female unless she arches her back to adopt this posture, which is regulated by the binding of estrogen during the fertile phase of her cycle (Pfaff, 1999; Blaustein, 2008). Further, some mammals use their snouts when eating and others their snouts paws/hands, but the core circuits described above nevertheless regulate the various homeostatic and behavioral functions required to regulate energy and nutritional supplies.
Thus, the responses used by survival circuits to achieve survival goals can be species-specific even though the circuit is largely species-general (obviously, there must be some differences in circuitry, at least in terms of motor output circuitry for different kinds of behaviors, but the core circuit is conserved). By focusing on the evolved function of a circuit (defense, reproduction, energy and nutrition maintenance, fluid balance, thermoregulation), rather than on the actual responses controlled by the circuit, a species-independent set of criteria emerge for defining brain systems that detect significant events and control responses that help meet the challenges and opportunities posed by those events.
Information Processing By Survival Circuits: Computation of Stimulus Significance
A key component of a survival circuits is a mechanism for computing circuit-specific stimulus information. A defense circuit needs to be activated by stimuli related to predators, potentially harmful conspecifcs, and other potential sources of harm, and not be triggered by potential mates or food items. The goal of such computational networks is to determine whether circuit-specific triggers are present in the current situation, and, if a trigger is detected, to initiate hard-wired (innate) responses that are appropriate to the computed evaluation. Such responses are automatically released (in the ethological sense — see Tinbergen, 1951; Lorenz, 1981; Manning, 1967) by trigger stimuli.
The nature of behavioral responses released by survival circuit triggers should be briefly discussed. Activation of a survival circuit elicits behavioral responses on the spot in some cases (e.g. in the presence of defense triggers) but in other cases unless the goal object (sexual partner, food, drink) is immediately present, the more general effect is the alteration of information processing throughout the brain in such a way as to mobilize resources for bringing the organism into proximity with suitable goal objects and thus dealing with the opportunity or challenge signaled by the trigger. We will consider a number of different consequences of survival circuit activation below. Here, we focus on information processing related to trigger detection.
Above we briefly noted the species-specific nature of innate trigger stimuli. While the original idea of the ethologists focused on complex Gestalt configural stimuli and pattern recognition, simpler features are now emphasized. Thus, a rat can recognize a predator (cat, fox) by specific chemical constituents of predator odors (Wallace and Rosen, 2000; Vyas et al, 2007; Dielenberg et al, 2001; Markham et al, 2004; Blanchard et al, 2003), and does not have to recognize the predator as a complex perceptual pattern. Moreover, humans can recognize certain emotions by the eyes alone, and do not need to process the face as a whole (e.g. Whalen et al, 2004), and evidence exists that this can be handled subcortically (Liddell et al, 2005; Whalen, 1998; Morris et al, 1999; Tamietto et al, 2009; Luo et al, 2007). These findings are consistent with the notion that that relatively simple sensory processing by subcortical areas can provide the requisite inputs to structures such as the amygdala, by passing or short-circuiting cortical areas (LeDoux, 1996). In contrast to innate trigger stimuli, learned triggers are less restricted by species characteristics. Thus, many (though not all, as noted above) stimuli can be associated with harm and become a trigger of defense circuits later.
In the field of emotion, the term automatic appraisal is sometimes used when discussing how significant stimuli elicit so-called emotional responses automatically (without deliberate control), and is contrasted with cognitive or reflective appraisal, where processing that is deliberate, controlled and often conscious, determines stimulus meaning and predisposes actions (e.g. Arnold, 1960; Bowlby, 1969; Frijda, 1986; Lazarus, 1991a, b; Leventhal and Scherer, 1987; Lazarus and Folkman, 1984; Smith and Ellsworth, 1985; Scherer, 1988; Scherer et al, 2001; Sander et al, 2005; Jarymowicz, 2009).
The stimulus significance evaluations by survival circuits are obviously more in line with automatic, unconscious appraisal mechanisms. However, while stimulus evaluations by survival circuits is clearly an example of automatic appraisal, one should not be too quick to assume that what psychologists refer to as automatic appraisals in humans is identical to survival circuit processing. The latter probably refers to a narrower set of phenomena than the former, at least in humans, if not other species, though the range of phenomena in question clearly overlap.
Multiple Roles of Innate and Learned Stimuli
So far we’ve seen that unconditioned and conditioned emotional stimuli can be thought of in other terms, as unconditioned and conditioned survival circuit triggers. In addition, though, they can also be described as incentives — stimuli that motivate instrumental behavior. The same stimuli additionally function as reinforcers — stimuli that strengthen the probability that an instrumental response will be learned and later performed. Motivation and reinforcement are obviously closely aligned with the topic of emotion, though these are often studied separately today. Let’s look more closely at how closely intertwined these processes are to one another (Fig 2).
Figure 2. Multiple Roles for a Conditioned Stimulus.
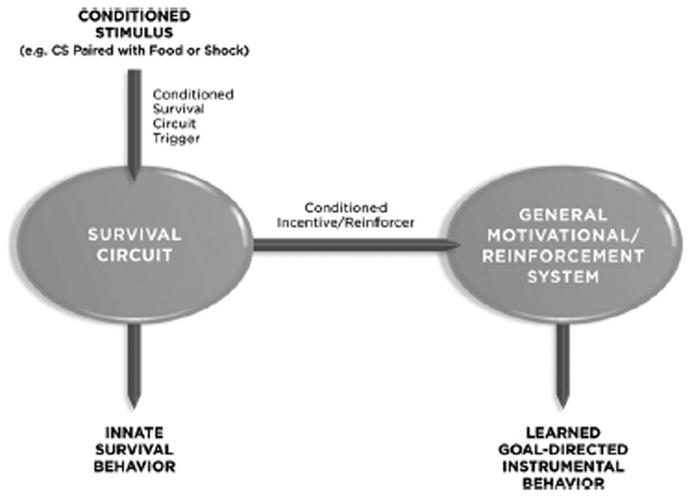
A CS functions as a survival circuit trigger (by activating a specific survival circuit related to the US that was used during conditioning), and as a conditioned incentive and a conditioned reinforcer (by way of connections from the survival circuit to motivational and reinforcement systems). Other routes by which a CS might influence motivational and reinforcement circuitry are not shown.
Consider a tone that is paired with food. This is a typical paradigm used to study positive emotional states in animals. The tone in other words is an appetitive Pavlovian CS that elicits innate approach behavior. However, it is also a survival circuit trigger, as it can stimulate eating, even in satiated rats, by activating hypothalamic circuits involved in energy management (Petrovich, 2011). The same CS will also function as a conditioned incentive that can modulate instrumental behaviors (in contrast to the ability of a CS to elicit Pavlovian innate responses (such as approach behaviors). Thus, a CS associated with food will facilitate performance of an instrumental response that is also maintained by food (e.g. bar-pressing for food) (Corbit and Balleine, 2005; Cardinal et al, 2002; Balleine and Killcross, 2006). This is called Pavlovain-to-instrumental transfer since the value of the Pavlovian CS is transferred to (alters performance of) the instrumental response. The degree of transfer depends in part on the similarity of the US in the Pavlovian and instrumental tasks. A tone CS can also be used to reinforce the learning of a new instrumental response (e.g. Holland and Rescorla, 1975). Thus, a hungry rat will learn to press a bar simply to receive the tone CS. In this case the tone is considered a reinforcer, a second-order or conditioned reinforcer (a first order or primary reinforcer would be something like food itself rather than a stimulus associated with food).
Similar relations hold for a tone paired with an aversive US, footshock. The tone CS elicits innate freezing behavior (see above) and is thus often described as a conditioned emotional stimulus (conditioned fear stimulus in this case). And just as an appetitive CS enhances bar pressing for food, and aversive CS suppresses food-maintained bar pressing (Estes and Skinner, 1941; Hammond, 1970; Cardinal et al, 2002; Balleine and Killcross, 2006). However, an aversive CS will also facilitate performance of an aversively motivated behavior (Hammond, 1970; Lazaro-Munoz et al, 2010). Further, just as rats wil l learn to perform new instrumental responses for the sole reward of receiving an appetitive CS, they will also learn new instrumental responses that are rewarded by the elimination of an aversive CS (e.g. Cain and LeDoux, 2007).
Although we’ve focused on multiple roles of CSs a similar argument can be made for USs. These are simply stimuli that innately activate survival circuits, promote the performance of consummatory responses (food is eaten, sex is consummated) in their presence, or that support Pavlovain associative conditioning or instrumental conditioning.
If we choose, we can thus describe a variety of the effects of so-called “emotional” stimuli without the use of the adjective “emotional.” These are innate or learned stimuli that activate survival circuits and trigger the expression of the innate responses controlled by these circuits, that modulate the performance of learned (previously reinforced) instrumental behaviors, and that lead to the reinforcement of new instrumental behaviors (Table 1).
TABLE 1.
Multiple Roles for so-called “Emotional” Stimuli
| 1. Survival Circuit Trigger Stimulus | Activates a specific survival circuit |
| Innate (Unconditioned) trigger | Elicits innate responses to stimuli without the need for prior exposure to the stimulus and mobilizes other brain resources to deal with the opportunity or challenge presented by the innate trigger |
| Learned (Conditioned) trigger | Potentially elicits innate responses to stimuli after being associated (via Pavlovian conditioning) with an innate trigger; more generally, mobilizes brain resources to deal with the challenge or opportunity signaled by the learned trigger |
| 2. Incentive | Modulates instrumental goal-directed behavior to help meet the opportunity or challenge signaled by the stimulus that is triggering activation of a specific survival circuit |
| Innate (unconditioned or primary) incentive | Increases approach toward or avoidance of the stimulus in an effort to resolve the challenge or opportunity present |
| Learned (conditioned or secondary) incentive | Invigorates and guides behavior toward situations where the challenge or opportunity present can be resolved |
| 3. Reinforcer | Supports the learning of Pavlovian or instrumental associations |
| Innate (unconditioned or primary) reinforce | Induces the formation of associations with neutral stimuli that occur in its presence (through Pavlovian conditioning) and to the formation of associations with responses that lead to the presentation (appetitive stimuli) or removal (aversive stimuli) of the stimulus (through instrumental conditioning) |
| Learned (conditioned or second-order reinforce | Induces formation of associations with other stimuli (through Pavlovian second-order conditioning) or with goal directed responses (through second-order instrumental conditioning) |
Motivation in the Survival Circuit Scheme
The bottom line of the preceding discussion is that emotion, motivation, and reinforcement are closely intertwined processes. Let’s look a bit more closely at the interrelation between these, focusing on motivation here, and reinforcement in the following section.
Emotion and motivation were traditionally treated as separate topics. Emotion was viewed as a reaction (e.g. a fearful, angry, disgusted, joyful, or sad emotional reaction) to some environmental situation, and motivation as a drive from within (e.g. hunger, thirst, or sexual drive) (e.g. Hull, 1943; Stellar, 1954). In the late 1960s, the emergence of the concept of incentives helped bring these together (Bindra, 1969; Trowill et al, 1969). Bindra (1969), for example, argued that emotion, like motivation, is influenced by internal factors (e.g. hormones) and motivation, like emotion, is impacted by external stimuli (incentives).
Motivation, as assessed behaviorally, involves approach towards desired outcomes and avoidance of undesired outcomes (Tolman, 1932; McClelland et al, 1953; Schneirla 1959, Elliot and Church, 1997; Cofer, 1972; Cofer and Appley, 1964; Miller, 1944; Trowill et al, 1969; Bindra, 1969; Davidson, 1993; Gray, 1982; Lang et al., 1990; Berridge, 2004; Cardinal et al, 2002; Balleine and Dickinson, 1998; Holland and Gallagher, 2004; Gallagher and Holland, 1994). So-called approach/avoidance motivation often occurs in two stages: an anticipatory/exploratory/search for goal objects and the performance and consummatory responses (innate responses controlled by surivial circuits) once goal objects are in reach (Sherrington, 1906; Tinbergen, 1951; Cardinal et al, 2002; Berridge, 1999, 2007).
The anticipatory/exploratory/search phase is guided by incentives (Bindra, 1968; Trowill et al, 1969; Balleine and Dickinson, 1998; Cardinal et al, 2002; Johnson et al, 2009; Petrovich et al, 2002; Berridge, 1999, 2007, 2004; Rolls, 1999, 2005; Glimcher, 2003). Incentives, as noted, are essentially innate or conditioned emotional stimuli; in other words, stimuli with the potential to activate survival circuits.
One of the key discoveries that led to the rise of incentive views was that stimuli that lacked the ability to satisfy needs and reduce drives (for example, the non-nutritive sugar substitute saccharin) were nevertheless motivating (Sheffield and Roby, 1950, Cofer, 1972). A major consequence was that the connection between motivation and specific functional circuits (what we are calling survival circuits) began to be deemphasized. Motivation became a somewhat generic process by which behavior was invigorated and guided towards goals by incentives.
The nucleus accumbens emerged as a key focal point of this general motivational system (Graybiel, 1976; Mogenson et al, 1980; Balleine and Killcross, 1994; Killcross and Robbins, 1993; Everitt et al, 1999; Cardinal et al, 2002; Ikemoto and Panksepp, 1999; Parkinson et al, 1999; Koob, 2009; Sesack and Grace, 2010; Berridge, 2007, 2009; Berridge and Robinson, 1998; Hyman et al, 2006; Nestler, 2004; Kelley, 2004). Behavioral invigoration or energization was said to be a function of dopamine release in the accumbens and incentive processing by the accumbens was thought to guide behavior towards goals. Other areas involved in incentive motivation, such as the obrbito-frontal cortex, are not considered here (see Rolls, 1999, 2005).
A key question is whether motivation is a generic process or whether motivationally specific processing by survival circuits might be significant as well. While there may indeed be generic aspects of motivation (e.g. behavioral invigoration), evidence also supports motivationally specific information processing as well. At the behavioral level, bar pressing for food by a hungry obtain food is facilitated by a conditioned incentive that signals food, is facilitated less by one that signals water, and is inhibited by one that signals shock (Corbit and Balleine, 2005; Hammond, 1970), indicating that motivation is tied to specific survival functions. Lateral hypothalamic circuits that control energy maintenace through feeding modulate nucleus accumbens activity (Sears et al, 2010). The accumbens, once thought to be mainly involved in processing appetitive stimuli is now know to contribute to the processing of aversive incentives as well (Salamone, 1994; Schoenbaum and Setlow, 2003; Roitman et al, 2005; Reynolds and Berridge, 2008). Within the accumbens information processing segregated along motivational lines — aversive and appetitive stimuli are processed separately at the cellular and molecular level (Roitman et al, 2005, 2008). While most work is at the level of appetitive vs. aversive states, it would be important to determine whether incentive related to different appetitive survival circuits (e.g. incentives related to food vs. sex) are processed separately.
Once incentives have guided the organism to goal objects, innate consummatory responses, which are specific to the particular survival circuit and function, are initiated. Their termination essentially ends the survival (emotional) episode — food is eaten, liquid is drunk, sex is consummated, safety is reached.
Before leaving the topic of motivation of instrumental goal-directed behavior it is important to mention that such behaviors, when repeatedly performed in recurring situations, can become habitual and divorced from the actual attainment of the goal. In such cases of stimulus-response habit formation, the neural control switches from the ventral to the dorsal striatum (Everitt and Robbins, 2005; Wickens et al, 2007; Packard and Knowlton, 2002).
Reinforcement and Survival Circuits
Reinforcement and motivation are closely related. Things that motivate are ofen reinforcing, and vice versa. Like motivation, reinforcement was once linked to drive states (Hull, 1943), but drifted towards generic mechanisms over the years. The discovery that behavior could be reinforced by electrical stimulation of brain areas (Olds and Milner, 1954), and the finding that electrical reinforcement could summate with different natural reinforcers (Coons and White, 1977; Conover and Shizgal, 1994), were compatible with a generic mechanism of reinforcement. Similarly, that addictive drugs and natural or electrical reinforcers interact is also consistent with a generic mechanism (Wise, 2006). Further, iunfluential mathematical models of reinforcement (e.g. Rescorla and Wagner, 1972; Sutton and Barto, 1987) explained learning with singular learning rules. The modern paradigmatic example of a generic reinforcement mechanism is the role of dopamine in the striatum as a reward prediction error signal (Schultz, 1997).
Nevertheless, there have from time to time been calls for linking reinforcement more directly to specific neurobiological systems. For example, Glickman and Schiff (1967) poroposed that reinforcement is a facilitation of activity in neural systems that mediate species-specific consummatory acts. In other words, they proposed a link between reinforcement and motivationally-specific survival circuits. It is therefore of great interest that recent work on the role of dopamine as a reward prediction error signal is beginning to recognize the importance of specific motivational states in modulating the effects of dopamine as a reward prediction error signal (Schultz, 2006; Glimcher, 2011).
The expression of reinforcement as a change in the probability that an instrumental response will be performed may well occur via a generic system in which the reinforcer strengthens the response (e.g. via contributions of dopamine in the striatum to reward prediction errors). But, in addition, survival circuit-specific motivational information is likely to contribute at a fundamental level, providing the stimulus with the motivational value that allows it to ultimately engage the more generic mechanisms that strengthen instrumental responses and that motivate their performance.
Reinforcement principles have been used by some authors to classify emotional states (e.g. Gray, 1982; Rolls, 1999, 2005; Cardinal et al, 2002; Hammond, 1970; Mowrer, 1960). In these models various emotions defined in terms of the presentation or removal of reinforcers. Mowrer (1960), for example, proposed a theory in which fear, hope, relief, and disappointment were explained in these terms. Later authors have attempted to account for more conventional emotions (fear, sadness, anger, pleasure, etc) as products of the presentation or removal of reinforcement. This approach suffers from some of the same problems as basic emotions theory in that it focuses on common language words related to human feelings as the way to identify emotion mechanisms in the brain. Perhaps reinforcement, like motivation, might be fruitfully linked to emotional phenomena through the survival circuit conception.
Survival Circuits and Arousal
Survival circuits are engaged in situations in which challenges and/or opportunities exist, in other words what we commonly call emotional or motivated situations. So far we have focused on two major consequence of survival circuit activation. One is the elicitation of specific kinds of hard-wired behavioral reactions. The second is an increase in the probability that instrumental goal-directed actions relevant to the opportunity or challenge will be learned (reinforced) and performed (motivated) — or, if the situation has been experienced by the individual repeatedly in the past, stimulus-response habits may substitute for incentive guided instrumental goal-directed action.
A third consequence of survival circuit activation is “generalized arousal” (Moruzzi and Magoun, 1949; Lindsley, 1951; Schober et al, 2011; Lang, 1994; Pfaff et al, 2008). As originally conceived, generalized arousal was a function of the brainstem reticular activating system (Moruzzi and Magoun, 1949; Lindsley, 1951). Later, the undifferentiated reticular activating system concept gave way to the notion that distinct populations of chemically-specific neurons that underlie sleep-wake cycles and the degree of arousal, attention and vigilance while awake (Jouvet, 1969, 1999; Steriade, 1995, 2004; Jacobs et al, 1990; Jones, 2003; Aston-Jones, 2005; Monti and Jantos, 2008; Sarter et al, 2005; Arnsten and Li, 2005; Robbins, 2005; Nieuwenhuys, 1985; Nishino, 2011). Specifically, neurons that synthesize and release biogenic amines (norepinephrine, dopamine, serotoinin, or acetylcholine) and peptides (e.g. orexins) are believed to make significant contributions to brain arousal. While these transmitters are released in widespread areas of the brain, their effects are especially profound on neurons that are actively engaged in information processing (Aston-Jones et al, 1991; Foote et al, 1983, 1991; Aston-Jones and Bloom, 1981). That is, they modulate rather than initiate neural activity, regulating neuronal excitability and neurotransmission (Schildkraut and Kety, 1967; Hasselmo, 1995; Lopez and Brown, 1992). Also contributing to generalized arousal are peripheral systems that release hormones into the circulation (e.g. cortisol released from the adrenal cortex, adrenergic hormones, epinephrine and nor-epinephrine, from the adrenal medulla; and others) (Axelrod and Reisine, 1984; McEwen, 2009; Sapolsky et al, 1986). Cortisol crosses the blood brain barrier and binds to receptors in a variety of areas, while adrenergic hormones affect the CNS indirectly (McGaugh, 2000). The modulatory effects of central modulators are relatively rapid, whereas the effects of peripheral hormones are considerably slower, allowing the prolongation of the survival state for extended periods of time.
Generalized arousal has played a key role in a number of theories of emotion over the years (e.g. Duffy, 1941; Lindsley, 1951; Schachter and Singer, 1962; Schachter, 1975; Schildkraut and Kety, 1967; Mandler, 1975; Lang, 1994; Robbins, 1997) and is also important in contemporary dimensional theories of emotion (Russell, 1980, 2003; Russell and Barrett, 1999) and some neural models of emotion (e.g. Whalen, 1998; Davis and Whalen, 2001; Gallagher and Holland, 1994; Kapp et al, 1994; Lang and Davis, 2006). However, it is important to ask how generalized arousal is triggered in emotional situations, and how the arousal, once present, affects further processing. Again, the defense circuit is useful for illustrative purposes.
The detection of a threat by defense circuits of the amygdala leads to the activation of central neuromodulatory and peripheral hormonal systems (see Gray, 1993; LeDoux, 1992, 1995; Davis, 1992; Rodrigues et al, 2009;). Thus, central amygdala outputs target dendritic areas of norpeiphrine, dopamine, serotonin, and acetylcholine containing neurons and cause these to release their chemical products in widespread brain areas (e.g. Reyes et al, 2011; Gray, 1993; Weinberger, 1995; Kapp et al, 1994). Central amygdala outputs also target neurons that activate the sympathetic division of the autonomic nervous system, which releases adrenergic hormones from the adrenal medulla, and the hypothalamic-pituitary-adrenal axis, which releases cortisol from the adrenal cortex (Gray, 1993; Talarovicova et al, 2007; Loewy, 1991; Reis and LeDoux, 1987). Threats thus not only elicit specific defense responses but also initiate generalized arousal in the brain and body. Body feedback has played an important role in emotion theory for more than a century (James, 1884; Lange, 1885/1922; Schachter and Singer, 1962; Tomkins, 1962; Adelmann and Zajonc, 1989; Buck, 1980; Damasio, 1994, 1999).
One consequence of this pattern of connectivity is that central and peripheral arousal signals facilitate processing in the survival circuit that triggered the activation of arousal. This establishes a loop in which continued activation of the survival circuit by external stimuli produces continued activation of the modulator release, which in turn facilitates the ability of external stimuli to continue to drive the survival circuit. Indeed, modulators facilitate activity in sensory processing areas (e.g. Hurley et al, 2004), which should enhance attention to external stimuli present during survival circuit activation. Modulators also facilitate processing areas involved in retrieving forming, and storing memories (McGaugh, 2003; Roozendaal et al, 2009). All of these effects are recapitulated in motivational circuits once the initial reaction begins to give way to goal-directed instrumental actions. For example, dopamine contributes to the invigoration or activation of behavior during the exploratory search phase of a motivated state (Berridge 2004; Berridge and Robinson, 1998; Robbins and Everitt, 2007). Norepinephrine, serotoin, acetylcholine, orexins and other modulators also contribute. While arousal is often discussed in terms of generic (generalized) mechanisms, the possibility that some aspects of arousal might be survival circuit specific should also be explored (Pfaff et al, 2008; Schober et al, 2011).
Global Organismic States
Survival circuit activation leads to the triggering of arousal responses in the CNS, and to the potential expression of innate behaviors (depending on the circumstances), as well as expression of autonomic nervous system and hormonal responses in the body. Behavioral, autonomic, and endocrine responses feedback to the brain and also contribute to arousal. In addition, motivational systems are activated, potentially leading to goal-directed behaviors (Figure 3). The overall result of survival circuit-specific activity, motivational activity, and generalized arousal is the establishment of a state in which brain resources are coordinated and monopolized for the purpose of enhancing the organism’s ability to cope with a challenge and/or benefit from opportunities. The organism becomes especially attentive to and sensitive to stimuli relevant to the survival function, memories relevant to the survival function are retrieved, and previously learned instrumental responses relevant to the survival function are potentiated. New learning occurs and new explict memories (via the hippocampus and related cortical areas) and implicit memories (memories stored in the survival circuit) are formed. Such states will be referred to here as global organismic states. The fact that these states are global does not mean that they completely lack specificity. They include survival circuit-specific components, as well as general motivational components that control instrumental behavior and components that control non-specific or generalized arousal within the brain and body.
Figure 3. Consequences of Survival Circuit Activation.
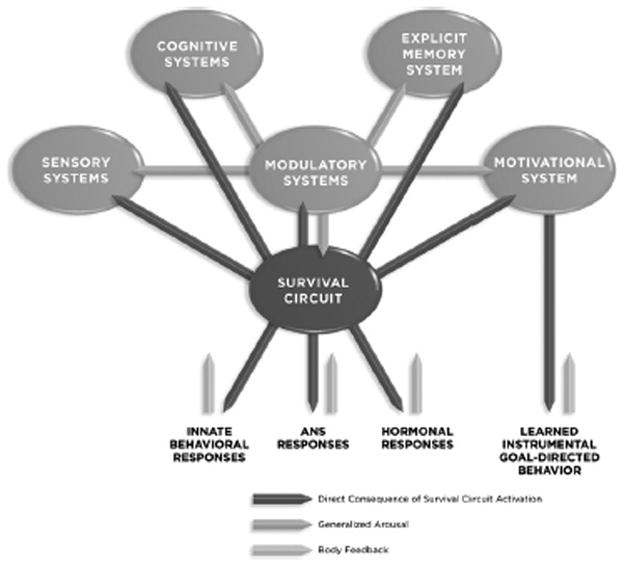
When a survival circuit trigger activates a survival circuit, a number of consequences follow. (1) Innate behavioral responses are potentially activated, as well as autonomic nervous system (ANS) responses and hormonal responses. These each generate feedback to the brain. (2) Neuromodulator systems are activated and begin to regulate excitability and neurotransmission throughout the brain. (3) Goal directed instrumental behavior is initiated by the motivation system. (4) Sensory, cognitive, and explicit memory systems are also affected, leading to enhanced attention to relevant stimuli and the formation of new explicit memories (memories formed by the hippocampus and related cortical areas) and implicit memories (memories formed within the survival circuit).
The notion that emotional and motivated states have profound effects on the brain, recruiting widespread areas into the service of the immediate situation, monopolizing and/or synchronizing brain resources, has been proposed previously (Gallistel, 1980; Maturana and Varela, 1987; Scherer, 2000; LeDoux, 2002, 2008). Particularly relevant is the “central motive state” hypothesis (Morgan, 1943; Hebb, 1949; Bindra, 1969). Yet, the exact nature of global organismic states is poorly understood. In part this is likely attributable to the lack of techniques for assessing neural activity across widespread areas of the brain at a sufficiently detailed level of resolution. Measurement of BOLD activity in the brains of humans or animals with fMRI allows whole brain analysis of functional activity, but lacks spatial resolution at the level of cells and circuits. Use of molecular markers, such as the expression of immeditate early gene activity, in relation to behavior holds promise. Particularly important would be the development of techniques that could provide widespread assessment of changes in body physiology and brain activation and related to survival circuit processing, general-purpose motivational processing, and generalized arousal.
In point of fact, we may need a new kind of neuroscience to come to grips with whole brain states that reflect the activity and inactivity of multiple interacting brain systems. The field has been extremely good at studying individual brain systems but has not done as well in understanding more global functions. If we are to ever truly relate notions of self and personality to the brain, we will need these kinds of approaches (LeDoux, 2002).
Transcending Neuroanatomical Homology: Survival Throughout the Animal World
Invertebrates do not have the same conserved circuits that vertebrates have. However, they face many of the same problems of survival that vertebrates do: they must defend against danger, satisfy energy and nutritional needs, maintain fluid balance and body temperature, and reproduce. As in vertebrates, specific circuits are associated with such functions, though different invertebrates have different nervous systems and different circuits.
The fact invertebrate nervous systems are diverse and differ from the canonical vertebrate nervous system does not mean the invertebrates are irrelevant to understanding survival functions (and thus so-called emotional behavior) in vertebrates. Much progress is being made in understanding innate behaviors related to survival functions such as defense, reproduction and arousal in invertebrates such as Drosophila (Wang et al, 2011; Lebetsky et al, 2009; Dickson, 2008) and C. elegans (McGrath et al, 2009; Pirri and Alkema, 2012; Garrity et al, 2010). In these creatures, as in mammals and other vertebrates, G-protein coupled receptors and their regulators play key roles in modulating neuronal excitability and synaptic strength, and in setting the threshold for behavioral responses to incentives associated with specific motivational/emotional states (Bendesky and Bargmann, 2011). Biogenic amines and their G-protein coupled receptors also play a key role in arousal and behavioral decision-making in Drosophila (Lebetsky et al, 2009) and C. elegans (Bendesky et al, 2011) as in vertebrates (see above). Indeed, it would not be too far fetched to suggest that biogenic amines and their G-protein coupled receptors create global organismic that coordinate the activity multiple subsystems within the nervous system in invertebrates as well as vertebrates.
Much has been also been learned about the cellular, molecular and genetic mechanisms underlying survival based learning in invertebrates. For example, such as Aplysia californica many of the neurotransmitters (e.g. glutamate), neuromodulators (e.g. serotonin, dopamine), intracellular signals (e.g. protein kinase A, map kinase), transcription factors (e.g. cyclic AMP response element binding protein) involved in defense conditioning Aplysia (e.g. Hawkins et al, 2006; Kandel, 2001; Carew and Sutton, 2001; Glanzman, 2010; Mozzachiodi and Byrne, 2010) have been implicated in defense conditioning in the mammalian amygdala (see Johansen et al, 2011). Further, studies in Drosophila have implicated some of the same intracellular signals and transcription factors in defense based learning (Dudai, 1988; Yovell et al, 1992; Yin and Tully, 1996; Margulies et al, 2005).
Similarities at the cellular and molecular level, and presumably at the level of genes that encode these processes, across diverse groups of animals is impressive evidence for conserved principles of organization underlying survival functions. However, an important question is whether there might be more fundamental circuit principles that are instantiated at the microcircuit level in nervous systems that are superficially distinct. If so, the key to understanding the relation of survival functions across invertebrates and vertebrates is likely to involve conserved principles of organization at the microcircuit level rather similarity of anatomical structures or molecules (David Anderson, personal communication). Very interesting examples are emerging from studies of olfactory processing, for which analogies in behaviorally-relevant peripheral odor-encoding and central representation occur using similar organizational principles in anatomically distinct (non-homologous) structures in Drosophila and rodents (see Bargmann, 2006; Sosulski et al, 2011; Wang et al, 2011).
Survival functions instantiated in specific neural circuits likely reflect conserved neural principles. We should at least be amenable to the possibility that defense, reproduction, and other survival functions in humans, may be related to survival functions in invertebrates. This notion is not likely to be surprising to card carrying comparative neurobiologist, but might meet more resistance from researchers who study humans since survival functions account for some fundamental emotional functions in humans, and in humans emotions are often equated with or closely tied to feelings. But the thrust of what has been said here is that survival functions should not be treated as qualitatively differently in humans and other mammals, in mammals and other vertebrates, in vertebrates and invetebrates. As noted earlier, a case can even be made that solutions to fundamental problems of survival are in the final analysis derived form solutions to these problems present primordial single cell organisms.
Survival Circuits and Human Feelings: What Is An Emotional State?
When the term “emotional state” is used, the user typically has the notion of “feeling” in mind. This article is an attempt to redefine the nature of such states, at least the components of such states that are shared across mammalian species (and likely across vertebrates, and to some extent in invertebrates as well). Nevertheless, the history of emotion research and theory is for the most part the history of trying to understand what feelings are and how they come about. It is thus important to comment on the nature of feelings and their relation to survival circuits.
One might be tempted to conclude that global organismic states, or at least the central representation of such sates, constitute neural correlates of feelings. Global organismic states make major contributions to conscious feelings but the two are not the same. Global organismic states are part of the raw material from which certain classes of feelings are constructed (those feelings associated with survival circuit activation). But they could, and likely do, exist, independent of feelings, at least in relation to what humans call feelings. My proposal is that these kinds of feelings (those associated with survival circuit activation) occur in humans when consciousness (a) detects that a survival circuit is active or witnesses the existence of a global organismic state initiated by the activation of a survival circuit in the presence of particular kind of challenge or opportunity and (b) appraises and labels this state. These are not the only kinds of feelings that can occur in humans. Other kinds include feelings associated with higher order or social emotions (guilt, shame, envy, pride) or sensory feelings (a pleasant touch or an annoying itch).
What about other animals? To the extent that non-human organisms have consciousness and cognition, capacities that allow the observation, appraisal, and categorization of survival circuit activity or global organismic states, they can have feelings when survical circuit activity or global organismic states occur. To the extent that the mechanisms of consciousness and cognition differ in different animals (with humans included as an animal), and to the extent that the mechanisms underlying survival circuit or global organismic states themselves differ, feelings will be different. This leaves open the possibility that conscious feelings can be present in other mammals, other vertebrates, or even in invertebrates. But rather than engaging in idle speculation about this, criteria can be offered that can help address the question. Specifically, if we can understand what underlies conscious feelings in humans, we can then search for whether those mechanisms are present, and to what extent they are present, in other animals.
This, you probably noticed, is a different approach from the one advocated earlier for survival circuits. We now ask whether processes in humans are present in other animals. But just as the survival circuit question should be asked about whether mechanisms in other animals are present in humans, the question of whether mechanisms shown to be present in humans are present in other animals seems only addressable in the other direction. We can never know whether another animal has conscious emotional feelings, but we might be able to determine whether the mechanisms that make of consciousness and feelings possible in humans also present in other animals.
The fact is that the brain mechanisms that underlie conscious emotional feelings in humans are still poorly understood. However, this should not stand in the way of understanding survival functions and the states that occur in the brain when the circuits mediating survival functions are activated. There is much work to be done even if we don’t have viable solutions to the problems of conscious feelings.
Research on feelings is complicated because feelings cannot be measured directly. We rely on the outward expression of emotional responses, or on verbal declarations by the person experiencing the feeling, as ways of assessing what that person is feeling. This is true both when scientists do research on emotions, and when people judge emotions in their social interactions with one another.
When not wearing a scientific hat, most of us apply introspectively based concepts to other animals. When a deer freezes to the sound of a shotgun we say it is afraid, and when a kitten purrs or a dog wags its tail, we say it is happy. In other words, we use words that refer to human subjective feelings to describe our interpretation of what is going on in the animal’s mind when it acts in way that has some similarity to the way we act when we have those feelings. Some authors also claim that similarity of behavior is strongly suggestive of similarity at the level of subjective experience (Panksepp, 1998, 2005) or more generally that humans know what an animal feels from observing their behavior (Bekoff, 2007; Masson and McCarthy, 1996). But it’s hard to justify anthropomorphic speculation in science. Panksepp has attempted this (Panksepp, 1982; 1998, 2000; 2005), but few scientists are convinced that this is the way to go, as there is no way to objectively verify what another organism experiences.
So what’s the difference, if any, between attributing feelings to other people and to other animals? There is a strong rationalization for assuming all humans have subjective mental states, such as feelings, that are similar in kind. In the absence of genetic mutations of the nervous system or acquired brain damage, each human possesses the same basic kind of brain, a brain with the same basic neural systems, as every other human. As a result we expect that other people have the same kinds of basic brain functions, and corresponding mental capacities, that we have, and we can assume with some confidence that other people experience the same kinds of feelings we do when we they behave the way we behave when we have those feelings (unless they are being intentionally deceitful). We can therefore fairly comfortably apply our introspections about our own feelings to the mental states of other people on the basis of their behavior.
We should not, however, be so comfortable in talking about the mental states of other species because their brains differ from ours. A key question, of course, is whether their brains differ from ours in ways that matter. In other words, do the brain areas responsible for states of consciousness, such as feelings, differ in humans and other animals?
There is considerable support for the idea that states of consciousness are made possible, at least in part, through the representation of experience in a cognitive workspace involving neocortical areas, including prefrontal (especially dorso-lateral prefrontal) and parietal cortical areas (Crick and Koch, 1990, 2004, Dehaene and Changeux, 2004, Baars, 2005; Frith and Dolan, 1996; Frith et al, 1999; Frith, 2008; Shallice, 1988; Shallice et al, 2008; Goldman-Rakic, xxx). To the extent that feelings are states of consciousness about emotional situations, they should be represented in these cognitive workspace circuits (LeDoux, 1996, 2002, 2008).
The idea proposed here is that conscious feelings result when global organismic states are represented in the cognitive workspace. The basic ingredients of the global organismic state would include information about the stimulus and other aspects of the social and physical environment, the survival circuit the stimulus activates, CNS arousal initiated by the survival circuit, feedback from survival responses that are expressed in the body, and long-term memories (episodic and semantic) about the stimulus and about the resulting state (Fig 4). Thus, in the presence of a survival circuit trigger (a.k.a. an emotional stimulus) the various ingredients would be integrated, and the resulting state categorized by matching the state with long-term memory stores. When this occurs, a conscious feeling of the global organismic state begins to exist. Such a state, having been categorized on the basis of memories of similar states, could be dimensional in nature (just based on arousal and valence) or could take on specific qualities (could be more like what one felt when previously in danger than when frustrated or when enjoying a tasty meal). Labeling of the state with emotion words adds additional specificity to the experience, creating specific feelings (fear, pleasure, disgust, etc).
Figure 4.
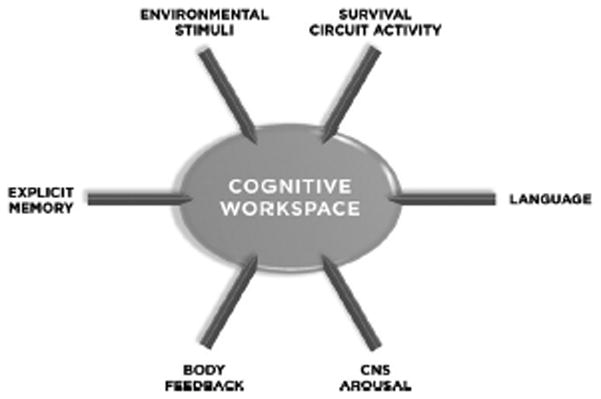
Ingredients of Feelings in a Cognitive Workspace. An emotional feeling is hypothesized to be a representation of a global organismic state initiated by an external stimulus. The representation includes sensory information about the stimulus and the social and physical context, information about the survival circuit that is active, information about CNS arousal, body feedback information, and mnemonic information about the stimulus situation and the state itself. When such a global organismic state is categorized and labeled a conscious feeling of a certain type (e.g. a feeling of fear, pleasure, disgust, etc) results. To the extent that any of these components differ in human and non-human species, the nature of the resulting state would differ as well.
Dorsolateral prefrontal cortex, a key component of the cognitive workspace, is lacking in most other mammals, and is less developed in non-human primates than in humans (Reep, 1984; Braak, 1980; Preuss, 1995; Wise, 2008). In humans, granular prefrontal cortex also has unique cellular features (Semendeferi et al, 2011). Given that feelings are a category of conscious experience, the usual mechanisms of conscious experience should be at work when we have emotional experiences (LeDoux, 1996, 2002; 2008). And given that some of the neural mechanisms involved in conscious representations may be different in humans and other animals, we should be cautious in assuming that the subjectively experienced phenomena that humans label as feelings are experienced by other animals when they engage in behaviors that have some similarity to human emotional behavior. In short, if the circuits that give rise to conscious representations are different in two species, we cannot use behavioral similarity to argue for similarity of conscious feelings functionally. These observations add neurobiological substance to the point famously argued by the philosopher Thomas Nagel. He proposed that only a bat can experience the world like a bat, and only a human can experience the world like a human (Nagel, 1974). We should resist the inclination to apply our introspections to other species.
Also, given that humans are the only organisms with natural language, and language allows a unique mode of information processing in the human brain, we need to be very cautious when we make assumptions about non-human behavior regarding processes that language affects. While the idea that language affects thought and conscious experience (Whorf, 1956) was out of favor for a while, it has reemerged as an important principle in recent times (Lakoff, 1987; Lucy, 1997). One way that language is important is that it allows the semantic categorization of experience, including emotional experience. For example, there are more than 30 words in English for gradations of fear (fear, panic, anxiety, worry, trepidation, consternation, etc) (Marks, 1987). The human brain may be able to categorize emotional states in broad strokes without language but it is unlikely that specific emotions (fear, anger, sadness, joy) could come about without words. Accordingly, lacking language and emotion words, an animal brain cannot partition emotional experience in this way. In short, the language of emotion likely contributes to the experiences one has in emotional situations (Schachter, 1975; Johnson-Laird and Oatley, 1989; Scherer, 1984; Reisenzein, 1995). Indeed, different cultures and their languages express emotions differently (Kitayama and Markus, 1994; Wierzbicka, 1994; Averill, 1980). The dimensional theory of emotion views emotion words as markers in a multidimensional semantic space of feelings (Russell, 1980; Russell and Barrett, 1999). The dimensional theory is incompatible with a basic emotions view, since the latter argues that feelings associated with basic emotions are due to hard-wired circuits, but is compatible with the survival circuit view, which posits indirect and non-obligatory, as opposed to casual, links between survival circuits and feelings.
But the impact of language goes far beyond simple semantics and labeling. We use syntactic processes to evaluate the logical truth of propositional statements. While not all human thought involves propositional statements and logic, syntactic processing provides the human brain and mind with unique features and advantages. Through syntax, the human mind can simulate who will do what to whom in a social situation instantaneously rather than having to learn by trial and error.
So what then might a bat or a rat experience without the kind of cerebral hardware that is characteristic of the human brain? Some have proposed that in addition to full blown feelings that humans talk about, more basic, less differentiated feelings (crude states of positive or negative valence, or maybe even somewhat finer categories based on memory of feelings from the past in similar situations) may exist in other animals. Such states have been called core affects (Panksepp, 1998, 2005; Damasio. 1994, 1999; Barrett et al, 2007; Russell, 2003). While we cannot ask other animals about their feelings, studies of humans can begin to unravel how such states are experienced. Similarity of the structure of these circuits in animals might then provide insight into their function in other animals (Panksepp, 1998, 2005), provided that we do a good job of clarifying the function of the circuits in question (see survival circuit discussion above).
Consciousness and feelings are topics that are best studied in humans. Research on the neural basis of feelings in humans is in its infancy (Panksepp, 1998; 2005; Damasio, 2003; Damasio et al, 2000; Ochsner et al, 2002; Barrett et al, 2007; Rudrauf et al, 2009; Critchley et al, 2004; Pollatos et al, 2007). We will never know what an animal feels. But if we can find neural correlates of conscious feelings in humans (and distinguish them from correlates of unconscious emotional computations in survival circuits), and show that similar correlates exists in homologous brain regions in animals, then some basis for speculating about animal feelings and their nature would exist. While such speculations would be empirically based, they would nevertheless remain speculations.
Future Directions of Research
There are many topics that need further exploration in the study of emotional phenomena in the brain. The following list is meant to point out a few of the many examples, and is not meant to be exhaustive.
The circuits underlying defense in rodents is fairly well characterized and provides a good starting point for further advancement. An important first step is elucidation of the exact relation between innate and learned defense circuits. Paradigms should be devised that directly compare circuits that are activated by innate and learned cues of the same sensory modality and that elicit similar behavioral defense responses (freezing, escape, attack, etc). Comparisons should proceed in stepwise fashion within a species, with variation in the stimulus and response modalities (though mundane, systematic studies are important).
More information is also needed regarding the manner in which external stimuli function as defense triggers, incentives, and reinforcers within defense circuits. Tasks should be developed that can be readily applied across species, but at the same time easily tailored to species-specific factors. Further exploration of whether there are different circuits for defense in different contexts would also be useful.
How do innate and learned stimuli trigger, motivate and reinforce behaviors in non-defense survival circuits? What are the evaluative mechanisms that process such stimuli within specific survival circuits (e.g. energy/nutrition, fluid balance, reproduction, etc)? While incentives and reinforcers have been studied extensively in the context of generic appetitive processes, also of interest is the relation of such stimuli to the specific survival circuit on which they depend.
Can survival circuits be further differentiated? For example, to what extent do different forms of defense utilize different circuits? Beyond classifications based on the sensory modality that detects threats and whether the threat is learned or unlearned, are there different circuits for threats related to conspecifics, predators, injested substances, territory, etc. Similar questions arises for each survival circuit category.
When a given brain area is involved in multiple survival functions, an effort should be made to determine the extent to which underlying cellular mechanisms might make distinct contributions. For example, the LA and BA and nucleus accumbens have been implicated in defensive and appetitive behaviors. As noted above, single unit recordings suggest some independence of responses to aversive and appetitive stimuli in these areas. However, questions remain. Do amygdala or accumbens cells simply encode positive and negative valence or are they tied to more specific survival functions? This is readily explored by examining cellular responses to incentive stimuli related different forms of appetitive motivation. For example, do the cells that respond to incentives related to food, drink and reproduction overlap or are they survival-function specific? Single unit recordings are often restricted to one or a small number of brain areas. New molecular imaging techniques are beginning to allow such differences to be explored at the cellular level across the whole brain. A recent study by Lin et al (2011) showing distinct populations of cells in the ventromedial hypothalamus that contribute to mating and intruder attack is a prime example. Genetic tools can also be used to provide more detailed information about connectivity, including connectivity at the level of brain areas but also between specific cell types. Such approaches have begun to be used but systematic studies are needed.
It is generally assumed that circuits underlying defense, energy, fluid balance, reproduction, thermoregulation, and other survival functions interact, but this has been not been studied to any significant degree. This is a particularly important topic that is best pursued by methods that allow evaluation of concurrent activity in the multiple brain areas, such as fMRI in humans and fMRI and molecular imaging in animals. Studies comparing activity patterns across the whole brain in response processing signals related to various survival circuit functions could provide very important information, especially if animal and human projects use related behavioral paradigms. If homologies are found at the level of brain areas between humans and other mammals, molecular imaging can be used in animal studies to search for unique microcircuits that differentiate between functions and the cellular and synaptic level.
Techniques are needed to assess physiological activity at the cellular level across the whole brain and throughout the body (global organismic states) in the presence of biological significant stimuli (triggers of survival circuit activity or motivating incentives,) and during the performance of innate or learned survival responses as well as goal directed responses. It will be especially useful to develop analytic tools that will be able to separate contributions that are survival circuit specific from more general purpose mechanisms, such as non-specific arousal, and generic aspects of reinforcement and instrumental behavior control.
More comparative work is needed to elucidate similarities and differences in survival functions and circuits between various groups of vertebrates. Particularly pressing are studies of non-mammalian vertebrates.
What is the relation of survival functions in invertebrates to vertebrates? Are there conserved molecules or genes, or conserved computational principles, that underlie anatomically distinct kinds of circuits but that perform similar survival functions in vertebrates and invertebrates?
Explorations of the mechanisms underlying conscious emotional feelings in humans should be pursued more vigorously, including full-blown conscious feelings (feelings of fear, joy, sadness, shame, embarrassment) and coarser conditions (pleasant or unpleasant feelings). This information is important because feelings are such a defining feature of human mental life. But in addition with such information it will then be possible to ask if the required mechanisms of a given kind of conscious state are present in other animals. Even if the mechanisms are present, the results would not allow the conclusion that other animals have feelings that are homologous with human feelings. However, such a result would at least provide a basis for saying whether there is a physical possibility for such states in other animals.
Conclusion
The survival circuit concept provides a conceptualization of an important set of phenomena that are often studied under the rubric of emotion — those phenomena that reflect circuits and functions that are conserved across mammals. Included are circuits responsible for defense, energy/nutrition management, fluid balance, thermoregulation, and procreation, among others. With this approach, key phenomena relevant to the topic of emotion can be accounted for without assuming that the phenomena in question are fundamentally the same or even similar to the phenomena people refer to when they use emotion words to characterize subjective emotional feelings (like feeling afraid, angry, or sad). This approach shifts the focus away from questions about whether emotions that humans consciously experience (feel) are also present in other mammals, and towards questions about the extent to which circuits and corresponding functions that are relevant to the field of emotion and that are present in other mammals are also present in humans. By reassembling ideas about emotion, motivation, reinforcement, and arousal in the context of survival circuits, hypotheses emerge about how organisms negotiate behavioral interactions with the environment in process of dealing with challenges and opportunities in daily life.
References
- Adelmann PK, Zajonc RB. Facial efference and the experience of emotion. Annu Rev Psychol. 1989;40:249–280. doi: 10.1146/annurev.ps.40.020189.001341. [DOI] [PubMed] [Google Scholar]
- Adolphs R. Fear, faces, and the human amygdala. Curr Opin Neurobiol. 2008;18:166–172. doi: 10.1016/j.conb.2008.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amaral DG. The amygdala, social behavior, and danger detection. Ann N Y Acad Sci. 2003;1000:337–347. doi: 10.1196/annals.1280.015. [DOI] [PubMed] [Google Scholar]
- Antoniadis EA, Winslow JT, Davis M, Amaral DG. Role of the primate amygdala in fear-potentiated startle: effects of chronic lesions in the rhesus monkey. J Neurosci. 2007;27:7386–7396. doi: 10.1523/JNEUROSCI.5643-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnold MB. Emotion and Personality. New York: Columbia University Press; 1960. [Google Scholar]
- Arnsten AF, Li BM. Neurobiology of executive functions: catecholamine influences on prefrontal cortical functions. Biol Psychiatry. 2005;57:1377–1384. doi: 10.1016/j.biopsych.2004.08.019. [DOI] [PubMed] [Google Scholar]
- Aston-Jones G. Brain structures and receptors involved in alertness. Sleep Med. 2005;6(Suppl 1):S3–7. doi: 10.1016/s1389-9457(05)80002-4. [DOI] [PubMed] [Google Scholar]
- Aston-Jones G, Bloom FE. Norepinephrine-containing locus coeruleus neurons in behaving rats exhibit pronounced responses to non-noxious environmental stimuli. J Neurosci. 1981;1:887–900. doi: 10.1523/JNEUROSCI.01-08-00887.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aston-Jones G, Shipley MT, Chouvet G, Ennis M, van Bockstaele E, Pieribone V, Shiekhattar R, Akaoka H, Drolet G, Astier B, Charlety P, Valentino RJ, Williams JT. Afferent regulation of locus coeruleus neurons: Anatomy, physiology, and pharmacology. Progress in Brain Research. 1991;88:47–75. doi: 10.1016/s0079-6123(08)63799-1. [DOI] [PubMed] [Google Scholar]
- Averill J. Emotion and anxiety: Sociocultural, biological, and psychological determinants. In: Rorty AO, editor. Explaining emotions. Berkeley: University of California Press; 1980. [Google Scholar]
- Axelrod J, Reisine TD. Stress hormones: their interaction and regulation. Science. 1984;224:452–459. doi: 10.1126/science.6143403. [DOI] [PubMed] [Google Scholar]
- Baars BJ. Global workspace theory of consciousness: toward a cognitive neuroscience of human experience. Prog Brain Res. 2005;150:45–53. doi: 10.1016/S0079-6123(05)50004-9. [DOI] [PubMed] [Google Scholar]
- Balleine B, Killcross S. Effects of ibotenic acid lesions of teh nucleus accumbens on instrumental action. Behavioral Brain Research. 1994;65:181–193. doi: 10.1016/0166-4328(94)90104-x. [DOI] [PubMed] [Google Scholar]
- Balleine BW, Dickinson A. Goal-directed instrumental action: contingency and incentive learning and their cortical substrates. Neuropharmacology. 1998;37:407–419. doi: 10.1016/s0028-3908(98)00033-1. [DOI] [PubMed] [Google Scholar]
- Balleine BW, Killcross S. Parallel incentive processing: an integrated view of amygdala function. Trends Neurosci. 2006;29:272–279. doi: 10.1016/j.tins.2006.03.002. [DOI] [PubMed] [Google Scholar]
- Bargmann CI. Comparative chemosensation from receptors to ecology. Nature. 2006;444:295–301. doi: 10.1038/nature05402. [DOI] [PubMed] [Google Scholar]
- Barrett LF. Are Emotions Natural Kinds? Perspectives on Psychological Science. 2006a;1:28–58. doi: 10.1111/j.1745-6916.2006.00003.x. [DOI] [PubMed] [Google Scholar]
- Barrett LF. Solving the emotion paradox: categorization and the experience of emotion. Pers Soc Psychol Rev. 2006b;10:20–46. doi: 10.1207/s15327957pspr1001_2. [DOI] [PubMed] [Google Scholar]
- Barrett LF, Lindquist KA, Bliss-Moreau E, Duncan S, Gendron M, Mize J, Brennan L. Of Mice and Men: Natural Kinds of Emotions in the Mammalian Brain? A Response to Panksepp and Izard. Perspect Psychol Sci. 2007;2:297–311. doi: 10.1111/j.1745-6916.2007.00046.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baxter MG, Murray EA. The amygdala and reward. Nat Rev Neurosci. 2002;3:563–573. doi: 10.1038/nrn875. [DOI] [PubMed] [Google Scholar]
- Bekoff M. The Emotional Lives of Animals: A leading scientist explores animal joy, sorrow, and empathy - and why they matter. Novato, CA: New World Library; 2007. [Google Scholar]
- Belova MA, Paton JJ, Salzman CD. Moment-to-moment tracking of state value in the amygdala. J Neurosci. 2008;28:10023–10030. doi: 10.1523/JNEUROSCI.1400-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belova MA, Paton JJ, Morrison SE, Salzman CD. Expectation modulates neural responses to pleasant and aversive stimuli in primate amygdala. Neuron. 2007;55:970–984. doi: 10.1016/j.neuron.2007.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bendesky A, Bargmann CI. Genetic contributions to behavioural diversity at the gene-environment interface. Nat Rev Genet. 2011;12:809–820. doi: 10.1038/nrg3065. [DOI] [PubMed] [Google Scholar]
- Bendesky A, Tsunozaki M, Rockman MV, Kruglyak L, Bargmann CI. Catecholamine receptor polymorphisms affect decision-making in C. elegans. Nature. 2011;472:313–318. doi: 10.1038/nature09821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernard C. Lecons sur les phenomenes de la vie communs aux animaus et aux vegetaux. Paris: Bailliere; 1878–1879. [Google Scholar]
- Berridge KC. Pleasure, pain, desire, and dread: hidden core processes of emotion. In: Kahnemann D, Diener E, Schwarz N, editors. Well-Being. New York: Russell Sage Foundation; 1999. [Google Scholar]
- Berridge KC. Motivation concepts in behavioral neuroscience. Physiol Behav. 2004;81:179–209. doi: 10.1016/j.physbeh.2004.02.004. [DOI] [PubMed] [Google Scholar]
- Berridge KC. The debate over dopamine’s role in reward: the case for incentive salience. Psychopharmacology (Berl) 2007;191:391–431. doi: 10.1007/s00213-006-0578-x. [DOI] [PubMed] [Google Scholar]
- Berridge KC. ‘Liking’ and ‘wanting’ food rewards: brain substrates and roles in eating disorders. Physiol Behav. 2009;97:537–550. doi: 10.1016/j.physbeh.2009.02.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berridge KC, Robinson TE. What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? Brain Res Brain Res Rev. 1998;28:309–369. doi: 10.1016/s0165-0173(98)00019-8. [DOI] [PubMed] [Google Scholar]
- Bindra D. Neuropsychological interpretation of the effects of drive and incentive-motivation on general activity and instrumental behavior. Psychol Rev. 1968;75:1–22. [Google Scholar]
- Bindra D. A unfied interpretation of emotion and motivation. Ann N Y Acad Sci. 1969;159:1071–1083. doi: 10.1111/j.1749-6632.1969.tb12998.x. [DOI] [PubMed] [Google Scholar]
- Blanchard CD, Blanchard RJ. Innate and conditioned reactions to threat in rats with amygdaloid lesions. Journal of Comparative Physiological Psychology. 1972;81:281–290. doi: 10.1037/h0033521. [DOI] [PubMed] [Google Scholar]
- Blanchard DC, Blanchard RJ, Rodgers RJ. Pharmacological and neural control of anti-predator defense in the rat. Aggressive Behavior. 1990;16:165–175. [Google Scholar]
- Blanchard DC, Li CI, Hubbard D, Markham CM, Yang M, Takahashi LK, Blanchard RJ. Dorsal premammillary nucleus differentially modulates defensive behaviors induced by different threat stimuli in rats. Neurosci Lett. 2003;345:145–148. doi: 10.1016/s0304-3940(03)00415-4. [DOI] [PubMed] [Google Scholar]
- Blaustein JD. Neuroendocrine regulation of feminine sexual behavior: lessons from rodent models and thoughts about humans. Annu Rev Psychol. 2008;59:93–118. doi: 10.1146/annurev.psych.59.103006.093556. [DOI] [PubMed] [Google Scholar]
- Bolles RC, Fanselow MS. A perceptual-defensive-recuperative model of fear and pain. Behavioral and Brain Sciences. 1980;3:291–323. [Google Scholar]
- Bordi F, LeDoux J. Sensory tuning beyond the sensory system: an initial analysis of auditory response properties of neurons in the lateral amygdaloid nucleus and overlying areas of the striatum. J Neurosci. 1992;12:2493–2503. doi: 10.1523/JNEUROSCI.12-07-02493.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowlby J. Attachment. Vol. 1. New York: Basic Books; 1969. Attachment and Loss. [Google Scholar]
- Braak E. On the structure of IIIab-pyramidal cells in the human isocortex. A Golgi and electron microscopical study with special emphasis on the proximal axon segment. J Hirnforsch. 1980;21:437–442. [PubMed] [Google Scholar]
- Brown SM, Hariri AR. Neuroimaging studies of serotonin gene polymorphisms: exploring the interplay of genes, brain, and behavior. Cogn Affect Behav Neurosci. 2006;6:44–52. doi: 10.3758/cabn.6.1.44. [DOI] [PubMed] [Google Scholar]
- Bruce LL, Neary TJ. The limbic system of tetrapods: a comparative analysis of cortical and amygdalar populations. Brain Behav Evol. 1995;46:224–234. doi: 10.1159/000113276. [DOI] [PubMed] [Google Scholar]
- Buchel C, Dolan RJ. Classical fear conditioning in functional neuroimaging. Curr Opin Neurobiol. 2000;10:219–223. doi: 10.1016/s0959-4388(00)00078-7. [DOI] [PubMed] [Google Scholar]
- Buck R. Nonverbal behavior and the theory of emotion: the facial feedback hypothesis. J Pers Soc Psychol. 1980;38:811–824. doi: 10.1037//0022-3514.38.5.811. [DOI] [PubMed] [Google Scholar]
- Butler AB, Hodos W. Comparative Vertebrate Neuroanatomy: Evolution and Adaptation. 2. Hoboken: John Wiley & Sons, Inc; 2005. [Google Scholar]
- Cain CK, LeDoux JE. Escape from fear: a detailed behavioral analysis of two atypical responses reinforced by CS termination. J Exp Psychol Anim Behav Process. 2007;33:451–463. doi: 10.1037/0097-7403.33.4.451. [DOI] [PubMed] [Google Scholar]
- Canli T, Sivers H, Whitfield SL, Gotlib IH, Gabrieli JD. Amygdala response to happy faces as a function of extraversion. Science. 2002;296:2191. doi: 10.1126/science.1068749. [DOI] [PubMed] [Google Scholar]
- Cannon WB. The James-Lange theory of emotions: a critical examination and an alternative theory. Am J Psychol. 1927;39:106–124. [PubMed] [Google Scholar]
- Cannon WB. Bodily changes in pain, hunger, fear, and rage. New York: Appleton; 1929. [Google Scholar]
- Cannon WB. Again the James-Lange and the thalamic theories of emotion. Psychol Rev. 1931;38:281–295. [Google Scholar]
- Canteras NS. The medial hypothalamic defensive system: hodological organization and functional implications. Pharmacol Biochem Behav. 2002;71:481–491. doi: 10.1016/s0091-3057(01)00685-2. [DOI] [PubMed] [Google Scholar]
- Canteras NS, Resstel LB, Bertoglio LJ, de Carobrez AP, Guimaraes FS. Neuroanatomy of anxiety. Curr Top Behav Neurosci. 2010;2:77–96. doi: 10.1007/7854_2009_7. [DOI] [PubMed] [Google Scholar]
- Cardinal RN, Parkinson JA, Hall J, Everitt BJ. Emotion and motivation: the role of the amygdala, ventral striatum, and prefrontal cortex. Neurosci Biobehav Rev. 2002;26:321–352. doi: 10.1016/s0149-7634(02)00007-6. [DOI] [PubMed] [Google Scholar]
- Carew TJ, Sutton MA. Molecular stepping stones in memory consolidation. Nat Neurosci. 2001;4:769–771. doi: 10.1038/90458. [DOI] [PubMed] [Google Scholar]
- Casey BJ, Ruberry EJ, Libby V, Glatt CE, Hare T, Soliman F, Duhoux S, Frielingsdorf H, Tottenham N. Transitional and translational studies of risk for anxiety. Depress Anxiety. 2011;28:18–28. doi: 10.1002/da.20783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chi CC, Flynn JP. Neural pathways associated with hypothalamically elicited attach behavior in cats. Science. 1971;171:703–706. doi: 10.1126/science.171.3972.703. [DOI] [PubMed] [Google Scholar]
- Choi GB, Dong HW, Murphy AJ, Valenzuela DM, Yancopoulos GD, Swanson LW, Anderson DJ. Lhx6 delineates a pathway mediating innate reproductive behaviors from the amygdala to the hypothalamus. Neuron. 2005;46:647–660. doi: 10.1016/j.neuron.2005.04.011. [DOI] [PubMed] [Google Scholar]
- Choi JS, Brown TH. Central amygdala lesions block ultrasonic vocalization and freezing as conditional but not unconditional responses. J Neurosci. 2003;23:8713–8721. doi: 10.1523/JNEUROSCI.23-25-08713.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cofer C, Appley M. Motivation, Theory and Research. New York: John Wiley & Sons, Inc; 1964. [Google Scholar]
- Cohen DH. The neural pathways and informational flow mediating a conditioned autonomic response. In: Di Cara LV, editor. Limbic and Autonomic Nervous System Research. New York: Plenum Press; 1974. pp. 223–275. [Google Scholar]
- Conover KL, Shizgal P. Differential effects of postingestive feedback on the reward value of sucrose and lateral hypothalamic stimulation in rats. Behav Neurosci. 1994;108:559–572. doi: 10.1037//0735-7044.108.3.559. [DOI] [PubMed] [Google Scholar]
- Coons EE, White HA. Tonic properties of orosensation and the modulation of intracranial self-stimulation: the CNS weighting of external and internal factors governing reward. Ann NY Acad Sci. 1977;290:158–179. doi: 10.1111/j.1749-6632.1977.tb39725.x. [DOI] [PubMed] [Google Scholar]
- Corbit LH, Balleine BW. Double dissociation of basolateral and central amygdala lesions on the general and outcome-specific forms of pavlovian-instrumental transfer. J Neurosci. 2005;25:962–970. doi: 10.1523/JNEUROSCI.4507-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cousens G, Otto T. Both pre- and posttraining excitotoxic lesions of the basolateral amygdala abolish the expression of olfactory and contextual fear conditioning. Behav Neurosci. 1998;112:1092–1103. doi: 10.1037//0735-7044.112.5.1092. [DOI] [PubMed] [Google Scholar]
- Crick F, Koch C. Toward a neurobiological theory of consciousness. Neurosciences. 1990;2:263–275. [Google Scholar]
- Crick FC, Koch C. A Framework for Consciousness. In: Gazzaniga MS, editor. The Cognitive Neurosciences. 3. Cambridge: MIT Press; 2004. pp. 1133–1143. [Google Scholar]
- Critchley HD, Wiens S, Rotshtein P, Ohman A, Dolan RJ. Neural systems supporting interoceptive awareness. Nat Neurosci. 2004;7:189–195. doi: 10.1038/nn1176. [DOI] [PubMed] [Google Scholar]
- Damasio A. Descarte’s error: Emotion, reason, and the human brain. New York: Gosset/Putnam; 1994. [Google Scholar]
- Damasio A. Feelings of emotion and the self. Ann N Y Acad Sci. 2003;1001:253–261. doi: 10.1196/annals.1279.014. [DOI] [PubMed] [Google Scholar]
- Damasio AR. The feeling of what happens: Body and emotion in the making of consciousness. New York: Harcourt Brace; 1999. [Google Scholar]
- Damasio AR, Grabowski TJ, Bechara A, Damasio H, Ponto LL, Parvizi J, Hichwa RD. Subcortical and cortical brain activity during the feeling of self- generated emotions. Nat Neurosci. 2000;3:1049–1056. doi: 10.1038/79871. [DOI] [PubMed] [Google Scholar]
- Darwin C. The expression of the emotions in man and animals. London: Fontana Press; 1872. [Google Scholar]
- Davidson RJ. Cerebral asymmetry and emotion: Conceptual and methodological conundrums. Cognition and Emotion. 1993;7:115–138. [Google Scholar]
- Davies DC, Martinez-Garcia F, Lanuza E, Novejarque A. Striato-amygdaloid transition area lesions reduce the duration of tonic immobility in the lizard Podarcis hispanica. Brain Res Bull. 2002;57:537–541. doi: 10.1016/s0361-9230(01)00687-6. [DOI] [PubMed] [Google Scholar]
- Davis FC, Somerville LH, Ruberry EJ, Berry AB, Shin LM, Whalen PJ. A tale of two negatives: differential memory modulation by threat-related facial expressions. Emotion. 2011;11:647–655. doi: 10.1037/a0021625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis M. The role of the amygdala in conditioned fear. In: Aggleton JP, editor. The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction. NY: Wiley-Liss, Inc; 1992. pp. 255–306. [Google Scholar]
- Davis M, Whalen PJ. The amygdala: vigilance and emotion. Mol Psychiatry. 2001;6:13–34. doi: 10.1038/sj.mp.4000812. [DOI] [PubMed] [Google Scholar]
- Davis M, Walker DL, Lee Y. Amygdala and bed nucleus of the stria terminalis: differential roles in fear and anxiety measured with the acoustic startle reflex. Philosophical Transactions of the Royal Society of London Series B: Biological Sciences. 1997;352:1675–1687. doi: 10.1098/rstb.1997.0149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Oca BM, Fanselow MS. Amygdala and periaqueductal gray lesions only partially attenuate unconditional defensive responses in rats exposed to a cat. Integr Physiol Behav Sci. 2004;39:318–333. doi: 10.1007/BF02734170. [DOI] [PubMed] [Google Scholar]
- de Oca BM, Minor TR, Fanselow MS. Brief flight to a familiar enclosure in response to a conditional stimulus in rats. J Gen Psychol. 2007;134:153–172. doi: 10.3200/GENP.134.2.153-172. [DOI] [PubMed] [Google Scholar]
- De Oca BM, DeCola JP, Maren S, Fanselow MS. Distinct regions of the periaqueductal gray are involved in the acquisition and expression of defensive responses. J Neurosci. 1998;18:3426–3432. doi: 10.1523/JNEUROSCI.18-09-03426.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dehaene S, Changeux J-P. Neural Mechanisms for Access to Consciousness. In: Gazzaniga MS, editor. The Cognitive Neurosciences. 3. Cambridge: MIT Press; 2004. pp. 1145–1157. [Google Scholar]
- Dickson BJ. Wired for sex: the neurobiology of Drosophila mating decisions. Science. 2008;322:904–909. doi: 10.1126/science.1159276. [DOI] [PubMed] [Google Scholar]
- Dielenberg RA, Hunt GE, McGregor IS. “When a rat smells a cat”: the distribution of Fos immunoreactivity in rat brain following exposure to a predatory odor. Neuroscience. 2001;104:1085–1097. doi: 10.1016/s0306-4522(01)00150-6. [DOI] [PubMed] [Google Scholar]
- Dixon BA. Animal Emotion. Ethics and the Environment. 2001;6:22–30. [Google Scholar]
- Dudai Y. Neurogenetic dissection of learning and short-term memory in Drosophila. Annu Rev Neurosci. 1988;11:537–563. doi: 10.1146/annurev.ne.11.030188.002541. [DOI] [PubMed] [Google Scholar]
- Duffy E. Emotion: an example of the need for reorientation in psychology. Psychol Rev. 1934;41:184–198. [Google Scholar]
- Duffy E. An explanation of “emotional” phemomena without the use of the concept “emotion”. Journal of General Psychology. 1941;25:283–293. [Google Scholar]
- Ekman P. Universals and Cultural Differences in Facial Expressions of Emotions. In: Cole J, editor. Nebraska Symposium on Motivation. Lincoln, Nebraska: University of Nebraska Press; 1972. pp. 207–283. [Google Scholar]
- Ekman P. Biological and cultural contributions to body and facial movement in the expression of emotions. In: Rorty AO, editor. Explaining Emotions. Berkeley: University of California Press; 1980. [Google Scholar]
- Ekman P. Expression and nature of emotion. In: Scherer K, Ekman P, editors. Approaches to Emotion. Hillsdale, NJ: Earlbaum; 1984. pp. 319–343. [Google Scholar]
- Ekman P. An argument for basic emotions. Cognition and Emotion. 1992;6:169–200. [Google Scholar]
- Ekman P. Basic Emotions. In: Dalgleish T, Power M, editors. Handbook of Cognition and Emotion. Chichester: John Wiley and Sons, Co; 1999a. pp. 45–60. [Google Scholar]
- Ekman P. Annotated update of Charles Darwin’s “The Expression of the Emotions in Man and Animals”. New York: Harpercollins; 1999b. [Google Scholar]
- Ekman P, Davidson R. The nature of emotion: fundamental questions. New York: Oxford University Press; 1994. [Google Scholar]
- Elliot A, Church M. A hierarchical model of approach motivation and avoidance achievement motivation. Journal of Personality and Social Psychology. 1997;72:218–232. doi: 10.1037//0022-3514.76.4.628. [DOI] [PubMed] [Google Scholar]
- Ellsworth PC. William James and emotion: is a century of fame worth a century of misunderstanding? Psychol Rev. 1994;101:222–229. doi: 10.1037/0033-295x.101.2.222. [DOI] [PubMed] [Google Scholar]
- Elmquist JK, Coppari R, Balthasar N, Ichinose M, Lowell BB. Identifying hypothalamic pathways controlling food intake, body weight, and glucose homeostasis. J Comp Neurol. 2005;493:63–71. doi: 10.1002/cne.20786. [DOI] [PubMed] [Google Scholar]
- Estes WK, Skinner BF. Some quantitative properties of anxiety. Journal of Experimental Psychology. 1941;29:390–400. [Google Scholar]
- Everitt BJ, Robbins TW. Neural systems of reinforcement for drug addiction: from actions to habits to compulsion. Nat Neurosci. 2005;8:1481–1489. doi: 10.1038/nn1579. [DOI] [PubMed] [Google Scholar]
- Everitt BJ, Cardinal RN, Parkinson JA, Robbins TW. Appetitive behavior: impact of amygdala-dependent mechanisms of emotional learning. Ann N Y Acad Sci. 2003;985:233–250. [PubMed] [Google Scholar]
- Everitt BJ, Parkinson JA, Olmstead MC, Arroyo M, Robledo P, Robbins TW. Associative processes in addiction and reward. The role of amygdala- ventral striatal subsystems. Ann N Y Acad Sci. 1999;877:412–438. doi: 10.1111/j.1749-6632.1999.tb09280.x. [DOI] [PubMed] [Google Scholar]
- Fanselow MS. Neural organization of the defensive behavior system responsible for fear. Psychonomic Bulletin and Review. 1994;1:429–438. doi: 10.3758/BF03210947. [DOI] [PubMed] [Google Scholar]
- Fanselow MS, Poulos AM. The neuroscience of mammalian associative learning. Annu Rev Psychol. 2005;56:207–234. doi: 10.1146/annurev.psych.56.091103.070213. [DOI] [PubMed] [Google Scholar]
- Ferris CF, Stolberg T, Kulkarni P, Murugavel M, Blanchard R, Blanchard DC, Febo M, Brevard M, Simon NG. Imaging the neural circuitry and chemical control of aggressive motivation. BMC Neurosci. 2008;9:111. doi: 10.1186/1471-2202-9-111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fitzsimons JT. The physiology of thirst and sodium appetite. Monogr Physiol Soc. 1979:1–572. [PubMed] [Google Scholar]
- Flynn JP. The neural basis of aggression in cats. In: Glass DG, editor. Biology and Behavior: Neurophysiology and Emotion. New York: Rockefeller University Press and Russell Sage Foundation; 1967. pp. 40–60. [Google Scholar]
- Foote SL, Bloom FE, Aston-Jones G. Nucleus locus ceruleus: new evidence of anatomical and physiological specificity. Physiol Rev. 1983;63:844–914. doi: 10.1152/physrev.1983.63.3.844. [DOI] [PubMed] [Google Scholar]
- Foote SL, Berridge CW, Adams LM, Pineda JA. Electrophysiological evidence for the involvement of the locus coeruleus in alerting, orienting, and attending. Prog Brain Res. 1991;88:521–532. doi: 10.1016/s0079-6123(08)63831-5. [DOI] [PubMed] [Google Scholar]
- Frijda N. The Emotions. Cambridge: Cambridge University Press; 1986. [Google Scholar]
- Frith C, Dolan R. The role of the prefrontal cortex in higher cognitive functions. Brain Res Cogn Brain Res. 1996;5:175–181. doi: 10.1016/s0926-6410(96)00054-7. [DOI] [PubMed] [Google Scholar]
- Frith C, Perry R, Lumer E. The neural correlates of conscious experience: an experimental framework. Trends in Cognitive Sciences. 1999;3:105–114. doi: 10.1016/s1364-6613(99)01281-4. [DOI] [PubMed] [Google Scholar]
- Frith CD. The social functions of consciousness. In: Weiskrantz L, Davies M, editors. Frontiers of Consciousness: Chichele lectures. Oxford: Oxford University Press; 2008. pp. 225–244. [Google Scholar]
- Fusar-Poli P, Placentino A, Carletti F, Landi P, Allen P, Surguladze S, Benedetti F, Abbamonte M, Gasparotti R, Barale F, Perez J, McGuire P, Politi P. Functional atlas of emotional faces processing: a voxel-based meta-analysis of 105 functional magnetic resonance imaging studies. J Psychiatry Neurosci. 2009;34:418–432. [PMC free article] [PubMed] [Google Scholar]
- Gallagher M, Holland PC. The amygdala complex: multiple roles in associative learning and attention. Proc Natl Acad Sci U S A. 1994;91:11771–11776. doi: 10.1073/pnas.91.25.11771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallistel R. The organization of action: A new synthesis. Hillsdale, NJ: Erlbaum; 19800. [Google Scholar]
- Garcia J, McGowan BK, Ervin FR, Koelling RA. Cues: their relative effectiveness as a function of the reinforcer. Science. 1968;160:794–795. doi: 10.1126/science.160.3829.794. [DOI] [PubMed] [Google Scholar]
- Garrity PA, Goodman MB, Samuel AD, Sengupta P. Running hot and cold: behavioral strategies, neural circuits, and the molecular machinery for thermotaxis in C. elegans and Drosophila. Genes Dev. 2010;24:2365–2382. doi: 10.1101/gad.1953710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glanzman DL. Common mechanisms of synaptic plasticity in vertebrates and invertebrates. Curr Biol. 2010;20:R31–36. doi: 10.1016/j.cub.2009.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glickman SE, Schiff BB. A biological theory of reinforcement. Psychol Rev. 1967;74:81–109. doi: 10.1037/h0024290. [DOI] [PubMed] [Google Scholar]
- Glimcher PW. Decisions, Uncertainty, and the Brain: The science of neuroeconomics. Cambridge: MIT Press; 2003. [Google Scholar]
- Glimcher PW. Understanding dopamine and reinforcement learning: the dopamine reward prediction error hypothesis. Proc Natl Acad Sci U S A. 2011;108(Suppl 3):15647–15654. doi: 10.1073/pnas.1014269108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray JA. The Neuropsychology of Anxiety. New York: Oxford University Press; 1982. [Google Scholar]
- Gray JA. The Psychology of Fear and Stress. New York: Cambridge University Press; 1987. [Google Scholar]
- Gray TS. Amygdaloid CRF pathways. Role in autonomic, neuroendocrine, and behavioral responses to stress. Annal NY Academy of Science. 1993;697:53–60. doi: 10.1111/j.1749-6632.1993.tb49922.x. [DOI] [PubMed] [Google Scholar]
- Graybiel A. Input-output anatomy of the basal ganglia. Toronto, Canada: Lecture at the Society for Neuroscience; 1976. [Google Scholar]
- Griffiths PE. What Emotions Really Are: The problem of psychological categories. Chicago: University of Chicago Press; 1997. [Google Scholar]
- Griffiths PE. Is emotion a natural kind? In: Solomon RC, editor. Thinking about feeling: Contemporary philosophers on emotions. Oxford: Oxford University Press; 2004. pp. 233–249. [Google Scholar]
- Hamann SB, Ely TD, Hoffman JM, Kilts CD. Ecstasy and agony: activation of the human amygdala in positive and negative emotion. Psychol Sci. 2002;13:135–141. doi: 10.1111/1467-9280.00425. [DOI] [PubMed] [Google Scholar]
- Hammond LJ. Conditional emotional states. In: Black P, editor. Physiological Correlates of Emotion. New York: Academic Press; 1970. [Google Scholar]
- Hasselmo ME. Neuromodulation and cortical function: modeling the physiological basis of behavior. Behav Brain Res. 1995;67:1–27. doi: 10.1016/0166-4328(94)00113-t. [DOI] [PubMed] [Google Scholar]
- Hawkins RD, Kandel ER, Bailey CH. Molecular mechanisms of memory storage in Aplysia. Biol Bull. 2006;210:174–191. doi: 10.2307/4134556. [DOI] [PubMed] [Google Scholar]
- Hebb DO. The Organization of Behavior. New York: John Wiley and Sons; 1949. [Google Scholar]
- Hess WR. Functional organization of the diencephalon. New York: Grune and Stratton; 1954. [Google Scholar]
- Holland PC, Rescorla RA. Second-order conditioning with food unconditioned stimulus. J Comp Physiol Psychol. 1975;88:459–467. doi: 10.1037/h0076219. [DOI] [PubMed] [Google Scholar]
- Holland PC, Gallagher M. Amygdala-frontal interactions and reward expectancy. Curr Opin Neurobiol. 2004;14:148–155. doi: 10.1016/j.conb.2004.03.007. [DOI] [PubMed] [Google Scholar]
- Hull CL. Principles of behavior. New York: Appleton-Century-Crofts; 1943. [Google Scholar]
- Hurley LM, Devilbiss DM, Waterhouse BD. A matter of focus: monoaminergic modulation of stimulus coding in mammalian sensory networks. Curr Opin Neurobiol. 2004;14:488–495. doi: 10.1016/j.conb.2004.06.007. [DOI] [PubMed] [Google Scholar]
- Hyman SE, Malenka RC, Nestler EJ. Neural mechanisms of addiction: the role of reward-related learning and memory. Annu Rev Neurosci. 2006;29:565–598. doi: 10.1146/annurev.neuro.29.051605.113009. [DOI] [PubMed] [Google Scholar]
- Ikemoto S, Panksepp J. The role of nucleus accumbens dopamine in motivated behavior: a unifying interpretation with special reference to reward-seeking. Brain Res Brain Res Rev. 1999;31:6–41. doi: 10.1016/s0165-0173(99)00023-5. [DOI] [PubMed] [Google Scholar]
- Izard CE. Basic emotions, relations among emotions, and emotion-cognition relations. Psychological Review. 1992;99:561–565. doi: 10.1037/0033-295x.99.3.561. [DOI] [PubMed] [Google Scholar]
- Izard CE. Basic emotions, natural kinds, emotion schemas, and a new paradigm. Perspectives on Psychological Science. 2007;2:260–280. doi: 10.1111/j.1745-6916.2007.00044.x. [DOI] [PubMed] [Google Scholar]
- Jacobs BL, Wilkinson LO, Fornal CA. The role of brain serotonin. A neurophysiologic perspective. Neuropsychopharmacology. 1990;3:473–479. [PubMed] [Google Scholar]
- James W. What is an emotion? Mind. 1884;9:188–205. [Google Scholar]
- Jarymowicz M. Emotions as evaluative processes from primary affects to appraisals based on deliberative thinking. In: Blachnio A, Przepiorka A, editors. Closer to emotions. Lublin: Wydawnictwo KUL; 2009. pp. 55–72. [Google Scholar]
- Johansen JP, Cain CK, Ostroff LE, LeDoux JE. Molecular mechanisms of fear learning and memory. Cell. 2011;147:509–524. doi: 10.1016/j.cell.2011.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson-Laird PN, Oatley K. The language of emotions: An analysis of a semantic field. Cognition and Emotion. 1989;3:81–123. [Google Scholar]
- Johnson AK. The sensory psychobiology of thirst and salt appetite. Med Sci Sports Exerc. 2007;39:1388–1400. doi: 10.1249/mss.0b013e3180686de8. [DOI] [PubMed] [Google Scholar]
- Johnson AW, Gallagher M, Holland PC. The basolateral amygdala is critical to the expression of pavlovian and instrumental outcome-specific reinforcer devaluation effects. J Neurosci. 2009;29:696–704. doi: 10.1523/JNEUROSCI.3758-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones BE. Arousal systems. Front Biosci. 2003;8:s438–451. doi: 10.2741/1074. [DOI] [PubMed] [Google Scholar]
- Jouvet M. Biogenic amines and the states of sleep. Science. 1969;163:32–41. doi: 10.1126/science.163.3862.32. [DOI] [PubMed] [Google Scholar]
- Jouvet M. Sleep and serotonin: an unfinished story. Neuropsychopharmacology. 1999;21:24S–27S. doi: 10.1016/S0893-133X(99)00009-3. [DOI] [PubMed] [Google Scholar]
- Kagan J. What is emotion? history, measures, and meanings. New Haven: Yale University Press; 2007. [Google Scholar]
- Kalin NH, Shelton SE, Davidson RJ. The role of the central nucleus of the amygdala in mediating fear and anxiety in the primate. J Neurosci. 2004;24:5506–5515. doi: 10.1523/JNEUROSCI.0292-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalin NH, Shelton SE, Davidson RJ, Kelley AE. The primate amygdala mediates acute fear but not the behavioral and physiological components of anxious temperament. J Neurosci. 2001;21:2067–2074. doi: 10.1523/JNEUROSCI.21-06-02067.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kandel ER. The molecular biology of memory storage: a dialog between genes and synapses. Biosci Rep. 2001;21:565–611. doi: 10.1023/a:1014775008533. [DOI] [PubMed] [Google Scholar]
- Kapp BS, Supple WF, Jr, Whalen PJ. Effects of electrical stimulation of the amygdaloid central nucleus on neocortical arousal in the rabbit. Behav Neurosci. 1994;108:81–93. doi: 10.1037//0735-7044.108.1.81. [DOI] [PubMed] [Google Scholar]
- Kapp BS, Whalen PJ, Supple WF, Pascoe JP. Amygdaloid contributions to conditioned arousal and sensory information processing. In: Aggleton JP, editor. The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction. New York: Wiley-Liss; 1992. pp. 229–254. [Google Scholar]
- Kelley AE. Ventral striatal control of appetitive motivation: role in ingestive behavior and reward-related learning. Neurosci Biobehav Rev. 2004;27:765–776. doi: 10.1016/j.neubiorev.2003.11.015. [DOI] [PubMed] [Google Scholar]
- Killcross AS, Robbins TW. Differential effects of intra-accumbens and systemic amphetamine on latent inhibition using an on-baseline, within-subject conditioned suppression paradigm. Psychopharmacology (Berl) 1993;110:479–489. doi: 10.1007/BF02244656. [DOI] [PubMed] [Google Scholar]
- Kitayama S, Markus HR, editors. Emotion and culture: Empirical studies of mutual influence. Washington, DC: American Psychological Association; 1994. [Google Scholar]
- Koob GF. Brain stress systems in the amygdala and addiction. Brain Res. 2009;1293:61–75. doi: 10.1016/j.brainres.2009.03.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaBar KS, Cabeza R. Cognitive neuroscience of emotional memory. Nat Rev Neurosci. 2006;7:54–64. doi: 10.1038/nrn1825. [DOI] [PubMed] [Google Scholar]
- Lakoff G. Women, Fire, and Dangerous Things: What Categories Reveal about the mind. Chicago: The University of Chicago Press; 1987. [Google Scholar]
- Lamprecht R, Dudai Y. The amygdala in conditioned taste aversion: it’s there, but where? In: Aggleton J, editor. The Amygdala. Oxford: Oxford U. Press; 2000. [Google Scholar]
- Lane RD, Chua PM, Dolan RJ. Common effects of emotional valence, arousal and attention on neural activation during visual processing of pictures. Neuropsychologia. 1999;37:989–997. doi: 10.1016/s0028-3932(99)00017-2. [DOI] [PubMed] [Google Scholar]
- Lang PJ. The varieties of emotional experience: a meditation on James-Lange theory. Psychol Rev. 1994;101:211–221. doi: 10.1037/0033-295x.101.2.211. [DOI] [PubMed] [Google Scholar]
- Lang PJ, Davis M. Emotion, motivation, and the brain: reflex foundations in animal and human research. Prog Brain Res. 2006;156:3–29. doi: 10.1016/S0079-6123(06)56001-7. [DOI] [PubMed] [Google Scholar]
- Lang PJ, Bradley MM, Cuthbert BN. Emotion, Attention, and the Startle Reflex. Psychological Review. 1990;97:377–395. [PubMed] [Google Scholar]
- Lange CG. The emotions. In: Dunlap E, editor. The Emotions. Baltimore: Williams and Wilkins; 1885/1922. [Google Scholar]
- Lashley KS. Experimental analysis of instinctive behavior. Psychol Rev. 1938;45:445–471. [Google Scholar]
- Lazaro-Munoz G, LeDoux JE, Cain CK. Sidman instrumental avoidance initially depends on lateral and Basal amygdala and is constrained by central amygdala-mediated Pavlovian processes. Biol Psychiatry. 2010;67:1120–1127. doi: 10.1016/j.biopsych.2009.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lazarus RS. A cognitivist’s reply to Zajonc on emotion and cognition. American Psychologist. 1981;36:222–223. [Google Scholar]
- Lazarus RS. Cognition and Motivation in Emotion. American Psychologist. 1991a;46:352–367. doi: 10.1037//0003-066x.46.4.352. [DOI] [PubMed] [Google Scholar]
- Lazarus RS. Emotion and Adaptation. New York: Oxford University Press; 1991b. [Google Scholar]
- Lazarus RS, Folkman S. Stress, Appraisal and Coping. New York: Springer; 1984. [Google Scholar]
- Lebestky T, Chang JS, Dankert H, Zelnik L, Kim YC, Han KA, Wolf FW, Perona P, Anderson DJ. Two different forms of arousal in Drosophila are oppositely regulated by the dopamine D1 receptor ortholog DopR via distinct neural circuits. Neuron. 2009;64:522–536. doi: 10.1016/j.neuron.2009.09.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LeDoux JE. Emotion and the Amygdala. In: Aggleton JP, editor. The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction. New York: Wiley-Liss, Inc; 1992. pp. 339–351. [Google Scholar]
- LeDoux JE. Seting “Stress” into Motion. In: Friedman MJ, Charney DS, Deutch AY, editors. Neurobiological and Clinical Consequences of Stress: from Normal Adaptation to PTSD. New York: Raven Press; 1995. pp. 125–134. [Google Scholar]
- LeDoux JE. The Emotional Brain. New York: Simon and Schuster; 1996. [Google Scholar]
- LeDoux JE. Emotion circuits in the brain. Annu Rev Neurosci. 2000;23:155–184. doi: 10.1146/annurev.neuro.23.1.155. [DOI] [PubMed] [Google Scholar]
- LeDoux JE. Synaptic Self: How our brains become who we are. New York: Viking; 2002. [Google Scholar]
- LeDoux JE. Emotional colouration of consciousness: how feelings come about. In: Weiskrantz L, Davies M, editors. Frontiers of Consciousness: Chichele Lectures. Oxford: Oxford University Press; 2008. pp. 69–130. [Google Scholar]
- LeDoux JE. Evolution of human emotion A view through fear. Prog Brain Res. 2012;195:431–442. doi: 10.1016/B978-0-444-53860-4.00021-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leventhal H, Scherer K. The relationship of emotion to cognition: a functional approach to a semantic controversy. Cognition and Emotion. 1987;1:3–28. [Google Scholar]
- Liddell BJ, Brown KJ, Kemp AH, Barton MJ, Das P, Peduto A, Gordon E, Williams LM. A direct brainstem-amygdala-cortical ‘alarm’ system for subliminal signals of fear. Neuroimage. 2005;24:235–243. doi: 10.1016/j.neuroimage.2004.08.016. [DOI] [PubMed] [Google Scholar]
- Lima SL, Dill LM. Behavioral decision made under the risk of predation: a review and prospectus. Can J Zool. 1990;68:619–640. [Google Scholar]
- Lin D, Boyle MP, Dollar P, Lee H, Lein ES, Perona P, Anderson DJ. Functional identification of an aggression locus in the mouse hypothalamus. Nature. 2011;470:221–226. doi: 10.1038/nature09736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindsley DB. Emotions. In: Stevens SS, editor. Handbook of Experimental Psychology. New York: Wiley; 1951. pp. 473–516. [Google Scholar]
- Litvin Y, Blanchard DC, Blanchard RJ. Rat 22kHz ultrasonic vocalizations as alarm cries. Behav Brain Res. 2007;182:166–172. doi: 10.1016/j.bbr.2006.11.038. [DOI] [PubMed] [Google Scholar]
- Loewy AD. Forebrain nuclei involved in autonomic control. Prog Brain Res. 1991;87:253–268. doi: 10.1016/s0079-6123(08)63055-1. [DOI] [PubMed] [Google Scholar]
- Lopez HS, Brown AM. Neuromodulation. Curr Opin Neurobiol. 1992;2:317–322. doi: 10.1016/0959-4388(92)90122-2. [DOI] [PubMed] [Google Scholar]
- Lorenz KZ. The Foundations of Ethology: The principal ideas and discoveries in animal behavior. New York: Simon and Schuster; 1981. [Google Scholar]
- Lucy JA. Linguistic Relativity. Annu Rev Anthropol. 1997;26:291–312. [Google Scholar]
- Luo Q, Holroyd T, Jones M, Hendler T, Blair J. Neural dynamics for facial threat processing as revealed by gamma band synchronization using MEG. Neuroimage. 2007;34:839–847. doi: 10.1016/j.neuroimage.2006.09.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyons W. Emotion. Cambridge: Cambridge University Press; 1980. [Google Scholar]
- MacLean PD. Psychosomatic disease and the “visceral brain”: recent developments bearing on the Papez theory of emotion. Psychosomatic Medicine. 1949;11:338–353. doi: 10.1097/00006842-194911000-00003. [DOI] [PubMed] [Google Scholar]
- MacLean PD. Some psychiatric implications of physiological studies on frontotemporal portion of limbic system (visceral brain) Electroencephalography and Clinical Neurophysiology. 1952;4:407–418. doi: 10.1016/0013-4694(52)90073-4. [DOI] [PubMed] [Google Scholar]
- Macnab RM, Koshland DE., Jr The gradient-sensing mechanism in bacterial chemotaxis. Proc Natl Acad Sci U S A. 1972;69:2509–2512. doi: 10.1073/pnas.69.9.2509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mandler G. Mind and Emotion. New York: Wiley; 1975. [Google Scholar]
- Manning A. An introduction to animal behaviour. Reading, MA: Addison-Wesley; 1967. [Google Scholar]
- Maren S. Neurobiology of Pavlovian fear conditioning. Annu Rev Neurosci. 2001;24:897–931. doi: 10.1146/annurev.neuro.24.1.897. [DOI] [PubMed] [Google Scholar]
- Maren S, Quirk GJ. Neuronal signalling of fear memory. Nat Rev Neurosci. 2004;5:844–852. doi: 10.1038/nrn1535. [DOI] [PubMed] [Google Scholar]
- Margulies C, Tully T, Dubnau J. Deconstructing memory in Drosophila. Curr Biol. 2005;15:R700–713. doi: 10.1016/j.cub.2005.08.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Markham CM, Blanchard DC, Canteras NS, Cuyno CD, Blanchard RJ. Modulation of predatory odor processing following lesions to the dorsal premammillary nucleus. Neurosci Lett. 2004;372:22–26. doi: 10.1016/j.neulet.2004.09.006. [DOI] [PubMed] [Google Scholar]
- Marks I. Fears, Phobias, and Rituals: Panic, Anxiety and Their Disorders. New York: Oxford University Press; 1987. [Google Scholar]
- Martinez-Garcia F, Martinez-Marcos A, Lanuza E. The pallial amygdala of amniote vertebrates: evolution of the concept, evolution of the structure. Brain Res Bull. 2002;57:463–469. doi: 10.1016/s0361-9230(01)00665-7. [DOI] [PubMed] [Google Scholar]
- Masson JM, McCarthy S. When elephants weep: The emotional lives of animals. New York: Delacorte; 1996. [Google Scholar]
- Maturana H, Varela F. The tree of knowledge: The biological roots of human understanding. Boston: New Science Library; 1987. [Google Scholar]
- McClelland D, Atkinson J, Clark R, Lowell E. The achievement motive. New York: Irvington Publishers; 1953. [Google Scholar]
- McEwen BS. The brain is the central organ of stress and adaptation. Neuroimage. 2009;47:911–913. doi: 10.1016/j.neuroimage.2009.05.071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McEwen BS, Eiland L, Hunter RG, Miller MM. Stress and anxiety: structural plasticity and epigenetic regulation as a consequence of stress. Neuropharmacology. 2012;62:3–12. doi: 10.1016/j.neuropharm.2011.07.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGaugh JL. Memory--a century of consolidation. Science. 2000;287:248–251. doi: 10.1126/science.287.5451.248. [DOI] [PubMed] [Google Scholar]
- McGaugh JL. Memory and Emotion: The making of lasting memories. London: The Orion Publishing Group; 2003. [Google Scholar]
- McGrath PT, Rockman MV, Zimmer M, Jang H, Macosko EZ, Kruglyak L, Bargmann CI. Quantitative mapping of a digenic behavioral trait implicates globin variation in C. elegans sensory behaviors. Neuron. 2009;61:692–699. doi: 10.1016/j.neuron.2009.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McNaughton N. Biology and emotion. Cambridge: Cambridge University Press; 1989. [Google Scholar]
- Miller N. Experimental studies of conflict. In: Endler NS, Hunt JM, editors. Personality and the Behavioral Disorders. New York: Ronald Press; 1944. pp. 431–465. [Google Scholar]
- Mineka S, Ohman A. Phobias and preparedness: the selective, automatic, and encapsulated nature of fear. Biol Psychiatry. 2002;52:927–937. doi: 10.1016/s0006-3223(02)01669-4. [DOI] [PubMed] [Google Scholar]
- Mobbs D, Marchant JL, Hassabis D, Seymour B, Tan G, Gray M, Petrovic P, Dolan RJ, Frith CD. From threat to fear: the neural organization of defensive fear systems in humans. J Neurosci. 2009;29:12236–12243. doi: 10.1523/JNEUROSCI.2378-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mogenson GJ, Jones DL, Yim CY. From motivation to action: functional interface between the limbic system and the motor system. Prog Neurobiol. 1980;14:69–97. doi: 10.1016/0301-0082(80)90018-0. [DOI] [PubMed] [Google Scholar]
- Monsey MS, Ota KT, Akingbade IF, Hong ES, Schafe GE. Epigenetic alterations are critical for fear memory consolidation and synaptic plasticity in the lateral amygdala. PLoS One. 2011;6:e19958. doi: 10.1371/journal.pone.0019958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monti JM, Jantos H. The roles of dopamine and serotonin, and of their receptors, in regulating sleep and waking. Prog Brain Res. 2008;172:625–646. doi: 10.1016/S0079-6123(08)00929-1. [DOI] [PubMed] [Google Scholar]
- Morgan CT. Physiological Psychology. New York: McGraw-Hill; 1943. [Google Scholar]
- Morris JS, Ohman A, Dolan RJ. A subcortical pathway to the right amygdala mediating “unseen” fear. Proceedings of the National Academy of Sciences, USA. 1999;96:1680–1685. doi: 10.1073/pnas.96.4.1680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrison SE, Salzman CD. Re-valuing the amygdala. Curr Opin Neurobiol. 2010;20:221–230. doi: 10.1016/j.conb.2010.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morton GJ, Cummings DE, Baskin DG, Barsh GS, Schwartz MW. Central nervous system control of food intake and body weight. Nature. 2006;443:289–295. doi: 10.1038/nature05026. [DOI] [PubMed] [Google Scholar]
- Moruzzi G, Magoun HW. Brain Stem Reticular Formation and Activation of the EEG. Electroencephalography and Clinical Neurophysiology. 1949;1:455–473. [PubMed] [Google Scholar]
- Moscarello JM, Ben-Shahar O, Ettenberg A. Effects of food deprivation on goal-directed behavior, spontaneous locomotion, and c-Fos immunoreactivity in the amygdala. Behav Brain Res. 2009;197:9–15. doi: 10.1016/j.bbr.2008.07.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motta SC, Goto M, Gouveia FV, Baldo MV, Canteras NS, Swanson LW. Dissecting the brain’s fear system reveals the hypothalamus is critical for responding in subordinate conspecific intruders. Proc Natl Acad Sci U S A. 2009;106:4870–4875. doi: 10.1073/pnas.0900939106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mowrer OH. Learning theory and behavior. New York: Wiley; 1960. [Google Scholar]
- Moyer KE. The psychobiology of aggression. New York: Harper & Row; 1976. [Google Scholar]
- Mozzachiodi R, Byrne JH. More than synaptic plasticity: role of nonsynaptic plasticity in learning and memory. Trends Neurosci. 2010;33:17–26. doi: 10.1016/j.tins.2009.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagel T. What is it like to be a bat? Philosophical Review. 1974;83:4435–4450. [Google Scholar]
- Nakamura K, Morrison SF. Central efferent pathways mediating skin cooling-evoked sympathetic thermogenesis in brown adipose tissue. Am J Physiol Regul Integr Comp Physiol. 2007;292:R127–136. doi: 10.1152/ajpregu.00427.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nauta WJH, Karten HJ. A general profile of the vertebrate brain, with sidelights on the ancestry of cerebral cortex. In: Schmitt FO, editor. The Neurosciences: Second Study Program. New York: The Rockefeller University Press; 1970. pp. 7–26. [Google Scholar]
- Nesse R. Evolutionary explanations of emotions. Human Nature. 1990;1:261–289. doi: 10.1007/BF02733986. [DOI] [PubMed] [Google Scholar]
- Nestler EJ. Molecular mechanisms of drug addiction. Neuropharmacology. 2004;47(Suppl 1):24–32. doi: 10.1016/j.neuropharm.2004.06.031. [DOI] [PubMed] [Google Scholar]
- Nieuwenhuys R. Chemoarchitecture of the Brain. Berlin: Springer-Verlag; 1985. [Google Scholar]
- Nishino S. Hypothalamus, hypocretins/orexin, and vigilance control. Handb Clin Neurol. 2011;99:765–782. doi: 10.1016/B978-0-444-52007-4.00006-0. [DOI] [PubMed] [Google Scholar]
- Northcutt RG, Kaas JH. The emergence and evolution of mammalian neocortex. Trends in Neurosciences. 1995;18:373–379. doi: 10.1016/0166-2236(95)93932-n. [DOI] [PubMed] [Google Scholar]
- Ochsner KN, Gross JJ. The cognitive control of emotion. Trends Cogn Sci. 2005;9:242–249. doi: 10.1016/j.tics.2005.03.010. [DOI] [PubMed] [Google Scholar]
- Ochsner KN, Bunge SA, Gross JJ, Gabrieli JD. Rethinking feelings: an FMRI study of the cognitive regulation of emotion. J Cogn Neurosci. 2002;14:1215–1229. doi: 10.1162/089892902760807212. [DOI] [PubMed] [Google Scholar]
- Ohman A. Of snakes and faces: an evolutionary perspective on the psychology of fear. Scand J Psychol. 2009;50:543–552. doi: 10.1111/j.1467-9450.2009.00784.x. [DOI] [PubMed] [Google Scholar]
- Öhman A. Face the beast and fear the face: animal and social fears as prototypes for evolutionary analyses of emotion. Psychophysiology. 1986;23:123–145. doi: 10.1111/j.1469-8986.1986.tb00608.x. [DOI] [PubMed] [Google Scholar]
- Olds J. Drives and Reinforcement. New York: Raven; 1977. [Google Scholar]
- Olds J, Milner P. Positive reinforcement produced by electrical stimulation of septal and other regions of the brain. J Comp Physiol Psychol. 1954;47:419–427. doi: 10.1037/h0058775. [DOI] [PubMed] [Google Scholar]
- Ono T, Nishijo H. Neurophysiological Basis of the Kluver-Bucy Syndrome: Responses of Monkey Amygdaloid Neurons to Biologically Significant Objects. In: Aggleton JP, editor. The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction. New York: Wiley-Liss, Inc; 1992. pp. 167–190. [Google Scholar]
- Oomura Y, Aou S, Koyama Y, Fujita I, Yoshimatsu H. Central control of sexual behavior. Brain Res Bull. 1988;20:863–870. doi: 10.1016/0361-9230(88)90103-7. [DOI] [PubMed] [Google Scholar]
- Ortony A, Turner TJ. What’s basic about basic emotions? Psychological Review. 1990;97:315–331. doi: 10.1037/0033-295x.97.3.315. [DOI] [PubMed] [Google Scholar]
- Packard MG, Knowlton BJ. Learning and memory functions of the Basal Ganglia. Annu Rev Neurosci. 2002;25:563–593. doi: 10.1146/annurev.neuro.25.112701.142937. [DOI] [PubMed] [Google Scholar]
- Pagani JH, Rosen JB. The medial hypothalamic defensive circuit and 2,5-dihydro-2,4,5-trimethylthiazoline (TMT) induced fear: comparison of electrolytic and neurotoxic lesions. Brain Res. 2009;1286:133–146. doi: 10.1016/j.brainres.2009.06.062. [DOI] [PubMed] [Google Scholar]
- Panksepp J. Toward a general psychobiological theory of emotions. Behavioral and Brain Sciences. 1982;5:407–467. [Google Scholar]
- Panksepp J. Affective Neuroscience. New York: Oxford U. Press; 1998. [Google Scholar]
- Panksepp J. Emotions as natural kinds within the mammalian brain. In: Lewis M, Haviland-Jones JM, editors. Handbook of Emotions. 2. New York: The Guilford Press; 2000. pp. 137–156. [Google Scholar]
- Panksepp J. Affective consciousness: Core emotional feelings in animals and humans. Conscious Cogn. 2005;14:30–80. doi: 10.1016/j.concog.2004.10.004. [DOI] [PubMed] [Google Scholar]
- Pare D, Quirk GJ, LeDoux JE. New vistas on amygdala networks in conditioned fear. J Neurophysiol. 2004;92:1–9. doi: 10.1152/jn.00153.2004. [DOI] [PubMed] [Google Scholar]
- Parkinson JA, Olmstead MC, Burns LH, Robbins TW, Everitt BJ. Dissociation in effects of lesions of the nucleus accumbens core and shell on appetitive pavlovian approach behavior and the potentiation of conditioned reinforcement and locomotor activity by D-amphetamine. J Neurosci. 1999;19:2401–2411. doi: 10.1523/JNEUROSCI.19-06-02401.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paton JJ, Belova MA, Morrison SE, Salzman CD. The primate amygdala represents the positive and negative value of visual stimuli during learning. Nature. 2006;439:865–870. doi: 10.1038/nature04490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pavesi E, Canteras NS, Carobrez AP. Acquisition of Pavlovian fear conditioning using beta-adrenoceptor activation of the dorsal premammillary nucleus as an unconditioned stimulus to mimic live predator-threat exposure. Neuropsychopharmacology. 2011;36:926–939. doi: 10.1038/npp.2010.231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrovich GD. Learning and the motivation to eat: forebrain circuitry. Physiol Behav. 2011;104:582–589. doi: 10.1016/j.physbeh.2011.04.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrovich GD, Risold PY, Swanson LW. Organization of projections from the basomedial nucleus of the amygdala: a PHAL study in the rat. J Comp Neurol. 1996;374:387–420. doi: 10.1002/(SICI)1096-9861(19961021)374:3<387::AID-CNE6>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- Petrovich GD, Canteras NS, Swanson LW. Combinatorial amygdalar inputs to hippocampal domains and hypothalamic behavior systems. Brain Res Brain Res Rev. 2001;38:247–289. doi: 10.1016/s0165-0173(01)00080-7. [DOI] [PubMed] [Google Scholar]
- Petrovich GD, Setlow B, Holland PC, Gallagher M. Amygdalo-hypothalamic circuit allows learned cues to override satiety and promote eating. J Neurosci. 2002;22:8748–8753. doi: 10.1523/JNEUROSCI.22-19-08748.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfaff D, Ribeiro A, Matthews J, Kow LM. Concepts and mechanisms of generalized central nervous system arousal. Ann N Y Acad Sci. 2008;1129:11–25. doi: 10.1196/annals.1417.019. [DOI] [PubMed] [Google Scholar]
- Pfaff DW. Drive: neurobiological and molecular mechanisms of sexual motivation. Cambridge, MA: MIT Press; 1999. [Google Scholar]
- Phelps EA. Emotion and cognition: insights from studies of the human amygdala. Annu Rev Psychol. 2006;57:27–53. doi: 10.1146/annurev.psych.56.091103.070234. [DOI] [PubMed] [Google Scholar]
- Phelps EA, LeDoux JE. Contributions of the amygdala to emotion processing: from animal models to human behavior. Neuron. 2005;48:175–187. doi: 10.1016/j.neuron.2005.09.025. [DOI] [PubMed] [Google Scholar]
- Pinker S. How the Mind Works. New York: Norton; 1997. [DOI] [PubMed] [Google Scholar]
- Pirri JK, Alkema MJ. The neuroethology of C. elegans escape. Curr Opin Neurobiol. 2012 doi: 10.1016/j.conb.2011.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitkänen A, Savander V, LeDoux JE. Organization of intra-amygdaloid circuitries in the rat: an emerging framework for understanding functions of the amygdala. Trends Neurosci. 1997;20:517–523. doi: 10.1016/s0166-2236(97)01125-9. [DOI] [PubMed] [Google Scholar]
- Pollatos O, Gramann K, Schandry R. Neural systems connecting interoceptive awareness and feelings. Hum Brain Mapp. 2007;28:9–18. doi: 10.1002/hbm.20258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preuss TM. Do rats have prefrontal cortex? The Rose-Woolsey-Akert program reconsidered. Journal of Cognitive Neuroscience. 1995;7:1–24. doi: 10.1162/jocn.1995.7.1.1. [DOI] [PubMed] [Google Scholar]
- Prinz J. Which emotions are basic? In: Cruise P, Evans D, editors. Emotion, Evolution and Rationality. Oxford: Oxford University Press; 2004. pp. 69–87. [Google Scholar]
- Quirk GJ, Mueller D. Neural mechanisms of extinction learning and retrieval. Neuropsychopharmacology. 2008;33:56–72. doi: 10.1038/sj.npp.1301555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reep R. Relationship between prefrontal and limbic cortex: a comparative anatomical review. Brain, Behavior and Evolution. 1984;25:5–80. doi: 10.1159/000118849. [DOI] [PubMed] [Google Scholar]
- Reis DJ, LeDoux JE. Some central neural mechanisms governing resting and behaviorally coupled control of blood pressure. Circulation. 1987;76:2–19. [PubMed] [Google Scholar]
- Reisenzein R. On Oatley and Johnson-Laird’s Theory of emotion and hierarchical structures in the affective lexicon. Cognition and Emotion. 1995;9:383–416. [Google Scholar]
- Rescorla RA, Wagner AR. A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. In: Black AA, Prokasy WF, editors. Classical Conditioning II: Current Research and Theory. New York: Appleton-Centry-Crofts; 1972. pp. 64–99. [Google Scholar]
- Reyes BA, Carvalho AF, Vakharia K, Van Bockstaele EJ. Amygdalar peptidergic circuits regulating noradrenergic locus coeruleus neurons: linking limbic and arousal centers. Exp Neurol. 2011;230:96–105. doi: 10.1016/j.expneurol.2011.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reynolds SM, Berridge KC. Emotional environments retune the valence of appetitive versus fearful functions in nucleus accumbens. Nat Neurosci. 2008;11:423–425. doi: 10.1038/nn2061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robbins TW. Arousal systems and attentional processes. Biol Psychol. 1997;45:57–71. doi: 10.1016/s0301-0511(96)05222-2. [DOI] [PubMed] [Google Scholar]
- Robbins TW. Chemistry of the mind: neurochemical modulation of prefrontal cortical function. J Comp Neurol. 2005;493:140–146. doi: 10.1002/cne.20717. [DOI] [PubMed] [Google Scholar]
- Robbins TW, Everitt BJ. A role for mesencephalic dopamine in activation: commentary on Berridge (2006) Psychopharmacology (Berl) 2007;191:433–437. doi: 10.1007/s00213-006-0528-7. [DOI] [PubMed] [Google Scholar]
- Rodrigues SM, LeDoux JE, Sapolsky RM. The influence of stress hormones on fear circuitry. Annu Rev Neurosci. 2009;32:289–313. doi: 10.1146/annurev.neuro.051508.135620. [DOI] [PubMed] [Google Scholar]
- Roitman MF, Wheeler RA, Carelli RM. Nucleus accumbens neurons are innately tuned for rewarding and aversive taste stimuli, encode their predictors, and are linked to motor output. Neuron. 2005;45:587–597. doi: 10.1016/j.neuron.2004.12.055. [DOI] [PubMed] [Google Scholar]
- Roitman MF, Wheeler RA, Wightman RM, Carelli RM. Real-time chemical responses in the nucleus accumbens differentiate rewarding and aversive stimuli. Nat Neurosci. 2008;11:1376–1377. doi: 10.1038/nn.2219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rolls ET. Neurophysiology and Functions of the Primate Amygdala. In: Aggleton JP, editor. The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction. New York: Wiley-Liss, Inc; 1992. pp. 143–165. [Google Scholar]
- Rolls ET. The Brain and Emotion. Oxford: Oxford University Press; 1999. [Google Scholar]
- Rolls ET. Emotion Explained. New York: Oxford University Press; 2005. [Google Scholar]
- Roozendaal B, McEwen BS, Chattarji S. Stress, memory and the amygdala. Nat Rev Neurosci. 2009;10:423–433. doi: 10.1038/nrn2651. [DOI] [PubMed] [Google Scholar]
- Rorty AO. Explaining emotions. In: Rorty AO, editor. Explaining emotions. Berkeley: University of California Press; 1980. [Google Scholar]
- Rosenkranz JA, Grace AA. Dopamine-mediated modulation of odour-evoked amygdala potentials during pavlovian conditioning. Nature. 2002;417:282–287. doi: 10.1038/417282a. [DOI] [PubMed] [Google Scholar]
- Rudrauf D, Lachaux JP, Damasio A, Baillet S, Hugueville L, Martinerie J, Damasio H, Renault B. Enter feelings: somatosensory responses following early stages of visual induction of emotion. Int J Psychophysiol. 2009;72:13–23. doi: 10.1016/j.ijpsycho.2008.03.015. [DOI] [PubMed] [Google Scholar]
- Russell JA. A Circumplex Model of Affect. J Pers Soc Psychol. 1980;39:1161–1178. [Google Scholar]
- Russell JA. Natural language concepts of emotion. In: Hogan R, Jones WH, Stewart AJ, Healy JM, Ozer DJ, editors. Perspectives in Personality. London: Jessica Kingsley; 1991. [Google Scholar]
- Russell JA. Core affect and the psychological construction of emotion. Psychol Rev. 2003;110:145–172. doi: 10.1037/0033-295x.110.1.145. [DOI] [PubMed] [Google Scholar]
- Russell JA, Barrett LF. Core affect, prototypical emotional episodes, and other things called emotion: dissecting the elephant. J Pers Soc Psychol. 1999;76:805–819. doi: 10.1037//0022-3514.76.5.805. [DOI] [PubMed] [Google Scholar]
- Salamone JD. The involvement of nucleus accumbens dopamine in appetitive and aversive motivation. Behav Brain Res. 1994;61:117–133. doi: 10.1016/0166-4328(94)90153-8. [DOI] [PubMed] [Google Scholar]
- Sander D, Grandjean D, Scherer KR. A systems approach to appraisal mechanisms in emotion. Neural Netw. 2005;18:317–352. doi: 10.1016/j.neunet.2005.03.001. [DOI] [PubMed] [Google Scholar]
- Saper CB. Staying awake for dinner: hypothalamic integration of sleep, feeding, and circadian rhythms. Prog Brain Res. 2006;153:243–252. doi: 10.1016/S0079-6123(06)53014-6. [DOI] [PubMed] [Google Scholar]
- Saper CB, Chou TC, Elmquist JK. The need to feed: homeostatic and hedonic control of eating. Neuron. 2002;36:199–211. doi: 10.1016/s0896-6273(02)00969-8. [DOI] [PubMed] [Google Scholar]
- Sapolsky RM, Krey LC, McEwen BS. The neuroendocrinology of stress and aging: The glucocorticoid cascade hypothesis. Endocrine Reviews. 1986;7:284–301. doi: 10.1210/edrv-7-3-284. [DOI] [PubMed] [Google Scholar]
- Sarter M, Hasselmo ME, Bruno JP, Givens B. Unraveling the attentional functions of cortical cholinergic inputs: interactions between signal-driven and cognitive modulation of signal detection. Brain Res Brain Res Rev. 2005;48:98–111. doi: 10.1016/j.brainresrev.2004.08.006. [DOI] [PubMed] [Google Scholar]
- Scarantino A. Core affect and natural affective kinds. Philosophy of Science. 2009;76:940–957. [Google Scholar]
- Schachter S. Cognition and centralist-peripheralist controversies in motivation and emotion. In: Gazzaniga MS, Blakemore CB, editors. Handbook of Psychobiology. New York: Academic Press; 1975. pp. 529–564. [Google Scholar]
- Schachter S, Singer JE. Cognitive, social, and physiological determinants of emotional state. Psychol Rev. 1962;69:379–399. doi: 10.1037/h0046234. [DOI] [PubMed] [Google Scholar]
- Scherer K. Emotions as episodes of subsystem synchronization driven by nonlinear appraisal processes. In: Lewis M, Granic I, editors. Emotion, development, and self-organization: Dynamic systems approaches to emotional development. New York: Cambridge University Press; 2000. pp. 70–99. [Google Scholar]
- Scherer KR. Emotion as a multicomponent process: A model and some cross-cultural data. Review of Personality and Social Psychology. 1984;5:37–63. [Google Scholar]
- Scherer KR. Criteria for emotion-antecedent appraisal: A review. In: Hamilton V, Bower GH, Frijda NH, editors. Cognitive Perspectives on Emotion and Motivation. Norwell, MA: Kluwer Academic Publishers; 1988. pp. 89–126. [Google Scholar]
- Scherer KR, Schorr A, Johnstone T, editors. Appraisal processes in emotion: Theory, Methods, Research. London: London University Press; 2001. [Google Scholar]
- Schildkraut JJ, Kety SS. Biogenic amines and emotion. Science. 1967;156:21–37. doi: 10.1126/science.156.3771.21. [DOI] [PubMed] [Google Scholar]
- Schiller D, Delgado MR. Overlapping neural systems mediating extinction, reversal and regulation of fear. Trends Cogn Sci. 2010;14:268–276. doi: 10.1016/j.tics.2010.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schimitel FG, de Almeida GM, Pitol DN, Armini RS, Tufik S, Schenberg LC. Evidence of a suffocation alarm system within the periaqueductal gray matter of the rat. Neuroscience. 2011 doi: 10.1016/j.neuroscience.2011.10.032. [DOI] [PubMed] [Google Scholar]
- Schneirla TC. An evolutionary theory of biphasic processes underlying approach and withdrawal. In: Jones MR, editor. Nebraska Symposium on Motivation. University of Nebraska Press; 1959. pp. 1–42. [Google Scholar]
- Schober J, Weil Z, Pfaff D. How generalized CNS arousal strengthens sexual arousal (and vice versa) Horm Behav. 2011;59:689–695. doi: 10.1016/j.yhbeh.2010.10.005. [DOI] [PubMed] [Google Scholar]
- Schoenbaum G, Setlow B. Lesions of nucleus accumbens disrupt learning about aversive outcomes. J Neurosci. 2003;23:9833–9841. doi: 10.1523/JNEUROSCI.23-30-09833.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schulkin J. Rethinking homeostasis: Allostatic regulation in physiology and pathophysiology. Cambridge: MIT Press; 2003. [Google Scholar]
- Schultz W. Dopamine neurons and their role in reward mechanisms. Curr Opin Neurobiol. 1997;7:191–197. doi: 10.1016/s0959-4388(97)80007-4. [DOI] [PubMed] [Google Scholar]
- Schultz W. Behavioral theories and the neurophysiology of reward. Annu Rev Psychol. 2006;57:87–115. doi: 10.1146/annurev.psych.56.091103.070229. [DOI] [PubMed] [Google Scholar]
- Sears RM, Liu RJ, Narayanan NS, Sharf R, Yeckel MF, Laubach M, Aghajanian GK, DiLeone RJ. Regulation of nucleus accumbens activity by the hypothalamic neuropeptide melanin-concentrating hormone. J Neurosci. 2010;30:8263–8273. doi: 10.1523/JNEUROSCI.5858-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seligman MEP. Phobias and Preparedness. Behavior therapy. 1971;2:307–320. [Google Scholar]
- Selye H. Stress and disease. Science. 1955;122:625–631. doi: 10.1126/science.122.3171.625. [DOI] [PubMed] [Google Scholar]
- Semendeferi K, Teffer K, Buxhoeveden DP, Park MS, Bludau S, Amunts K, Travis K, Buckwalter J. Spatial organization of neurons in the frontal pole sets humans apart from great apes. Cereb Cortex. 2011;21:1485–1497. doi: 10.1093/cercor/bhq191. [DOI] [PubMed] [Google Scholar]
- Sesack SR, Grace AA. Cortico-Basal Ganglia reward network: microcircuitry. Neuropsychopharmacology. 2010;35:27–47. doi: 10.1038/npp.2009.93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seymour B, Dolan R. Emotion, decision making, and the amygdala. Neuron. 2008;58:662–671. doi: 10.1016/j.neuron.2008.05.020. [DOI] [PubMed] [Google Scholar]
- Shallice T. Information processing models of consiousness. In: Marcel A, Bisiach E, editors. Consciousness in contemporary science. Oxford: Oxford U. Press; 1988. pp. 305–333. [Google Scholar]
- Shallice T, Stuss DT, Alexander MP, Picton TW, Derkzen D. The multiple dimensions of sustained attention. Cortex. 2008;44:794–805. doi: 10.1016/j.cortex.2007.04.002. [DOI] [PubMed] [Google Scholar]
- Sheffield FD, Roby TB. Reward value of a non-nutritive sweet taste. Journal of Comparative Physiology and Psychology. 1950;43:471–481. doi: 10.1037/h0061365. [DOI] [PubMed] [Google Scholar]
- Shepherd G. Neurobiology. 2. New York: Oxford; 1988. [Google Scholar]
- Sherrington CS. The Integrative Action of the Nervous System. New Haven: Yale U. Press; 1906. [Google Scholar]
- Siegel A, Edinger H. Neural control of aggression and rage behavior. In: Morgane PJ, Panksepp J, editors. Handbook of the Hypothalamus, Vol. 3, Behavioral Studies of the Hypothalamus. New York: Marcel Dekker; 1981. pp. 203–240. [Google Scholar]
- Skinner BF. The behavior of organisms: An experimental analysis. New York: Appleton-Century-Crofts; 1938. [Google Scholar]
- Smith CA, Ellsworth PC. Patterns of cognitive appraisal in emotion. Journal of Personality and Social Psychology. 1985;56:339–353. [PubMed] [Google Scholar]
- Sosulski DL, Bloom ML, Cutforth T, Axel R, Datta SR. Distinct representations of olfactory information in different cortical centres. Nature. 2011;472:213–216. doi: 10.1038/nature09868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stellar E. The physiology of motivation. Psychol Rev. 1954;61:5–22. doi: 10.1037/h0060347. [DOI] [PubMed] [Google Scholar]
- Steriade M. Brain activation, then (1949) and now: coherent fast rhythms in corticothalamic networks. Arch Ital Biol. 1995;134:5–20. [PubMed] [Google Scholar]
- Steriade M. Acetylcholine systems and rhythmic activities during the waking--sleep cycle. Prog Brain Res. 2004;145:179–196. doi: 10.1016/S0079-6123(03)45013-9. [DOI] [PubMed] [Google Scholar]
- Striedter GF. Principles of Brain Evolution. Sunderland: Sinauer Associates; 2005. [Google Scholar]
- Sutton RS, Barto AG. A temporal-difference model of classical conditioning. Proceedings of the Ninth Annual Conference of the Cognitive Science Society; 1987. pp. 355–378. [Google Scholar]
- Swanson LW. Cerebral hemisphere regulation of motivated behavior. Brain Res. 2000;886:113–164. doi: 10.1016/s0006-8993(00)02905-x. [DOI] [PubMed] [Google Scholar]
- Swanson LW. Brain Architecture: Understanding the Basic Plan. Oxford: Oxford University Press; 2002. [Google Scholar]
- Talarovicova A, Krskova L, Kiss A. Some assessments of the amygdala role in suprahypothalamic neuroendocrine regulation: a minireview. Endocr Regul. 2007;41:155–162. [PubMed] [Google Scholar]
- Tamietto M, Castelli L, Vighetti S, Perozzo P, Geminiani G, Weiskrantz L, de Gelder B. Unseen facial and bodily expressions trigger fast emotional reactions. Proc Natl Acad Sci U S A. 2009;106:17661–17666. doi: 10.1073/pnas.0908994106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson R, LeDoux JE. Common brain regions essential for the expression of learned and instinctive visual habits in the albino rat. Bull Psychon Soc. 1974;4:78–80. [Google Scholar]
- Tinbergen N. The Study of Instinct. New York: Oxford University Press; 1951. [Google Scholar]
- Tolman EC. Purposive behavior. New York: Appleton-Century-Crofts; 1932. [Google Scholar]
- Tomkins SS. Affect, Imagery, Consciousness. New York: Springer; 1962. [Google Scholar]
- Tooby J, Cosmides L. The past explains the present: Emotional adaptations and the structure of ancestral environments. Ethological Sociobiology. 1990;11:375–424. [Google Scholar]
- Trowill JA, Panksepp J, Gandelman R. An incentive model of rewarding brain stimulation. Psychol Rev. 1969;76:264–281. doi: 10.1037/h0027295. [DOI] [PubMed] [Google Scholar]
- von Holst E, von Saint-Paul U. Electrically controlled behavior. Sci Am. 1962;206:50–59. doi: 10.1038/scientificamerican0362-50. [DOI] [PubMed] [Google Scholar]
- Vyas A, Kim SK, Giacomini N, Boothroyd JC, Sapolsky RM. Behavioral changes induced by Toxoplasma infection of rodents are highly specific to aversion of cat odors. Proc Natl Acad Sci U S A. 2007;104:6442–6447. doi: 10.1073/pnas.0608310104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker DL, Davis M. Light-enhanced startle: further pharmacological and behavioral characterization. Psychopharmacology (Berl) 2002;159:304–310. doi: 10.1007/s002130100913. [DOI] [PubMed] [Google Scholar]
- Wallace KJ, Rosen JB. Predator odor as an unconditioned fear stimulus in rats: elicitation of freezing by trimethylthiazoline, a component of fox feces. Behav Neurosci. 2000;114:912–922. doi: 10.1037//0735-7044.114.5.912. [DOI] [PubMed] [Google Scholar]
- Wang L, Han X, Mehren J, Hiroi M, Billeter JC, Miyamoto T, Amrein H, Levine JD, Anderson DJ. Hierarchical chemosensory regulation of male-male social interactions in Drosophila. Nat Neurosci. 2011;14:757–762. doi: 10.1038/nn.2800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weinberger NM. Retuning the brain by fear conditioning. In: Gazzaniga MS, editor. The Cognitive Neurosciences. Cambridge, MA: The MIT Press; 1995. pp. 1071–1090. [Google Scholar]
- Whalen PJ. Fear, vigilance and ambiguity: initial neuroimaging studies of the human amygdala [manuscript] 1998 [Google Scholar]
- Whalen PJ, Phelps EA. The human amygdala. New York: Guilford Press; 2009. [Google Scholar]
- Whalen PJ, Kagan J, Cook RG, Davis FC, Kim H, Polis S, McLaren DG, Somerville LH, McLean AA, Maxwell JS, Johnstone T. Human amygdala responsivity to masked fearful eye whites. Science. 2004;306:2061. doi: 10.1126/science.1103617. [DOI] [PubMed] [Google Scholar]
- Whorf BL. Language, thought, and reality. Cambridge: Technology Press of MIT; 1956. [Google Scholar]
- Wickens JR, Horvitz JC, Costa RM, Killcross S. Dopaminergic mechanisms in actions and habits. J Neurosci. 2007;27:8181–8183. doi: 10.1523/JNEUROSCI.1671-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wierzbicka A. Emotion, language, and cultural scripts. In: Kitayama S, Markus HR, editors. Emotion and Culture: Empirical studies of mutual influence. Washington, DC: American Psychological Association; 1994. pp. 133–196. [Google Scholar]
- Wise RA. Role of brain dopamine in food reward and reinforcement. Philos Trans R Soc Lond B Biol Sci. 2006;361:1149–1158. doi: 10.1098/rstb.2006.1854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wise SP. Forward frontal fields: phylogeny and fundamental function. Trends Neurosci. 2008;31:599–608. doi: 10.1016/j.tins.2008.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin JC, Tully T. CREB and the formation of long-term memory. Curr Opin Neurobiol. 1996;6:264–268. doi: 10.1016/s0959-4388(96)80082-1. [DOI] [PubMed] [Google Scholar]
- Yovell Y, Kandel ER, Dudai Y, Abrams TW. A quantitative study of the Ca2+/calmodulin sensitivity of adenylyl cyclase in Aplysia, Drosophila, and rat. J Neurochem. 1992;59:1736–1744. doi: 10.1111/j.1471-4159.1992.tb11005.x. [DOI] [PubMed] [Google Scholar]
- Zachar P. The classification of emotion and scientific realism. J Theor Philosoph Psychol. 2006;26:120–138. [Google Scholar]
- Zajonc R. Feeling and thinking: preferences need no inferences. American Psychologist. 1980;35:151–175. [Google Scholar]
- Zhang TY, Parent C, Weaver I, Meaney MJ. Maternal programming of individual differences in defensive responses in the rat. Ann N Y Acad Sci. 2004;1032:85–103. doi: 10.1196/annals.1314.007. [DOI] [PubMed] [Google Scholar]