

Abstract
Rationale
Chronic dysregulation of hypothalamus-pituitary-adrenal (HPA) axis activity is related to several neuropsychiatric disorders. Studies suggest that cortisol response to stress has a strong genetic etiology, and that FK506 binding protein 5 (FKBP5) and G-protein coupled type-I CRH receptor (CRHR1) are key proteins regulating response. Variations in the genes encoding these proteins, FKBP5 and CRHR1, have been associated with several neuropsychiatric disorders.
Objectives
We examined variation in these genes in relation to cortisol response to psychological stress in one of the largest Trier Social Stress Test (TSST) cohorts yet examined.
Methods
Three hundred sixty-eight healthy, young adults underwent the TSST. Salivary cortisol was measured at multiple time points before and after the stressor. Nine variants in FKBP5 and four in CRHR1 were assessed. Single marker analyses were conducted. Secondary analyses assessed haplotypes and interaction with stress-related variables.
Results
The strongest association was for rs4713902 in FKBP5 with baseline cortisol (pdom=0.0004). We also identified a male-specific effect of FKBP5 polymorphisms on peak response and response area under the curve (p=0.0028 for rs3800374). In CRHR1, rs7209436, rs110402, and rs242924 were nominally associated with peak response (prec=0.0029–0.0047). We observed interactions between trait anxiety and rs7209436 and rs110402 in CRHR1 in association with baseline cortisol (pLRT=0.0272 and pLRT=0.0483, respectively).
Conclusions
We show association of variants in FKBP5 and CRHR1 with cortisol response to psychosocial stress. These variants were previously shown to be associated with neuropsychiatric disorders. These findings have implications for interindividual variation in HPA axis activity and potentially for the etiology of neuropsychiatric disorders.
Keywords: Human, HPA axis, cortisol, psychosocial stress, genetics, FKBP5, CRHR1
INTRODUCTION
Following the perception of stress, the hypothalamus-pituitary-adrenal (HPA) axis is activated. Several important neurotransmitter systems signal the hypothalamic corticotropin releasing hormone (CRH) neuron to activate the glucocorticoid component of the stress response. CRH stimulates pituitary adrenocorticotropic hormone (ACTH) secretion, which in turn stimulates cortisol release from the adrenal glands. Cortisol participates in its own production through sensitive negative feedback loops which are in part governed by binding to glucocorticoid receptors (GR) and FK506 binding protein 5 (FKBP5). There is marked interindividual variability in the cortisol response to stress.
Both enhanced and attenuated HPA axis responses are maladaptive. Dysregulation of the cortisol response to stress may be a risk factor for psychiatric illness (Holsboer, 2000; Tsigos and Chrousos, 2002). Depression is associated with an elevated cortisol response (Chopra et al, 2009; Heim et al, 2000), and blunted cortisol responses to stress have been observed in panic disorder and PTSD (McFarlane et al, 2011; Petrowski et al, 2010). Because chronic dysregulation of HPA axis activity is related to the onset and course of several stress-related disorders (McEwen, 1998), the identification of relevant sources of interindividual variability is an important goal in psychoendocrinological research.
Both genetic (Federenko et al, 2004) and environmental determinants, individually and through interaction, have been shown to influence an individual’s cortisol response to stress (Foley and Kirschbaum, 2010; Kudielka and Wüst, 2010). Estimates of the heritability of cortisol response to certain forms of stress are 0.62 (Bartels et al, 2003), increasing up to 0.98 with repeated administration of the Trier Social Stress Test (TSST) (Federenko et al, 2004). Several HPA axis candidate genes have been studied for association with cortisol response to psychosocial stress, including NR3C1 (van West et al, 2010; Wüst et al, 2004), NR3C2 (DeRijk et al, 2006), GABRA6 (Uhart et al, 2004), OPRM1 (Chong et al, 2006), SLC6A4 (Mueller et al, 2011; Wüst et al, 2009), and BDNF (Shalev et al, 2009). Here we focus on two genes that bookend the HPA axis, FKPB5 and CRHR1.
The CRHR1 gene encodes the G-protein coupled type I CRH receptor (CRHR1), a protein that is essential for the activation of signal transduction pathways that activate mesolimbic and HPA axis responses to many types of stress. The CRHR1 gene was selected for study for several reasons. CRH is one of the key stress factors in the CNS. The CRHR1 receptor is crucial for establishing the initial HPA axis response to stressful events. Variation in the CRHR1 gene has been shown to moderate the relationship between childhood maltreatment and cortisol response to the Dex/CRH test (Tyrka et al, 2009). CRHR1 polymorphisms have been linked to major depression in interaction with childhood maltreatment or adverse experiences (Bradley et al, 2008; Grabe et al, 2010; Kranzler et al, 2011; Polanczyk et al, 2009). An interaction between CRHR1 and stressful life events has also been shown to influence alcohol use (Blomeyer et al, 2008; Schmid et al, 2010).
Whereas CRHR1 helps establish the initial HPA axis response to a stressful episode, the FKBP5 protein assists in terminating the stress response. FKBP5 is a co-chaperone of the GR heterocomplex, part of the negative feedback loop regulating GR activity. When cortisol binds to the GR, FKBP5 is replaced by a positive regulator of GR signaling, FKBP4, and the complex is shuttled into the nucleus for transcriptional regulatory activity (Riggs et al, 2003; Wochnik et al, 2005). Overexpression of FKBP5 reduces nuclear translocation of the GR complex by sequestering it in the cytosol (Wochnik et al, 2005). Interestingly, glucocorticoid exposure increases expression of FKBP5, dampening glucocorticoid negative feedback and allowing cortisol to participate in its own regulation (Lee et al, 2011).
Like CRHR1, the FKBP5 gene was selected for study based on several lines of evidence. The FKBP5 protein plays a crucial role in determining sensitivity to glucocorticoid negative feedback, a key mechanism for terminating the HPA axis response to a stressful episode. Alterations in the expression or function of FKBP5 could increase cortisol burden and contribute to the allostatic shift in cortisol regulation that occurs with repeated exposure to stress (Lee et al, 2011). Indeed, multiple polymorphisms in the FKBP5 gene have been shown to be associated with cortisol response to the TSST (Ising et al, 2008). Variations in the gene have also been associated with mood disorder (Lavebratt et al, 2010; Lekman et al, 2008; Velders et al, 2011; Willour et al, 2009; Zobel et al, 2010), suicidality (Brent et al, 2010; Perroud et al, 2011) and response to antidepressant treatment (Binder et al, 2004; Lekman et al, 2008). Variation in FKBP5 has also been shown to interact with child abuse or trauma to predict adult depression (Appel et al, 2011), suicidality (Roy et al, 2010) and PTSD (Binder et al, 2008; Xie et al, 2010).
Since examining genetic variation in HPA axis stress response may shed light on the mechanisms underlying dysregulation of the stress response, the goal of this study was to assess whether variants in the HPA axis candidate genes FKBP5 and CRHR1 are differentially associated with cortisol responses to psychosocial stress in a large sample of healthy adults. We studied healthy young adults in order to minimize the influences of significant psychiatric burden on the stress response and highlight any potential genetic effects. We selected SNPs in both genes primarily based on their previous associations with neuropsychiatric disorders.
METHODS AND MATERIALS
We recruited healthy adults, aged 18–30 years, by newspaper advertisements and posted flyers. Participants were told they were being recruited for a stress response study. Screening was done by telephone and then in person at Johns Hopkins University School of Medicine (JHU) or University of Maryland (UMD). Participants gave written informed consent after complete description of the study. The study was approved by the Institutional Review Boards at both JHU and UMD. Participant assessments included a medical history, physical examination, blood chemistry profile, complete blood count, alcohol breathalyzer test, and urine toxicology screen. DSM-IV axis I psychiatric diagnoses were determined by a Master’s level interviewer administering The Semi-Structured Assessment for the Genetics of Alcoholism (Bucholz et al, 1994). Exclusion criteria included: (a) current medical conditions and/or use of prescription medications, (b) diagnosis of DSM-IV axis I disorder, including alcohol/drug dependence or abuse, (c) use of any psychoactive medication within the past 30 days, (d) treatment in the last six months with antidepressants, neuroleptics, sedative hypnotics, glucocorticoids, appetite suppressants, estrogens, opiate, or dopamine medications, (e) seizure disorder or history of closed head trauma, (f) a report of drinking more than 30 drinks per month in women or more than 60 drinks per month in men, (g) inability to provide clean urine drug screens at intake or during study participation, (h) nicotine dependence measured by the Fagerstom Nicotine Dependence Test and (i) pregnancy or hormonal methods of birth control in females. Female subjects were tested in the follicular phase of their menstrual cycle determined by diary and confirmed with serum progesterone level (<2ng/mL).
Trier Social Stress Test (TSST)
After overnight fasting, participants reported to the study room at 1000h for a modified version of the TSST (Kirschbaum et al, 1993) and were instructed to refrain from use of any alcohol, illicit drugs, or over the counter medications for 48h prior. A toxicology screen was performed and participants were disqualified if positive. Participants consumed a calorie-controlled breakfast between 1000–1015h and sat in a quiet room until testing began. A pressure cuff was used to record vital signs. Baseline heart rate, blood pressure and saliva samples were obtained at 1200, 1215, and 1230h. In addition to saliva collection at the JHU site, an intravenous catheter was placed in a forearm vein at 1000h and baseline blood samples were obtained at 1200, 1215 and 1230h. Quality control between sites was performed routinely. Participants then listened to audio-taped instructions of the performance task. The instructions indicated that the participant would be taking on the role of a job applicant for the position of hospital administrator and that they should convince a panel of interviewers they were the best candidate for the job in a 5-min speech. Instructions also stated they would be asked to perform a 5-min oral mental arithmetic challenge judged on speed and accuracy. They were given 10-min to mentally prepare for the task, and then escorted to another room. Participants were instructed to stand at one end of a long table with two interviewers sitting at the other end. One interviewer asked the participant to describe his/her qualifications for the job, while the other operated a video camera. Participants were expected to utilize the entire five minutes for the speech and were prompted as needed by the interviewers. For the mental arithmetic task, participants were told to repeat a four-digit number after the interviewer, subtract 13 from it, and speak their answer aloud. The participant was asked to start again if they made a mistake. After the tasks, subjects were escorted back to the study room and asked to sit quietly. Blood pressure and heart rate were recorded immediately after the arithmetic task at 1300h and again at 1315, 1330, 1355, 1410, 1425, 1440, and 1455h. Saliva and blood (only JHU site) were obtained at the same time intervals. Each participant underwent a baseline passive session 1–4 weeks before the active TSST, which was identical to the active session except for the absence of the administration of the stress procedure. This was done to minimize the effects of environmental novelty and maximize the genetic effects of the TSST on the cortisol response (Federenko et al, 2004).
Psychometric Instruments
Subjects completed self-report measures of personality traits, depression, and anxiety approximately one week before undergoing the TSST. These measures included the Revised NEO Personality Inventory (NEO-PI-R; Costa and McCrae, 1992), Beck Depression Inventory II (BDI-II; Beck et al, 1996), State-Trait Anxiety Inventory (STAI; Spielberger et al, 1983), Life Events Scale (LES; (Holmes and Rahe, 1967) and Perceived Stress Scale (PSS; Cohen et al, 1983).
Hormone Assays
Salivettes were used to collect saliva for cortisol measurement. Salivary cortisol measurements, as well as plasma concentrations of progesterone in female participants on the day of TSST, were performed in our laboratory by RIA (Diagnostics Product Co., Los Angeles, CA) using a model 1470 –counter (PerkinElmer, Shelton, CT). The inter- and intraassay coefficients of variation for all assays are less than 10%.
SNPs and Genotyping
A total of 368 individuals were genotyped at two candidate genes in the HPA axis pathway, CRHR1 and FKBP5 (see Table 1 for details). We selected SNPs for genotyping based on previous literature. We selected nine SNPs in FKBP5 for genotyping. Seven of the nine SNPs are tag SNPs (rs3800374, rs7757037, rs3798346, rs9296158, rs9380525, rs4713902, and rs6912833; Willour et al, 2009) and the other two were previously identified as being associated with response to antidepressant treatment (rs3800373 and rs1360780; Binder et al 2004). Five SNPs in CRHR1 were selected for genotyping. Three of these SNPs (rs7209436, rs110402, rs242924) were previously identified as interacting with child abuse to influence adult depression (Bradley et al, 2008). The remaining two SNPs were previously identified as interacting with stressful life events to influence alcohol use and initiation (rs242938, rs1876831 (Blomeyer et al, 2008; Schmid et al, 2010)). We examined the genotype data for deviations from Hardy-Weinberg equilibrium (HWE) and based on those results removed rs1876831 in CRHR1 from the analysis. The average missing data rate across the experiment was 0.1%.
Table 1.
Genotyped SNPs and summary measures
| Gene | SNP | BP location | Allelesa | MAF | HWE p-value | Call rate | Genotypes | ||
|---|---|---|---|---|---|---|---|---|---|
| FKBP5 | rs3800374 | 35537405 | T/C | 0.19 | 0.235 | 0.995 | 17 | 106 | 244 |
| FKBP5 | rs7757037 | 35548235 | A/G | 0.46 | 0.402 | 1.00 | 83 | 175 | 111 |
| FKBP5 | rs3798346 | 35562639 | G/A | 0.21 | 0.122 | 0.997 | 22 | 114 | 232 |
| FKBP5 | rs9296158 | 35567081 | A/G | 0.32 | 0.905 | 0.997 | 38 | 159 | 171 |
| FKBP5 | rs1360780 | 35607570 | T/C | 0.30 | 0.805 | 0.997 | 35 | 153 | 180 |
| FKBP5 | rs4713902 | 35614025 | C/T | 0.25 | 0.162 | 0.997 | 28 | 127 | 213 |
| FKBP5 | rs6912833 | 35617584 | A/T | 0.27 | 0.597 | 1.00 | 29 | 141 | 199 |
| FKBP5 | rs9380525 | 35633037 | G/C | 0.34 | 0.816 | 1.00 | 43 | 163 | 163 |
| FKBP5 | rs3800373 | 35650204 | C/A | 0.30 | 0.901 | 0.997 | 33 | 153 | 182 |
| CRHR1 | rs7209436 | 43870141 | T/C | 0.46 | 0.209 | 1.00 | 83 | 171 | 115 |
| CRHR1 | rs110402 | 43880046 | A/G | 0.47 | 0.145 | 1.00 | 89 | 170 | 110 |
| CRHR1 | rs242924 | 43885366 | T/G | 0.47 | 0.532 | 1.00 | 86 | 178 | 105 |
| CRHR1 | rs242938 | 43895935 | A/G | 0.06 | 0.635 | 1.00 | 2 | 41 | 326 |
| CRHR1 | rs1876831 | 43907744 | T/C | 0.18 | 3.53×10−07 | 1.00 | 0 | 135 | 234 |
Minor allele/major allele
Additionally, we selected 96 ancestry informative markers (AIMs) for genotyping. Four of these AIMs were unable to be called and one additional AIM was out of HWE (p=6.96×10−7), resulting in a panel of 91 AIMs available for ancestry analysis. All genotyping was conducted using the Illumina GoldenGate platform.
Statistical Analysis
Cortisol Curve Components
We assessed three components of the cortisol response curve, baseline, peak and response area under the curve (AUC). Mean baseline cortisol was calculated by averaging the three levels obtained before the performance tasks were begun. Peak cortisol was defined as the highest level reached after completion of the performance tasks. Task AUC was calculated by trapezoidal approximation from the last sample obtained prior to the speech to the end of the session. Baseline AUC was calculated using the first three cortisol levels obtained before the performance tasks were begun. Response AUC was calculated by subtracting the baseline AUC from the task AUC. Cortisol values were log-transformed for analysis. Raw values are displayed in figures.
Ancestry-Informative Marker Analysis
Ancestry analysis using the 91 genotyped AIMs was conducted in STRUCTURE 2.3 (Pritchard et al, 2000). Under an admixture model with correlated allele frequencies, 10,000 burnin length and 10,000 steps, we identified 4 ancestry clusters. We then assigned each individual to the cluster for which they had the highest estimated membership. We will henceforth refer to this coding as the population ancestry.
Single SNP Analysis
For our primary analysis, we conducted single-locus association with each of the three cortisol curve components under an additive genotype model in PLINK 1.07 (Purcell et al, 2007; http://pngu.mgh.harvard.edu/purcell/plink/). The major allele was the reference allele in all analyses. Linear regression models were run and each model was adjusted for site, sex and dummy coded indicators for population ancestry. In addition, analyses for peak cortisol included adjustment for baseline cortisol. Age and education did not contribute significantly and were not included in the final model. We adjusted the p-values for testing eight effectively independent SNPs, the number of which we estimated using the program SNPSpD (Nyholt, 2004). As baseline, peak and AUC cortisol are all highly correlated (pairwise r=0.41–0.91, all p<0.001), we did not adjust for the three curve components tested. Post hoc tests were used to identify the underlying genetic model from among dominant, recessive or additive. All p-values reported below are nominal unless otherwise specified as adjusted for multiple testing.
Haplotype Analysis
In secondary analyses, we tested whether haplotypes in FKBP5 and CRHR1 were associated with cortisol response components in PLINK 1.07. Haplotypes were defined using 2,3 and 4 SNP sliding windows across each gene. Tests of haplotype-based association were conducted using linear regression with models including terms to adjust for site, sex and dummy coded indicators for population ancestry. In addition, analyses for peak cortisol included adjustment for baseline cortisol. We conducted an omnibus test, jointly estimating and testing all haplotype effects at that position.
Interaction Effects
In other secondary analyses, we tested for differences in the associations between genotype and cortisol response by sex. We also evaluated the potential moderating effects on these associations of stress-related variables captured by scores on the BDI-II, STAI, PSS or LES. No main effect on cortisol response was observed for any of these instruments (results not shown). Moderating effects were tested by including interaction terms between genotype and the covariate of interest in the regression models. Likelihood ratio tests (LRTs) were used to test whether including the interaction term provided a better fit to the data than a model without the interaction term. These analyses were conducted in STATA 11.0 (StataCorp, 2009). All analyses were adjusted for the covariates of interest listed above.
Power
Assuming a minor allele frequency of 0.2, an additive genotype model and a p-value corrected for 8 effectively independent tests, our sample had 80% power to detect an effect size of 0.19 for baseline cortisol. The detectable effect size for peak cortisol was 0.2 and for response AUC was 0.26.
RESULTS
Sample characteristics
A total of 368 subjects were included in this study, 193 from JHU and 175 from UMD (see Supplementary text and Table S1 for characteristics by site). Subjects were between 18 and 30 years of age (mean age=23) and predominantly Caucasian (82%). The UMD site recruited only female participants and thus the combined sample is predominantly female (79%). Males had greater cortisol response than females on average (males mean peak=0.51, females mean peak=0.43, t366=−3.28, p=0.007). Scores on the depression and anxiety measures indicated that on average the subjects were not depressed or anxious (BDI mean=2.8, range=0–17; STAI trait anxiety mean=31.4, range=20–64).
Genetic Association
FKBP5
The single-SNP association results under an additive model can be seen in Table 2. The strongest evidence was for an association between rs4713902 in FKBP5 and baseline cortisol that was significant even after accounting for multiple testing (p=0.001; adjusted p=0.008). Further examination suggested a dominant model with carriers of the CC or CT genotypes having higher baseline cortisol levels on average than TT (common) homozygotes (β=0.20, p=0.0004; see Figure 1). Another SNP, rs7757037, was also associated with baseline cortisol, but only marginally so after correcting for multiple testing (p=0.008; adjusted p=0.064). Post hoc analysis suggested a recessive model with individuals carrying the rarer genotype having higher baseline cortisol levels compared to those with either heterozygous or common homozygous genotypes (β=0.21, p=0.0019). These results were consistent across sites (see Supplementary Table S2) and within the Caucasian portion of the sample (results not shown).
Table 2.
Associations between cortisol response to the TSST and polymorphisms in FKBP5 and CRHR1 under an additive genotype model
| Baseline | Peak | AUC | |||||
|---|---|---|---|---|---|---|---|
| Gene | SNP | B | pa | β | pa | β | pa |
| FKBP5 | rs3800374 | −0.05 | 0.326 | −0.05 | 0.288 | −0.06 | 0.440 |
| FKBP5 | rs7757037 | 0.11 | 0.008 | 0.01 | 0.811 | 0.01 | 0.922 |
| FKBP5 | rs3798346 | −0.07 | 0.133 | −0.01 | 0.863 | 0.02 | 0.795 |
| FKBP5 | rs9296158 | −0.07 | 0.105 | −0.04 | 0.369 | −0.06 | 0.351 |
| FKBP5 | rs1360780 | −0.07 | 0.112 | −0.03 | 0.452 | −0.04 | 0.492 |
| FKBP5 | rs4713902 | 0.15 | 0.001 | 0.002 | 0.973 | −0.001 | 0.985 |
| FKBP5 | rs6912833 | −0.08 | 0.064 | 0.01 | 0.905 | −0.02 | 0.777 |
| FKBP5 | rs9380525 | −0.07 | 0.088 | −0.0002 | 0.995 | −0.01 | 0.899 |
| FKBP5 | rs3800373 | −0.06 | 0.150 | −0.04 | 0.352 | −0.07 | 0.301 |
| CRHR1 | rs7209436 | −0.03 | 0.478 | −0.10 | 0.014 | −0.13 | 0.026 |
| CRHR1 | rs110402 | −0.01 | 0.875 | −0.09 | 0.018 | −0.11 | 0.066 |
| CRHR1 | rs242924 | −0.02 | 0.588 | −0.10 | 0.011 | −0.12 | 0.044 |
| CRHR1 | rs242938 | −0.08 | 0.369 | 0.03 | 0.749 | −0.04 | 0.851 |
Nominal p-values shown, bold indicates p-value significant after adjusting for multiple testing. Results from a linear regression model adjusted for site, sex and dummy coded indicators for population ancestry.
Figure 1.
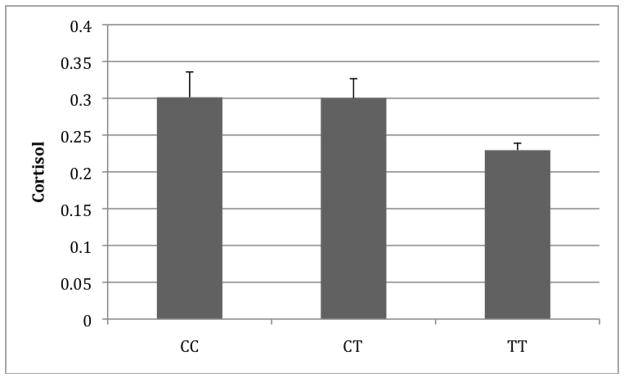
Baseline cortisol by FKBP5 rs4713902 genotype. Shown are the unadjusted mean baseline cortisol levels (μg/dl).
In the haplotype analysis, the most significant association was between baseline cortisol and the 2 SNP haplotype rs4713902-rs6912833 (p=0.0036). Examination of specific haplotypes suggested the CT haplotype (frequency 24.7%) versus all others was positively associated with baseline cortisol (p=0.001). This haplotype contained the C allele of rs4713902 and did not appear to add any information over the single-SNP analysis. No FKBP5 haplotypes were associated with peak cortisol or response AUC under the omnibus test.
We identified a sex-specific effect for three SNPs in FKBP5 associated with peak response and response AUC (rs3800374, rs7757037 and rs3800373; LRT p=0.001–0.041). Specifically, there were significant associations between FKBP5 genotypes and peak cortisol response in males (pmin=0.0028 for rs3800374), but not females (pmin=0.3498; See Figure 2). Similarly, males displayed significant associations between FKBP5 genotypes and response AUC (pmin=0.0429 for rs3800374), whereas females did not (pmin=0.4157). There was no evidence of a sex-specific effect for rs4713902, the SNP with the most significant overall association with baseline cortisol. None of the stress-related variables we tested (BDI, STAI, PSS and LES) were significant effect modifiers of the association between variants in FKBP5 and cortisol response (results not shown).
Figure 2.

Cortisol response over time by genotype for rs3800374 in FKBP5, by sex. Shown are unadjusted cortisol curves (μg/dl) for a) females and b) males.
CRHR1
Three SNPs in CRHR1 (rs7209436, rs110402 and rs242924) were associated with peak cortisol response to the TSST under an additive model (p=0.011–0.018). The association with rs242924 was marginally significant after accounting for multiple testing (adjusted p=0.088; see Figure 3). Post hoc testing suggested that the underlying genetic model for all three SNPs was recessive (p=0.0029–0.0047). For each SNP, individuals with the rare homozygous genotype had lower peak cortisol response on average than those with either a heterozygous or common homozygous genotype. These results were consistent across sites (see Supplementary Table S1) and within the Caucasian portion of the sample (results not shown). Rs7209436 was also associated with response AUC, although this association was no longer significant after accounting for multiple testing (p=0.026; adjusted p=0.208).
Figure 3.
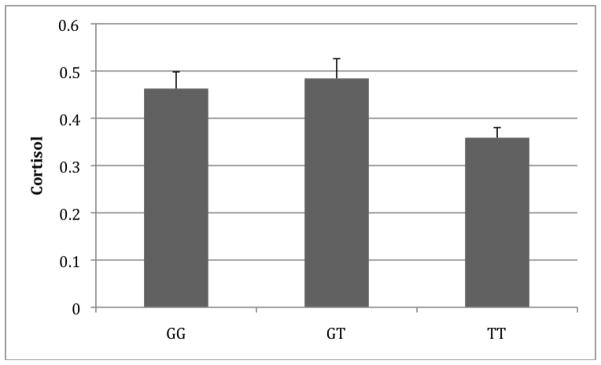
Peak cortisol response by CRHR1 rs242924 genotype. Shown are the unadjusted mean peak cortisol levels (μg/dl).
Haplotype analysis provided further evidence of associations between CRHR1 and peak cortisol and response AUC. All 2-,3-, and 4-SNP haplotypes formed by the four SNPs in CRHR1 were associated with peak cortisol by the omnibus test (p<0.05). The most significant haplotype was the 2-SNP haplotype rs110402-rs242924 (p=0.0099). The 4-SNP haplotype was also significantly associated with response AUC in the omnibus test (p=0.0433). No haplotypes were found to be associated with baseline cortisol.
There was evidence for an interaction between STAI trait anxiety and variation in CRHR1 in association with baseline cortisol (LRT p=0.0272 and p=0.0483 for rs7209436 and rs110402, respectively). In particular, higher anxiety was associated with higher baseline cortisol only among individuals with the common homozygous genotype (frequency 33%) at rs7209436 (p=0.038). This same pattern was observed for rs110402. None of the other stress-related variables (BDI, PSS and LES) or sex were significant effect modifiers of the relationship between polymorphisms in CRHR1 and cortisol response measures (results not shown).
DISCUSSION
Results of this study indicate that variation in two important HPA axis genes may influence cortisol response to psychosocial stress. The SNP associations with cortisol were consistent across study sites. The strongest evidence was for an association between FKBP5 and baseline cortisol, with homozygous or heterozygous carriers of the C allele in rs4713902 having higher baseline cortisol on average than individuals with the more common TT genotype. In addition, we noted a sex-specific effect in the association with FKBP5 such that SNPs were associated with peak response and AUC only among males. SNPs in CRHR1 were also nominally associated with peak cortisol response to the TSST, and there was evidence of an interaction between STAI trait anxiety and CRHR1 genotypes influencing baseline cortisol.
One previous study by Ising et al (2008) examined the effect of genetic variation in FKBP5 on HPA axis regulation using the TSST. That study examined three SNPs in FKBP5, rs3800373, rs1360780 or rs4713916, two of which overlapped with ours. Consistent with our study, it found significant associations with altered cortisol response to the TSST. They reported that homozygous carriers of the rare allele at each of the three SNPs tended to have higher cortisol levels during recovery after the TSST and that homozygous carriers of the rare allele at rs4713916 tended to have higher cortisol levels during the anticipation period., while we found that homozygous or heterozygous carriers of the rare allele at rs4713902 had higher baseline cortisol. We did not observe a significant association for our SNPs with peak cortisol or response AUC. In addition, the previous study did not find any differences in these associations by sex as we did. There are several possible explanations for the differences. We assessed salivary cortisol while the previous study assayed serum cortisol. While serum and salivary cortisol are highly correlated (Gozansky et al, 2005; Kirschbaum and Hellhammer, 1994), they are not perfectly so and this may have contributed to differences. Additionally, our study was over five times larger and therefore was less susceptible to findings by random chance.
Interestingly, rs4713902 in FKBP5 has previously been shown to be associated with bipolar disorder (Willour et al, 2009) and suicide attempt (Roy et al, 2010). Willour et al (2009) reported over transmission of the major allele in bipolar disorder using a family sample. The rare allele was shown to be associated with suicide attempt by Roy et al (2010). Mechanistically, the association of FKBP5 SNPs with baseline cortisol is consistent with FKBP5’s influence on glucocorticoid negative feedback. Increased expression or function of FKBP5 imparts reduced sensitivity to glucocorticoid negative feedback (Hartmann et al, 2012). This can bring about higher daily cortisol levels. This increase in cortisol burden may alter neural circuitry in a manner that predisposes to neuropsychiatric disorders.
Of the three FKBP5 SNPs that interacted with sex in our study, two have previously been reported to be associated with psychiatric disorder or treatment response (rs3800373 and rs7757037; Binder et al, 2004, 2008; Brent et al, 2010; Koenen et al, 2005; Lekman et al, 2008; Roy et al, 2010; Willour et al, 2009; Zobel et al, 2010), although these findings have not been consistent (Fallin et al, 2005; Gawlik et al, 2006; for a review see Binder, 2009). One previous study reported a male-specific association of the rare homozygous genotype in rs1360780 with depression (Lavebratt et al, 2010). However, several other studies have not observed a sex-specific effect of FKBP5 in psychiatric disorder (e.g. Binder et al, 2004, 2008; Velders et al, 2011; Willour et al, 2009). This male-specific effect of FKBP5 on measures of cortisol response (peak and AUC) may relate to the observed pattern of higher cortisol response in males than in females. This pattern of higher response in males has been reported previously (Kirschbaum et al, 1992; Uhart et al, 2006). It may be that dependent on FKBP5 genotype, males are especially vulnerable to psychosocial stress. Pre-clinically, sex hormones have been shown to affect FKBP5 expression (e.g. Hubler et al., 2003) and FKBP5 expression has been shown to be up-regulated in pregnant women and women being treated with hormone therapy (Dahm et al., 2012; Katz et al., 2012). The exact mechanisms for the observed sex-specific association of variation in FKBP5 with peak cortisol response is unclear; however FKBP5 functions as a co-chaperone for androgen and progesterone receptors in addition to glucocorticoid and mineralocorticoid receptors (Jääskeläinen et al, 2011). In addition, recent studies have shown that acute stress in adulthood has a differential sex-dependent effect on the expression of CRH and the CRHR1 receptors (Zohar & Weinstock, 2011). It is possible the sexual dimorphism in peak response is related to differential effects of male and female sex hormones on these genes and other HPA axis response genes.
In regard to the other gene examined in this study, there are some precedents for our CRHR1 SNP findings. Using a Dex/CRH test paradigm and examining variation in CRHR1, Tyrka et al (2009) found that both rs110402 and rs242924 interact with childhood maltreatment to affect HPA axis reactivity. Specifically, subjects homozygous for the common allele had higher cortisol response to the Dex/CRH test than those with heterozygous or rare homozygous genotypes, but only among those subjects reporting moderate to severe childhood maltreatment. We did not have a measure of childhood maltreatment available in our study. However, we did identify an interaction between trait anxiety and SNPs rs110402 and rs7209436 in CRHR1. Higher trait anxiety was associated with higher baseline cortisol only among individuals with the common homozygous genotype.
Variation in CRHR1 has been reported to be associated with depression (Ishitobi et al, 2012; Liu et al, 2006). In particular, the three SNP haplotype rs7209436-rs110402-rs242924 has been repeatedly shown to interact with childhood maltreatment to influence adult depression (Bradley et al, 2008; Grabe et al, 2010; Laucht et al, 2012; Polanczyk et al, 2009; Ressler et al, 2010). CRHR1 polymorphisms have also been shown to influence panic disorder (Ishitobi et al, 2012), alcohol use (Blomeyer et al, 2008; Chen et al, 2010; Schmid et al, 2010; Treutlein et al, 2006), suicide attempt (Ben-Efraim et al, 2011; Roy et al, 2012; Wasserman et al, 2008), and antidepressant treatment response (Licinio et al, 2004; Liu et al, 2007; Papiol et al, 2007).
Our study has several important strengths. First, this is one of the largest TSST studies with humans to examine the relationship between common variants in HPA axis genes and cortisol stress response to date. Second, participants in our study were carefully and extensively screened to include only healthy individuals. By focusing on this population we are able to minimize state effects (smoking, alcohol and other drug use, mood disorders, medical conditions, etc.) in favor of examining trait effects (genetics) for association with cortisol stress response. Third, in our study the cortisol response to TSST displayed typical characteristics, suggesting that the TSST was effective in activating the physiological stress response (Foley and Kirschbaum, 2010). The cortisol response to the TSST in our sample exhibited the characteristic gradual increase of cortisol level with peak concentration being reached within 15 minutes of stress cessation. As has been previously shown, cortisol response in males in our study was greater than in females (Kirschbaum et al, 1992; Uhart et al, 2006). Fourth, use of ancestry informative markers allowed us to adjust for potential population stratification. Last, women were studied only during the follicular phase of the menstrual cycle documented with serum progesterone levels drawn on the day of the TSST.
There are several issues to consider when interpreting these results. The functional influences of the variants that were found to be associated with cortisol are unknown. Also, because we selected SNPs for inclusion based on previous findings in the literature we did not have full LD coverage of the candidate genes. For FKBP5 our SNPs covered 48% of the common variation in the gene and the SNPs in CRHR1 covered only 9% (HapMap 27, CEU+TSI, MAF>0.01, r2>0.8). Our observation of a sex by genotype interaction should be interpreted taking into account the smaller proportion of males in our sample as compared to females (21% male, 79% female). Although we attempted to control for potential confounding due to ethnicity through the inclusion of the AIMs, it is possible that there remains residual confounding. Finally, we used two sites to conduct the TSST and the sites did differ in their distribution on some subject characteristics. Quality control between sites was performed routinely. Moreover, we found that SNP associations for cortisol were consistent across the two sites.
In summary, we found that differences in response to psychological stress in healthy, young adults were associated with common variants in the genes CRHR1 and FKBP5. Many of the SNPs examined in the present study have been previously associated with neuropsychiatric disorders, thus strengthening the proposed relationship between cortisol dynamics and predisposition to neuropsychiatric illnesses. Replication is needed to see if the relationship between these genes and cortisol stress response is seen in other populations. Future directions for research include a more complete assessment of HPA axis genes in relation to cortisol response to stress. Also, studies will be needed to determine the mechanism underlying these genetic influences on HPA axis activity. Identifying sources of interindividual variation in HPA axis activity has important implications for illuminating the etiology of neuropsychiatric and stress-related disorders. Genetic variation like that described above may lay the neural circuitry foundation that then interacts with early life events to determine resilience to stress later in life. Such genetic variation could impact cortisol burden over the life cycle thus influencing the risk and severity of neuropsychiatric illness associated with aberrant stress responses.
Acknowledgments
This work was supported by the grants R01MH076953, U01 AA020890, and K05AA020342 (GSW) and a gift from the Kenneth Lattman Foundation.
Footnotes
FINANCIAL DISCLOSURES
All authors report no competing interests.
Contributor Information
Pamela Belmonte Mahon, The Johns Hopkins Mood Disorders Center, Baltimore, MD, Department of Psychiatry and Behavioral Sciences, Johns Hopkins School of Medicine, Baltimore, MD.
Peter P. Zandi, Department of Mental Health, Johns Hopkins Bloomberg School of Public Health, Baltimore, MD, The Johns Hopkins Mood Disorders Center, Baltimore, MD, Department of Psychiatry and Behavioral Sciences, Johns Hopkins School of Medicine, Baltimore, MD
James B. Potash, Department of Psychiatry, University of Iowa, Iowa City, IA
Gerald Nestadt, Department of Psychiatry and Behavioral Sciences, Johns Hopkins School of Medicine, Baltimore, MD.
Gary S. Wand, Email: gwand@jhmi.edu, Department of Medicine, Johns Hopkins School of Medicine, 720 Rutland Avenue, Ross 863, Baltimore, MD 21205, USA. Tel.: 410 955 7225 Fax: 410 955 0841.
References
- Appel K, Schwahn C, Mahler J, Schulz A, Spitzer C, Fenske K, et al. Moderation of adult depression by a polymorphism in the FKBP5 gene and childhood physical abuse in the general population. Neuropsychopharmacology. 2011;36:1982–91. doi: 10.1038/npp.2011.81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartels M, Van den Berg M, Sluyter F, Boomsma DI, de Geus EJC. Heritability of cortisol levels: review and simultaneous analysis of twin studies. Psychoneuroendocrinology. 2003;28:121–37. doi: 10.1016/s0306-4530(02)00003-3. [DOI] [PubMed] [Google Scholar]
- Beck AT, Steer RA, Ball R, Ranieri W. Comparison of Beck Depression Inventories -IA and -II in psychiatric outpatients. Journal of Personality Assessment. 1996;67:588–97. doi: 10.1207/s15327752jpa6703_13. [DOI] [PubMed] [Google Scholar]
- Ben-Efraim YJ, Wasserman D, Wasserman J, Sokolowski M. Gene-environment interactions between CRHR1 variants and physical assault in suicide attempts. Genes, Brain, and Behavior. 2011;10:663–72. doi: 10.1111/j.1601-183X.2011.00703.x. [DOI] [PubMed] [Google Scholar]
- Binder EB. The role of FKBP5, a co-chaperone of the glucocorticoid receptor in the pathogenesis and therapy of affective and anxiety disorders. Psychoneuroendocrinology. 2009;34(Suppl 1):S186–95. doi: 10.1016/j.psyneuen.2009.05.021. [DOI] [PubMed] [Google Scholar]
- Binder EB, Bradley RG, Liu W, Epstein MP, Deveau TC, Mercer KB, et al. Association of FKBP5 polymorphisms and childhood abuse with risk of posttraumatic stress disorder symptoms in adults. JAMA. 2008;299:1291–305. doi: 10.1001/jama.299.11.1291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Binder EB, Salyakina D, Lichtner P, Wochnik GM, Ising M, Pütz B, et al. Polymorphisms in FKBP5 are associated with increased recurrence of depressive episodes and rapid response to antidepressant treatment. Nature Genetics. 2004;36:1319–25. doi: 10.1038/ng1479. [DOI] [PubMed] [Google Scholar]
- Blomeyer D, Treutlein J, Esser G, Schmidt MH, Schumann G, Laucht M. Interaction between CRHR1 gene and stressful life events predicts adolescent heavy alcohol use. Biological Psychiatry. 2008;63:146–51. doi: 10.1016/j.biopsych.2007.04.026. [DOI] [PubMed] [Google Scholar]
- Bradley RG, Binder EB, Epstein MP, Tang Y, Nair HP, Liu W, et al. Influence of child abuse on adult depression: moderation by the corticotropin-releasing hormone receptor gene. Archives of General Psychiatry. 2008;65:190–200. doi: 10.1001/archgenpsychiatry.2007.26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brent D, Melhem N, Ferrell R, Emslie G, Wagner KD, Ryan N, et al. Association of FKBP5 polymorphisms with suicidal events in the Treatment of Resistant Depression in Adolescents (TORDIA) study. The American Journal of Psychiatry. 2010;167:190–7. doi: 10.1176/appi.ajp.2009.09040576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bucholz KK, Cadoret R, Cloninger CR, Dinwiddie SH, Hesselbrock VM, Nurnberger JI, et al. A new, semi-structured psychiatric interview for use in genetic linkage studies: a report on the reliability of the SSAGA. Journal of Studies on Alcohol. 1994;55:149–58. doi: 10.15288/jsa.1994.55.149. [DOI] [PubMed] [Google Scholar]
- Chen ACH, Manz N, Tang Y, Rangaswamy M, Almasy L, Kuperman S, et al. Single-nucleotide polymorphisms in corticotropin releasing hormone receptor 1 gene (CRHR1) are associated with quantitative trait of event-related potential and alcohol dependence. Alcoholism, Clinical and Experimental Research. 2010;34:988–96. doi: 10.1111/j.1530-0277.2010.01173.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chong RY, Oswald L, Yang X, Uhart M, Lin P-I, Wand GS. The mu-opioid receptor polymorphism A118G predicts cortisol responses to naloxone and stress. Neuropsychopharmacology. 2006;31:204–11. doi: 10.1038/sj.npp.1300856. [DOI] [PubMed] [Google Scholar]
- Chopra KK, Ravindran A, Kennedy SH, Mackenzie B, Matthews S, Anisman H, et al. Sex differences in hormonal responses to a social stressor in chronic major depression. Psychoneuroendocrinology. 2009;34:1235–41. doi: 10.1016/j.psyneuen.2009.03.014. [DOI] [PubMed] [Google Scholar]
- Cohen S, Kamarck T, Mermelstein R. A global measure of perceived stress. Journal of Health and Social Behavior. 1983;24:385–96. [PubMed] [Google Scholar]
- Costa P, McCrae R. Revised NEO Personality Inventory (NEO-PI-R) and NEO Five-Factor Inventory (NEO-FFI) professional manual. Psychological Assessmnet Resources, Inc; Odessa, FL: 1992. [Google Scholar]
- Dahm AEA, Eilertsen AL, Goeman J, Olstad OK, Ovstebø R, Kierulf P, Mowinckel M-C, et al. A microarray study on the effect of four hormone therapy regimens on gene transcription in whole blood from healthy postmenopausal women. Thrombosis Research. 2012;130(1):45–51. doi: 10.1016/j.thromres.2011.12.009. [DOI] [PubMed] [Google Scholar]
- DeRijk RH, Wüst S, Meijer OC, Zennaro M-C, Federenko IS, Hellhammer DH, et al. A common polymorphism in the mineralocorticoid receptor modulates stress responsiveness. The Journal of Clinical Endocrinology and Metabolism. 2006;91:5083–9. doi: 10.1210/jc.2006-0915. [DOI] [PubMed] [Google Scholar]
- Fallin MD, Lasseter VK, Avramopoulos D, Nicodemus KK, Wolyniec PS, McGrath JA, et al. Bipolar I disorder and schizophrenia: a 440-single-nucleotide polymorphism screen of 64 candidate genes among Ashkenazi Jewish case-parent trios. American Journal of Human Genetics. 2005;77:918–36. doi: 10.1086/497703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Federenko IS, Nagamine M, Hellhammer DH, Wadhwa PD, Wüst S. The heritability of hypothalamus pituitary adrenal axis responses to psychosocial stress is context dependent. The Journal of Clinical Endocrinology and Metabolism. 2004;89:6244–50. doi: 10.1210/jc.2004-0981. [DOI] [PubMed] [Google Scholar]
- Foley P, Kirschbaum C. Human hypothalamus-pituitary-adrenal axis responses to acute psychosocial stress in laboratory settings. Neuroscience and Biobehavioral Reviews. 2010;35:91–6. doi: 10.1016/j.neubiorev.2010.01.010. [DOI] [PubMed] [Google Scholar]
- Gawlik M, Moller-Ehrlich K, Mende M, Jovnerovski M, Jung S, Jabs B, et al. Is FKBP5 a genetic marker of affective psychosis? A case control study and analysis of disease related traits. BMC Psychiatry. 2006;6:52. doi: 10.1186/1471-244X-6-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gozansky WS, Lynn JS, Laudenslager ML, Kohrt WM. Salivary cortisol determined by enzyme immunoassay is preferable to serum total cortisol for assessment of dynamic hypothalamic--pituitary--adrenal axis activity. Clinical Endocrinology. 2005;63:336–41. doi: 10.1111/j.1365-2265.2005.02349.x. [DOI] [PubMed] [Google Scholar]
- Grabe HJ, Schwahn C, Appel K, Mahler J, Schulz A, Spitzer C, et al. Childhood maltreatment, the corticotropin-releasing hormone receptor gene and adult depression in the general population. American Journal of Medical Genetics Part B, Neuropsychiatric Genetics. 2010;153B:1483–93. doi: 10.1002/ajmg.b.31131. [DOI] [PubMed] [Google Scholar]
- Hartmann J, Wagner KV, Liebl C, Scharf SH, Wang X-D, Wolf M, et al. The involvement of FK506-binding protein 51 (FKBP5) in the behavioral and neuroendocrine effects of chronic social defeat stress. Neuropharmacology. 2012;62:332–9. doi: 10.1016/j.neuropharm.2011.07.041. [DOI] [PubMed] [Google Scholar]
- Heim C, Newport DJ, Heit S, Graham YP, Wilcox M, Bonsall R, et al. Pituitary-adrenal and autonomic responses to stress in women after sexual and physical abuse in childhood. JAMA. 2000;284:592–7. doi: 10.1001/jama.284.5.592. [DOI] [PubMed] [Google Scholar]
- Holmes TH, Rahe RH. The Social Readjustment Rating Scale. Journal of Psychosomatic Research. 1967;11:213–8. doi: 10.1016/0022-3999(67)90010-4. [DOI] [PubMed] [Google Scholar]
- Holsboer F. The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology. 2000;23:477–501. doi: 10.1016/S0893-133X(00)00159-7. [DOI] [PubMed] [Google Scholar]
- Hubler TR, Denny WB, Valentine DL, Cheung-Flynn J, Smith DF, Scammell JG. The FK506-binding immunophilin FKBP51 is transcriptionally regulated by progestin and attenuates progestin responsiveness. Endocrinology. 2003;144(6):2380–7. doi: 10.1210/en.2003-0092. [DOI] [PubMed] [Google Scholar]
- Ishitobi Y, Nakayama S, Yamaguchi K, Kanehisa M, Higuma H, Maruyama Y, et al. Association of CRHR1 and CRHR2 with major depressive disorder and panic disorder in a Japanese population. American Journal of Medical Genetics Part B, Neuropsychiatric Genetics. 2012;159B:429–36. doi: 10.1002/ajmg.b.32046. [DOI] [PubMed] [Google Scholar]
- Ising M, Depping A-M, Siebertz A, Lucae S, Unschuld PG, Kloiber S, et al. Polymorphisms in the FKBP5 gene region modulate recovery from psychosocial stress in healthy controls. The European Journal of Neuroscience. 2008;28:389–98. doi: 10.1111/j.1460-9568.2008.06332.x. [DOI] [PubMed] [Google Scholar]
- Jääskeläinen T, Makkonen H, Palvimo JJ. Steroid up-regulation of FKBP51 and its role in hormone signaling. Current Opinion in Pharmacology. 2011;11(4):326–31. doi: 10.1016/j.coph.2011.04.006. [DOI] [PubMed] [Google Scholar]
- Katz ER, Stowe ZN, Newport DJ, Kelley ME, Pace TW, Cubells JF, Binder EB. Regulation of mRNA expression encoding chaperone and co-chaperone proteins of the glucocorticoid receptor in peripheral blood: association with depressive symptoms during pregnancy. Psychological Medicine. 2012;42(5):943–56. doi: 10.1017/S0033291711002121. [DOI] [PubMed] [Google Scholar]
- Kirschbaum C, Hellhammer DH. Salivary cortisol in psychoneuroendocrine research: recent developments and applications. Psychoneuroendocrinology. 1994;19:313–33. doi: 10.1016/0306-4530(94)90013-2. [DOI] [PubMed] [Google Scholar]
- Kirschbaum C, Pirke KM, Hellhammer DH. The “Trier Social Stress Test”--a tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology. 1993;28:76–81. doi: 10.1159/000119004. [DOI] [PubMed] [Google Scholar]
- Kirschbaum C, Wüst S, Hellhammer D. Consistent sex differences in cortisol responses to psychological stress. Psychosomatic Medicine. 1992;54:648–57. doi: 10.1097/00006842-199211000-00004. [DOI] [PubMed] [Google Scholar]
- Koenen KC, Saxe G, Purcell S, Smoller JW, Bartholomew D, Miller A, et al. Polymorphisms in FKBP5 are associated with peritraumatic dissociation in medically injured children. Molecular Psychiatry. 2005;10:1058–9. doi: 10.1038/sj.mp.4001727. [DOI] [PubMed] [Google Scholar]
- Kranzler HR, Feinn R, Nelson EC, Covault J, Anton RF, Farrer L, et al. A CRHR1 haplotype moderates the effect of adverse childhood experiences on lifetime risk of major depressive episode in African-American women. American Journal of Medical Genetics Part B, Neuropsychiatric Genetics. 2011;156B:960–8. doi: 10.1002/ajmg.b.31243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kudielka BM, Wüst S. Human models in acute and chronic stress: assessing determinants of individual hypothalamus-pituitary-adrenal axis activity and reactivity. Stress. 2010;13:1–14. doi: 10.3109/10253890902874913. [DOI] [PubMed] [Google Scholar]
- Laucht M, Treutlein J, Blomeyer D, Buchmann AF, Schmidt MH, Esser G, et al. Interactive effects of corticotropin-releasing hormone receptor 1 gene and childhood adversity on depressive symptoms in young adults: Findings from a longitudinal study. European Neuropsychopharmacology. 2012 doi: 10.1016/j.euroneuro.2012.06.002. [DOI] [PubMed] [Google Scholar]
- Lavebratt C, Aberg E, Sjöholm LK, Forsell Y. Variations in FKBP5 and BDNF genes are suggestively associated with depression in a Swedish population-based cohort. Journal of Affective Disorders. 2010;125:249–55. doi: 10.1016/j.jad.2010.02.113. [DOI] [PubMed] [Google Scholar]
- Lee RS, Tamashiro KLK, Yang X, Purcell RH, Huo Y, Rongione M, et al. A measure of glucocorticoid load provided by DNA methylation of Fkbp5 in mice. Psychopharmacology. 2011;218:303–12. doi: 10.1007/s00213-011-2307-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lekman M, Laje G, Charney D, Rush AJ, Wilson AF, Sorant AJM, et al. The FKBP5-gene in depression and treatment response--an association study in the Sequenced Treatment Alternatives to Relieve Depression (STAR*D) Cohort. Biological Psychiatry. 2008;63:1103–10. doi: 10.1016/j.biopsych.2007.10.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Licinio J, O’Kirwan F, Irizarry K, Merriman B, Thakur S, Jepson R, et al. Association of a corticotropin-releasing hormone receptor 1 haplotype and antidepressant treatment response in Mexican-Americans. Molecular Psychiatry. 2004;9:1075–82. doi: 10.1038/sj.mp.4001587. [DOI] [PubMed] [Google Scholar]
- Liu Z, Zhu F, Wang G, Xiao Z, Tang J, Liu W, et al. Association study of corticotropin-releasing hormone receptor 1 gene polymorphisms and antidepressant response in major depressive disorders. Neuroscience Letters. 2007;414:155–8. doi: 10.1016/j.neulet.2006.12.013. [DOI] [PubMed] [Google Scholar]
- Liu Z, Zhu F, Wang G, Xiao Z, Wang H, Tang J, et al. Association of corticotropin-releasing hormone receptor 1 gene SNP and haplotype with major depression. Neuroscience Letters. 2006;404:358–62. doi: 10.1016/j.neulet.2006.06.016. [DOI] [PubMed] [Google Scholar]
- McEwen BS. Stress, adaptation, and disease. Allostasis and allostatic load. Annals of the New York Academy of Sciences. 1998;840:33–44. doi: 10.1111/j.1749-6632.1998.tb09546.x. [DOI] [PubMed] [Google Scholar]
- McFarlane AC, Barton CA, Yehuda R, Wittert G. Cortisol response to acute trauma and risk of posttraumatic stress disorder. Psychoneuroendocrinology. 2011;36:720–7. doi: 10.1016/j.psyneuen.2010.10.007. [DOI] [PubMed] [Google Scholar]
- Mueller A, Armbruster D, Moser DA, Canli T, Lesch K-P, Brocke B, et al. Interaction of serotonin transporter gene-linked polymorphic region and stressful life events predicts cortisol stress response. Neuropsychopharmacology. 2011;36:1332–9. doi: 10.1038/npp.2011.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nyholt DR. A simple correction for multiple testing for single-nucleotide polymorphisms in linkage disequilibrium with each other. American Journal of Human Genetics. 2004;74:765–9. doi: 10.1086/383251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papiol S, Arias B, Gastó C, Gutiérrez B, Catalán R, Fañanás L. Genetic variability at HPA axis in major depression and clinical response to antidepressant treatment. Journal of Affective Disorders. 2007;104:83–90. doi: 10.1016/j.jad.2007.02.017. [DOI] [PubMed] [Google Scholar]
- Perroud N, Bondolfi G, Uher R, Gex-Fabry M, Aubry J-M, Bertschy G, et al. Clinical and genetic correlates of suicidal ideation during antidepressant treatment in a depressed outpatient sample. Pharmacogenomics. 2011;12:365–77. doi: 10.2217/pgs.10.189. [DOI] [PubMed] [Google Scholar]
- Petrowski K, Herold U, Joraschky P, Wittchen H-U, Kirschbaum C. A striking pattern of cortisol non-responsiveness to psychosocial stress in patients with panic disorder with concurrent normal cortisol awakening responses. Psychoneuroendocrinology. 2010;35:414–21. doi: 10.1016/j.psyneuen.2009.08.003. [DOI] [PubMed] [Google Scholar]
- Polanczyk G, Caspi A, Williams B, Price TS, Danese A, Sugden K, et al. Protective effect of CRHR1 gene variants on the development of adult depression following childhood maltreatment: replication and extension. Archives of General Psychiatry. 2009;66:978–85. doi: 10.1001/archgenpsychiatry.2009.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pritchard JK, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data. Genetics. 2000;155:945–59. doi: 10.1093/genetics/155.2.945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MAR, Bender D, et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. American Journal of Human Genetics. 2007;81:559–75. doi: 10.1086/519795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ressler KJ, Bradley B, Mercer KB, Deveau TC, Smith AK, Gillespie CF, et al. Polymorphisms in CRHR1 and the serotonin transporter loci: gene x gene x environment interactions on depressive symptoms. American Journal of Medical Genetics Part B, Neuropsychiatric Genetics. 2010;153B:812–24. doi: 10.1002/ajmg.b.31052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riggs DL, Roberts PJ, Chirillo SC, Cheung-Flynn J, Prapapanich V, Ratajczak T, et al. The Hsp90-binding peptidylprolyl isomerase FKBP52 potentiates glucocorticoid signaling in vivo. The EMBO Journal. 2003;22:1158–67. doi: 10.1093/emboj/cdg108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy A, Gorodetsky E, Yuan Q, Goldman D, Enoch M-A. Interaction of FKBP5, a stress-related gene, with childhood trauma increases the risk for attempting suicide. Neuropsychopharmacology. 2010;35:1674–83. doi: 10.1038/npp.2009.236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy A, Hodgkinson CA, Deluca V, Goldman D, Enoch M-A. Two HPA axis genes, CRHBP and FKBP5, interact with childhood trauma to increase the risk for suicidal behavior. Journal of Psychiatric Research. 2012;46:72–9. doi: 10.1016/j.jpsychires.2011.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmid B, Blomeyer D, Treutlein J, Zimmermann US, Buchmann AF, Schmidt MH, et al. Interacting effects of CRHR1 gene and stressful life events on drinking initiation and progression among 19-year-olds. The International Journal of Neuropsychopharmacology. 2010;13:703–14. doi: 10.1017/S1461145709990290. [DOI] [PubMed] [Google Scholar]
- Shalev I, Lerer E, Israel S, Uzefovsky F, Gritsenko I, Mankuta D, et al. BDNF Val66Met polymorphism is associated with HPA axis reactivity to psychological stress characterized by genotype and gender interactions. Psychoneuroendocrinology. 2009;34:382–8. doi: 10.1016/j.psyneuen.2008.09.017. [DOI] [PubMed] [Google Scholar]
- Spielberger C, Gorsuch R, Lushene R, Vagg P, Jacobs G. Manual for the State-Trait Anxiety Inventory. Consulting Psychologists Press; Palo Alto, CA: 1983. [Google Scholar]
- StataCorp. Stata Statistical Software. 2009. [Google Scholar]
- Treutlein J, Kissling C, Frank J, Wiemann S, Dong L, Depner M, et al. Genetic association of the human corticotropin releasing hormone receptor 1 (CRHR1) with binge drinking and alcohol intake patterns in two independent samples. Molecular Psychiatry. 2006;11:594–602. doi: 10.1038/sj.mp.4001813. [DOI] [PubMed] [Google Scholar]
- Tsigos C, Chrousos GP. Hypothalamic-pituitary-adrenal axis, neuroendocrine factors and stress. Journal of Psychosomatic Research. 2002;53:865–71. doi: 10.1016/s0022-3999(02)00429-4. [DOI] [PubMed] [Google Scholar]
- Tyrka AR, Price LH, Gelernter J, Schepker C, Anderson GM, Carpenter LL. Interaction of childhood maltreatment with the corticotropin-releasing hormone receptor gene: effects on hypothalamic-pituitary-adrenal axis reactivity. Biological Psychiatry. 2009;66:681–5. doi: 10.1016/j.biopsych.2009.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uhart M, Chong RY, Oswald L, Lin P-I, Wand GS. Gender differences in hypothalamic-pituitary-adrenal (HPA) axis reactivity. Psychoneuroendocrinology. 2006;31:642–52. doi: 10.1016/j.psyneuen.2006.02.003. [DOI] [PubMed] [Google Scholar]
- Uhart M, McCaul ME, Oswald LM, Choi L, Wand GS. GABRA6 gene polymorphism and an attenuated stress response. Molecular Psychiatry. 2004;9:998–1006. doi: 10.1038/sj.mp.4001535. [DOI] [PubMed] [Google Scholar]
- Velders FP, Kuningas M, Kumari M, Dekker MJ, Uitterlinden AG, Kirschbaum C, et al. Genetics of cortisol secretion and depressive symptoms: a candidate gene and genome wide association approach. Psychoneuroendocrinology. 2011;36:1053–61. doi: 10.1016/j.psyneuen.2011.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wasserman D, Sokolowski M, Rozanov V, Wasserman J. The CRHR1 gene: a marker for suicidality in depressed males exposed to low stress. Genes, Brain, and Behavior. 2008;7:14–9. doi: 10.1111/j.1601-183X.2007.00310.x. [DOI] [PubMed] [Google Scholar]
- van West D, Del-Favero J, Deboutte D, Van Broeckhoven C, Claes S. Associations between common arginine vasopressin 1b receptor and glucocorticoid receptor gene variants and HPA axis responses to psychosocial stress in a child psychiatric population. Psychiatry Research. 2010;179:64–8. doi: 10.1016/j.psychres.2009.04.002. [DOI] [PubMed] [Google Scholar]
- Willour VL, Chen H, Toolan J, Belmonte P, Cutler DJ, Goes FS, et al. Family-based association of FKBP5 in bipolar disorder. Molecular Psychiatry. 2009;14:261–8. doi: 10.1038/sj.mp.4002141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wochnik GM, Rüegg J, Abel GA, Schmidt U, Holsboer F, Rein T. FK506-binding proteins 51 and 52 differentially regulate dynein interaction and nuclear translocation of the glucocorticoid receptor in mammalian cells. The Journal of Biological Chemistry. 2005;280:4609–16. doi: 10.1074/jbc.M407498200. [DOI] [PubMed] [Google Scholar]
- Wüst S, Kumsta R, Treutlein J, Frank J, Entringer S, Schulze TG, et al. Sex-specific association between the 5-HTT gene-linked polymorphic region and basal cortisol secretion. Psychoneuroendocrinology. 2009;34:972–82. doi: 10.1016/j.psyneuen.2009.01.011. [DOI] [PubMed] [Google Scholar]
- Wüst S, Van Rossum EFC, Federenko IS, Koper JW, Kumsta R, Hellhammer DH. Common polymorphisms in the glucocorticoid receptor gene are associated with adrenocortical responses to psychosocial stress. The Journal of Clinical Endocrinology and Metabolism. 2004;89:565–73. doi: 10.1210/jc.2003-031148. [DOI] [PubMed] [Google Scholar]
- Xie P, Kranzler HR, Poling J, Stein MB, Anton RF, Farrer LA, et al. Interaction of FKBP5 with childhood adversity on risk for post-traumatic stress disorder. Neuropsychopharmacology. 2010;35:1684–92. doi: 10.1038/npp.2010.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zobel A, Schuhmacher A, Jessen F, Höfels S, von Widdern O, Metten M, et al. DNA sequence variants of the FKBP5 gene are associated with unipolar depression. The International Journal of Neuropsychopharmacology. 2010;13:649–60. doi: 10.1017/S1461145709991155. [DOI] [PubMed] [Google Scholar]
- Zohar I, Weinstock M. Differential effect of prenatal stress on the expression of corticotrophin-releasing hormone and its receptors in the hypothalamus and amygdala in male and female rats. Journal of Neuroendocrinology. 2011;23(4):320–8. doi: 10.1111/j.1365-2826.2011.02117.x. [DOI] [PubMed] [Google Scholar]