

Abstract
Dendritic cells with tolerogenic function (tolDC) have become a promising immunotherapeutic tool for reinstating immune tolerance in rheumatoid arthritis (RA) and other autoimmune diseases. The concept underpinning tolDC therapy is that it specifically targets the pathogenic autoimmune response while leaving protective immunity intact. Findings from human in-vitro and mouse in-vivo studies have been translated into the development of clinical grade tolDC for the treatment of autoimmune disorders. Recently, two tolDC trials in RA and type I diabetes have been carried out and other trials are in progress or are imminent. In this review, we provide an update on tolDC therapy, in particular in relation to the treatment of RA, and discuss the challenges and the future perspectives of this new experimental immunotherapy.
Keywords: autoimmune disease, dendritic cells, immunotherapy, rheumatoid arthritis, tolerance
OTHER ARTICLES PUBLISHED IN THIS SERIES
T cell depletion in paediatric stem cell transplantation. Clinical and Experimental Immunology 2013, 172: 139–47.
Promoting transplantation tolerance; adoptive regulatory T cell therapy. Clinical and Experimental Immunology 2013, 172: 158–68.
Whole body imaging of adoptively transferred T cells using magnetic resonance imaging, single photon emission computed tomography and positron emission tomography techniques, with a focus on regulatory T cells. Clinical and Experimental Immunology 2013, 172: 169–77.
Introduction
Rheumatoid arthritis (RA) is an autoimmune disease characterized by chronic synovial inflammation, leading to destruction of joint cartilage and bone. Although the precise aetiology remains to be established, it is thought that RA results from a breach in immune tolerance. T cell responses to several (joint-associated) autoantigens, including ‘altered self’ citrullinated peptides, can be detected in a proportion of RA patients 1–8, and the function of peripheral blood regulatory T cells (Tregs) is impaired in RA patients with active disease 9. Immunosuppressive drugs (including biological drugs) can relieve disease symptoms effectively, but none of the currently available treatments provide a cure, i.e. a long-lasting and drug-free remission of RA 10,11. Moreover, these drugs can increase the risk of serious infections 12–14. The ‘holy grail’ of the immunotherapy field is to develop a therapy that targets and rectifies the pathological autoimmune response specifically and effectively, while leaving protective immunity intact. A new immunotherapeutic approach aims to achieve restoration of immune tolerance by treatment with autologous dendritic cells (DC) with tolerogenic function [tolerogenic DC (tolDC)]. Here we review recent progress in this field.
Role of DC in maintaining immune tolerance
Destructive autoimmunity is normally prevented through active silencing of autoreactive T cells, a process in which DC play a central role. In the thymus self-reactive T cells are deleted, but this process of ‘central tolerance’ has limitations and some autoreactive T cells escape to peripheral tissues. Here they are kept under control by a variety of mechanisms, termed collectively ‘peripheral tolerance’. When tolerance mechanisms break down, autoreactive T cells can acquire proinflammatory properties [e.g. become T helper type 1 (Th1) or Th17 cells] and mount an attack on the body's own tissues, causing an autoinflammatory, destructive immune response 15. For example, a shift from a tolerogenic to a proinflammatory T cell response in RA has been reported by van Bilsen et al. 3. They detected CD4+ T cells specific for the autoantigen human cartilage gp39 (HCgp39) in both healthy individuals and RA patients. However, HCgp39-reactive T cells from healthy individuals exhibited a regulatory phenotype [interleukin (IL)-10 production, forkhead box protein 3 (FoxP3) expression, capability to suppress T cell responses], whereas HCgp39-reactive T cells from RA patients produced the proinflammatory cytokine interferon (IFN)-γ and lacked suppressive activity.
DC are professional antigen-presenting cells that, in terms of function, were defined initially by their potent ability to activate naive T cells and instigate effective T cell responses. However, it is now recognized that DC are also important for the induction and maintenance of peripheral T cell tolerance 15. For instance, mice in which both conventional and plasmacytoid DC subsets have been ablated develop severe, fatal autoimmunity 16. Notably, patients with the recently identified combined mononuclear cell deficiency DCML [DC, monocyte, B and natural killer (NK) lymphoid-deficient], virtually lacking DC in the blood and interstitial tissues, have a reduced number of Tregs, and a quarter of these patients develop autoimmune disorders 17.
The dual function of DC in initiating immunity, on one hand, and maintaining T cell tolerance on the other hand, can be explained, in part, by the different maturation stages of DC 18,19. In the absence of danger signals provided by infection or inflammation (also referred to as ‘steady state’), DC are largely in an immature differentiation state. They can capture and present antigens to T cells, but in so doing will induce tolerance rather than immunity 20–22. Maturation of DC can be induced by pathogen-associated molecular patterns (PAMP), e.g. bacterial lipopolysaccharide (LPS) or viral double-stranded RNA 23. The process of DC maturation enhances their immunogenicity by up-regulation of major histocompatibility complex (MHC)–peptide complexes and T cell co-stimulatory molecules (e.g. CD80, CD86) on the plasma membrane, and by inducing the production of proinflammatory cytokines (e.g. IL-12) that help and polarize T cell differentiation 24,25. However, the notion that immature DC induce tolerance and mature DC induce immunity has been revised in recent years, as it has become clear that mature DC can also exert pro-tolerogenic effects. For example, DC matured in response to certain PAMP display a typical mature DC surface phenotype but produce anti-inflammatory IL-10 and promote the development of IL-10-producing Tregs 26,27. It is now generally accepted that the tolerogenic function of DC is determined by the signals that they receive during maturation; these signals can be derived either from the microenvironment in which DC maturation takes place or from invading pathogens. For instance, anti-inflammatory cytokines [IL-10, transforming growth factor (TGF)-β], immunosuppressive substances (e.g. corticosteroids) or certain PAMP (e.g. schistosomal lysophosphatidylserine) have all been shown to promote the tolerogenic function of DC 27–31.
Several mechanisms by which tolDC induce immune peripheral tolerance have been described, including blocking of T cell clonal expansion and induction of T cell anergy, deletion of T cells and the induction of Tregs. Two major groups of Tregs have been defined: naturally occurring Tregs (nTregs) that arise in the thymus, and adaptive Tregs, that are induced in the periphery (iTregs) 32,33. nTregs are thought to suppress immune responses by mainly contact-dependent mechanisms, including down-regulation of CD80 and CD86 expression by CTLA4-mediated transendocytosis 34. One of the best-characterized types of iTreg is the type 1 regulatory T cell (Tr1). These cells are induced from naive T cells and control immune responses mainly through the production of immunosuppressive cytokines (IL-10 and TGF-β), but they can also act by lysing target cells of myeloid origin 35. The mechanisms by which tolDC operate have been described amply in detail by others (e.g. 18,36,37); only a few examples will be mentioned here. DC producing the tryptophan-degrading enzyme indoleamine 2,3 dioxygenase (IDO) block T cell clonal expansion 38. Plasmacytoid DC in the liver promote antigen-specific tolerance through T cell deletion and/or the induction of T cell anergy 39. Mucosal CD103+ DC induce FoxP3+ Tregs through secretion of TGF-β and/or retinoic acid 40,41, whereas mucosal CD8+ DC induce Tr1-like cells with regulatory properties 41. Interestingly, it has been shown that Tregs, in turn, suppress DC maturation and enhance the expression of immunosuppressive molecules (e.g. IL-10, B7-H4), thus inducing tolerogenic function in DC 42,43. This bidirectional cross-talk between Tregs and DC further supports immune tolerance.
Tolerogenic dendritic cells as an immunotherapeutic tool
The concept that maturation conditions determine the tolerogenicity of DC has facilitated the development of tolDC therapies for disorders that are characterized by a failure in immune tolerance. TolDC treatment for the prevention of graft rejection in transplantation has been reviewed extensively elsewhere 44,45; the current review focuses on development of this tolerogenic immunotherapy for autoimmune diseases, in particular RA. TolDC have been developed as an autologous cellular therapy, in which DC precursors are isolated from the patient, differentiated ex vivo into tolDC, loaded with appropriate autoantigens (optional), and injected back into the patient. Many different methods are available for the ex-vivo generation of DC with potent tolerogenic function. One of the most important considerations in choosing the appropriate method is that the final tolDC product should be stable, i.e. tolDC should not differentiate into immunogenic DC in vivo when exposed to proinflammatory mediators. The stability of tolDC is, therefore, an especially important consideration if they are going to be used for the treatment of autoimmune diseases that are characterized by chronic inflammation, as is the case in RA. Certain types of tolDC (e.g. partially matured DC, also referred to as semi-mature DC) have indeed been shown to become immunogenic in vivo 46,47, which is undesirable, as presentation of autoantigen by immunogenic DC can induce or exacerbate autoimmune disease 48,49.
Methods for stable tolDC generation have been reviewed elsewhere 50, and will be summarized only briefly here. One of the most popular methods is to maintain DC in an permanently immature state by inhibiting the expression or function of nuclear factor (NF)κB, a transcription factor pivotal for DC maturation 49,51–55. Another option is to engineer DC genetically to either constitutively express immunosuppressive [e.g. IL-4, IL-10, cytotoxic T lymphocyte antigen (CTLA)-4; 56–60] or apoptosis-inducing [e.g. Fas, tumour necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL); 61–63] molecules or, conversely, to inhibit expression of immunostimulatory molecules (e.g. CD80/CD86, IL-12; 64–66). Other methods of tolDC generation include treatment of DC with immunosuppressive cytokines IL-10/TGF-β 67–69 or rapamycin 70, short-term stimulation with LPS 71, induction of microRNA-23b expression 72 or increasing Wnt signalling by treatment with Wnt-5a 73. Many of these ex-vivo-generated tolDC are capable of inhibiting pathogenic autoreactive T cell responses in vivo 50.
A variety of tolDC have been tested in animal models of RA. Importantly, a number of tolDC have been shown to have immunotherapeutic potential, i.e. can suppress established arthritis 50,74). Not surprisingly, the in-vivo mechanism of action by which these tolDC exert their beneficial effects depends on the type of tolDC administered (reviewed in 74). For instance, FasL-transduced DC act by depletion of autoreactive T cells 62, IDO- or CTLA 4 immunoglobulin (Ig)-transduced DC induce FoxP3+ Tregs 75, and dexamethasone/vitamin D3-modulated DC inhibit Th17 cells and enhance IL-10-producing T cells 74.
Clinical trials with tolDC in autoimmune disease
The positive results from preclinical animal models have provided strong support for the concept that tolDC can be applied as an immunotherapeutic agent for the treatment of autoimmune diseases. However, animal models of autoimmune disease do not reflect human disease completely and ultimately the safety, feasibility and effectiveness of tolDC therapy can be tested only through clinical trials. Two tolDC trials (in type I diabetes and RA) have been conducted recently 76,77, and our tolDC trial in RA has also started recently – see section below for more detail. A tolDC trial in MS has not yet been reported, but a recent study by the Martinez-Caceres/Borras group 78 has shown that myelin peptide-pulsed tolDC can induce anergy in myelin-specific T cells from relapsing–remitting MS patients. The group are currently preparing for a tolDC trial in MS in the near future (Eva Martinez-Caceres, personal communication).
The first clinical trial with tolDC was carried out by the Giannoukakis/Trucco team at the University of Pittsburgh School of Medicine, and the results were published in 2011 76. They conducted a randomized, double-blind, Phase I study with tolDC in patients who had insulin-requiring type I diabetes for at least 5 years. Patients were injected with autologous, monocyte-derived DC that were either unmanipulated (control DC; three patients treated) or were treated ex vivo with anti-sense oligonucleotides targeting the CD40, CD80 and CD86 co-stimulatory molecules (tolDC; seven patients treated). This type of tolDC is defined by low surface levels of CD40, CD80 and CD86, and because of their low expression of CD40 they do not produce high levels of cytokine (IL-12, TNF) upon CD40 ligation 79. Furthermore, mouse analogues of these co-stimulatory-attenuated tolDC have been shown to prevent diabetes onset in non-obese diabetic (NOD) mice 79. Ten million control DC or tolDC were injected intradermally into the abdominal wall once every 2 weeks for a total of four administrations, and patients were monitored subsequently for a period of 12 months. DC treatment was well tolerated without any adverse events. DC treatment did not increase or induce autoantibodies (e.g. insulinoma-associated protein-2 antibodies). Furthermore, despite the fact that serum levels of IL-10 and IL-4 were increased, patients did not lose their capability to mount T cell responses to viral peptides or allogeneic cells, indicating that DC treatment did not result in systemic immunosuppression. The percentages of immune cell subsets in peripheral blood did not change after DC treatment, with the notable exception of B220+/CD11c– B cells. The proportions of this subset were increased significantly after DC treatment, although their levels returned to baseline after 6 months of treatment. This subset of B cells displayed suppressive activity in vitro and their proportional enhancement may be a beneficial effect of DC treatment. Overall, there were no notable differences between treatment with control DC and tolDC. Control DC were immature and therefore in a tolerogenic state; thus, it is not surprising that both types of DC exerted similar, potentially ‘pro-tolerogenic’ effects, i.e. enhancing IL-4 and IL-10 and the proportion of regulatory B cells. However, as it cannot be excluded that immature DC may become immunogenic DC in vivo, treatment with stable tolDC remains the preferred option.
A Phase I study with autologous tolDC in patients with RA has been carried out by Ranjeny Thomas and colleagues at the University of Queensland. Preliminary data were reported at the European League against Rheumatism meeting (EULAR) in 2011 77. In this study tolDC were generated by treatment of monocyte-derived DC with an inhibitor of NFκB signalling, BAY 11–7082, shown previously to maintain mouse DC in a tolerogenic state by preventing DC maturation 54,80. BAY-treated tolDC are deficient for CD40 expression but express high levels of CD86 80,81; thus, they are phenotypically different from the co-stimulation-attenuated tolDC developed by the Giannoukakis/Trucco team 79. Furthermore, unlike the trial in type I diabetes, in which tolDC were not loaded with a relevant autoantigen, in this trial tolDC were pulsed with four citrullinated peptide antigens. The final, antigen-pulsed, tolDC product is referred to as ‘Rheumavax’. A total of 18 patients [all human leucocyte antigen D-related (HLA-DR) shared epitope-positive] received a single dose (either 1 million or 5 million) of Rheumavax intradermally and were evaluated at baseline and after 3 and 6 months of treatment. Similarly to the tolDC trial in type I diabetes, Rheumavax was well tolerated; no major adverse effects were observed, and treatment did not appear to enhance the autoinflammatory response. Further assessments on how Rheumavax treatment has modulated anti-citrullinated peptide-specific immunity will be highly informative for understanding how tolDC affect antigen-specific T cell responses.
The main conclusion that can be drawn from these trials is that intradermal injection of autologous tolDC that are maturation-resistant appears to be safe – the autoimmune response was not enhanced. Although these trials were primarily safety trials, not designed to measure efficacy, they represent an important step forward in the field, and will pave the way for future tolDC trials.
Developing tolDC for the treatment of RA
We have developed a protocol to produce tolDC for the treatment of RA (Fig. 1) by pharmacological modulation of monocyte-derived DC with the immunosuppressive agents dexamethasone (Dex) and vitamin D3 [1,25 dihydroxyvitamin D3 (VitD3)], together with a Toll-like receptor (TLR)-4 agonist [Escherichia coli LPS or monophosphoryl lipid A (MPLA); see below]. Compared to mature DC, our tolDC are characterized by (i) high expression of MHC class II (i.e. similar levels as mature DC); (ii) intermediate expression of co-stimulatory molecules CD80 and CD86 and low expression of CD40 and CD83; and (iii) an anti-inflammatory cytokine production profile with high levels of IL-10 and TGF-β and low or undetectable levels of IL-12, IL-23 and TNF (55,82,83 and unpublished data). There are two reasons for including a TLR-4 ligand in the tolDC generation protocol. First, activation through TLR-4 is required for tolDC to process and present exogenous antigen efficiently on MHC class II 82; a similar observation has been reported for immunogenic DC 84. Thus, MHC class II–peptide complexes do not form efficiently unless the (tol)DC also receives a proinflammatory signal (e.g. LPS) during antigen uptake 82,84. The ability of tolDC to present antigens is clearly critical to the success of tolDC therapy, because the main goal of tolDC therapy is to induce T cell tolerance to relevant autoantigens. Secondly, TLR-4-mediated activation is also required for tolDC to acquire the ability to migrate in a CCR7-dependent manner 82, thus enabling them to migrate to secondary lymphoid tissues, where they can interact with T cells. Whether this migratory capacity is required for tolDC therapy to be successful in RA is not entirely clear, but there is evidence from the transplant setting that CCR7 expression by tolDC is required to prolong the survival of allografts in an animal model 85. These data fit the paradigm that secondary lymphoid tissues are an important site for the induction of immune tolerance 86,87, at least under normal, steady state conditions. However, the situation may be different in RA, in which diseased joint tissues are heavily infiltrated by T cells and antigen-presenting cells, including DC 88,89. Under these circumstances it is highly likely that presentation of autoantigen also takes place in the joint. Therefore, it could be speculated that, in RA, tolDC would ideally have the ability to act in several locations: in the rheumatoid joint to anergize autoantigen-specific effector T cells locally, and in the draining lymph node to induce Tregs from autoantigen-specific naive T cells. However, it should be noted that T cells from RA patients can be resistant to at least some tolerogenic signals; for instance, they can resist IL-10- and IDO-mediated suppression 90,91. Our tolDC operate, at least partially, via a TGF-β-dependent mechanism and inhibit proliferation and IFN-γ production of peripheral blood RA T cells in vitro (unpublished data); however, whether they can inhibit autoreactive T cells in the rheumatoid joint remains to be determined.
Fig. 1.
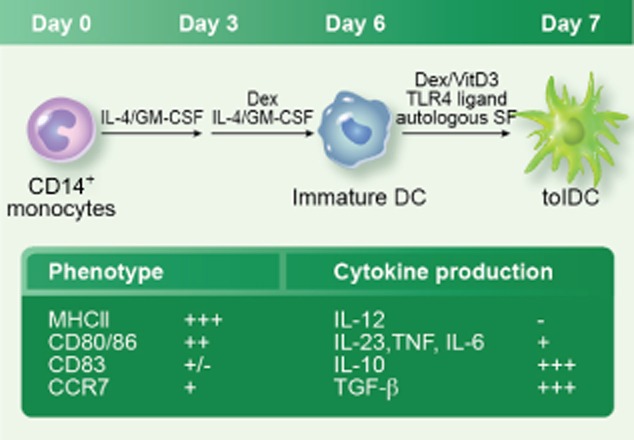
Generation of tolerogenic dendritic cells (tolDC). Peripheral blood CD14+ monocytes are cultured with interleukin (IL)-4 and granulocyte–macrophage colony-stimulating factor (GM-CSF) to generate immature DC. On day 3 of culture, cells receive fresh medium containing IL-4, GM-CSF and dexamethasone (Dex). After 6 days of culture monocytes will have differentiated into immature DC. These immature DC are then treated with Dex, vitamin D3 (VitD3) and a Toll-like receptor (TLR)-4 agonist [e.g. lipopolysaccharide (LPS) or for therapeutic grade tolDC, the current good manufacturing practice (cGMP)-compatible monophosphoryl lipid A] for 24 h to generate tolDC. During the final 24 h synovial fluid (SF) can be added to the cultures as a source of joint-associated autoantigen. The box summarizes typical tolDC features: +++ high expression; ++ intermediate expression; + low expression; +/– very low but detectable expression; – undetectable expression.
Despite the fact that our tolDC have similar ability as mature DC to process and present exogenous antigen, tolDC have lower T cell stimulatory capacity than mature DC, in line with their lower expression of co-stimulatory molecules and low production of proinflammatory cytokines 55,82. Moreover, tolDC induce hyporesponsiveness (‘anergy’) in antigen-experienced memory T cells while polarizing naive T cells towards an anti-inflammatory cytokine profile 55. We have also shown that, in a mouse in-vivo model of collagen-induced arthritis, murine bone marrow-derived tolDC generated with Dex, VitD3 and LPS have a therapeutic effect: treatment of arthritic mice with tolDC (1 million cells injected intravenously three times over 8 days) reduced significantly the severity and progression of arthritis, whereas treatment with immunogenic mature DC did not reduce disease and, in fact, exacerbated arthritis 49. Interestingly, tolDC exerted a therapeutic effect only if they had been loaded with the immunizing antigen, type II collagen. Treatment with tolDC was associated with a reduction in Th17 cells and an enhancement of IL-10-producing T cells, and a reduction in type II collagen-specific T cell proliferation, possibly explaining their therapeutic effect. Thus, this type of tolDC is a potentially powerful tool for the treatment of RA and other autoimmune diseases.
Clinical grade tolDC
Before tolDC can be applied in a clinical trial, a protocol to generate clinical grade tolDC, compliant with current good manufacturing practice (cGMP) regulations, had to be established. For this purpose, the research-grade fetal calf serum (FCS)-containing medium was replaced with cGMP-grade medium specialized for DC (CellGro® DC medium from CellGenix, Freiburg, Germany) and LPS was replaced with MPLA, a synthetic cGMP-grade TLR-4 ligand (from Avanti Polar Lipids, Alabaster, AL, USA). MPLA has strong adjuvant capacity but without the LPS-associated toxicity; it has been used safely in clinical trials testing next-generation vaccines 92. Clinical-grade tolDC have typical pro-tolerogenic features, including intermediate expression of co-stimulatory molecules and an anti-inflammatory cytokine profile. They induce T cell hyporesponsiveness and have the ability to inhibit T cell responses induced by mature DC 83. Despite the fact that monocyte-derived DC from RA patients with active disease are in an enhanced proinflammatory state 93,94, our protocol robustly generates tolDC from RA patients that are indistinguishable from healthy donor DC 83.
Importantly, tolDC exposed to proinflammatory cytokines, TLR ligands or RA synovial fluid retain their pro-tolerogenic features in vitro (83 and our unpublished data); whether they remain stable in vivo remains to be determined. However, it should be noted that equivalent Dex/VitD3/LPS-modulated mouse tolDC exerted their pro-tolerogenic in vivo in a proinflammatory environment, suggesting that their tolerogenic phenotype and function was not reverted in vivo 49. Furthermore, it has been shown that mouse tolDC generated with anti-sense oligonucleotides for CD40, CD80 and CD86 remained co-stimulatory-deficient in vivo, even after 3 weeks of injection 79.
Because tolDC therapy is designed to target autoantigen-specific T cells, a major consideration is the choice of autoantigen. However, reactivity to known autoantigens varies between RA patients and no universal autoantigen has yet been identified to which all RA patients respond. Furthermore, there is no validated, robust and reliable technique for defining autoantigen-responsiveness for an individual RA patient. We have therefore chosen to use autologous synovial fluid (SF) as a source of autoantigen, because a wide range of self-proteins are present in the SF of RA patients, including proteins containing autoantigenic T cell epitopes (e.g. HCgp39 and type II collagen) that can be processed efficiently and presented by DC 95–97.
The final tolDC product needs to conform to a list of predefined quality control (QC) criteria, which relate to the sterility, viability, purity and the ‘functionality’ of the product. Functional essays (e.g. induction of IL-10-producing Tr1 cells) are unsuitable for establishing the latter QC as they require at least 10 days to complete, whereas a rapid read-out is needed for QC testing. What is required is an assay that predicts product functionality with a read-out within hours, rather than days, as was established recently for Tregs 98. In the case of tolDC, low expression of CD83, non-detectable production of IL-12 and high secretion levels of IL-10 were chosen as QC markers as they correlate with tolDC function.
Autologous tolerogenic dendritic cells for RA (AUTODECRA trial)
We have designed a clinical trial to study autologous tolDC in RA (AUTODECRA), for which we are currently recruiting patients. It is a randomized, unblinded, placebo-controlled, dose-escalation Phase I study. Three dosing cohorts are planned: 1 × 106, 3 × 106 and 10 × 106 viable TolDC per patient. A major difference between this trial and the previous tolDC trials is the route of administration. TolDC will be injected intra-articularly, under arthroscopic guidance. Before tolDC are administered the joint will be irrigated with saline; ‘placebo’ patients will receive saline irrigation alone. The reason that tolDC will be administered directly into an affected knee joint is not only that it is beneficial from a safety perspective (if the joint flares up it can be irrigated again, followed by an intra-articular injection with corticosteroids) but also allows the collection of synovial biopsies for the analysis of potential response biomarkers. Intra-articular administration may also provide benefits compared with systemic administration, as tolDC are targeted to the diseased tissue. Furthermore, tolDC may migrate to the regional lymph nodes, where they could provide immunoregulatory signals required for immune tolerance induction.
The primary objective of AUTODECRA is to assess the safety of intra-articular administration of tolDC in patients with RA. The secondary objective is to assess the tolerability/acceptability to patients and feasibility of tolDC treatment. The trial also has a number of exploratory objectives, including assessing the effects of intra-articular tolDC administration on RA disease activity (locally and systemically) and investigating prospective response biomarkers in both synovial tissue and peripheral blood, taken at several time-points (see Fig. 2). The mechanisms underlying induction of immune tolerance in vivo are still poorly understood, and therefore no comprehensive set of suitable biomarkers can be predicted. Our biomarker analyses will therefore utilize a hypothesis-free approach and include leucocyte subset analysis by flow cytometry (e.g. DC subsets, T/B cell subsets), transcriptional profiling and immunohistochemistry. The latter will assess semi-quantitatively synovitis and cell subsets in the synovial membrane. Findings from the transplantation field have suggested that we are more likely to find tolerance biomarkers in the synovial tissue than in the peripheral blood, and that unexpected signals may emerge, hence the need for approaches such as transcriptional profiling 99. While we will attempt to study systemic autoreactivity before and after therapy, the uncertain nature of RA autoantigens renders this approach challenging.
Fig. 2.
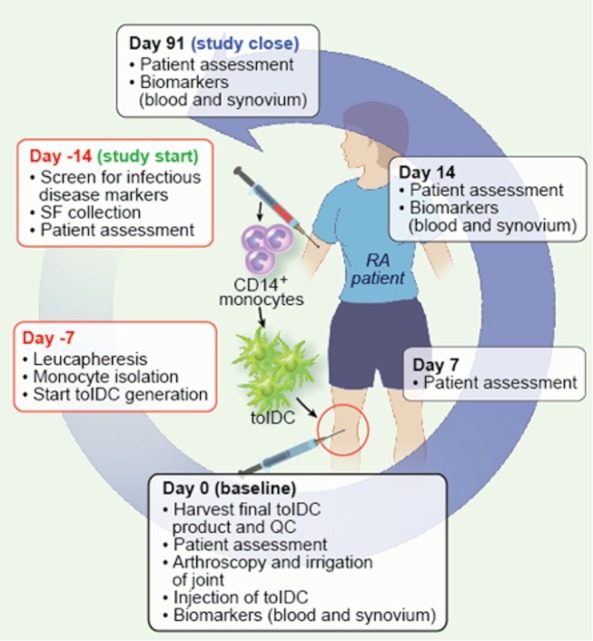
Autologous dendritic cells (DC) for rheumatoid arthritis (RA) (AUTODECRA) trial. The diagram depicts the autologous nature of tolerogenic dendritic cell (tolDC) treatment: monocytes are isolated from the patient's own peripheral blood product obtained by leucopheresis, cultured ex vivo under current good manufacturing practice (cGMP) conditions to generate tolDC (as depicted in Fig. 1) and injected into the patient's knee joint under arthroscopic guidance. The boxes summarize the procedures at different time-points during the trial (anti-clockwise). The trial starts 14 days before the patient is injected with tolDC (day −14). The patient is screened for mandatory disease markers to ensure a clean blood product in the cGMP facilities. Synovial fluid (SF) is collected as a source of joint-associated antigens and is added to the tolDC culture (see Fig. 1). Patient assessment takes place at this and other time-points during the trial (see boxes) and includes a knee assessment, health questionnaire and establishment of the disease activity score of 28 joints (DAS28). Peripheral blood samples (for collection of immune cell and serum) and synovial biopsies for biomarker analyses are taken before and after tolDC treatment (see boxes). If no serious adverse effects of tolDC are observed (e.g. knee flare) the study closes on day 91.
Challenges for design of tolDC trials
In addition to issues relating to the development and manufacture of tolDC for clinical application, there are a number of challenges relating to the design of clinical trials. The timing of tolDC treatment is an important issue. In the transplantation setting tolerogenic therapies can be applied before transplanting the graft, allowing for tolerance induction in an unprimed immune system. However, in the autoimmune setting this is not the case, and tolDC will be administered to patients with ongoing autoimmune disease, in whom dysregulated autoimmune responses have already been established. In RA, in particular, breach of tolerance to autoantigens may develop many years before the first symptoms of arthritis appear 100. It is generally thought that tolerogenic treatments, including tolDC therapy, will have the greatest chance of success if they are applied early on in the disease process 101. However, for safety reasons, new experimental therapies are being tested in patients with established disease who have failed other treatments and have a poor prognosis. Whether tolerogenic strategies can be successful under these conditions remains to be seen, and an obvious risk is that further development of tolDC therapy may not take place if initial trials show no or little efficacy. A related concern, therefore, is how to measure efficacy. The goal of tolDC therapy is to induce immune tolerance, but this may take time to develop and may not necessarily result in an immediate reduction of inflammation or other chronic disease symptoms. It has been observed that some immunomodulatory therapies that were ineffective in the short term appeared to provide benefits to RA patients in the longer term 102. Therefore, the timing of the end-points as well as what outcomes are being measured need careful consideration; current outcome measures for clinical trials in RA measure the consequences of inflammation, but this is unlikely to be an appropriate marker for the short-term ‘success’ of tolDC therapy. What is badly needed is the development of appropriate biomarkers of tolerance induction, which could then be used to monitor and guide tolerogenic therapies such as tolDC. Collecting data on expression of tolerance-related genes and the function of relevant immune subsets pre- and post-treatment will be essential for the design of a robust and quantifiable biomarker set. Such a set would enable us to measure the short-term therapeutic response in future tolerogenic therapy trials and, if standardized, would enable comparisons between different trials.
Future perspectives and concluding remarks
Over the last decade a variety of methods have been developed to generate tolDC in the laboratory. The characteristics of these tolDC have been defined extensively in in-vitro studies and their therapeutic potential has been demonstrated in experimental animal models of autoimmune disease. The field has now moved into a new era, translating these findings towards clinical application of tolDC. The first clinical trials have indicated that tolDC administration is tolerated and appears safe, and further studies now need to be conducted to establish their efficacy in treating autoimmune disorders, including RA, type 1 diabetes and MS. A major drawback of tolDC therapy is that it is a highly customized ‘bespoke’ therapy, which not only makes it expensive but also limits its application to centres that have appropriate facilities and are specialized in cellular therapies. Nevertheless, if tolDC trials in humans are successful, they will provide important ‘proof-of-principle’ data on therapeutic tolerance induction in humans. Lessons learned from tolDC trials, relating particularly to biomarker identification, should assist the development and clinical translation of new tolerance-inducing strategies, e.g. strategies that directly target and enhance the tolerogenic function of DC in vivo, or strategies that combine tolDC therapy with other treatments. For example, it has been shown that the combination of tolDC treatment with CTLA-4Ig prolongs allograft survival significantly in an animal model 31. The success of human tolDC trials will be enhanced by the definition of a robust set of biomarkers; without such a set it may prove difficult to establish if immune tolerance has been achieved. Furthermore, defining and standardizing biomarker analyses will be important to compare the results from different therapeutic tolerance strategies and trials.
Acknowledgments
The authors are supported by grants from Arthritis Research UK, Medical Research Council (MRC), Biotechnology and Biological Sciences Research Council (BBSRC) and the J.G.W. Patterson Foundation. Research in the Musculoskeletal Research Group is supported by the National Institute for Health Research Newcastle Biomedical Research Centre based at Newcastle Hospitals Foundation Trust and Newcastle University. The views expressed are those of the author(s) and not necessarily those of the NHS, the NIHR or the Department of Health.
Disclosure
The authors have no competing interests.
References
- 1.Londei M, Savill CM, Verhoef A, et al. Persistence of collagen type II-specific T-cell clones in the synovial membrane of a patient with rheumatoid arthritis. Proc Natl Acad Sci USA. 1989;86:636–640. doi: 10.1073/pnas.86.2.636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fritsch R, Eselbock D, Skriner K, et al. Characterization of autoreactive T cells to the autoantigens heterogeneous nuclear ribonucleoprotein A2 (RA33) and filaggrin in patients with rheumatoid arthritis. J Immunol. 2002;169:1068–1076. doi: 10.4049/jimmunol.169.2.1068. [DOI] [PubMed] [Google Scholar]
- 3.van Bilsen JH, van Dongen H, Lard LR, et al. Functional regulatory immune responses against human cartilage glycoprotein-39 in health vs. proinflammatory responses in rheumatoid arthritis. Proc Natl Acad Sci USA. 2004;101:17180–17185. doi: 10.1073/pnas.0407704101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.de Jong H, Berlo SE, Hombrink P, et al. Cartilage proteoglycan aggrecan epitopes induce proinflammatory autoreactive T-cell responses in rheumatoid arthritis and osteoarthritis. Ann Rheum Dis. 2010;69:255–262. doi: 10.1136/ard.2008.103978. [DOI] [PubMed] [Google Scholar]
- 5.Feitsma AL, van der Voort EI, Franken KL, et al. Identification of citrullinated vimentin peptides as T cell epitopes in HLA-DR4-positive patients with rheumatoid arthritis. Arthritis Rheum. 2010;62:117–125. doi: 10.1002/art.25059. [DOI] [PubMed] [Google Scholar]
- 6.James EA, Moustakas AK, Bui J, et al. HLA-DR1001 presents ‘altered-self’ peptides derived from joint-associated proteins by accepting citrulline in three of its binding pockets. Arthritis Rheum. 2010;62:2909–2918. doi: 10.1002/art.27594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.von Delwig A, Locke J, Robinson JH, et al. Response of Th17 cells to a citrullinated arthritogenic aggrecan peptide in patients with rheumatoid arthritis. Arthritis Rheum. 2010;62:143–149. doi: 10.1002/art.25064. [DOI] [PubMed] [Google Scholar]
- 8.Catalan D, Aravena O, Zuniga R, et al. Weak CD4+ T-cell responses to citrullinated vimentin in rheumatoid arthritis patients carrying HLA-DR9 alleles. Rheum Int. 2012;32:1819–1825. doi: 10.1007/s00296-011-2039-z. [DOI] [PubMed] [Google Scholar]
- 9.Ehrenstein MR, Evans JG, Singh A, et al. Compromised function of regulatory T cells in rheumatoid arthritis and reversal by anti-TNFalpha therapy. J Exp Med. 2004;200:277–285. doi: 10.1084/jem.20040165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Singh JA, Christensen R, Wells GA, et al. Biologics for rheumatoid arthritis: an overview of Cochrane reviews. Cochrane Database Syst Rev. 2009;(4) doi: 10.1002/14651858.CD007848.pub2. CD007848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Isaacs JD. Therapeutic agents for patients with rheumatoid arthritis and an inadequate response to tumour necrosis factor-alpha antagonists. Exp Opin Biol Ther. 2009;9:1463–1475. doi: 10.1517/14712590903379494. [DOI] [PubMed] [Google Scholar]
- 12.Salliot C, Gossec L, Ruyssen-Witrand A, et al. Infections during tumour necrosis factor-alpha blocker therapy for rheumatic diseases in daily practice: a systematic retrospective study of 709 patients. Rheumatology. 2007;46:327–334. doi: 10.1093/rheumatology/kel236. [DOI] [PubMed] [Google Scholar]
- 13.Gomez-Reino JJ, Carmona L, Angel Descalzo M. Risk of tuberculosis in patients treated with tumor necrosis factor antagonists due to incomplete prevention of reactivation of latent infection. Arthritis Rheum. 2007;57:756–761. doi: 10.1002/art.22768. [DOI] [PubMed] [Google Scholar]
- 14.Carson KR, Evens AM, Richey EA, et al. Progressive multifocal leukoencephalopathy after rituximab therapy in HIV-negative patients: a report of 57 cases from the Research on Adverse Drug Events and Reports project. Blood. 2009;113:4834–4840. doi: 10.1182/blood-2008-10-186999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Steinman RM, Nussenzweig MC. Avoiding horror autotoxicus: the importance of dendritic cells in peripheral T cell tolerance. Proc Natl Acad Sci USA. 2002;99:351–358. doi: 10.1073/pnas.231606698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ohnmacht C, Pullner A, King SB, et al. Constitutive ablation of dendritic cells breaks self-tolerance of CD4 T cells and results in spontaneous fatal autoimmunity. J Exp Med. 2009;206:549–559. doi: 10.1084/jem.20082394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Collin M, Bigley V, Haniffa M, et al. Human dendritic cell deficiency: the missing ID? Nature reviews. Immunology. 2011;11:575–583. doi: 10.1038/nri3046. [DOI] [PubMed] [Google Scholar]
- 18.Steinman RM, Hawiger D, Nussenzweig MC. Tolerogenic dendritic cells. Annu Rev Immunol. 2003;21:685–711. doi: 10.1146/annurev.immunol.21.120601.141040. [DOI] [PubMed] [Google Scholar]
- 19.Moser M. Dendritic cells in immunity and tolerance – do they display opposite functions? Immunity. 2003;19:5–8. doi: 10.1016/s1074-7613(03)00182-1. [DOI] [PubMed] [Google Scholar]
- 20.Jonuleit H, Schmitt E, Schuler G, et al. Induction of interleukin 10-producing, nonproliferating CD4(+) T cells with regulatory properties by repetitive stimulation with allogeneic immature human dendritic cells. J Exp Med. 2000;192:1213–1222. doi: 10.1084/jem.192.9.1213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hawiger D, Inaba K, Dorsett Y, et al. Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo. J Exp Med. 2001;194:769–779. doi: 10.1084/jem.194.6.769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Probst HC, Lagnel J, Kollias G, et al. Inducible transgenic mice reveal resting dendritic cells as potent inducers of CD8+ T cell tolerance. Immunity. 2003;18:713–720. doi: 10.1016/s1074-7613(03)00120-1. [DOI] [PubMed] [Google Scholar]
- 23.Diebold SS. Activation of dendritic cells by Toll-like receptors and C-type lectins. Handb Exp Pharmacol. 2009;188:3–30. doi: 10.1007/978-3-540-71029-5_1. [DOI] [PubMed] [Google Scholar]
- 24.Kalinski P, Hilkens CM, Wierenga EA, et al. T-cell priming by type-1 and type-2 polarized dendritic cells: the concept of a third signal. Immunol Today. 1999;20:561–567. doi: 10.1016/s0167-5699(99)01547-9. [DOI] [PubMed] [Google Scholar]
- 25.Diebold SS. Determination of T-cell fate by dendritic cells. Immunol Cell Biol. 2008;86:389–397. doi: 10.1038/icb.2008.26. [DOI] [PubMed] [Google Scholar]
- 26.McGuirk P, McCann C, Mills KH. Pathogen-specific T regulatory 1 cells induced in the respiratory tract by a bacterial molecule that stimulates interleukin 10 production by dendritic cells: a novel strategy for evasion of protective T helper type 1 responses by Bordetella pertussis. J Exp Med. 2002;195:221–231. doi: 10.1084/jem.20011288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.van der Kleij D, Latz E, Brouwers JF, et al. A novel host–parasite lipid cross-talk. Schistosomal lyso-phosphatidylserine activates Toll-like receptor 2 and affects immune polarization. J Biol Chem. 2002;277:48122–48129. doi: 10.1074/jbc.M206941200. [DOI] [PubMed] [Google Scholar]
- 28.de Jong EC, Vieira PL, Kalinski P, et al. Corticosteroids inhibit the production of inflammatory mediators in immature monocyte-derived DC and induce the development of tolerogenic DC3. J Leukoc Biol. 1999;66:201–204. doi: 10.1002/jlb.66.2.201. [DOI] [PubMed] [Google Scholar]
- 29.Steinbrink K, Graulich E, Kubsch S, et al. CD4(+) and CD8(+) anergic T cells induced by interleukin-10-treated human dendritic cells display antigen-specific suppressor activity. Blood. 2002;99:2468–2476. doi: 10.1182/blood.v99.7.2468. [DOI] [PubMed] [Google Scholar]
- 30.Sato K, Yamashita N, Baba M, et al. Regulatory dendritic cells protect mice from murine acute graft-versus-host disease and leukemia relapse. Immunity. 2003;18:367–379. doi: 10.1016/s1074-7613(03)00055-4. [DOI] [PubMed] [Google Scholar]
- 31.Lan YY, Wang Z, Raimondi G, et al. ‘Alternatively activated’ dendritic cells preferentially secrete IL-10, expand Foxp3+CD4+ T cells, and induce long-term organ allograft survival in combination with CTLA4-Ig. J Immunol. 2006;177:5868–5877. doi: 10.4049/jimmunol.177.9.5868. [DOI] [PubMed] [Google Scholar]
- 32.Josefowicz SZ, Lu LF, Rudensky AY. Regulatory T cells: mechanisms of differentiation and function. Annu Rev Immunol. 2012;30:531–564. doi: 10.1146/annurev.immunol.25.022106.141623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bilate AM, Lafaille JJ. Induced CD4+Foxp3+ regulatory T cells in immune tolerance. Annu Rev Immunol. 2012;30:733–758. doi: 10.1146/annurev-immunol-020711-075043. [DOI] [PubMed] [Google Scholar]
- 34.Qureshi OS, Zheng Y, Nakamura K, et al. Trans-endocytosis of CD80 and CD86: a molecular basis for the cell-extrinsic function of CTLA-4. Science. 2011;332:600–603. doi: 10.1126/science.1202947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gregori S, Goudy KS, Roncarolo MG. The cellular and molecular mechanisms of immuno-suppression by human type 1 regulatory T cells. Front Immunol. 2012;3:30. doi: 10.3389/fimmu.2012.00030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Maldonado RA, von Andrian UH. How tolerogenic dendritic cells induce regulatory T cells. Adv Immunol. 2010;108:111–165. doi: 10.1016/B978-0-12-380995-7.00004-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Manicassamy S, Pulendran B. Dendritic cell control of tolerogenic responses. Immunol Rev. 2011;241:206–227. doi: 10.1111/j.1600-065X.2011.01015.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mellor AL, Baban B, Chandler P, et al. Cutting edge: induced indoleamine 2,3 dioxygenase expression in dendritic cell subsets suppresses T cell clonal expansion. J Immunol. 2003;171:1652–1655. doi: 10.4049/jimmunol.171.4.1652. [DOI] [PubMed] [Google Scholar]
- 39.Goubier A, Dubois B, Gheit H, et al. Plasmacytoid dendritic cells mediate oral tolerance. Immunity. 2008;29:464–475. doi: 10.1016/j.immuni.2008.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Coombes JL, Siddiqui KR, Arancibia-Carcamo CV, et al. A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-beta and retinoic acid-dependent mechanism. J Exp Med. 2007;204:1757–1764. doi: 10.1084/jem.20070590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sun CM, Hall JA, Blank RB, et al. Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 Treg cells via retinoic acid. J Exp Med. 2007;204:1775–1785. doi: 10.1084/jem.20070602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Misra N, Bayry J, Lacroix-Desmazes S, et al. Cutting edge: human CD4+CD25+ T cells restrain the maturation and antigen-presenting function of dendritic cells. J Immunol. 2004;172:4676–4680. doi: 10.4049/jimmunol.172.8.4676. [DOI] [PubMed] [Google Scholar]
- 43.Kryczek I, Wei S, Zou L, et al. Cutting edge: induction of B7-H4 on APCs through IL-10: novel suppressive mode for regulatory T cells. J Immunol. 2006;177:40–44. doi: 10.4049/jimmunol.177.1.40. [DOI] [PubMed] [Google Scholar]
- 44.Ezzelarab M, Thomson AW. Tolerogenic dendritic cells and their role in transplantation. Semin Immunol. 2011;23:252–263. doi: 10.1016/j.smim.2011.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Beriou G, Moreau A, Cuturi MC. Tolerogenic dendritic cells: applications for solid organ transplantation. Curr Opin Organ Transplant. 2012;17:42–47. doi: 10.1097/MOT.0b013e32834ee662. [DOI] [PubMed] [Google Scholar]
- 46.Voigtlander C, Rossner S, Cierpka E, et al. Dendritic cells matured with TNF can be further activated in vitro and after subcutaneous injection in vivo which converts their tolerogenicity into immunogenicity. J Immunother. 2006;29:407–415. doi: 10.1097/01.cji.0000210081.60178.b4. [DOI] [PubMed] [Google Scholar]
- 47.Lim DS, Kang MS, Jeong JA, et al. Semi-mature DC are immunogenic and not tolerogenic when inoculated at a high dose in collagen-induced arthritis mice. Eur J Immunol. 2009;39:1334–1343. doi: 10.1002/eji.200838987. [DOI] [PubMed] [Google Scholar]
- 48.Ludewig B, Ochsenbein AF, Odermatt B, et al. Immunotherapy with dendritic cells directed against tumor antigens shared with normal host cells results in severe autoimmune disease. J Exp Med. 2000;191:795–804. doi: 10.1084/jem.191.5.795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Stoop JN, Harry RA, von Delwig A, et al. Therapeutic effect of tolerogenic dendritic cells in established collagen-induced arthritis is associated with a reduction in Th17 responses. Arthritis Rheum. 2010;62:3656–3665. doi: 10.1002/art.27756. [DOI] [PubMed] [Google Scholar]
- 50.Hilkens CM, Isaacs JD, Thomson AW. Development of dendritic cell-based immunotherapy for autoimmunity. Int Rev Immunol. 2010;29:156–183. doi: 10.3109/08830180903281193. [DOI] [PubMed] [Google Scholar]
- 51.Matasic R, Dietz AB, Vuk-Pavlovic S. Dexamethasone inhibits dendritic cell maturation by redirecting differentiation of a subset of cells. J Leukoc Biol. 1999;66:909–914. doi: 10.1002/jlb.66.6.909. [DOI] [PubMed] [Google Scholar]
- 52.Giannoukakis N, Bonham CA, Qian S, et al. Prolongation of cardiac allograft survival using dendritic cells treated with NF-kB decoy oligodeoxyribonucleotides. Mol Ther. 2000;1:430–437. doi: 10.1006/mthe.2000.0060. [DOI] [PubMed] [Google Scholar]
- 53.Penna G, Adorini L. 1 Alpha,25-dihydroxyvitamin D3 inhibits differentiation, maturation, activation, and survival of dendritic cells leading to impaired alloreactive T cell activation. J Immunol. 2000;164:2405–2411. doi: 10.4049/jimmunol.164.5.2405. [DOI] [PubMed] [Google Scholar]
- 54.Martin E, Capini C, Duggan E, et al. Antigen-specific suppression of established arthritis in mice by dendritic cells deficient in NF-kappaB. Arthritis Rheum. 2007;56:2255–2266. doi: 10.1002/art.22655. [DOI] [PubMed] [Google Scholar]
- 55.Anderson AE, Sayers BL, Haniffa MA, et al. Differential regulation of naive and memory CD4+ T cells by alternatively activated dendritic cells. J Leukoc Biol. 2008;84:124–133. doi: 10.1189/jlb.1107744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lu L, Gambotto A, Lee WC, et al. Adenoviral delivery of CTLA4Ig into myeloid dendritic cells promotes their in vitro tolerogenicity and survival in allogeneic recipients. Gene Ther. 1999;6:554–563. doi: 10.1038/sj.gt.3300862. [DOI] [PubMed] [Google Scholar]
- 57.Kim SH, Kim S, Evans CH, et al. Effective treatment of established murine collagen-induced arthritis by systemic administration of dendritic cells genetically modified to express IL-4. J Immunol. 2001;166:3499–3505. doi: 10.4049/jimmunol.166.5.3499. [DOI] [PubMed] [Google Scholar]
- 58.Morita Y, Yang J, Gupta R, et al. Dendritic cells genetically engineered to express IL-4 inhibit murine collagen-induced arthritis. J Clin Invest. 2001;107:1275–1284. doi: 10.1172/JCI11490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Coates PT, Krishnan R, Kireta S, et al. Human myeloid dendritic cells transduced with an adenoviral interleukin-10 gene construct inhibit human skin graft rejection in humanized NOD-scid chimeric mice. Gene Ther. 2001;8:1224–1233. doi: 10.1038/sj.gt.3301513. [DOI] [PubMed] [Google Scholar]
- 60.Tan PH, Yates JB, Xue SA, et al. Creation of tolerogenic human dendritic cells via intracellular CTLA4: a novel strategy with potential in clinical immunosuppression. Blood. 2005;106:2936–2943. doi: 10.1182/blood-2005-05-1826. [DOI] [PubMed] [Google Scholar]
- 61.Min WP, Gorczynski R, Huang XY, et al. Dendritic cells genetically engineered to express Fas ligand induce donor-specific hyporesponsiveness and prolong allograft survival. J Immunol. 2000;164:161–167. doi: 10.4049/jimmunol.164.1.161. [DOI] [PubMed] [Google Scholar]
- 62.Kim SH, Kim S, Oligino TJ, et al. Effective treatment of established mouse collagen-induced arthritis by systemic administration of dendritic cells genetically modified to express FasL. Mol Ther. 2002;6:584–590. [PubMed] [Google Scholar]
- 63.Liu Z, Xu X, Hsu HC, et al. CII-DC-AdTRAIL cell gene therapy inhibits infiltration of CII-reactive T cells and CII-induced arthritis. J Clin Invest. 2003;112:1332–1341. doi: 10.1172/JCI19209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Liang X, Lu L, Chen Z, et al. Administration of dendritic cells transduced with antisense oligodeoxyribonucleotides targeting CD80 or CD86 prolongs allograft survival. Transplantation. 2003;76:721–729. doi: 10.1097/01.TP.0000076470.35404.49. [DOI] [PubMed] [Google Scholar]
- 65.Xu H, Chen T, Wang HQ, et al. Prolongation of rat intestinal allograft survival by administration of donor interleukin-12 p35-silenced bone marrow-derived dendritic cells. Transplant Proc. 2006;38:1561–1563. doi: 10.1016/j.transproceed.2006.03.022. [DOI] [PubMed] [Google Scholar]
- 66.Li R, Zheng X, Popov I, et al. Gene silencing of IL-12 in dendritic cells inhibits autoimmune arthritis. J Transl Med. 2012;10:19. doi: 10.1186/1479-5876-10-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Steinbrink K, Wolfl M, Jonuleit H, et al. Induction of tolerance by IL-10-treated dendritic cells. J Immunol. 1997;159:4772–4780. [PubMed] [Google Scholar]
- 68.Yarilin D, Duan R, Huang YM, et al. Dendritic cells exposed in vitro to TGF-beta1 ameliorate experimental autoimmune myasthenia gravis. Clin Exp Immunol. 2002;127:214–219. doi: 10.1046/j.1365-2249.2002.01748.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Boks MA, Kager-Groenland JR, Haasjes MS, et al. IL-10-generated tolerogenic dendritic cells are optimal for functional regulatory T cell induction – a comparative study of human clinical-applicable DC. Clin Immunol. 2012;142:332–342. doi: 10.1016/j.clim.2011.11.011. [DOI] [PubMed] [Google Scholar]
- 70.Turnquist HR, Raimondi G, Zahorchak AF, et al. Rapamycin-conditioned dendritic cells are poor stimulators of allogeneic CD4+ T cells, but enrich for antigen-specific Foxp3+ T regulatory cells and promote organ transplant tolerance. J Immunol. 2007;178:7018–7031. doi: 10.4049/jimmunol.178.11.7018. [DOI] [PubMed] [Google Scholar]
- 71.Salazar L, Aravena O, Abello P, et al. Modulation of established murine collagen-induced arthritis by a single inoculation of short-term lipopolysaccharide-stimulated dendritic cells. Ann Rheum Dis. 2008;67:1235–1241. doi: 10.1136/ard.2007.072199. [DOI] [PubMed] [Google Scholar]
- 72.Zheng J, Jiang HY, Li J, et al. MicroRNA-23b promotes tolerogenic properties of dendritic cells in vitro through inhibiting Notch1/NF-kappaB signalling pathways. Allergy. 2012;67:362–370. doi: 10.1111/j.1398-9995.2011.02776.x. [DOI] [PubMed] [Google Scholar]
- 73.Valencia J, Hernandez-Lopez C, Martinez VG, et al. Wnt5a skews dendritic cell differentiation to an unconventional phenotype with tolerogenic features. J Immunol. 2011;187:4129–4139. doi: 10.4049/jimmunol.1101243. [DOI] [PubMed] [Google Scholar]
- 74.Stoop JN, Robinson JH, Hilkens CM. Developing tolerogenic dendritic cell therapy for rheumatoid arthritis: what can we learn from mouse models? Ann Rheum Dis. 2011;70:1526–1533. doi: 10.1136/ard.2011.151654. [DOI] [PubMed] [Google Scholar]
- 75.Bianco NR, Kim SH, Ruffner MA, et al. Therapeutic effect of exosomes from indoleamine 2,3-dioxygenase-positive dendritic cells in collagen-induced arthritis and delayed-type hypersensitivity disease models. Arthritis Rheum. 2009;60:380–389. doi: 10.1002/art.24229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Giannoukakis N, Phillips B, Finegold D, et al. Phase I (safety) study of autologous tolerogenic dendritic cells in type 1 diabetic patients. Diabetes Care. 2011;34:2026–2032. doi: 10.2337/dc11-0472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Thomas R, Street S, Ramnoruth N, et al. Safety and preliminary evidence of efficacy in a phase I clinical trial of autologous tolerising dendritic cells exposed to citrullinated peptides (Rheumavax) in patients with rheumatoid arthritis. Ann Rheum Dis. 2011;70(Suppl 3):169. [Google Scholar]
- 78.Raich-Regue D, Grau-Lopez L, Naranjo-Gomez M, et al. Stable antigen-specific T-cell hyporesponsiveness induced by tolerogenic dendritic cells from multiple sclerosis patients. Eur J Immunol. 2012;42:771–782. doi: 10.1002/eji.201141835. [DOI] [PubMed] [Google Scholar]
- 79.Machen J, Harnaha J, Lakomy R, et al. Antisense oligonucleotides down-regulating costimulation confer diabetes-preventive properties to nonobese diabetic mouse dendritic cells. J Immunol. 2004;173:4331–4341. doi: 10.4049/jimmunol.173.7.4331. [DOI] [PubMed] [Google Scholar]
- 80.Martin E, O'Sullivan B, Low P, et al. Antigen-specific suppression of a primed immune response by dendritic cells mediated by regulatory T cells secreting interleukin-10. Immunity. 2003;18:155–167. doi: 10.1016/s1074-7613(02)00503-4. [DOI] [PubMed] [Google Scholar]
- 81.Thompson AG, O'Sullivan BJ, Beamish H, et al. T cells signaled by NF-kappa B-dendritic cells are sensitized not anergic to subsequent activation. J Immunol. 2004;173:1671–1680. doi: 10.4049/jimmunol.173.3.1671. [DOI] [PubMed] [Google Scholar]
- 82.Anderson AE, Swan DJ, Sayers BL, et al. LPS activation is required for migratory activity and antigen presentation by tolerogenic dendritic cells. J Leukoc Biol. 2009;85:243–250. doi: 10.1189/jlb.0608374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Harry RA, Anderson AE, Isaacs JD, et al. Generation and characterisation of therapeutic tolerogenic dendritic cells for rheumatoid arthritis. Ann Rheum Dis. 2010;69:2042–2050. doi: 10.1136/ard.2009.126383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Inaba K, Turley S, Iyoda T, et al. The formation of immunogenic major histocompatibility complex class II-peptide ligands in lysosomal compartments of dendritic cells is regulated by inflammatory stimuli. J Exp Med. 2000;191:927–936. doi: 10.1084/jem.191.6.927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Garrod KR, Chang CK, Liu FC, et al. Targeted lymphoid homing of dendritic cells is required for prolongation of allograft survival. J Immunol. 2006;177:863–868. doi: 10.4049/jimmunol.177.2.863. [DOI] [PubMed] [Google Scholar]
- 86.Bai Y, Liu J, Wang Y, et al. L-selectin-dependent lymphoid occupancy is required to induce alloantigen-specific tolerance. J Immunol. 2002;168:1579–1589. doi: 10.4049/jimmunol.168.4.1579. [DOI] [PubMed] [Google Scholar]
- 87.Ochando JC, Yopp AC, Yang Y, et al. Lymph node occupancy is required for the peripheral development of alloantigen-specific Foxp3+ regulatory T cells. J Immunol. 2005;174:6993–7005. doi: 10.4049/jimmunol.174.11.6993. [DOI] [PubMed] [Google Scholar]
- 88.Cush JJ, Pietschmann P, Oppenheimer-Marks N, et al. The intrinsic migratory capacity of memory T cells contributes to their accumulation in rheumatoid synovium. Arthritis Rheum. 1992;35:1434–1444. doi: 10.1002/art.1780351206. [DOI] [PubMed] [Google Scholar]
- 89.Thomas R, Davis LS, Lipsky PE. Rheumatoid synovium is enriched in mature antigen-presenting dendritic cells. J Immunol. 1994;152:2613–2623. [PubMed] [Google Scholar]
- 90.Yamana J, Yamamura M, Okamoto A, et al. Resistance to IL-10 inhibition of interferon gamma production and expression of suppressor of cytokine signaling 1 in CD4+ T cells from patients with rheumatoid arthritis. Arthritis Res Ther. 2004;6:R567–577. doi: 10.1186/ar1445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Zhu L, Ji F, Wang Y, et al. Synovial autoreactive T cells in rheumatoid arthritis resist IDO-mediated inhibition. J Immunol. 2006;177:8226–8233. doi: 10.4049/jimmunol.177.11.8226. [DOI] [PubMed] [Google Scholar]
- 92.Cluff CW. Monophosphoryl lipid A (MPL) as an adjuvant for anti-cancer vaccines: clinical results. Adv Exp Med Biol. 2009;667:111–123. doi: 10.1007/978-1-4419-1603-7_10. [DOI] [PubMed] [Google Scholar]
- 93.Radstake TR, van Lent PL, Pesman GJ, et al. High production of proinflammatory and Th1 cytokines by dendritic cells from patients with rheumatoid arthritis, and down regulation upon FcgammaR triggering. Ann Rheum Dis. 2004;63:696–702. doi: 10.1136/ard.2003.010033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Baldwin HM, Ito-Ihara T, Isaacs JD, et al. Tumour necrosis factor alpha blockade impairs dendritic cell survival and function in rheumatoid arthritis. Ann Rheum Dis. 2010;69:1200–1207. doi: 10.1136/ard.2009.110502. [DOI] [PubMed] [Google Scholar]
- 95.Tsark EC, Wang W, Teng YC, et al. Differential MHC class II-mediated presentation of rheumatoid arthritis autoantigens by human dendritic cells and macrophages. J Immunol. 2002;169:6625–6633. doi: 10.4049/jimmunol.169.11.6625. [DOI] [PubMed] [Google Scholar]
- 96.Seward RJ, Drouin EE, Steere AC, et al. Peptides presented by HLA-DR molecules in synovia of patients with rheumatoid arthritis or antibiotic-refractory Lyme arthritis. Mol Cell Proteomics. 2011;10:M110–002477. doi: 10.1074/mcp.M110.002477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.van Beers JJ, Schwarte CM, Stammen-Vogelzangs J, et al. The RA synovial fluid citrullinome reveals novel citrullinated epitopes in apolipoprotein E, myeloid nuclear differentiation antigen and beta-actin. Arthritis Rheum. 2013;65:69–80. doi: 10.1002/art.37720. [DOI] [PubMed] [Google Scholar]
- 98.Canavan JB, Afzali B, Scotta C, et al. A rapid diagnostic test for human regulatory T-cell function to enable regulatory T-cell therapy. Blood. 2012;119:e57–66. doi: 10.1182/blood-2011-09-380048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Bohne F, Martinez-Llordella M, Lozano JJ, et al. Intra-graft expression of genes involved in iron homeostasis predicts the development of operational tolerance in human liver transplantation. J Clin Invest. 2012;122:368–382. doi: 10.1172/JCI59411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Nielen MM, van Schaardenburg D, Reesink HW, et al. Specific autoantibodies precede the symptoms of rheumatoid arthritis: a study of serial measurements in blood donors. Arthritis Rheum. 2004;50:380–386. doi: 10.1002/art.20018. [DOI] [PubMed] [Google Scholar]
- 101.Isaacs JD. The changing face of rheumatoid arthritis: sustained remission for all? Nat Rev Immunol. 2010;10:605–611. doi: 10.1038/nri2804. [DOI] [PubMed] [Google Scholar]
- 102.Morgan AW, Hale G, Rebello PR, et al. A pilot study of combination anti-cytokine and anti-lymphocyte biological therapy in rheumatoid arthritis. Q J Med. 2008;101:299–306. doi: 10.1093/qjmed/hcn006. [DOI] [PubMed] [Google Scholar]