

Abstract
Parmotrema A. Massal. is a common lichen genus scattered throughout the Korean Peninsula; however, no detailed taxonomic or revisionary study of this genus has been conducted for nearly two decades. Therefore, this study revised the taxonomy of this genus based on specimens deposited in the lichen herbarium at the Korean Lichen Research Institute and samples wereidentified using recent literature. In this revisionary study, a total of eighteen species of Parmotrema including eight new records [Parmotrema cetratum (Ach.) Hale, Parmotrema cristiferum (Taylor) Hale, Parmotrema grayanum (Hue) Hale, Parmotrema defectum (Hale) Hale, Parmotrema dilatatum (Vain.) Hale, Parmotrema margaritatum (Hue) Hale, Parmotrema pseudocrinitum (Abbayes) Hale, and Parmotrema subsumptum (Nyl.) Hale] are documented. Detailed descriptions of each species with their morphological, anatomical and chemical characteristics are also given and a key to the known Parmotrema species of the Korean Peninsula is presented.
Keywords: Key, New record, Parmeliaceae, Parmotrema, South Korea
Parmotrema A. Massal. (previously known as Parmelia s. lat.) is one of the largest genera of parmelioid core in the family Parmeliaceae [1]. According to Bayesian analysis of nuclear internal transcribed spacer (ITS), large-subunit rDNA and mitochondrial small subunit (SSU) rDNA sequences carried out by Blanco et al. [1], the parmotremoid genera Rimelia, Canomaculina, and Concamerella are synonymized with Parmotrema. These findings were further confirmed by morphological analysis of the same groups by Louwhoff and Crisp [2]. Molecular phylogenetic analyses carried out by Divakar et al. [3] revealed that P. reticulatum and P. clavuliferum formed a monophyletic clade based on nuclear ITS rDNA and mitochondrial SSU rDNA sequences. Thus, P. reticulatum and P. clavuliferum were combined under the older name, P. reticulatum.
The genus Parmotrema is usually characterized by large foliose thalli with broad lobes, commonly with a broad erhizinate marginal zone on the lower surface, pored epicortex, thick-walled hyaline ellipsoid ascospores, sublageniform or filiform conidia and with or without marginal cilia. The greatest distribution of the genus is in tropical regions, where more than 220 species are known [4]. Currently, this genus is composed of c. 350 species worldwide [1, 5, 6]. To date, only 12 species have been reported from South Korea: Parmotrema arnoldii (Du Rietz) Hale 1974, Parmotrema austrosinense (Zahlbr.) Hale 1974, Parmotrema clavuliferum (Räsänen) Streimann 1986 [previously known as Rimelia clavulifera (Räsänen) Kurok. 1991], Parmotrema crinitum (Ach.) M. Choisy 1974, Parmotrema eciliatum (Nyl.) Hale 1974, Parmotrema mellissii (C. W. Dodge) Hale 1974, Parmotrema perlatum (Huds.) M. Choisy 1952 (previously known as Parmotrema chinense sensu auct.), Parmotrema praesorediosum (Nyl.) Hale 1974, Parmotrema reticulatum (Taylor) M. Choisy 1952 [previously known as Rimelia reticulata (Taylor) Hale & A. Fletcher, 1990], Parmotrema subtinctorium (Zahlbr.) Hale 1974 [previously known as Canomaculina subtinctoria (Zahlbr.) Elix 1997], Parmotrema tinctorum (Despr. ex Nyl.) Hale 1974 and Parmotrema ultralucens (Krog) Hale 1974 [7].
The current study resulted eight species, Parmotrema cetratum (Ach.) Hale 1974, Parmotrema grayanum (Hue) Hale 1974, Parmotrema defectum (Hale) Hale 1974, Parmotrema dilatatum (Vain.) Hale 1974, Parmotrema margaritatum (Hue) Hale 1974, Parmotrema pseudocrinitum (Abbayes) Hale 1974, and Parmotrema subsumptum (Nyl.) Hale 1977 as new records from South Korea. Further, Parmotrema cristiferum (Taylor) Hale 1974, which was identified incorrectly as Parmotrema ultralucens (Krog) Hale 1974 [8], also is added as a new record. Thus, a total of 18 species of Parmotrema are reported from South Korea. The first world monograph of Parmotrema was published under the name Parmelia subgenus Amphigymnia by Hale in 1965 [9]. Later, monographic studies of the genus Parmotrema were carried out by Krog and Swinscow [10], Elix [11, 12], and Divakar and Upreti [13].
MATERIALS AND METHODS
The study was based on specimens deposited in Korean Lichen Research Institute (KoLRI). The lichen samples were identified using stereo and light microscopes. Specifically, a dissecting microscope (SMZ645; Nikon, Tokyo, Japan) was used to identify morphological characteristics of the thallus and the reproductive structures, while a compound microscope (Zeiss Scope. A1; Carl Zeiss, Jena, Germany) was used to study the anatomy of thalli and fruiting bodies. Spot test reactions were carried out on thalli under the compound microscope. Thin layer chromatography (TLC) was performed in solvent system A (toluene:dioxin: acetic acid = 180:45:5) [14]. All examined localities of specimens were mapped using the open source GIS software Quantum GIS 1.7.0 (QGIS). Voucher specimens have been deposited in the herbarium of the Lichen and Allied Bio-resource Center at the KoLRI, Sunchon National University, South Korea. In the identification key, the newly reported species are indicated in bold.
Key to the South Korean species of Parmotrema
1. Thallus lacking isidia and soredia ... 2
1a. Thallus isidiate or sorediate ... 3
2. Thallus eciliate, stictic acid present ... P. eciliatum
2a. Thallus ciliate, salazinic acid present ... P. cetratum
3. Thallus isidiate ... 4
3a. Thallus sorediate ... 8
4. Lobe margins ciliate ... 5
4a. Lobe margins eciliate, isidia cylindrical, medulla C+ red, lecanoric acid present ... P. tinctorum
5. Isidia ciliate ... 6
5a. Isidia eciliate or very rarely ciliate ... 7
6. Isidia coralloid branched and sorediate, medulla K-, C-, alectoronic acid present ... P. mellissii
6a. Isidia cylindrical, simple and not sorediate, medulla K+ yellow, P+ orange red, stictic acid present ... P. crinitum
7. Medulla C+ rose red, gyrophoric acid present ... P. pseudocrinitum
7a. Medulla C-, salazinic acid present ... P. subtinctorium
8. Lobe margins ciliate ... 9
8a. Lobe margins eciliate ... 14
9. Medulla K+ red, salazinic acid present ... 10
9a. Medulla K+ (yellow/orange) or K- ... 12
10. Upper surface faintly maculate, lobes dimorphic, secondary lobes arising from margins in the center of the thallus ... P. margaritatum
10a. Upper surface highly maculate, dimorphic lobes absent ... 11
11. Maculae forming an intricate reticulate network and fissuring into fine cracks ... P. reticulatum
11a. Maculae not forming reticulate cracks, effigurate ... P. subsumptum
12. Medulla P+ orange or red ... 13
12a. Medulla P-, saxicolous ... P. grayanum
13. Stictic acid present ... P. perlatum
13a. Alectoronic acid present ... P. arnoldii
14. Medulla K+ red, salazinic acid present ... P. cristiferum
14a. Medulla K- ... 15
15. Medulla P+ red, protocetraric acid present ... P. dilatatum
15a. Madulla P- ... 16
16. Medulla C+ red, KC+ red, lecanoric acid present ... 17
16a. Medulla C-, KC- ... P. praesorediosum
17. Thallus rugose, lobes 6~12 mm wide, margins crenate, linear soralia < 1 cm long ... P. defectum
17a. Thallus more or less smooth, lobes 6~20 mm wide, margins ascending imbricate, linear soralia > 1 cm long ... P. austrosinense
Species descriptions
Parmotrema austrosinense (Zahlbr.) Hale, Phytologia 28: 335 (1974)
Parmelia austrosinensis Zahlbr., in Handel-Mazzetti, Symb. Sinic. 3: 180 (1930).
Thallus foliose, 6~10 cm broad, loosely attached to the substratum. Lobes rotund, 5~15 mm wide; margins ascending imbricate, sinuous, entire, eciliate. Upper surface slightly maculate, ash grey to grey green, shine, smooth and older parts reticulately cracked, without isidia, sorediate. Soralia marginal, submarginal, linear, very wavy and continuous for a few centimeters in length. Soredia farinose to granular. Medulla white. Lower surface finely reticulately wrinkled, black with a broad 4~6 mm wide yellowish brown, light tan to white mottled, shiny erhizinate marginal zone. Rhizines, sparse, simple, unevenly distributed, short, up to 1 mm long. Apothecia and pycnidia not seen in the specimens examined (Fig. 1A~1E).
Fig. 1.

A~E, Parmotrema species. A, P. austrosinense (J. S. Hur, 86774); B, P. cetratum (J. S. Hur, 40060); C, P. cristiferum (J. S. Hur, 30473); D, P. defectum (J. S. Hur, 41654); E, P. dilatatum (J. S. Hur, 60349). F~J, Parmotrema species. F, P. grayanum (J. S. Hur, 40534); G, P. margaritatum (J. S. Hur, 70794); H, P. perlatum (J. S. Hur, 70649); I, P. praesorediosum (J. S. Hur, 70007); J, P. pseudocrinitum (J. S. Hur, 80797). K~N, Parmotrema species. K, P. reticulatum (J. S. Hur, 41659); L, P. subsumptum (J. S. Hur, 30431); M, P. subtinctorium (J. S. Hur, 30029); N, P. tinctorum (J. S. Hur, 70183) (scale bars: A~N = 1 cm).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K-C+ (red), KC+ (red), P-. TLC: atranorin, chloroatranorin, lecanoric acid, unknown 1 and unknown 2 (Fig. 2).
Fig. 2.
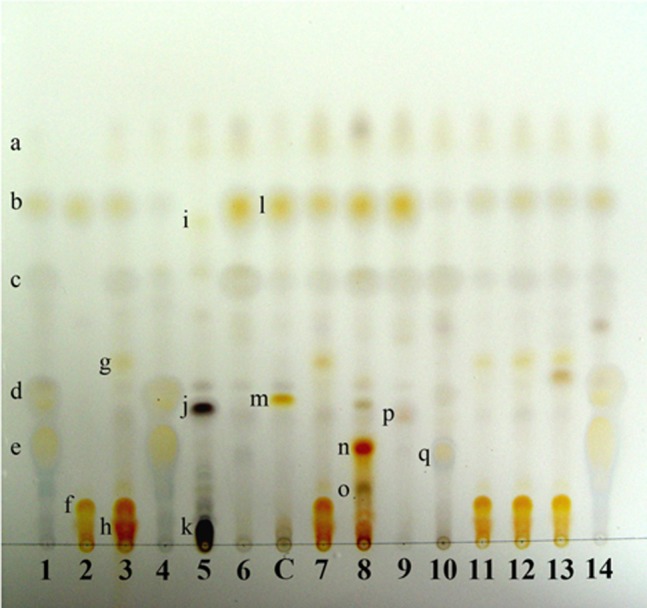
Thin layer chromatography profile of Parmotrema species in solvent system A. 1, P. austrosinense with chloroatranorin (a), atranorin (b), unknown 1 (c), unknown 2 (d), lecanoric acid (e); 2, P. cetratum with salazinic acid (f); 3, P. cristiferum with unknown 3 (g), consalazinic acid (h); 4, P. defectum; 5, P. dilatatum with usnic acid (i), unknown 4 (j), protocetraric acid (k); 6, P. grayanum; C, control [Lethariella cladonioides (Nyl.) Krog] with atranorin (l), norstictic acid (m); 7, P. margaritatum; 8, P. perlatum with stictic acid (n), menegazziaic acid (o); 9, P. praesorediosum with unknown 5 (p); 10, P. pseudocrinitum with gyrophoric acid (q); 11, P. reticulatum; 12, P. subsumptum; 13, P. subtinctorium; 14, P. tinctorum.
Remarks
P. austrosinensis has a thallus rather soft to the touch with eciliate, marginal soralia, and C+ red medulla. According to Divakar and Upreti [13], this species resembles P. sancti-angelii and P. hababianum in its sorediate condition. However, P. sancti-angelii is different from P. austrosinensis in that it has a coriaceous, ciliate, granular sorediate thallus with gyrophoric acid in the medulla. P. hababianum, which has a soft thallus, differs from P. austrosinensis in having cilia along the margins and C-medulla. P. cooperi also resembles P. austrosinensis, but it has ciliate lobe margins [12].
Ecology and distribution
In South Korea, this species is found on the bark of Pinus sp. and Cherry trees in Mt. Halla, Mt. Illim, Mt. Ungseokbong, Baeal Beach, Gwangpo Village and the Wondangbong area. This is a pantropical species that has been identified in tropical regions of Africa [9]. It has also been recorded from Asia, America and Central American regions.
Selected specimens examined
Mt. Halla, Jeju-si, on bark (Cherry), 33°22'17.7" N, 126°34'15.2" E, alt. 1,300 m, 9 Aug 2008, J. S. Hur, 086774; Baeal Beach, Taein-dong, Gwangyang-si, Jeollanam-do, on bark (Pinus sp.), 34°57'3.90" N, 127°45' 6.55" E, alt. 7 m, 16 Jan 2010, J. S. Hur, M. H. Jeong, GW1030; Gwangpo Village, Gadeok-ri, Hadong-gun, Gyeongsangnam-do, on bark (Pinus sp.), 34°58'2.91" N, 127°48'9.14" E, alt. 37 m, 25 Jan 2010, J. S. Hur, M. H. Jeong, GW1053; Wondangbong, Jeju Island, on bark (Pinus sp.), 33°31'7.49" N, 126°35'35.2" E, alt. 45 m, 19 Apr 2009, J. S. Hur, X. Y. Wang, J. A. Ryu, J. Y. Hur, 090004; Mt. Ungseokbong, Sancheong-eup, Sancheong-gun, Gyeongsangnam-do, on bark (Cherry tree), 35°23'25.7" N, 127°52'6.68" E, alt. 173 m, 11 Oct 2010, X. Y. Wang, J. A. Ryu, 101342; Mt. Illim, Ungchi-myeon, Boseong-gun, Jeollanam-do, on bark (Pinus sp.), 34°41'17.7" N, 127°00'57.3" E, alt. 220 m, 1 Sep 2005, J. S. Hur, 050374 (Fig. 3).
Fig. 3.

Distribution of Parmotrema species in South Korea. A, P. austrosinense; B, P. praesorediosum; C, P. cetratum (▲), P. grayanum (★), P. subsumptum (●), P. subtinctorium (×); D, P. reticulatum; E, P. cristiferum (+), P. defectum (▲), P. dilatatum (×), P. margaritatum (★), P. pseudocrinitum (■); F, P. tinctorum.
Parmotrema cetratum (Ach.) Hale, Phytologia 28: 335 (1974)
Parmelia cetrata Ach., Syn. Lich.: 198 (1814).
Rimelia cetrata (Ach.) Hale & A. Fletcher, Bryologist 93: 26 (1990).
Thallus foliose, loosely adnate to the substratum, 5~12 cm across. Lobes apically round, sub irregular, often laciniate, up to 8 mm wide; margins crenate, dentate, lateral margins in the central part of the thallus lacinules, subdichotomously divided and tapering, ciliate. Cilia sparse to dense, simple to sparingly branched, 0.5~2 mm long, black. Upper surface pale grey to mineral grey, without soredia, isidia or dactyls densely maculate. Maculae forming an intricate reticulate network and fissuring into fine cracks. Soredia, when present, developing either laminally or marginally. Medulla white. Lower surface black, glossy, with a narrow, 1~2 mm wide, erhizinate marginal zone. Rhizines dense, simple to squarrosely branched, black. Apothecia not observed. Conidiomata pycnidial, immersed, laminal, black. Conidia filiform, 9~12 × 1 µm (Fig. 1A~1E).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K+ (yellow to red), C-, KC+ (red), P+ (orange). TLC: atranorin, chloroatranorin, salazinic acid, consalazinic acid (Fig. 2).
Remarks
P. cetratum is characterized by a smooth cortex, reticulate maculate upper surface, and esorediate thallus with salazinic acid in the medulla. This species is similar to P. reticulatum in chemistry, but the latter species differs in having a sorediate thallus.
Ecology and distribution
In South Korea this species has only been recorded on a rock from a single location on Mt. Sonum. According to Elix [12], this is a very rare species that has been recorded from Asian, African and Central American regions.
Specimens examined
Mt. Sonun, Gochang-gun, on rock, 35°29'41.2" N, 126°34'14.1" E, alt. 295 m, 20 Apr 2004, 040060 (Fig. 3).
Parmotrema cristiferum (Taylor) Hale, Phytologia 28: 335 (1974)
Parmelia cristifera Taylor, Lond. J. Bot. 6: 165 (1847).
Parmelia perforata var. ulophylla Meyen & Flot., Nova Acta Acad. Caes. Leop. Carol. Nat. Curiosorum, Suppl. 119: 218 (1843).
Thallus foliose, loosely attached to the substratum, coriaceous, 4~6 cm broad, lobes rotund, 5~10 mm wide; margins entire, crenate, eciliate. Upper surface pale grey, dull, emaculate, without isidia, sorediate. Soralia marginal to submarginal, sinuous and revolute. Medulla white. Lower surface black, smooth with broad, pale brown to tan, erhizinate marginal zone. Rhizines, sparse, simple, short, up to 1 mm long. Apothecia and pycnidia not seen in the specimens examined (Fig. 1A~1E).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K+ (red), C-, KC-, P+ (orange-red). TLC: atranorin, chloroatranorin, salazinic acid and unknown (Fig. 2).
Remarks
This is the first record of this species in South Korea. P. cristiferum is characterized by the loosely adnate, coriaceous thallus, eciliate lobes, marginal soralia and salazinic acid in the medulla. According to Divakar and Upreti [13], this species is closely related to P. stuppeum in having marginal soralia and the presence of salazinic acid, but it differs in that it has ciliate margins.
During the current study, the specimens referred to under Parmotrema ultralucens (Krog) Hale [8] were identified as Parmotrema cristiferum.
Ecology and distribution
In South Korea this species has only been recorded on a rock from a single location on Mt. Naejang. However, this organism is a cosmopolitan species that is widely distributed in tropical and subtropical areas [10, 12].
Specimens examined
Mt. Naejang, on rock, 35°29'38" N, 126°54'28" E, alt. 590 m, 29 Jun 2003, J. S. Hur, 030473 (Fig. 3).
Parmotrema defectum (Hale) Hale, Phytologia 28: 335 (1974)
Parmelia defecta Hale, Contrib. U. S. Nat. Herb. 36: 244 (1965).
Thallus foliose, closely attached to the substratum, coriaceous, 5~8 cm broad, Lobes round to irregular, 5~10 mm wide; margins crenate, eciliate. Upper surface pale grey, smooth, somewhat shiny, rugose, cracked in the center, sorediate. Soralia marginal, linear, but not continuous for about 1 cm, sorediate margins revolute. Medulla white. Lower surface minutely wrinkled, black with smooth, pale brown to tan, erhizinate marginal zone. Rhizines, sparse, simple, short, up to 1 mm long. Apothecia and pycnidia not seen in the specimens examined (Fig. 1A~1E).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K-, C+ (rose to red), KC+ (red), P-. TLC: atranorin, chloroatranorin and lecanoric acid (Fig. 2).
Remarks
This is the first report of P. defectum from South Korea. This species is characterized by a rugose, emaculate, thallus with eciliate margins and lecanoric acid in the medulla. According to Divakar and Upreti [13], P. defectum is a saxicolous species, but this species was found on the bark of Pinus sp.
Ecology and distribution
In South Korea, this species was recorded on a rock and bark from two locations, Bokil-do and from a large bridge in Geumho. Outside of South Korea, this species has been recorded in African and South Asian regions.
Specimens examined
Bokil-do, on bark, 34°09'31.7" N, 126°33'27.7" E, alt. 5 m, 31 Dec 2004, J. S. Hur, 041654; Geumho, Geumho-dong, Gwangyang-si, Jeollanam-do, on rock, 34°56'6.12" N, 127°43'5.71" E, alt. 14 m, 16 Jan 2010, J. S. Hur, M. H. Jeong, GW1012 (Fig. 3).
Parmotrema dilatatum (Vain.) Hale, Phytologia 28: 335 (1974)
Parmelia dilatata Vain., Acta Soc. Fauna Flora Fenn. 7: 32 (1890).
Thallus foliose, loosely attached to the substratum, coriaceous, 4~6 cm broad. Lobes rotund, 5~10 mm wide; margins entire, crenate, eciliate. Upper surface pale grey, shiny, emaculate, without isidia, sorediate. Soralia marginal, globular or confluent, soredia granular. Medulla white. Lower surface black, smooth with broad, pale brown to tan, erhizinate marginal zone. Rhizines, sparse, simple, short, up to 1 mm long. Apothecia and pycnidia not seen in the specimens examined (Fig. 1A~1E).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K-, C-, KC+ (red), P+ (red). TLC: atranorin, chloroatranorin, usnic acid, protocetraric acid and unknown 4 (Fig. 2).
Remarks
P. dilatatum is characterized by coriaceous, eciliate lobes, marginal soralia and protocetraric acid in the medulla. Externally, this species externally resembles P. cristiferum, but it differs due to the presence of protocetraric acid in the medulla [13].
Ecology and distribution
In South Korea, this species has only been recorded on a rock from a single locality of Mt. Chiri. Apart from South Korea, this species has been reported from Asian and Central American regions.
Specimens examined
Mt. Chiri, on bark, 35°18'48.9" N, 127°35'13.5" E, alt. 1,120 m, 18 Jun 2006, J. S. Hur, 060349 (Fig. 3).
Parmotrema grayanum (Hue) Hale, Phytologia 28: 336 (1974)
Parmelia grayana Hue, Nouv. Arch. Mus. Hist. Nat., Paris, 4 Sér. 1: 184 (1899).
Thallus foliose, loosely attached to the substratum, 3~6 cm wide. Lobes rotund, crowded, rather narrow, 3~6 mm wide, margins ascending, crenate, ciliate; cilia simple to branched, black, thick, 0.5~2 mm long. Upper surface ashy-grey, smooth, epruinose, emaculate, sorediate near the margins, developed on apices of dents, linear to subglobose, soredia granular, often with a grey-brown tinge. Medulla white. Lower surface faintly wrinkled, black with a broad, brown, erhizinate marginal zone. Rhizines at the center, sparse, black, simple, 1~2 mm long. Apothecia not seen. Pycnidia present at the periphery of lobes, black, immersed. Conidia rod shaped, 6~8 µm long (Fig. 1F~1J).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; Medulla K-, C-, KC-, P-. TLC: atranorin and chloroatranorin (Fig. 2).
Remarks
P. grayanum closely resembles P. praesorediosum in that it has sorediate margins and negative chemical reactions, but P. grayanum can be differentiated by the presence of cilia along the margins.
Ecology and distribution
In South Korea this species has only been recorded from rock surfaces in Bogil Island, Cheongsan Island, Saryang Island, Geomun Island, Mt. Sobaek, the Yongbawi seaside and Daeki Valley. This species has also been reported in East Asian, African and Central American regions.
Selected specimens examined
Bogil Island, Shinan Co., Jeonam Prov., on rock, 34°07'8.62" N, 126°30'7.28" E, alt. 19 m, 23 Jun 2011, X. Y. Wang, J. A. Ryu, 110683; Cheongsan Island, Wando Co., Jeonam Prov., on rock, 34°12'21.8" N, 126°54'35.7" E, alt. 5 m, 23 Jun 2011, X. Y. Wang, J. A. Ryu, 110751; Yongbawi Seaside, Yongam Village, Ucheon-ri, Goheung-gun, Jeollanam-do, on rock, 34°35'7.65" N, 127°30'3.75" E, alt. 10 m, 19 Feb 2010, Y. Joshi, H. S. Jeon, G. S. Han, 100355; Daeki Valley, Nanhae-gun, Gyengshangnam-do, on rock, 34°45'38.3" N, 128°02'53.6" E alt., 280 m, 11 Nov 2007, J. S. Hur, 070977; Geomun Island, Samsan-myeon, Jeollanam-do, on rock, 34°00'28.9" N, 127°19'14.8" E alt. 80 m, 24 Mar 2007, J. S. Hur, 070172; Mt. Sobaek, on rock, 36°57'13.1" N, 128°28'38.2" E alt. 1,386 m, 2 Oct 2003, J. S. Hur, 030783; Saryang Island, Saryang-myeon, Sangnam-do, on rock, 34°50'13.9" N, 128°10'44.0" E, alt. 118 m, 17 Mar 2007, J. S. Hur, 070035; Fire Goheung, on rock, 34°29'17.3" N, 127°21'27.4" E, alt. 5 m, 6 Aug 2004, J. S. Hur, 040534 (Fig. 3).
Parmotrema margaritatum (Hue) Hale, Phytologia 28: 337 (1974)
Parmelia margaritata Hue, Nouv. Arch. Mus. Paris. Ser. 4 1:193 (1899).
Thallus foliose, loosely attached to the substratum, medium sized, 3~6 cm across, rather coriaceous to crisp. Lobes rotund, irregular, 5~8 mm wide, with several ascending, simple to dichotomously branched cilia. Cilia simple, 0.5 to 1 mm long, moderately dense. Upper surface pale grey to grey, smooth, dull, maculate, without isidia, sorediate. Soralia terminal on the lacinules, rounded or confluent and sinuous, sorediate lobes revolute. Medulla white. Lower surface black, with shiny, brown erhizinate marginal zone. Rhizines simple to slightly branched, dense in the central part. Apothecia and pycnidia not seen (Fig. 1F~1J).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K+ (red), C-, KC-, P+ (orange red). TLC: atranorin, chloroatranorin, salazinic acid (Fig. 2).
Remarks
P. margaritatum shows a close resemblance to P. stuppeum in the presence of marginal cilia, soralia and similar chemical substances, but the presence of secondary lobules and a faint maculate upper surface differentiate it from the latter. P. leucosemothetum also very closely resembles P. margaritatum, but has a highly maculate upper surface and lacks secondary lobules [13].
Ecology and distribution
In South Korea, this species has been recorded from Mt. Kongduck, Mt. Naejang, and the Asan Hwasun area. Outside of South Korea, this species has been reported from India [13], the United States [9], and Venezuela [15].
Specimens examined
Mt. Kongduck, on bark (Pinus sp.), 36°44'58.6" N, 128°15'52.1" E, alt. 628 m, 20 Jun 2007, J. S. Hur, 070794; Mt. Naejang, on rock, 29 Jun 2003, J. S. Hur, 030473; Hwasun, Asan, on rock, 34°10'14.0" N, 127°08'45.0" E, alt. 500 m, 8 Oct 2005, J. S. Hur, 050570 (Fig. 3).
Parmotrema perlatum (Huds.) M. Choisy, Bull. Mem. Soc. Linn. Lyon 21: 174 (1952)
Lichen perlatus Huds., Fl. Angl. 1: 448 (1762).
Imbricaria ciliata (DC.) Arnold, Flora, Jena 67: 158 (1884).
Imbricaria perlata (Huds.) Körb., Lichenogr. Germ. (Breslau): 8 (1846).
Lobaria perlata (Huds.) Hoffm., Deutschl. Fl., Zweiter Theil (Erlangen): 148 (1796).
Parmelia coniocarpa Laurer, Linnaea 2: 39 (1827).
Parmelia perlata (Huds.) Ach., Method. Lich.: 216 (1803).
Parmelia perlata var. ciliata (DC.) Duby, Bot. Gall., Ed. 2 (Paris) 2: 601 (1830).
Parmelia trichotera Hue, J. Bot., Paris 12: 245 (1898).
Parmotrema perlatum var. ciliata (DC.) M. Choisy, Bull. Mens. Soc. Linn. Lyon 21: 175 (1952).
Parmotrema trichotera (Hue) M. Choisy, Bull. Mens. Soc. Linn. Lyon 21: 175 (1952).
Platysma perlatum (Huds.) Frege, Deutsch. Bot. Taschenb. 2: 167 (1812).
Parmotrema chinense auct., non (Osbeck) Hale & Ahti, Taxon 35: 133 (1986).
Thallus foliose, membranaceous to coriaceous, loosely attached to the substratum, 5~8 cm broad. Lobes rotund, irregular, 5~8 mm wide; margins entire to crenate, but lacking sublinear lacinae, eciliate. Cilia simple to branched, rather long (0.5~2 mm), moderately dense to dense. Upper surface pale grey to mineral grey, dull, without isidia, sorediate. Soralia marginal to submarginal, causing lobe margins to become revolute and suberect, ultimately appearing labriform, soredia granular. Medulla white. Lower surface black, shiny with a broad 3~6 mm wide, brown to tan erhizinate marginal zone. Rhizines moderately dense, simple, up to 2 mm long. Apothecia and Pycnidia not seen (Fig. 1F~1J).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K+ (yellow), C-, KC-, P+ (orange-red). TLC: atranorin, chloroatranorin, stictic acid, constictic acid, menegazziaic acid (trace) and norstictic acid (trace) (Fig. 2).
Remarks
P. perlatum is characterized by a loosely attached thallus, marginal to sub marginal soralia with strongly revolute and suberect margins, and the presence of stictic acid in the medulla. The organism resembles Parmotrema reticulatum in the nature of soralia, but the latter species has a maculate, reticulately cracked upper surface and salazinic acid in the medulla. P. perlatum is similar to P. crinitum due to the presence of a brown to tan erhizinate marginal zone and stictic acid complex in the medulla, but the latter species has an isidiate upper surface [13]. According to Park [16], the species description was given under the species name P. chinense (Osbeck) Hale & Ahti.
According to Louwhoff's descriptions [17], P. perlatum is sensitive to air pollution and prefers well-lit, neutral to somewhat acid-barked, broad-leaved trees and siliceous rocks and coastal rocks where illumination is moderate to good. However, the recent revisionary work conducted by Jabłońska et al. [18] stated that P. perlatum is close to extinct from Poland.
Ecology and distribution
In South Korea this species has been recorded from Mt. Ungseok, Mt. Chiri, Mt. Hambeak, Mt. Taebaek, and the Sanchong area. P. perlatum is a cosmopolitan lichen and widespread in both the southern and northern hemisphere and tropical areas [9, 17]. This species has been identified in many European countries [18], as well as other Asian countries including Australia [12], India [13], Japan [19], and Taiwan [4].
Selected specimens examined
Mt. Ungseok, Sancheong-eup, Sancheong-gun, Gyeongsangnam-do, on bark (Quercus sp.), 35°22'53.3" N, 127°51'17.6" E, alt. 783 m, 16 Oct 2007, 070883; Mt. Chiri, on bark, 35°22'07.3" N, 127°34'52.7" E, alt. 480 m, J. S. Hur, 18 Jun 2006, 060376; Mt. Hambeak, on bark (Quercus sp.), 37°11'47.4" N, 128°54'53.6" E, alt. 1355 m, J. S. Hur, 19 Jun 2007, 070649; Jiri (Piagol, Market Place), on bark (Quercus sp.), 35°17'45.1" N, 127°33'38.5" E, alt. 1,202 m, 27 Sep 2006, J. S. Hur, 060774; Mt. Taebaek, on bark (Quercus sp.), 37°12'35.3" N, 128°55'11.9" E, alt. 1,399 m, J. S. Hur, 25 May 2008, 080279; Sanchong, Gyeongnam Province, Woong-Seok-Bong, on bark (Quercus sp.), 35°22'55.0" N, 127°51'20.7" E, alt. 795 m, 16 Oct 2007, J. S. Hur, 070885 (Fig. 3).
Parmotrema praesorediosum (Nyl.) Hale, Phytologia 28: 338 (1974)
Parmelia praesorediosa Nyl., Sert. Lich. Trop. Labuan Singapore: 18 (1891).
Thallus foliose, adnate to the substratum, 3~10 cm across. Lobes round, 4~10 mm wide; margins entire or crenate, eciliate, sorediate. Upper surface pale grey to grey, smooth, dull, emaculate, weakly rugose, lacking isidia, sorediate. Soralia marginal, linear to crescent shaped, granular. Medulla white. Lower surface black, minutely rugose, with shiny, mottled, ivory or brown, erhizinate marginal zone. Rhizines sparse, simple, short. Apothecia and pycnidia not seen (Fig. 1F~1J).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K-, C-, KC-, P- TLC: atranorin, chloroatranorin, fatty acids (protopraesorediosic acid, praesorediosic acid) [13] and unknown 5 (Fig. 2).
Remarks
P. praesorediosum is characterized by an adnate coriaceous thallus, eciliate lobes with ascending sorediate margins and fatty acid in the medulla. Externally, P. grayanum is similar to P. praesorediosum, but the former has ciliate margins [12]. According to Divakar and Upreti [13], the chemical composition of P. praesorediosum is similar to that of P. mesotropum, but the latter differs in having esorediate thalli.
Ecology and distribution
This species is very common on exposed rock surfaces [13] and has been reported from Geomun Island, Saryang Island, Jeob Island, Balpo, Changchon Village, Gopo Village and Mt. Nogudwit in South Korea. Apart from this country, this species has been reported from East Asia, South Asia and Central America.
Selected specimens examined
Geomun Island, Samsan-myeon, Jeollanam-do, on bark (Camellia japonica), 34°00'34.2" N, 127°19'12.4" E, alt. 61 m, 24 Mar 2007, J. S. Hur, 070151; Saryang Island, Saryang-myeon, Sangnam-do, on bark, 34°50'06.4" N, 128°10'53.7" E, alt. 26 m, 17 Mar 2007, J. S. Hur, 070007; Balpo, Goheung-gun, Jeollanam-do, on rock, 34°29'17.3" N, 127°21'27.4" E, alt. 5 m, 6 Aug 2004, J. S. Hur, 040536; Changchon Village, Jinsang-myeon, Gwangyang-si, Jeollanam-do, on rock, 34°58'5.92" N, 127°44'0.71" E, alt. 43 m, 16 Jan 2010, J. S. Hur, M. H. Jeong, GW1039; Gopo Village, Geumseong-myeon, Hadong-gun, Gyeongsangnam-do, on bark (Pinus sp.), 34°57'7.45" N, 127°46'7.36" E, alt. 4m, 25 Jan 2010, J. S. Hur, M. H. Jeong, GW1051; Jeob Island, Jeonam Prov., Jindo-Co., on rock, 34°23'6.80" N, 126°18'25.1" E, alt. 1 m, 3 Jun 2011, X. Y. Wang, J. A. Ryu, 110498; Mt. Nogudwit, Namhae-gun, Gyeongsangnam-do, on rock, 34°45'20.3" N, 128°02'39.4" E, alt. 210 m, 11 Nov 2007, 070942 (Fig. 3).
Parmotrema pseudocrinitum (Abbayes) Hale, Phytologia 28: 338 (1974)
Parmelia pseudocrinita des Abb., Bull. Inst. Fr. Afr. Noire 20: 19 (1958).
Thallus foliose, loosely attached to the substratum, coriaceous, 4~8 cm broad, lobes imbricate, 5~10 mm wide; margins crenate, ciliate. Cilia evenly distributed, coarse, 0.5~1 mm long. Upper surface pale grey, smooth, somewhat shiny, emaculate, esorediate, with isidia. Isidia laminal, sparse to dense, simple, cylindrical or coralloid. Medulla white. Lower surface black with smooth, pale brown to tan, erhizinate marginal zone. Rhizines, sparse, simple, short, up to 2 mm long. Apothecia and pycnidia not seen in the specimens examined (Fig. 1F~1J).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K-, C+ (rose to red), KC+ (red), P-. TLC: atranorin, chloroatranorin and gyrophoric acid (Fig. 2).
Remarks
P. pseudocrinitum is characterized by emaculate, isidiate upper surface, ciliate margin and the presence of gyrophoric acid in the medulla. P. crinitum also has similar ciliate and isidiate conditions, but differs from P. pseudocrinitum in having a stictic acid complex in the medulla.
Ecology and distribution
In South Korea, this species has been reported on bark in the Mt. Halla area. Outside South Korea, this species has been reported from East Africa [10], India [13], and Thailand [20].
Specimens examined
Mt. Halla, Jeju-si, on bark, 33°21'18.8" N, 126°30'00.4" E, alt. 1,492 m, 10 Aug 2008, J. S. Hur, 080787, 080797 (Fig. 3).
Parmotrema reticulatum (Taylor) M. Choisy, Bull. Mens. Soc. Linn. Lyon 21: 148 (1952)
Parmelia reticulata Taylor, in Mackay, Fl. Hibern. 2: 148 (1836).
Canomaculina leucosemotheta (Hue) Elix, Mycotaxon 65: 477 (1997).
Parmelia ciliata (DC.) Nyl., Flora, Jena 61: 247 (1878).
Parmelia ciliata (Nyl.) Gyeln., Feddes Repert. 30: 225 (1932).
Parmelia concors Kremp., Verh. Zool. Bot. Ges. Wien 30: 337 (1880).
Parmelia laevigata var. reticulata (Taylor) Linds., Trans. Linn. Soc. Lond., Bot. 25: 514 (1866).
Parmelia leucosemotheta Hue, Nouv. Arch. Mus. Hist. Nat., Paris, 4 Sér. 1: 192 (1899).
Parmelia macquariensis C.W. Dodge, Nova Hedwigia 19: 450 (1970).
Parmelia perforata var. ciliata Nyl., Annls Sci. Nat., Bot., Sér. 4 15: 373 (1861).
Parmelia pseudovirens Gyeln., Reprium Nov. Spec. Regni Veg. 29: 288 (1931).
Parmelia urceolata var. sorediifera Müll. Arg., Flora, Jena 63: 266 (1880).
Parmelia urceolata var. subcetrata Müll. Arg., Flora, Jena 66: 46 (1883).
Parmelia virens var. sorediata Müll. Arg., Flora, Jena 69: 256 (1886).
Parmotrema leucosemothetum (Hue) Hale, Phytologia 28: 337 (1974).
Parmotrema pseudovirens (Gyeln.) Elix, Mycotaxon 47: 127 (1993).
Rimelia reticulata (Taylor) Hale & A. Fletcher, Bryologist 93: 28 (1990).
Parmelia clavulifera Räsänen, Ann. Zool. Bot. Fenn., Vanamo 20: 4 (1944).
Parmotrema clavuliferum (Räsänen) Streimann, Bibliotheca Lich. 22: 93 (1986).
Rimelia clavulifera (Räsänen) Kurok., J. Jpn. Bot. 66: 158 (1991).
Thallus foliose, adnate to loosely adnate to the substratum, 5~15 cm across. Lobes apically round to sub rotund, imbricate 4~12 mm wide; central part of the thallus lacinules, lacunae up to 2 mm wide, ciliate. Cilia sparse to dense, simple to sparingly branched, 0.5~2 mm long, black. Upper surface pale grey to mineral grey, sorediate, isidia or dactyls absent, densely maculate. Maculae forming an intricate reticulate network and fissuring into fine cracks. Sometimes flaking off in older parts. Soredia marginal to submarginal. Linear to capitates, linear soralia on lateral margins and capitate soralia developing on apices of lacinae, which becomes involute, soredia granular. Medulla white. Lower surface black, glossy, with narrow, 1~2 mm wide, erhizinate marginal zone or rhizinate up to margins. Rhizines dense, simple to squarrosely branched, black. Apothecia and pycnidia not observed (Fig. 1K~1N).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K+ (yellow to red), C-, KC-, P+ (orange-red). TLC: atranorin, chloroatranorin, salazinic acid, consalazinic acid (Fig. 2).
Remarks
P. reticulatum is characterized by marginal to submarginal soralia and the presence of salazinic acid in the medulla. This species is similar to P. cetratum in chemistry, but the latter species differs in that it has an esorediate upper surface.
Ecology and distribution
In South Korea this species has been recorded from Mt. Halla, Mt. Talmasan, Mt. Hugseok, Mt. Naejang, Mt. Kongduck, Mt. Jumbong, Mt. Odae, and the Bokildo area. According to Elix [12] and Kurokawa and Lai [4], this species is widely distributed in tropical and temperate regions.
Selected specimens examined
Mt. Halla, Jeju-si, on bark, 33°22'17.7" N, 126°34'15.2" E, alt. 1,300 m, 9 Aug 2008, J. S. Hur, 080747, 080748, 080757, 080758, 080766, 080770, 080771; Mt. Talmasan, Songji-ri, Haenum-gun, Jeollanam-do, on rock, 34°22'45.1" N, 126°35'07.5" E, alt. 389 m, 26 Jul 2005, J. S. Hur, 050335; Mt. Hugseok, Gahak-ri, Haenam-gun, Jeollanam-do, on bark, 34°41'21.1"N, 126°40'47.5"E, alt. 230 m, 23 Sep 2005, J. S. Hur, 050499; Mt. Naejang, on rock, 35°29'41.0" N, 126°52'53.3" E, alt. 650 m, 8 Aug 2003, J. S. Hur, 030616; Mt. Kongduck, on rock, 36°45'12.4" N, 128°16'07.6" E, alt. 628 m, 20 Jun 2007, J. S. Hur, 070811; Jeju-do, on bark, 33°26'04.4" N, 126°34'01.7" E, 29 Aug 2004, J. S. Hur, 040872; Mt. Jumbong, on bark, 38°03'56.4" N, 128°26'51.7" E, alt. 600 m, 9 Oct 2004, J. S. Hur, 041324; Mt. Odae, on bark, 37°46'59.1" N, 128°33'59.1" E, alt. 890 m, 7 May 2004, J. S. Hur, 040418; Bokildo, on bark, 34°09'31.7" N, 126°33'27.7" E, alt. 5 m, 31 Dec 2004, J. S. Hur, 041659 (Fig. 3).
Parmotrema subsumptum (Nyl.) Hale, Mycotaxon 5: 434 (1977)
Parmelia subsumpta Nyl., Flora, Jena 52: 117 (1869).
Canomaculina subsumpta (Nyl.) Elix, Mycotaxon 65: 477 (1997).
Rimeliella subsumpta (Nyl.) Kurok., Ann. Tsukuba Bot. Gard. 10: 9 (1991).
Thallus foliose, loosely attached to the substratum, 6~12 cm broad. Lobes rotund, 7~15 mm wide; margins crenate, ciliate. Cilia dense, simple, black, markedly tapered, 0.5~1 mm long. Upper surface mineral grey to grey, white maculate, maculae effigurate, often cracked in older parts, without isidia, sorediate. Soralia linear, marginal in the central part, sorediate lobes involute. Medulla white, sometimes pale reddish with age due to salazinic acid in the medulla. Lower surface pale brown, rhizines dense. Rhizines black, short, dimorphic, marginal rhizines simple, central rhizines squarrosely branched. Apothecia and pycnidia not seen (Fig. 1K~1N).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K+ (yellow to red), C-, KC-, P+ (orange). TLC: atranorin, chloroatranorin and salazinic acid (Fig. 2).
Remarks
P. subsumptum is characterized by sorediate upper surface and presence of salazinic acid in the medulla. In chemistry, this species is similar to P. subtinctorium, but the latter has isidia on the thallus.
Ecology and distribution
According to Divakar and Upreti [13], this species is only known to be corticolous, but it has been reported as corticolous and saxicolous in areas of Jeju-do, Bogil Island, Mt. Sunwoon, Mt. Naejang, Mt. Sorodo, Mt. Wolchul, and the Haein Temple area in South Korea. Outside of South Korea, this species has been reported in East and South Asia and Central America.
Selected specimens examined
Jeju-do, on bark, 33°27'15.4" N, 126°33'41.1" E, alt. 370 m, 29 Aug 2004, J. S. Hur, 040865, 040868; Bogil Island, on rock, 34°09'14.7" N, 126°37'33.2" E, alt. 5 m, 31 Dec 2004, J. S. Hur, 041668, 041669, 041670; Mt. Sunwoon, on bark, 35°29'46.9" N, 126°53'40.7" E, alt. 140 m, 11 May 2003, J. S. Hur, 030233; Mt. Naejang, on rock, 36°57'13.1" N, 128°28'38.2" E, alt., 29 Jun 2003, J. S. Hur, 030431, 030431-2; Mt. Sorodo, on bark, 35°48'11.2" N, 129°18'47.3" E, alt. 15 m, 23 Mar 2003, J. S. Hur, 030071; Mt. Wolchul, on rock, 34°59'55.9" N, 127°16'42.10" E, alt. 816 m, 8 Jun 2003, J. S. Hur, 030324; Haein Temple, Mt. Gaya, 35°47'20.7" N, 128°05'51.5" E, 22 Apr 2003, J. S. Hur, 030151 (Fig. 3).
Parmotrema subtinctorium (Zahlbr.) Hale, Phytologia 28: 339 (1974)
Parmelia subtinctoria Zahlbr., in Handel-Mazzetti, Symb. Sinic. 3: 193 (1930).
Canomaculina subtinctoria (Zahlbr.) Elix, Mycotaxon 65: 477 (1997).
Parmelia virens f. isidiosa Müll. Arg., Annln K. K. Naturh. Hofmus. Wien 7: 303 (1892).
Rimelia subtinctoria (Zahlbr.) Kurok., Ann. Tsukuba Bot. Gard. 10: 10 (1991).
Thallus foliose, loosely attached to the substratum, 4~10 cm broad. Lobes rotund, 5~15 mm wide; margins crenate, dentate, ciliate. Cilia dense, simple to branched, markedly tapered, 0.5~2 mm long. Upper surface mineral grey to grey, turning buff in the herbarium with age, white maculate, maculae effigurate, often cracked in older parts, without soredia, isidiate. Isidia laminal, filiform, simple to branched, sometimes ciliate, dark brown tipped. Medulla white, sometimes reddish with age due to salazinic acid in the medulla. Lower surface dark to pale brown, with shiny, pale brown, narrow marginal zone. Rhizines simple, brown to black, uniformly distributed, up to 2mm long. Apothecia and pycnidia not seen (Fig. 1K~1N).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K+ (yellow to red), C-, KC-, P+ (orange). TLC: atranorin, chloroatranorin and salazinic acid (Fig. 2).
Remarks
P. subtinctorium is characterized by an isidiate upper surface and the presence of salazinic acid in the medulla. In chemistry, this species is similar to P. subsumptum, but the latter has a marginal soralia.
Ecology and distribution
This species has a saxicolous and corticolous habitat and has been recorded from areas of Mt. Jiri, Mt. Sunun, Mt. Sunwoon and Mt. Baekwoon. This species has also been reported in Asia, Africa and Central America.
Specimens examined
Mt. Jiri, Baemsagol, 35°29'44.3" N, 126°53'27.6" E, on bark, alt. 530 m, 22 Mar 2003, J. S. Hur, 030029; Mt. Sunun, 35°29'46.9" N, 126°53'40.7" E, 11 May 2003, J. S. Hur, 030260; Mt. Baekwoon, on rock, 1 Jun 2003, J. S. Hur, 030275; Mt. Sunwoon, 35°29'46.9" N, 126°53'40.7" E, alt. 45 m, 11 May 2003, J. S. Hur, 030259 (Fig. 3).
Parmotrema tinctorum (Despr. ex Nyl.) Hale, Phytologia 28: 339 (1974)
Parmelia tinctorum Despr. ex Nyl., Flora, Jena 55: 547 (1872).
Lichen chinensis Osbeck, Dagbok öfwer en Ostindisk Resa Aren 1750, 1751, 1752: 221 (1757).
Parmotrema chinense (Osbeck) Hale & Ahti, Taxon 35: 133 (1986).
Thallus foliose, loosely attached to the substratum, membranaceous to coriaceous, up to 18 cm across. Lobes rotund to irregular, 5~15 mm wide; margins entire to crenate, eciliate. Upper surface pale grey to grey green, shiny, becoming dull towards the thallus center, somewhat longitudinally folded in the marginal region, emaculate, without soredia, isidiate. Isidia sparse to dense, laminal and eventually marginal, granular to filiform, simple to coralloid branched, thin, brown tipped or concolorous. Medulla white. Lower surface black, minutely wrinkled, smooth, shiny, with a broad, erhizinate, pale brown to dark tan marginal zone. Rhizines sparse, simple, short. Apothecia and pycnidia not seen (Fig. 1K~1N).
Chemistry
Cortex K+ (yellow), C-, KC-, P-; medulla K-, C+ (red), KC+ (red), P- TLC: atranorin, chloroatranorin, lecanoric acid (Fig. 2).
Remarks
P. tinctorum is characterized by a large, loosely adnate thallus with broad, eciliate, laminaly isidiate lobes and lecanoric acid in the medulla. This species is very similar to P. pseudotinctorum, but the latter has inflated isidia. The species that grow under high humidity and moist conditions are larger in size, with large lobes and cylindrical, coralloid isidia, whereas species that grow under dry and stressed conditions are smaller in size with narrow lobes and granular, simple isidia [13].
During the current study, the specimens previously recorded as Parmotrema saccatilobum (Taylor) Hale [21] were identified as Parmotrema tinctorum.
Ecology and distribution
In South Korea, this species has been recorded from Mt. Sonun, Mt. Jogae, Mt. Halla, Mt. Cheontae, Mt. Jakseong, Mt. Nogudwit, and the Daeki Valley area. This is a cosmopolitan species [12] widely distributed throughout tropical, subtropical and temperate regions [13].
Selected specimens examined
Mt. Sonun, Gochang-gun, on bark, 35°29'40.4" N, 126°35'01.5" E, alt. 37 m, 20 Feb 2004, J. S. Hur, 040056-1; Mt. Jogae, on bark, 34°59'27.9" N, 127°20'01.8" E, alt. 210 m, 31 Jan 2004, J. S. Hur, 040008; Mt. Halla, Jeju Island, on rock, 33°30'17.8" N, 126°54'47.5" E, alt. 5 m, 17 Oct 2006, J. S. Hur, 061016; Mt. Cheontae, Deokhak-ri, Congju-si, Chungcheongnam-do, on rock, 36°09'24.4" N, 127°36'28.0" E, alt. 484 m, 3 Nov 2006, J. S. Hur, 061183; Mt. Jakseong, Seongnue-ri, Jechen-si, Chuncheongbuk-do, on rock, 37°01'44.5" N, 128°12'35.6" E, alt. 560 m, 28 Oct 2006, J. S. Hur, 0561115; Daeki Valley, Nanhae-gun, Gyengshangnam-do, on rock, 34°45'38.6" N, 128°02'54.0" E, alt. 262 m, 11 Nov 2007, J. S. Hur, 070972; Mt. Nogudwit, Namhae-gun, Gyeongsangnam-do, on bark, 34°45'38.6" N, 128°02'54.4" E, alt. 289 m, 11 Nov 2007, 070992 (Fig. 3).
Species excluded from this study
According to the literature, lichen species Parmotrema arnoldii (Du Rietz) Hale, Parmotrema crinitum (Ach.) M. Choisy, Parmotrema mellissii (C. W. Dodge) Hale and Parmotrema eciliatum (Nyl.) Hale have been reported from South Korea. However, the specimens of these species could not be traced; therefore, the description is based on previous literature.
Parmotrema arnoldii (Du Rietz) Hale, Phytologia 28: 335 (1974)
Parmelia arnoldii Du Rietz, Nytt Mag. Natur. 62: 80 (1924).
Imbricaria nilgherrensis Arnold, Verh. Zool. Bot. Ges. Wien 25: 472 (1875).
Parmelia nilgherrensis Nyl., Flora, Jena 57: 318 (1874).
Parmelia subarnoldii Abbayes, Mém. Inst. Sci. Madagascar, Sér. B 10: 113 (1961).
Parmotrema subarnoldii (Abbayes) Hale, Phytologia 28: 339 (1974).
P. arnoldii is characterized by the presence of a slightly maculate upper surface, coarsely sorediate margins of the lobes, revolute sorediate tips with irregular eruptions, and atranorin, alectorinic acid, and α-collatolic acid in the medulla [16, 18, 22]. This species is very closely related to P. mellisii, which is a panatropical species with both soredia and isidial initials. However, only soredia are present in P. arnoldii [16].
P. arnoldii is a temperate species [23] that has been recorded from several countries in Europe.
Parmotrema crinitum (Ach.) M. Choisy [as 'crinita'], Bull. Mens. Soc. Linn. Lyon 21: 175 (1952)
Parmelia crinita Ach., Syn. Meth. Lich. (Lund): 196 (1814).
Imbricaria crinita (Ach.) Arnold, Flora 67: 159 (1884).
Imbricaria proboscidea (Taylor) Jatta, Nuov. Giorn. Botan. Ital. 22: 50 (1890).
Parmelia proboscidea Taylor, in Mackay, Fl. Hibern. 2: 143 (1836).
P. crinitum is characterized by the presence of coralloid branched, epically ciliate isidia or often eciliate isidia and stictic acid complex in the medulla. P. mellissii also shows a coralloid isidiate condition, but differs from P. crinitum in having alectoronic and α-collatolic acid [13]. According to Elix [12], P. crinitum resembles P. ochrocrinitum both morphologically and chemically, but the latter shows pigments in the medulla. The specimen of the taxon is untraceable; therefore, the description is based on Kim [24], Divakr and Upreti [13] and Elix [12].
P. crinitum is a cosmopolitan species that is widespread in temperate, tropical regions and even subboreal forests, where high humidity is available [4, 12, 17]. Many European countries have reported the presence of P. crinitum [18]. Some Asian countries such as Japan [19], China [25], and Taiwan [26] have also reported the presence of P. crinitum.
Parmotrema mellissii (C. W. Dodge) Hale, Phytologia 28: 337 (1974)
Parmelia mellissii C. W. Dodge, Ann. Mo. Bot. Gard. 46: 134 (1959).
P. mellissii is characterized by a long abundant marginal cilia, marginal sorediate isidia and the presence of alectoronic acid in the medulla. According to Divakar and Upreti [13], this species shows great variation in the isidia. Coralloid isidia are sometimes difficult to differentiate from isidia, soredia or dactyls. Further, the specimens found from the E. Himalayan region are fertile, whereas the specimens from other regions are sterile. The specimen of the taxon could not be found; therefore, the description is based on Moon [27], Divakar and Upreti [13], and Elix [12].
Outside of South Korea, this species has also been reported in Asia, Africa and Central America.
Parmotrema eciliatum (Nyl.) Hale, Phytologia 28: 336 (1974)
Parmelia crinita var. eciliata Nyl. Flora 52: 291 (1869).
Parmelia eciliata (Nyl.) Nyl., in Fedde, Repert. 30: 225 (1932).
Parmelia platycarpa Stirt., Scott. Nat. 4: 252 (1878).
Remarks
P. eciliatum is characterized by a loosely attached thallus, dichotomous rhizines and the presence of lobules and a stictic acid complex in the medulla. P. eciliatum resembles P. crinitum in its medullary chemical nature, but the latter species has isidia on the upper surface [12]. The specimen of the taxon could not be found; therefore, the description is based on Park [16] and Elix [12].
P. eciliatum has been reported from Asia, Africa and Central America.
ACKNOWLEDGEMENTS
Author UJ thanks Ms. Jung-Shin Park for providing geographical locations of some specimens. This work was supported by a grant from the Korean National Research Resource Center Programme, and the Korean Forest Service Program (KNA 2012) through the Korea National Arboretum.
References
- 1.Blanco O, Crespol A, Divakar PK, Elix JA, Lumbsch HT. Molecular phylogeny of Parmotremoid lichens (Ascomycota, Parmeliaceae) Mycologia. 2005;97:150–159. doi: 10.3852/mycologia.97.1.150. [DOI] [PubMed] [Google Scholar]
- 2.Louwhoff SH, Crisp MD. Phylogenetic analysis of Parmotrema (Parmeliaceae: lichenized Ascomycotina) Bryologist. 2000;103:541–554. [Google Scholar]
- 3.Divakar PK, Blanco O, Hawksworth DL, Crespo A. Molecular phylogenetic studies on the Parmotrema reticulatum (syn. Rimelia reticulata) complex, including the confirmation of P. pseudoreticulatum as a distinct species. Lichenologist. 2005;37:55–65. [Google Scholar]
- 4.Kurokawa S, Lai MJ. Parmelioid lichen genera and species in Taiwan. Mycotaxon. 2001;77:225–284. [Google Scholar]
- 5.Kirk PM, Cannon PF, Minter DW, Stalpers JA. Dictionary of fungi. 10th ed. Wallingford: CABI Publishers; 2008. [Google Scholar]
- 6.Crespo A, Kauff F, Divakar PK, del Prado R, Pérez-Ortega S, de Paz GA, Ferencova Z, Blanco O, Roca-Valiente B, Núñez-Zapata J, et al. Phylogenetic generic classification of parmelioid lichens (Parmeliaceae, Ascomycota) based on molecular, morphological and chemical evidence. Taxon. 2010;59:1735–1753. [Google Scholar]
- 7.Hur JS, Koh YJ, Harada H. A checklist of Korean lichens. Lichenology. 2005;4:65–95. [Google Scholar]
- 8.Hur JS, Harada H, Oh SO, Lim KM, Kang ES, Lee SM, Kahng HY, Kim HW, Jung JS, Koh YJ. Distribution of lichen flora on South Korea. J Microbiol. 2004;42:163–167. [PubMed] [Google Scholar]
- 9.Hale ME. A monograph of Parmelia subgenus Amphigymnia. Contrib U S Natl Herb. 1965;36:193–358. [Google Scholar]
- 10.Krog H, Swinscow TD. Parmelia subgenus Amphigymnia (lichens) in East Africa. British Museum (Natural History) Botany series. 1981;9:143–231. [Google Scholar]
- 11.Elix JA. Progress in the generic delimitation of Parmelia sensu lato, lichens (Ascomycotina: Parmeliaceae) and a synoptic key to Parmeliaceae. Bryologist. 1993;96:359–383. [Google Scholar]
- 12.Elix JA. Flora of Australia. Vol. 55. Lichens: Lecanorales. 2. Parmeliaceae. Canberra: Australian Biological Resources Study; 1994. [Google Scholar]
- 13.Divakar PK, Upreti DK. Parmelinoid lichens in India (a revisionary study) Dehradun: Bishen Singh Mahendra Pal Singh, India; 2005. pp. 295–380. [Google Scholar]
- 14.Orange A, James PW, White FJ. Microchemical methods for the identification of lichens. 2nd ed. London: British Lichen Society; 2010. [Google Scholar]
- 15.López-Figueiras M. Censo de macrolíquenes venezolanos de los estados Falcón, Lara, Mérida, Táchira y Trujillo. Merida: Faculty of Pharmacy, Los Andes University; 1986. [Google Scholar]
- 16.Park YS. The macrolichen flora of South Korea. Bryologist. 1990;93:105–160. [Google Scholar]
- 17.Louwhoff SH. Parmotrema A. Massal. (1860) In: Smith CW, Aptroot A, Coppins BJ, Fletcher A, Gilbert OL, James PW, Wolseley PA, editors. The lichens of Great Britain and Ireland. London: Natural History Museum Publications; 2009. pp. 661–663. [Google Scholar]
- 18.Jabłońska A, Oset M, Kukwa M. The lichen family Parmeliaceae in Poland. I. The genus Parmotrema. Acta Mycol. 2009;44:211–222. [Google Scholar]
- 19.Yoshimura I. Lichen flora of Japan in colour. Osaka: Hoikusha; 1974. (in Japanese) [Google Scholar]
- 20.Wolseley PA, Aguirre-Hudson B, McCarthy PM. Catalogue of the lichens of Thailand. Bull Nat Hist Mus Lond (Bot) 2002;32:13–59. [Google Scholar]
- 21.Joshi Y, Wang XY, Lee YM, Byun BK, Koh YJ, Hur JS. Notes on some new records of macro- and micro-lichens from Korea. Mycobiology. 2009;37:197–202. doi: 10.4489/MYCO.2009.37.3.197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sipman H. Mason Hale's key to Parmotrema, revised edition: key to wide-lobed parmelioid species occurring in Tropical America (genera Canomaculina, Parmotrema, Rimelia, Rimeliella) [Internet] Berlin: Botanischer Garten und Botanisches Museum; 2005. [cited 2012 Sep 22]. Available from: http://www.bgbm.org/sipman/keys/Neoparmo.htm. [Google Scholar]
- 23.Jabłońska A, Oset M, Kukwa M. The lichen family Parmeliaceae in Poland. I. The genus Parmotrema. Acta Mycol. 2009;44:211–222. [Google Scholar]
- 24.Kim S. Floral studies on the lichens in Korea. Bull Kongju Teachers Coll. 1981;17:279–305. [Google Scholar]
- 25.Wei JC. An enumeration of lichens in China. Beijing International Academic Publishers; 1991. [Google Scholar]
- 26.Wang-Yang JR, Lai MJ. Additions and corrections to the lichen flora of Taiwan. Taiwania. 1976;21:226–228. [Google Scholar]
- 27.Moon KH. Lichens of Mt. Sorak in Korea. J Hattori Bot Lab. 1999;(86):187–220. [Google Scholar]