

Abstract
Senescence is a stable cell cycle arrest program that contributes to tumor suppression, organismal aging and certain wound healing responses. During liver fibrosis, for example, hepatic stellate cells initially proliferate and secrete extracellular matrix components that produce fibrosis; however, these cells eventually senesce and are cleared by immune cells, including natural killer (NK) cells. Here, we examine how NK cells target senescent cells and assess the impact of this process on liver fibrosis. We show that granule exocytosis, but not death-receptor-mediated apoptosis, is required for NK-cell-mediated killing of senescent cells. This pathway bias is due to upregulation of the decoy death receptor, Dcr2, an established senescence marker that attenuates NK-mediated cell death. Accordingly, mice with defects in granule exocytosis accumulate senescent stellate cells and display more liver fibrosis in response to a fibrogenic agent. Our results thus provide new insights into the immune surveillance of senescent cells and reveal how granule exocytosis has a protective role against liver fibrosis.
Keywords: cellular senescence, perforin, liver fibrosis
Introduction
Cellular senescence is a state of stable cell cycle arrest that limits tumor development in response to oncogenic mutations and certain chemotherapies.1, 2, 3 Recent studies suggest that senescence can also act to limit wound healing responses and may participate in organismal aging. For instance, in liver fibrosis, the senescence of activated hepatic stellate cells (HSCs), the main fibrogenic cell type in the liver, limits tissue damage and accelerates repair,4 and similar mechanisms regulate tissue damage during wound healing in the skin and oral submucous fibrosis.5, 6 Moreover, mice engineered to eliminate senescent cells from tissues show improved fitness during longevity,7 and clearance of pre-cancerous senescent hepatocytes is crucial for liver cancer suppression.8 Hence, factors that control senescent cell numbers in vivo may broadly impact cancer progression, tissue damage and functional decline.
Senescence is accompanied by phenotypic and transcriptional changes that identify senescent cells in vitro and in vivo. For example, senescent cells display a large and flat morphology in vitro and upregulate a senescence-associated β-galactosidase (SA-β-gal).9 Senescent cells often display global changes in chromatin structure10 that are associated with downregulation of cell cycle genes and components of the extracellular matrix and upregulation of immune modulators and matrix degrading enzymes.4 Comparative analyses of gene expression data have produced some markers that appear specific for senescence,11 including the p15ink4b cyclin-dependent kinase inhibitor and the decoy receptor 2 (Dcr2, formally TNFRSF10D). Although p15ink4b likely contributes to the senescence-associated cell cycle arrest,12 whether decoy receptors or some other senescence markers actively participate in the program remains unknown.
Senescence acts through a coordinated program involving cell autonomous and cell nonautonomous components.13 In a cell autonomous manner, the Rb and p53 tumor suppressor pathways act to produce the stable cell cycle arrest that is the hallmark of senescence.1 These proteins are activated by, or activate, cyclin-dependent kinase inhibitors, such as p15ink4b, p16ink4a and p21, which lead to stable suppression of E2F target genes.10, 14 Secreted proteins, regulated at least partially by NF-κB, enhance cell cycle arrest and are largely responsible for mediating the impact of senescent cells on tissue biology.15, 16, 17 These factors can attract immune cells, including natural killer (NK) cells, triggering the recognition and ultimate clearance of the senescent cells from tumors or tissue.4, 18 Such mechanisms may be necessary to prevent the long-term damage that might be produced by senescent cells, and to facilitate tissue repair and homeostasis.
The mechanisms whereby NK cells eliminate senescent cells from tissues are not known. NK cells rely on two independent mechanisms to eliminate a variety of external and internal threats, including tumor cells.19, 20 The ligands on the surface of NK cells, TRAIL and FAS ligand (FasL) bind corresponding receptors on target cells leading to caspase activation and cell death—a process that can be exquisitely controlled though the expression of various positive and negative regulators.21, 22 NK cells can also eliminate target cells through granule exocytosis, a process involving the production of perforin and granzyme (A, B) containing granules, which are secreted from the NK cell upon interaction with the target cell.21, 23 Perforin is responsible for perforating the cell membrane and thus enabling granzyme release into the target cells where it can induce cell death by both caspase-dependent and independent pathways.24 Both pathways are necessary for efficient NK-mediated defense of the liver from carcinogenesis and metastasis.25, 26
Here, we set out to understand how NK cells eliminate senescent cells from tissues and the implications of such mechanisms on liver fibrosis. Our results indicate that the granule exocytosis, and not death-receptor-mediated apoptosis, is essential for the NK-mediated surveillance of the senescent cells and that disruption of this pathway leads to the accumulation of senescent cells in damaged livers and increased fibrosis. Our study thus provides the key biological and mechanistic insights into the immune surveillance of senescent cells.
Results
Efficient killing of senescent cells by NK cells
In order to understand how NK cells target senescent cells, we employed an in vitro cytotoxicity assay whereby normal and senescent cells are co-cultured with the NK cells.4 We incorporated as one model the human NK cell line, YT. The cells express components of both the death receptor and granule exocytosis pathways and can engage both mechanisms to eliminate target cells.27, 28 To determine whether cytotoxicity could be assessed over a wide dynamic range, we assessed viability using different ratios of target cells to NK cells. IMR-90 human diploid fibroblasts, which have been widely used to study senescence,10 were triggered to senescence by treatment with the DNA damage agent, Etoposide. Within days, these cells displayed senescence features including a stable cell cycle arrest and upregulation of the NK cell receptor ligands and adhesion molecules that could mediate their interaction with the NK cells.4
Senescent and growing (control) IMR-90 cells were co-cultured with YT cells for 12 h at 1:5, 1:10 and 1:20 target cell to NK-cell ratios. Of note, we were unable to measure NK-mediated cytotoxicity using traditional assays that rely on the loading of the target cells with 51Cr, because efficient loading requires a threshold level of cell proliferation29 that cannot be achieved in senescent cells. As an alternative, we measured cell death indirectly by assessing cell viability using crystal violet staining to quantify the remaining viable cells at the end of the co-incubation period (Figure 1) and corroborated our results using propidium iodide-based viability assays and time lapse video microscopy.4 Combined, these studies confirmed that reductions in viable cells assessed by crystal violet staining were associated with cell death. As expected, NK cells eliminated senescent cells at least twice as efficiently as growing cells throughout the range of target to effector cell ratios tested (P<0.005). Such enhanced sensitivity was also observed with a second NK cell line, NK-92, and primary human NK cells (see Supplementary Figures S1 and 3C). Hence, this system provides a robust assay to study NK-mediated killing of senescent cells.
Figure 1.

NK cells preferentially recognize senescent cells in a wide range of target:effector cell ratios. Senescent or growing IMR-90 fibroblasts were co-incubated with YT cells for 12 h at the indicated ratios and cytotoxicity was determined. The graphs represent the average and the s.e. of triplicate measurements from at least three independent experiments. *P<0.005.
Death receptor signaling is dispensable for NK-mediated targeting of senescent cells
To determine whether the increased sensitivity of senescent cells to NK cell killing might involve death receptor signaling, we first asked whether senescent cells were more sensitive to the death receptor ligand, FasL. At a concentration of 2 ng/ml, FasL induced threefold more killing in growing cells than in senescent IMR-90 cells (P<0.001, Figure 2a), though at higher concentrations this difference was not significant. Similarly, TRAIL induced even more efficient killing of senescent cells at the concentrations of 10 and 100 ng/ml (P<0.001 and P<0.0001, respectively, Supplementary Figure S2)
Figure 2.

Caspases are dispensable for NK-mediated cell killing of senescent cells. Senescent or growing IMR-90 fibroblasts were incubated for 12 h with either 2 or 10 nℳ FasL (a). Caspase inhibitors Z-VAD-FMK or Z-IEDT-FMK were added at the concentration of 10 μℳ as indicated. Cytotoxicity was determined at the end of the co-incubation period. Senescent or growing IMR-90 fibroblasts were co-incubated with YT cells for 12 h in the presence of 10 μℳ of caspase inhibitors Z-VAD-FMK or Z-IEDT-FMK and then the cytotoxicity was determined (b). The graphs represent the average and the s.e. of triplicate measurements from at least three independent experiments. *P<0.05, ***P<0.001.
We also examined whether blocking the death-receptor-mediated apoptosis using either the caspase 8 inhibitor Z-IEDT-FMK or the pan-caspase inhibitor Z-VAD-FMK would attenuate NK-mediated cell death. As expected, both compounds significantly inhibited FasL-induced cytotoxicity in both growing (P<0.05 and P<0.001, for Z-IEDT and Z-VAD, respectively) and senescent (P<0.001 and P<0.0001 for Z-IEDT and Z-VAD, respectively) cells (Figure 2a). Thus, as anticipated from other systems, the caspase inhibitors, Z-VAD-FMK and Z-IEDT-FMK, efficiently inhibit cell death induced by a death receptor ligand in senescent cells.
We next evaluated whether the death receptor pathway contributes to NK-mediated elimination of senescent cells. We co-incubated NK (YT) cells with growing or senescent IMR-90 cells in the presence or absence of either the caspase inhibitor Z-VAD-FMK or Z-IEDT-FMK. Neither inhibitor suppressed NK-cell-mediated cytotoxicity towards senescent cells (Figure 2b). In fact, the viability of the NK cells themselves was slightly increased in the presence of the inhibitors, perhaps suggesting that these effector cells target each other through death-receptor-mediated mechanisms. Therefore, the death receptor pathway is dispensable for NK cell targeting of senescent IMR-90 cells.
Granule exocytosis mediates the targeting of senescent cells by NK cells
We next took a pharmacological approach to explore the role of granule exocytosis in NK cell targeting of senescent cells. When the NK-cell-mediated killing assay was performed in the presence of the granule exocytosis inhibitor Concanamycin A (CMA), the specific killing of the senescent cells was almost completely eliminated at all target to effector cell ratios tested (Supplementary Figure S3). For example, at a target to effector cell ratio of 1:10, the average killing of senescent cells was reduced fourfold in the presence of the CMA (P<0.001; Figure 3a). Similarly, pre-treatment of NK (YT) cells with the granzyme B inhibitor 3,4-Dichloroisocoumarin (3,4-DCI), dramatically reduced the cytotoxicity when cultured with senescent cell targets (12 h) compared with the effector cells incubated in the presence of a vehicle control (Figure 3a). Similar results were obtained using a different NK cell line (NK-92, Supplementary Figure S1). Together these data indicate that the granule exocytosis cell death pathway, rather than the death receptor pathway, mediates NK-cell-facilitated elimination of senescent cells.
Figure 3.
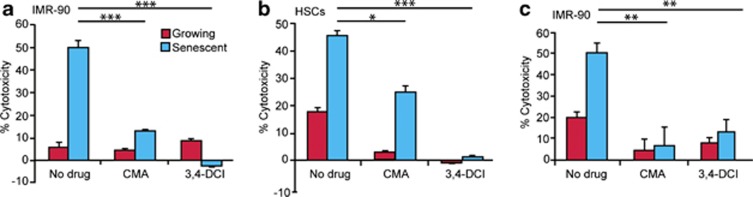
Granule exocytosis pathway is required for NK-cell-mediated killing of senescent cells. Senescent and growing IMR-90 fibroblasts (a, c) or HSCs (b) were co-incubated with YT cells for 12 h (a, b) or with primary NK cells for 2 h (c). Cytotoxicity assays were performed either in the presence of 100 nℳ granule exocytosis inhibitor, CMA or following pre-incubation of the YT or primary NK cells with 25 μℳ Granzyme B inhibitor 3,4-DCI. The graphs represent the average and the the s.e. of triplicate measurements from at least three independent experiments. *P<0.01, **P<0.001, ***P<0.0001.
To extend our studies to the use of primary cells, we co-cultured growing and senescent IMR-90 cells with primary human NK cells derived from whole blood and expanded in vitro as previously described.30 As primary NK cells are more potent than the YT NK cell line, the time of co-incubation was reduced to 2 h, where substantial levels of cytotoxicity were achieved (Figure 3). Despite this increased potency, treatment with either CMA or 3,4-DCI dramatically reduced NK-cell-mediated death of senescent IMR-90 cells (Figure 3c, P<0.001 for both treatments). The concordance between results from primary NK cells and two NK cell lines further underscores the importance of our models and the value of YT cells for studying NK-cell-mediated cytotoxicity.
A physiological setting in which clearance of senescent cells by NK cells has an important role is in liver fibrosis, where the NK-cells-mediated elimination of senescent activated HSCs limits the extent of fibrosis and facilitates repair.4 As occurs in senescent fibroblasts, both CMA and 3,4-DCI attenuated YT-mediated killing of senescent HSCs in 12 h cytotoxicity assays (Figure 3b, P<0.001 for both agents), suggesting a general role for this program in the immune surveillance of senescent cells.
Dcr2 attenuates NK-cell-mediated targeting of senescent cells
The fact that senescent cells are preferentially killed by granule exocytosis and are resistant to death receptor killing is intriguing in light of the senescence-specific upregulation of Dcr2—a decoy receptor that acts as a competitive inhibitor of death receptor signaling by death ligands, such as Fas or TRAIL.11, 31 Indeed, studies have identified and used high Dcr2 as a robust marker of senescence in tissues (for example, Collado et al. and Xue et al.11, 18) although its physiological contribution to the program is unknown. As expected, Dcr2 expression mRNA and protein was significantly increased in senescent IMR-90 cells and HSCs (P<0.0001 and P<0.05 for mRNA, respectively, Figures 4a-d, Supplementary Figure S4A). In principle, the observed increase in an inhibitor of death receptor signaling might explain the pathway bias towards granule exocytosis observed for NK-mediated killing of senescent cells.
Figure 4.

Dcr2 attenuates killing of senescent cells through the death receptor pathway. Dcr2 expression level in senescent and growing IMR-90 fibroblasts (a, b) and human HSCs (c, d) were evaluated by quantitative RT–PCR analysis (a, c) and immunoblotting (b, d). Dcr2-deficient senescent IMR-90 cells were incubated with either 10 or 100 ng/ml TRAIL and cytotoxicity was determined (e), and Dcr2 knockdown confirmed (f). Senescent IMR-90 cells with siDcr2 or siControl were incubated with YT cells for 12 h and cytotoxicity was determined (g). In the parallel approach IMR-90 cells were infected with short hairpin RNA (shRNA) targeting Dcr2 (shDcr2) or control shRNA targeting luciferase (shLuci) and induced to senescence by etoposide treatment. Dcr2 protein level was assessed by immunoblot (h). The cells were co-incubated for 12 h with YT cells and cytotoxicity was determined (i). The graphs represent the average and the s.e. of triplicate measurements from at least four independent experiments *P<0.05, **P<0.001, ***P<0.0001.
To test this hypothesis directly, we examined the requirement for Dcr2 in TRAIL-mediated cell death using small-interfering RNAs capable of potently suppressing Dcr2 protein in senescent IMR-90 cells (Figure 4f, Supplementary Figures S4B and C). Consistent with the ability of Dcr2 to modulate TRAIL-induced apoptosis, TRAIL induced twofold more killing in senescent IMR-90 cells transfected with siDcr2 comparing with siControl (Figure 4e, P<0.001, P<0.0001 for 10 and 100 ng/ml TRAIL, respectively). Moreover, senescent IMR-90 cells depleted of Dcr2 using either Dcr2 targeting small-interfering RNAs or a potent short hairpin RNA (Figure 4h) were more susceptible to killing by YT NK cells compared with cells transduced with control RNAi (Figure 4g, P<0.05, Figure 4i, P<0.05). Thus, NK cell cytotoxicity towards senescent cells is normally attenuated by the upregulation of Dcr2.
Granule exocytosis modulates liver fibrosis
If granule exocytosis is crucial for the immune surveillance of senescent cells in vivo, disruption of this program should alter the progression of liver fibrosis—a pathology where NK cell accumulation and senescent cell clearance has been shown to have a role.4 To examine this possibility, we induced fibrosis in wild-type (wt) and perforin knockout (Prf−/−) C57BL/6 mice by 12 consecutive bi-weekly injections of CCl4. To assess fibrosis progression, liver sections from treated and untreated mice were stained with Hematoxylin and Eosin (H&E) and the collagen-specific dye, Sirius red (Figure 5a). Fibrosis was not detected in untreated mice, nor were there any detectable differences in liver architecture between wt and Prf−/− mice. As expected, livers from CCl4-treated mice of both genotypes displayed a characteristic fibrotic histology.
Figure 5.

Perforin promotes senescent cell clearance and limits liver fibrosis. Perforin knockout (Prf−/−) and wt mice were treated with CCl4 to induce fibrosis. H&E and Sirius red staining show liver morphology and accumulation of fibrotic scar following the treatment (a). Morphometric analysis of Sirius red stained, entire liver sections (b). Expression of markers of activated HSCs, αSMA and Colagen1α, and senescence marker p15ink4b were tested by immunoblotting of whole-liver extracts (c). Four mice of each genotype are shown. SA-β-gal staining identified accumulation of senescent cells along the fibrotic scar areas in the livers (d). The presence of SA-β-gal-positive cells was quantified in the entire liver sections (e). At least five mice of each genotype were used for the analysis in B and E; **P<0.001, ***P<0.0001.
The extracellular matrix is deposited by the activated HSCs and their expansion is directly linked to the progression of fibrosis.32, 33 Deposits of extracellular matrix in fibrotic scars of these livers were identified by Sirius Red staining and quantified using morphometric analysis of whole-liver sections,4 and the relative abundance of activated HSCs was assessed by analyzing expression of the HSC markers αSMA (alpha smooth muscle actin) and Collagen1α in whole-liver lysates. Even in our short 12-week protocol, scarring was increased by 60% in Prf−/− mice comparing with wt (P<0.0001) (Figure 5a). Furthermore, immunoblotting showed an increased presence of both αSMA and Collagen1α in CCl4-treated Prf−/− livers compared with wt controls, indicating that the increase in fibrosis was also accompanied by enhanced expansion of the activated HSC compartment (Figure 5c). Therefore, perforin—and thus an intact granule exocytosis pathway—limits fibrosis progression.
To determine whether the granule exocytosis pathway influences the abundance of senescent cells, we examined wt and Prf−/− fibrotic livers for the senescent markers SA-β-gal, p15ink4b and Dcr2 following CCl4 treatment. As an additional control, we also examined these markers in mice deficient for senescent cells owing to deletion of p53 and Ink4a/Arf, a setting where enhanced fibrosis arises owing to defects in senescence induction.4 Consistent with our previous report, SA-β-gal-positive cells were found predominantly in the livers of CCl4-treated mice and in areas adjacent or within the fibrotic scars4 (Figure 5d). Using an imaging approach to quantify the tissue area occupied by SA-β-gal-positive cells (see Materials and methods), we noted a significant increase in senescent cells retained in Prf−/− livers compared with controls (Figure 5e, P<0.001). Similarly, Prf−/− livers from CCl4-treated mice showed enhanced expression of p15Ink4a and Dcr2 relative to treated livers from wt controls (Figure 5c, Supplementary Figures S5 and S6). By contrast, few SA-β-gal-positive cells and low p15Ink4b levels were observed in livers from CCl4-treated p53−/−;Ink4a/Arf−/− mice (Figure 5c, Supplementary Figure S5), even though these livers also showed more activated HSCs and fibrosis (Supplementary Figure S4).4 Together these data indicate that excessive liver fibrosis can arise from defects in the senescence of activated stellate cells or, as shown here, from the failure to eliminate these cells once they become senescent.
Discussion
NK-cell-mediated clearance of senescent cells is one component of the coordinated process whereby cellular senescence limits the extent of liver fibrosis and facilitates wound repair.4, 18 Recent studies also suggest that senescent cell clearance by immune cells promotes tumor regression in established tumors.18 Our results demonstrate that the granule exocytosis pathway, but not the death receptor pathway, is necessary for the specific killing of senescent fibroblasts and stellate cells by NK cells and participates in the clearance of senescent activated HSCs to limit liver fibrosis. Therefore, NK-cell-mediated cytotoxicity through granule exocytosis contributes to immune surveillance of senescent cells in vitro and in vivo.
Several types of cytotoxic lymphocytes can use granule exocytosis pathway to eliminate their target cells.21 For example, invariant NK T cells, which are abundant in the liver, limit the initial stages of liver fibrosis, presumably through targeting activated stellate cells.34 However, the functional contribution of invariant NK T cells for the clearance of senescent cells has not been examined. CD8+ cytotoxic T lymphocytes also rely on granule exocytosis for cell-mediated cytotoxicity23 and defects in their action could contribute to the enhanced phenotypes we observe in Prf−/− mice. Still, NK cells, which accumulate in the liver during fibrosis, are sufficient to eliminate senescent cells in vitro, and the primary NK cell populations used in these studies were 99% of CD56+ cells and devoid of T cells.30 However, we do not exclude the possibility that other immune cells contribute to the surveillance of senescent cells in tissues and, indeed, a recent report suggests that clearance of premalignant senescent hepatocytes is mediated by CD4 T cells and macrophages.8
In addition to the granule exocytosis pathway, most cytotoxic lymphocytes engage the death receptor pathway to eliminate target cells. This pathway is widely used by NK cells in the liver.21 NK cell express high levels of the death receptor ligand TRAIL upon activation with IL-2,26 are suggested to participate in the surveillance of the HSCs,35 and protect against tumor development following chemical carcinogenesis.36 Given this, we were surprised that death-receptor-mediated cytotoxicity was dispensable for the immune surveillance of senescent cells. Consistent with these findings, an anti-TRAIL antibody failed to inhibit immune system-mediated tumor clearance following p53 restoration in a liver carcinoma model18 (W Xue and SWL, unpublished data). Of course, we cannot rule out the possibility that death receptor pathways contribute to senescent cell clearance in other settings.
Why does granule exocytosis, and not the death-receptor signaling, mediate NK-cell surveillance of senescent cells? Mechanistically, this appears partly because of the accumulation of Dcr2 during senescence, which occurs in fibroblasts, certain epithelial cells11, 18 and, as shown here, also senescent activated HSCs. Dcr2 can bind death-receptor ligands, with higher affinity to TRAIL, but as it lacks the activation domain it prevents downstream signaling through the death receptor pathway31, 37 and, therefore, can protect senescent cells from death-receptor-ligand-mediated killing. Another decoy receptor, Dcr3, has higher affinity to FASL.38 However, in contrast to Dcr2, Dcr3 is a secreted receptor and is much less likely to have a role in direct interaction between senescent and NK cells. Although previously considered merely a senescence marker, our results establish a functional role for Dcr2 in protecting senescent cells from cytotoxicity through the death receptor pathway induced by NK cells and possibly other cells as well. The biological rationale for this regulation remains unclear, but may serve to prevent autoimmunity following short-term tissue damage.
In addition to blocking the death receptor pathway, senescent cells may also stimulate NK cells to induce the perforin-mediated killing. Senescent cells upregulate expression of several ligands of NK-cell receptor NKG2D4, 39 and ICAM-1, the ligand of NK-cell receptor LFA-1.40 Studies suggest that activation of the NKG2D receptor induces granule exocytosis to eliminate cancer cells, a process that might be reinforced by signaling from LFA-1.41 In this manner, ligands upregulated in senescent cells might activate multiple NK-cell receptors to trigger granule exocytosis.
The role of granule exocytosis in the surveillance of senescent cells has important ramifications for understanding and treating wound healing and cancer. Indeed, we show that the immune clearance of senescent activated HSCs has a significant impact on the pathophysiology of liver fibrosis in which the granule exocytosis pathway has been previously implicated.42, 43 Beyond the liver, immune surveillance of senescent cells might have a significant role in other fibrosis-related pathological conditions such as lung fibrosis, atopic dermatitis and atherosclerosis, where granule exocytosis has also been implicated in disease pathology.44, 45, 46
Still, the most prevalent conditions where senescence has been studied to date involve cancer and aging.3, 9 Senescent cells accumulate with age and contribute to functional decline of multiple tissues7, 9 while perforin-mediated granule exocytosis diminishes at that time.47, 48 Separate studies suggest that the integrity of the granule exocytosis pathway can modulate a variety of cancer phenotypes.49, 50 Though definitive proof will require further testing, we speculate that the granule exocytosis pathway contributes to immune surveillance of senescent cells in each of these conditions. In principle, pharmacological modulation of this pathway, as has been recently described using IL21,51 might increase the clearance of senescent cells from premalignant, damaged or aged tissues to limit carcinogenesis and the decline in tissue function accompanying the accumulation of senescent cells.
Materials and methods
Tissue culture and cytotoxicity assays
Human diploid fibroblasts IMR-90 (ATCC, Manassas, VA, USA) and primary human hepatic myofibroblasts (activated HSCs) (Dominion Pharmakine, Derio – Bizkaia, Spain) were grown in standard conditions. Senescence was induced by etoposide (Sigma, St Louis, MO, USA) treatment as described previously.4 In vitro cytotoxicity assays using the NK cell line, YT, were performed for 12 h as described previously.4 The long incubation period enables efficient execution of both granule exocytosis and death-receptor-mediated cytotoxicity.
For the cytotoxicity assays performed with primary human NK cells, target cells were plated in 12-well plates at 4 × 104 cells per well; 1 × 105 NK cells (more than 99% of CD56+; CD3− (Hanna et al.30) (gift from O Mandelboim, The Hebrew University Hadassah Medical School, Jerusalem, Israel.) were subsequently added to each well. Following 2 h of co-incubation, primary NK cells were washed gently and the cytotoxicity was determined based on quantification of remaining adherent cells.
To inhibit the granule exocytosis or granzyme B the cytotoxicity assays were performed in the presence of 100 nℳ CMA or the effector cells were pre-incubated with 25 μℳ of 3,4-Dichloroisocoumarin (3,4-DCI), respectively, (both Sigma). The caspase inhibitors Z-VAD-FMK and Z-IEDT-FMK (MBL, Woburn, MA, USA) were used at separate assays at a concentration of 10 μℳ.
In effector cell independent experiments, the target cells were incubated for 12 h with 2 or 10 ng/ml FasL in the presence of 10 μg/ml of a cross-linking antibody or for 24 h with 10 or 100 ng/ml TRAIL (both R&D Systems, Minneapolis, MN, USA). The cytotoxicity was determined using staining of the remaining adherent cells.
Mice
The Prf−/− mice were obtained from Jackson Laboratories (Bar Harbor, ME, USA) and C57Bl6 mice served as wt controls. For fibrosis induction, the mice were treated twice a week, with i.p. injection of 1 ml/kg CCl4, for 6 weeks as described.4
Formalin-fixed paraffin-embedded tissue was sectioned and stained either with hematoxylin–eosin for routine examination, or with Sirius red for visualization of fibrotic deposition. The relative fibrotic area was calculated based on Sirius red staining as previously described.4
Detection of SA-β-gal activity was performed as described previously.4 For quantification of the staining in the mouse tissue, images of whole-liver sections were taken from multiple slides (derived from at least 6 mice of each genotype) using 3D Histech Digital Microscopy scanner (3DHISTECH Kft., Budapest, Hungary). The images were quantified using Image-Pro software package (Media Cybernetics, Bethesda, MD, USA).
Detection and modification of gene expression
Detection of protein expression by immunoblotting was performed using anti-αSMA (DakoCytomation, Glostrup, Denmark), anti-p15 (Cell Signaling Technology, Beverly, MA, USA), anti-Collagen1α (Rockland, PA, USA), anti-β Actin (AC-15, Sigma), anti-DCR2 (Assay designs, Ann Arbor, MI, USA).
For quantitative RT–PCR total RNA was isolated using NucleoSpin kit (Macherey Nagel, Düren, Germany) and 1 μg was reverse transcribed using the RevertAid H Minus First Strand cDNA synthesis Kit (Fermentas, Glen Burnie, MD, USA). The cDNA samples were amplified using Fast SYBR Green Master Mix in StepOnePlus Real-Time PCR System (both Applied Biosystems, Foster City, CA, USA). The relative expression of Dcr2 was normalized using the expression levels of either β-Actin or GAPDH. Primer sequences are available upon request.
ON-TARGETplus SMARTpool small-interfering RNA targeting Dcr2 and the nontargeting (control) pool were transfected into senescent IMR-90 cells with Dharmafect 1 reagent (all from Dharmacon, Lafayette, CO, USA). The experiments using retroviral vectors (pMLP (targeting human Dcr2) and pMLP (targeting Luciferase)) were performed as described.14
Acknowledgments
We gratefully acknowledge O Mandelboim from Hadassah Medical Center for providing us with primary human NK cells; D Sabah-Israel for editorial suggestions; members of the Lowe and Krizhanovsky laboratories for stimulating discussions; K Lehet, L Bianco and the CSHL animal facility for help with animals; T Berkutzki from the Histology Facility at the Weizmann Institute for assistance with histology. This work was supported by a Binational Israel-USA Science foundation (VK and SWL) and grant AG16379 from the National Institutes of Health (SWL). VK was supported by Israel Science Foundation, Abisch-Frenkel foundation, Simms/Mann Family foundation and Marie Curie RG grant from EU FP7. VK is an incumbent of The Karl and Frances Korn Career Development Chair. SWL is a Howard Hughes Medical Institute investigator and Geoffrey Beene Chair for Cancer Biology (MSKCC).
The authors declare no conflict of interest.
Footnotes
Supplementary Information accompanies the paper on the Oncogene website (http://www.nature.com/onc)
Supplementary Material
References
- Serrano M, Lin AW, McCurrach ME, Beach D, Lowe SW. Oncogenic ras provokes premature cell senescence associated with accumulation of p53 and p16INK4a. Cell. 1997;88:593–602. doi: 10.1016/s0092-8674(00)81902-9. [DOI] [PubMed] [Google Scholar]
- Schmitt CA, Fridman JS, Yang M, Lee S, Baranov E, Hoffman RM, et al. A senescence program controlled by p53 and p16INK4a contributes to the outcome of cancer therapy. Cell. 2002;109:335–346. doi: 10.1016/s0092-8674(02)00734-1. [DOI] [PubMed] [Google Scholar]
- Narita M, Lowe SW. Senescence comes of age. Nat Med. 2005;11:920–922. doi: 10.1038/nm0905-920. [DOI] [PubMed] [Google Scholar]
- Krizhanovsky V, Yon M, Dickins RA, Hearn S, Simon J, Miething C, et al. Senescence of activated stellate cells limits liver fibrosis. Cell. 2008;134:657–667. doi: 10.1016/j.cell.2008.06.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jun JI, Lau LF. The matricellular protein CCN1 induces fibroblast senescence and restricts fibrosis in cutaneous wound healing. Nat Cell Biol. 2010;12:676–685. doi: 10.1038/ncb2070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitiyage GN, Slijepcevic P, Gabrani A, Chianea YG, Lim KP, Prime SS, et al. Senescent mesenchymal cells accumulate in human fibrosis by a telomere-independent mechanism and ameliorate fibrosis through matrix metalloproteinases. J Pathol. 2011;223:604–617. doi: 10.1002/path.2839. [DOI] [PubMed] [Google Scholar]
- Baker DJ, Wijshake T, Tchkonia T, LeBrasseur NK, Childs BG, van de Sluis B, et al. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature. 2011;479:232–236. doi: 10.1038/nature10600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang TW, Yevsa T, Woller N, Hoenicke L, Wuestefeld T, Dauch D, et al. Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature. 2011;479:547–551. doi: 10.1038/nature10599. [DOI] [PubMed] [Google Scholar]
- Dimri GP, Lee X, Basile G, Acosta M, Scott G, Roskelley C, et al. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc Natl Acad Sci USA. 1995;92:9363–9367. doi: 10.1073/pnas.92.20.9363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narita M, Nunez S, Heard E, Narita M, Lin AW, Hearn SA, et al. Rb-mediated heterochromatin formation and silencing of E2F target genes during cellular senescence. Cell. 2003;113:703–716. doi: 10.1016/s0092-8674(03)00401-x. [DOI] [PubMed] [Google Scholar]
- Collado M, Gil J, Efeyan A, Guerra C, Schuhmacher AJ, Barradas M, et al. Tumour biology: senescence in premalignant tumours. Nature. 2005;436:642. doi: 10.1038/436642a. [DOI] [PubMed] [Google Scholar]
- Krimpenfort P, Ijpenberg A, Song JY, van der Valk M, Nawijn M, Zevenhoven J, et al. p15Ink4b is a critical tumour suppressor in the absence of p16Ink4a. Nature. 2007;448:943–946. doi: 10.1038/nature06084. [DOI] [PubMed] [Google Scholar]
- Krizhanovsky V, Xue W, Zender L, Yon M, Hernando E, Lowe SW. Implications of cellular senescence in tissue damage response, tumor suppression, and stem cell biology. Cold Spring Harb Symp Quant Biol. 2008;73:513–522. doi: 10.1101/sqb.2008.73.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chicas A, Wang X, Zhang C, McCurrach M, Zhao Z, Mert O, et al. Dissecting the unique role of the retinoblastoma tumor suppressor during cellular senescence. Cancer Cell. 2010;17:376–387. doi: 10.1016/j.ccr.2010.01.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuilman T, Michaloglou C, Vredeveld LC, Douma S, van Doorn R, Desmet CJ, et al. Oncogene-induced senescence relayed by an interleukin-dependent inflammatory network. Cell. 2008;133:1019–1031. doi: 10.1016/j.cell.2008.03.039. [DOI] [PubMed] [Google Scholar]
- Acosta JC, O'Loghlen A, Banito A, Guijarro MV, Augert A, Raguz S, et al. Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell. 2008;133:1006–1018. doi: 10.1016/j.cell.2008.03.038. [DOI] [PubMed] [Google Scholar]
- Kortlever RM, Higgins PJ, Bernards R. Plasminogen activator inhibitor-1 is a critical downstream target of p53 in the induction of replicative senescence. Nat Cell Biol. 2006;8:877–884. doi: 10.1038/ncb1448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue W, Zender L, Miething C, Dickins RA, Hernando E, Krizhanovsky V, et al. Senescence and tumour clearance is triggered by p53 restoration in murine liver carcinomas. Nature. 2007;445:656–660. doi: 10.1038/nature05529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bottino C, Castriconi R, Moretta L, Moretta A. Cellular ligands of activating NK receptors. Trends Immunol. 2005;26:221–226. doi: 10.1016/j.it.2005.02.007. [DOI] [PubMed] [Google Scholar]
- Arnon TI, Markel G, Mandelboim O. Tumor and viral recognition by natural killer cells receptors. Semin Cancer Biol. 2006;16:348–358. doi: 10.1016/j.semcancer.2006.07.005. [DOI] [PubMed] [Google Scholar]
- Chan CW, Housseau F. The 'kiss of death' by dendritic cells to cancer cells. Cell Death Differ. 2008;15:58–69. doi: 10.1038/sj.cdd.4402235. [DOI] [PubMed] [Google Scholar]
- Wallach D, Varfolomeev EE, Malinin NL, Goltsev YV, Kovalenko AV, Boldin MP. Tumor necrosis factor receptor and Fas signaling mechanisms. Annu Rev Immunol. 1999;17:331–367. doi: 10.1146/annurev.immunol.17.1.331. [DOI] [PubMed] [Google Scholar]
- Chowdhury D, Lieberman J. Death by a thousand cuts: granzyme pathways of programmed cell death. Annu Rev Immunol. 2008;26:389–420. doi: 10.1146/annurev.immunol.26.021607.090404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cullen SP, Martin SJ. Mechanisms of granule-dependent killing. Cell Death Differ. 2008;15:251–262. doi: 10.1038/sj.cdd.4402244. [DOI] [PubMed] [Google Scholar]
- Kagi D, Ledermann B, Burki K, Seiler P, Odermatt B, Olsen KJ, et al. Cytotoxicity mediated by T cells and natural killer cells is greatly impaired in perforin-deficient mice. Nature. 1994;369:31–37. doi: 10.1038/369031a0. [DOI] [PubMed] [Google Scholar]
- Ishiyama K, Ohdan H, Ohira M, Mitsuta H, Arihiro K, Asahara T. Difference in cytotoxicity against hepatocellular carcinoma between liver and periphery natural killer cells in humans. Hepatology. 2006;43:362–372. doi: 10.1002/hep.21035. [DOI] [PubMed] [Google Scholar]
- Montel AH, Bochan MR, Hobbs JA, Lynch DH, Brahmi Z. Fas involvement in cytotoxicity mediated by human NK cells. Cell Immunol. 1995;166:236–246. doi: 10.1006/cimm.1995.9974. [DOI] [PubMed] [Google Scholar]
- Muta H, Boise LH, Fang L, Podack ER. CD30 signals integrate expression of cytotoxic effector molecules, lymphocyte trafficking signals, and signals for proliferation and apoptosis. J Immunol. 2000;165:5105–5111. doi: 10.4049/jimmunol.165.9.5105. [DOI] [PubMed] [Google Scholar]
- Chahroudi A, Silvestri G, Feinberg MB. Measuring T cell-mediated cytotoxicity using fluorogenic caspase substrates. Methods. 2003;31:120–126. doi: 10.1016/s1046-2023(03)00121-x. [DOI] [PubMed] [Google Scholar]
- Hanna J, Wald O, Goldman-Wohl D, Prus D, Markel G, Gazit R, et al. CXCL12 expression by invasive trophoblasts induces the specific migration of CD16- human natural killer cells. Blood. 2003;102:1569–1577. doi: 10.1182/blood-2003-02-0517. [DOI] [PubMed] [Google Scholar]
- Marsters SA, Sheridan JP, Pitti RM, Huang A, Skubatch M, Baldwin D, et al. A novel receptor for Apo2L/TRAIL contains a truncated death domain. Curr Biol. 1997;7:1003–1006. doi: 10.1016/s0960-9822(06)00422-2. [DOI] [PubMed] [Google Scholar]
- Bataller R, Brenner DA. Liver fibrosis. J Clin Invest. 2005;115:209–218. doi: 10.1172/JCI24282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo J, Friedman SL. Hepatic fibrogenesis. Semin Liver Dis. 2007;27:413–426. doi: 10.1055/s-2007-991517. [DOI] [PubMed] [Google Scholar]
- Park O, Jeong WI, Wang L, Wang H, Lian ZX, Gershwin ME, et al. Diverse roles of invariant natural killer T cells in liver injury and fibrosis induced by carbon tetrachloride. Hepatology. 2009;49:1683–1694. doi: 10.1002/hep.22813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radaeva S, Sun R, Jaruga B, Nguyen VT, Tian Z, Gao B. Natural killer cells ameliorate liver fibrosis by killing activated stellate cells in NKG2D-dependent and tumor necrosis factor-related apoptosis-inducing ligand-dependent manners. Gastroenterology. 2006;130:435–452. doi: 10.1053/j.gastro.2005.10.055. [DOI] [PubMed] [Google Scholar]
- Takeda K, Cretney E, Hayakawa Y, Ota T, Akiba H, Ogasawara K, et al. TRAIL identifies immature natural killer cells in newborn mice and adult mouse liver. Blood. 2005;105:2082–2089. doi: 10.1182/blood-2004-08-3262. [DOI] [PubMed] [Google Scholar]
- Pinkoski MJ, Brunner T, Green DR, Lin T. Fas and Fas ligand in gut and liver. Am J Physiol Gastrointest Liver Physiol. 2000;278:G354–G366. doi: 10.1152/ajpgi.2000.278.3.G354. [DOI] [PubMed] [Google Scholar]
- Colucci S, Brunetti G, Mori G, Oranger A, Centonze M, Mori C, et al. Soluble decoy receptor 3 modulates the survival and formation of osteoclasts from multiple myeloma bone disease patients. Leukemia. 2009;23:2139–2146. doi: 10.1038/leu.2009.136. [DOI] [PubMed] [Google Scholar]
- Soriani A, Zingoni A, Cerboni C, Iannitto ML, Ricciardi MR, Di Gialleonardo V, et al. ATM-ATR-dependent up-regulation of DNAM-1 and NKG2D ligands on multiple myeloma cells by therapeutic agents results in enhanced NK-cell susceptibility and is associated with a senescent phenotype. Blood. 2009;113:3503–3511. doi: 10.1182/blood-2008-08-173914. [DOI] [PubMed] [Google Scholar]
- Chien Y, Scuoppo C, Wang X, Fang X, Balgley B, Bolden JE, et al. Control of the senescence-associated secretory phenotype by NF-{kappa}B promotes senescence and enhances chemosensitivity. Genes Dev. 2011;25:2125–2136. doi: 10.1101/gad.17276711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayakawa Y, Smyth MJ. NKG2D and cytotoxic effector function in tumor immune surveillance. Semin Immunol. 2006;18:176–185. doi: 10.1016/j.smim.2006.03.005. [DOI] [PubMed] [Google Scholar]
- Muhanna N, Horani A, Doron S, Safadi R. Lymphocyte-hepatic stellate cell proximity suggests a direct interaction. Clin Exp Immunol. 2007;148:338–347. doi: 10.1111/j.1365-2249.2007.03353.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melhem A, Muhanna N, Bishara A, Alvarez CE, Ilan Y, Bishara T, et al. Anti-fibrotic activity of NK cells in experimental liver injury through killing of activated HSC. J Hepatol. 2006;45:60–71. doi: 10.1016/j.jhep.2005.12.025. [DOI] [PubMed] [Google Scholar]
- Yawalkar N, Schmid S, Braathen LR, Pichler WJ. Perforin and granzyme B may contribute to skin inflammation in atopic dermatitis and psoriasis. Br J Dermatol. 2001;144:1133–1139. doi: 10.1046/j.1365-2133.2001.04222.x. [DOI] [PubMed] [Google Scholar]
- Choy JC, McDonald PC, Suarez AC, Hung VH, Wilson JE, McManus BM, et al. Granzyme B in atherosclerosis and transplant vascular disease: association with cell death and atherosclerotic disease severity. Mod Pathol. 2003;16:460–470. doi: 10.1097/01.MP.0000067424.12280.BC. [DOI] [PubMed] [Google Scholar]
- Miyazaki H, Kuwano K, Yoshida K, Maeyama T, Yoshimi M, Fujita M, et al. The perforin mediated apoptotic pathway in lung injury and fibrosis. J Clin Pathol. 2004;57:1292–1298. doi: 10.1136/jcp.2003.015495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ogata K, Yokose N, Tamura H, An E, Nakamura K, Dan K, et al. Natural killer cells in the late decades of human life. Clin Immunol Immunopathol. 1997;84:269–275. doi: 10.1006/clin.1997.4401. [DOI] [PubMed] [Google Scholar]
- Rukavina D, Laskarin G, Rubesa G, Strbo N, Bedenicki I, Manestar D, et al. Age-related decline of perforin expression in human cytotoxic T lymphocytes and natural killer cells. Blood. 1998;92:2410–2420. [PubMed] [Google Scholar]
- van den Broek ME, Kagi D, Ossendorp F, Toes R, Vamvakas S, Lutz WK, et al. Decreased tumor surveillance in perforin-deficient mice. J Exp Med. 1996;184:1781–1790. doi: 10.1084/jem.184.5.1781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Street SE, Cretney E, Smyth MJ. Perforin and interferon-gamma activities independently control tumor initiation, growth, and metastasis. Blood. 2001;97:192–197. doi: 10.1182/blood.v97.1.192. [DOI] [PubMed] [Google Scholar]
- Davis ID, Skrumsager BK, Cebon J, Nicholaou T, Barlow JW, Moller NP, et al. An open-label, two-arm, phase I trial of recombinant human interleukin-21 in patients with metastatic melanoma. Clin Cancer Res. 2007;13:3630–3636. doi: 10.1158/1078-0432.CCR-07-0410. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.