

Abstract
Epithelial ovarian cancer (EOC) accounts for the most gynecological malignancy-associated deaths in the United States. Enhancer of zeste homolog 2 (EZH2), which silences gene expression through generating trimethylation on lysine 27 residue of histone H3 (H3K27Me3), is often overexpressed in EOC and has been suggested as a therapeutic target. However, the mechanism underlying EZH2 overexpression in EOC is unknown. Here we show that EZH2 is upregulated at the transcription level, and two CCAAT boxes in the proximal regions of the human EZH2 gene promoter are critical for its transcription in EOC cells. Indeed, NF-YA, the regulatory subunit of the CCAAT binding transcription factor NF-Y, is expressed at higher levels in human EOCs compared with primary human ovarian surface epithelial (HOSE) cells. In addition, there is a positive correlation between expression of NF-YA and EZH2 in EOCs. Notably, high NF-YA expression predicts shorter overall survival in EOC patients. The association of NF-YA with the promoter of the human EZH2 gene is enhanced in human EOC cells compared with primary HOSE cells. Significantly, knockdown of NF-YA downregulates EZH2, decreases H3K27Me3 levels, and suppresses the growth of human EOC cells both in vitro and in a xenograft mouse model. Notably, NF-YA knockdown induces apoptosis of EOC cells and ectopic EZH2 expression partially rescues apoptosis induced by NF-YA knockdown. Together, these data reveal that NF-Y is a key regulator of EZH2 expression and is required for EOC cell proliferation, thus representing a novel target for developing EOC therapeutics.
Introduction
Epithelial ovarian cancer (EOC) accounts for more deaths than any other gynecological malignancy in the United States (1). EOCs are classified into distinct histological types including serous, mucinous, endometrioid and clear cell. The most common histology of EOC is serous (~60% of all cancers) (2). Recently, an alternative classification has been proposed, in which EOC is broadly divided into two types (3). Type I EOC includes mucinous, low-grade serous, low-grade endometrioid and clear cell carcinomas, and type II EOC includes high-grade serous carcinomas, which is the most lethal histosubtype (3).
Enhancer of zeste homolog 2 (EZH2) is a histone methyltransferase that mediates gene silencing by catalyzing trimethylation of lysine 27 residue of histone H3 (H3K27Me3) (4). EZH2 is often expressed at higher levels in human EOCs, and its expression positively correlates with cell proliferation (5). Further underscoring the importance of EZH2 in EOC, EZH2 knockdown triggers apoptosis of human EOC cells (5). These findings identify EZH2 as a putative target for developing EOC therapeutics. Thus, it is important to elucidate the mechanism underlying EZH2 upregulation in EOCs to gain insights into the biology of the disease.
Gene amplification contributes to EZH2 upregulation in several types of cancers, including malignancies of the breast and prostate (6). However, based on the newly released the cancer genome atlas (TCGA) ovarian database (http://tcga-data.nci.nih.gov/) (7), EZH2 gene amplification (>4 copy) is rare (~2%) in EOC, suggesting that additional mechanisms make more significant contributions to EZH2 upregulation in EOC cells.
NF-Y is a transcription factor that specifically binds to the CCAAT consensus site (8). NF-Y is a heterotrimer, consisting of three subunits NF-YA, NF-YB and NF-YC. NF-YA is the regulatory subunit that is differentially expressed, while NF-YB and NF-YC are constitutively expressed (9–11). As a result of differential splicing, NF-YA has two isoforms, namely short and long (12). Both isoforms bind DNA and are equivalently active in transcriptional activation (9). NF-Y acts as a transcriptional activator by recruiting p300 histone acetyltransferase, which promotes gene expression by generating acetylation epigenetic marker on histone H3 (13, 14). Clinically, upregulated NF-Y target genes convey a poor prognosis in multiple cancers including those of the breast and lung (15). However, the role of NF-Y in EOC has never been investigated.
Here we demonstrate that EZH2 is upregulated at the transcriptional level, and two CCAAT sites at the proximal region of the human EZH2 gene promoter play a key role in regulating its transcription. NF-YA, the regulatory subunit of NF-Y transcription factor that binds to CCAAT sites, is upregulated in human EOCs compared with normal human ovarian surface epithelial (HOSE) cells. In addition, ectopic NF-YA upregulates EZH2 in normal HOSE cells. Importantly, there is a positive correlation between expression of NF-YA and EZH2 in human EOCs and a high level of NF-YA predicts poor overall survival in EOC patients. Chromatin immunoprecipitation analysis revealed that the interaction between NF-YA and the promoter of human EZH2 gene is enhanced in human EOC cells compared with normal HOSE cells. Knockdown of NF-YA downregulates EZH2, decreases the levels of H3K27Me3 and suppresses the growth of human EOC cells both in vitro and in a xenograft mouse model. Mechanistically, we find that NF-YA knockdown triggers apoptosis of human EOC cells and ectopic EZH2 expression partially rescues the apoptosis induced by NF-YA knockdown. Together, these data show that NF-Y plays a key role in regulating EZH2 transcription and is essential for proliferation of human EOC cells.
Material and Methods
Cell culture
Normal HOSE cells were cultured as previously described (5). Human EOC cell lines (PEO1, SKOV3 and OVCAR5) were cultured according to American Type Culture Collection (ATCC) in RPMI-1640 media supplemented with 10% FBS and as previously described (5). EOC cell line identification was further confirmed by DNA Diagnostic Center.
qRT-PCR and immunoblotting
RNA from cultured normal HOSE cells or human EOC cell lines was isolated using Trizol (Invitrogen) according to the manufacturer’s instructions. For qRT-PCR, Trizol-isolated RNA was further purified using the RNeasy kit (Qiagen) following the manufacturer’s instructions. Primers used for qRT-PCR were purchased from Applied Biosystems. Expression of the housekeeping gene β-2-microglobulin mRNA was used to normalize mRNA expression. The antibodies used for immunoblotting analysis were from the indicated suppliers as following: anti-NF-YA (Santa Cruz), anti-EZH2 (BD Bioscience), anti-H3K27Me3 (Cell Signaling), anti-H3K9Me3 (Abcam), anti-cleaved PARP p85 fragment (Promega), anti-cleaved caspase-3 (Cell signaling) and anti-histone H3 (Millipore).
Luciferase promoter activity and mutagenesis analysis
Indicated fragments of the proximal human EZH2 promoter were generated by PCR and cloned into the pGL2 basic luciferase reporter plasmid (Promega) following standard molecular cloning protocols. The NF-YA binding site single base pair substitution mutant was generated by PCR using standard molecular cloning protocols. Luciferase reporter plasmids were transfected into SKOV3, PEO1, OVCAR5 EOC cells or normal HOSE cells using Lipofectamine2000 (Invitrogen) following the manufacturer’s instructions. A luminescent β-galactosidase plasmid (Clontech) was included as an internal control to normalize the variation in transfection efficiency.
Chromatin immunoprecipitation (ChIP) analysis
ChIP was performed using a kit from Millipore following the manufacturer’s instructions. Briefly, SKOV3, PEO1, OVCAR5 EOC cells and normal HOSE cells were fixed with 1% formaldehyde. After cell lysis, genomic DNA was sheared into 300–500 bp fragments by sonication. Sheared chromatin was incubated with anti-NF-YA (Santa Cruz), anti-p300 (Santa Cruz), anti-histone H3 (Millipore), or anti-acetylated histone H3 (Millipore) antibodies overnight at 4°C. After reversing the cross-link, DNA was extracted using the phenol-chloroform method. The immunoprecipitated DNA then was amplified by PCR. The primers used for PCR amplification were: for distal EZH2 gene promoter: forward primer: 5 ′-GTCGGGAGTTCGAGAC-3′ and reverse primer: 5 ′-GTCGGCTCAGCTGTG-3′; and for proximal EZH2 gene promoter: forward primer: 5′-CTGTGATTGGACGGGC-3′ and reverse primer: 5′-ACTCGCGTTGTTCCC-3′
Inducible EZH2 overexpression, short hairpin RNA, lentivirus packaging and infection
Inducible EZH2 expression was achieved by using the Retro-X™ Tet-On® Advanced Inducible Expression System (Invitrogen) following the manufacturer’s instructions. Lentivirus packaging was performed using the Virapower kit (Invitrogen) according to the manufacturer’s instructions. Briefly, cells at 40%–50% confluence were infected with lentivirus-expressing shRNA to the human NF-YA gene (shNF-YA) or vector control. The infected cells were drug-selected with 3 μg/ml puromycin. The mature sense sequences of the three individual shNF-YA are: 5′-CCATCATGCAAGTACCTGTTT-3′, 5′-TTCTGTCCTGTAGTAAAGGGC-3′ and 5′-CGAGCTAAACTAGAGGCAGAA-3′. pLV-CMV-H4-puro lentiviral vector (kindly provided by Dr. Alexey Ivanov, West Virginia University, Morgantown, WV) was used to produce the lentivirus expressing NF-YA.
Anchorage-independent soft agar assay
Soft agar assays were performed as previously described (5). Briefly, 2500 cells were resuspended in 0.35% low melting point agarose dissolved in RPMI1640 medium supplemented with 10% FBS, and inoculated on top of 0.6% low melting point agarose base in 6-well plates. After 2 weeks in culture, the plates were stained with 0.005% crystal violet, and the number of colonies was counted.
Annexin V staining for detecting apoptotic cells (Guava assay)
Phosphatidylserine externalization was detected using an Annexin V staining kit (Millipore) following the manufacturer’s instructions. Annexin V positive cells were detected using the Guava® system and analyzed with the Guava Nexin software Module (Millipore).
Xenograft tumorigenesis assay
The protocol was approved by the Institutional Animal Care and Use Committee (IACUC). A total of 6×105 cells in PBS (pH 7.3) were injected subcutaneously into 6-week-old female nude athymic mice. The mice were sacrificed 30 days post-inoculation. Tumor size was measured, and the tumor volume (mm3) was calculated using the following formula: tumor volume [mm3] = a2 × b/2, where a is the smaller diameter and b is the larger diameter.
Data Sets
Gene expression microarray data sets for 53 cases of laser capture and microdissected high-grade serous EOC and 10 individual isolations of normal HOSE cells were obtained from Gene Expression Omnibus (GEO) (http://www.ncbi.nlm.nih.gov/geo/) (GEO accession number: GSE18521). Gene expression data from 24 cases of laser capture microdissected primary non-malignant fallopian tube epithelium and 13 cases of high-grade serous EOC (GEO accession number: GSE10971).
Statistical analysis
Quantitative data are expressed as mean ± SD unless otherwise stated. Analysis of variance (ANOVA) with Fisher’s Least Significant Difference (LSD) was used to identify significant differences in multiple comparisons. Spearman’s test was used to measure statistical dependence between EZH2 mRNA levels and NF-YA mRNA levels. For all statistical analyses, the level of significance was set at 0.05.
Results
EZH2 is upregulated at the transcriptional level and two CCAAT sites in the proximal region of the human EZH2 gene promoter are critical for its transcription in human EOC cells
It has been shown that EZH2 protein is overexpressed in EOC cells (5). To determine whether the upregulation occurs at the transcriptional level, we examined the mRNA level of EZH2 in human EOC cell lines and in normal HOSE cells by qRT-PCR. Compared with normal HOSE cells, the EZH2 mRNA level was significantly higher in human EOC cell lines (Figure 1A). This suggests that EZH2 is upregulated at the transcriptional level in human EOC cells. In agreement, a luciferase reporter driven by a 1152 bp (−1011 – +141bp) proximal fragment of the human EZH2 gene promoter was highly active in human EOC cell lines compared with normal HOSE cells (Figure S1A). E2F transcriptional factor is known to play a role in regulating the transcription of the human EZH2 gene in human EOC cells (6). In addition, Elk-1 transcription factor has also been implicated in regulating EZH2 gene transcription (16). Notably, deletion of both E2F and Elk-1 binding sites only caused a ~2-fold decrease in EZH2 gene promoter activity in human EOC cells (Figure 1B and S1B–C). However, the EZH2 mRNA levels are 30–50 times higher in human EOC cell lines compared with normal HOSE cells (Figure 1A) and the EZH2 gene promoter activity shows the similar fold changes in these cells (Figure S1A). These data suggest that mechanisms in addition to E2F and Elk-1 play a more important role in regulating the transcription of the human EZH2 gene in EOC cells.
Figure 1. EZH2 is upregulated at the transcriptional level and two CCAAT sites within proximal region of the human EZH2 gene promoter are critical for its transcriptional activation in human epithelial ovarian cancer (EOC) cells.

(A) EZH2 mRNA level was determined by qRT-PCR in human ovarian surface epithelial (HOSE) cells and indicated EOC cell lines. Mean of three independent experiments with SD. (B) SKOV3 EOC cells were transfected with a luciferase reporter driven by the indicated proximal promoter fragment of the human EZH2 gene. 12 hours post transfection, Luciferase reporter activity was determined. A luminescent β-gal reporter (Clontech) was used to normalize the transfection efficacy. Mean of three independent experiments with SD. (C) Two CCAAT sites in proximal region of the human EZH2 gene promoter were mutated to CCAAC, which is known to be sufficient to inhibit the binding of the CCAAT-binding transcription factor, NF-Y (38). Note that the first CCAAT site is inverted. (D) Luciferase activity of wild type EZH2 (WT) gene promoter compared to CCAAT sites mutated EZH2 (MT) gene promoter as diagramed in (C). Mean of three independent experiments with SD.
Systematic promoter serial deletion analysis revealed that two CCAAT sites in the proximal promoter of the human EZH2 gene are critical for its promoter activity (Figure 1B). We observed similar effects in multiple human EOC cell lines (Figure 1B and S1B–C), demonstrating this is not a cell line specific effect. Strikingly, a single base pair change in each of the CCAAT binding sites (changed to CCAAC) severely impaired the activity of the EZH2 gene promoter in human EOC cells (Figure 1C–D). Based on these findings, we conclude that two CCAAT sites in the proximal region of the human EZH2 gene promoter play a key role in regulating the transcriptional activity of the human EZH2 gene in EOC cells.
NF-YA is expressed at higher levels in human EOC cells and ectopic NF-YA expression drives EZH2 expression in normal HOSE cells
NF-Y is a transcription factor that binds specifically to the CCAAT consensus site (17). We thus sought to determine whether NF-YA, the regulatory subunit of NF-Y (8), is upregulated in human EOC cells. Indeed, NF-YA is expressed at higher levels in human EOC cells compared with normal HOSE cells (Figure 2A and S2). The anti-NF-YA antibody we used here can recognize both short and long isoforms of the NF-YA (Figure 2A and S2). Consistently, both EZH2 and H3K27Me3 are expressed at higher levels in human EOC cells compared with normal HOSE cells (Figure 2A). However, NF-YA upregulation is not a consequence of cell transformation since NF-YA is not upregulated in normal HOSE cells expressing hTERT, SV40 early region (s/L antigen) and oncogenic H-RASG12V (Figure S3), which has previously been shown to be sufficient for transforming normal HOSE cells (18). We next sought to determine whether NF-YA is sufficient to upregulate EZH2 in normal HOSE cells. Toward this goal, two independent isolates of normal HOSE cells were infected with a lentivirus encoding NF-YA and EZH2 expression was examined by immunoblotting analysis. Indeed, ectopic NF-YA expression induced EZH2 expression in normal HOSE cells (Figure 2B). Based on these results, we conclude that NF-YA is expressed at higher levels in human EOC cells compared with normal HOSE cells and ectopic NF-YA is sufficient to upregulate EZH2 in normal HOSE cells.
Figure 2. NF-YA expression positively correlates with EZH2 expression in human EOC cells and a high level of NF-YA predicts shorter overall survival in EOC patients.
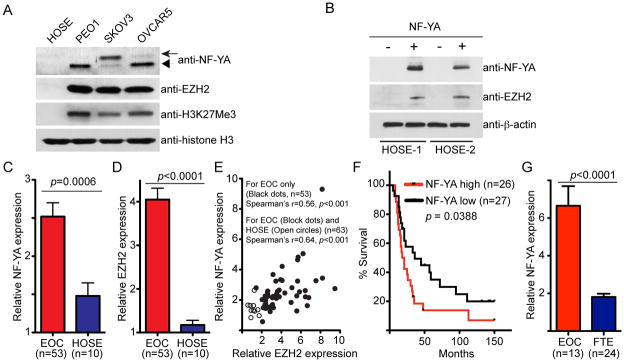
(A) Expression of NF-YA, EZH2, H3K27Me3 and histone H3 in indicated human EOC cell lines and normal HOSE cells determined by immunoblotting. Note that the arrow points to the long NF-YA isoform and the arrowhead points to the short NF-YA isoform. (B) Normal HOSE cells were infected with a lentivirus encoding NF-YA or control. Drug-selected cells were examined for expression of NF-YA, EZH2 and β-actin by immunoblotting. (C) NF-YA is expressed at higher levels in human EOCs compared with normal HOSE cells. Relative expression of NF-YA mRNA in laser capture and microdissected high-grade serous EOC (n = 53) and normal HOSE cells (n = 10). (D) Same as (C) but for expression of EZH2 mRNA. (E) NF-YA expression positively correlates with EZH2 expression. Correlation between expression of NF-YA and EZH2 was determined by Spearman statistical analysis. (F) High NF-YA expression predicts a shorter overall survival in EOC patients. Kaplan-Meier survival curve for EOC patients with low or high NF-YA expression. (G) NF-YA is expressed at higher levels in human EOCs compared with primary non-malignant human fallopian tube epithelial (FTE) cells. Relative expression of NF-YA mRNA in high-grade serous EOC (n = 13) and laser capture and microdissected primary non-malignant FTE cells (n = 24).
NF-YA expression positively correlates with the expression of EZH2 in human EOC cells and a high level of NF-YA predicts shorter overall survival in EOC patients
We next examined whether expression of NF-YA and EZH2 is positively correlated in human EOCs. In addition to EOC cells, EZH2 is also upregulated in ovarian tumor-associated stromal cells such as endothelial cells (19). To limit the confounding effects of EOC-associated stromal cells, we sought to correlate the expression of NF-YA and EZH2 in specimens from laser capture and microdissected (LCM) high-grade serous tumors, which account for a majority of EOC-associated mortalities (3). Toward this goal, we examined the expression of NF-YA and EZH2 in a published microarray database, which compares the gene expression profile in 53 cases of LCM high-grade serous EOC and 10 individual isolations of normal HOSE cells (20). Notably, NF-YA expression is significantly higher in human EOCs compared with normal HOSE cells (Figure 2C). Consistent with previous reports (5, 19), EZH2 was expressed at significantly higher levels in human EOCs compared with normal HOSE cells (Figure 2D). Indeed, there was a significant positive correlation between expression of NF-YA and EZH2 in a Spearman statistical analysis of the cases including EOC alone or both EOC and primary HOSE cells (Figure 2E, p < 0.001). Next, we determined whether there is an association between NF-YA expression and the survival of EOC patients. Upon dividing EOC cases into NF-YA high or low based on the median NF-YA expression, we found that a high level of NF-YA expression was significantly associated with shorter overall survival (Figure 2F, p = 0.0388). Notably, there is recent evidence to suggest that a proportion of serous histosubtype EOC may arise from distant fallopian tube epithelium (21, 22). Using a published microarray analysis (23), we found that NF-YA is also expressed at higher levels in serous ovarian carcinomas compared with primary non-malignant fallopian tube epithelial cells (Figure 2G, p < 0.001). Together, we conclude that NF-YA is expressed at higher levels in human EOC cells compared with normal HOSE cells or non-malignant fallopian tube epithelial cells and ectopic NF-YA is sufficient to drive EZH2 expression in normal HOSE cells. Significantly, expression of NF-YA positively correlates with expression of EZH2 in high-grade serous EOCs and high levels of NF-YA predicts shorter overall survival in EOC patients.
The association of NF-YA with the proximal promoter of the human EZH2 gene is enhanced in human EOC cells
Next, we determined whether NF-YA’s association with the human EZH2 gene promoter is enhanced in human EOC cells compared with normal HOSE cells by chromatin immunoprecipitation (ChIP) analysis using an anti-NF-YA antibody. An isotype matched IgG was used as a negative control and an antibody to the core histone H3 was used as a positive control in ChIP analysis. To further limit potential non-specific effects, we chose a distal region of the human EZH2 gene promoter that does not have NF-YA binding sites as a genomic region negative control (Figure 3A). There is evidence to suggest that NF-YA activates its target gene expression by recruiting p300 histone acetyltransferase (24, 25). Thus, we also examined the association of p300 with the human EZH2 gene promoter using an anti-p300 antibody. Indeed, both NF-YA and p300 showed an enhanced association with the proximal region of the human EZH2 gene promoter in human EOC cells compared with normal HOSE cells (Figure 3B). In contrast, NF-YA and p300 did not associate with the distal region of the human EZH2 promoter that lacks NF-YA binding sites (Figure 3B). Notably, NF-YA is associated with a known NF-YA target gene CCNA2 that has NF-YA binding sites (CCAAT) in its promoter but not with negative control genes such as a-tubulin, RPS19 and YBL1 that do not have NF-YA binding sites in their promoters, demonstrating the specificity of NF-YA antibody in ChIP analysis (Figure S4) (26–28). It has previously been reported that p300 binds NF-Y target gene promoters to promote the expression of these genes by generating acetylation epigenetic marker on histone H3 (25, 29, 30). Indeed, there was an increase in acetylated histone H3 in the proximal promoter of the human EZH2 gene where NF-YA and p300 bind in human EOC cells compared with normal HOSE cells (Figure 3C). Based on these results, we conclude that there is an association of NF-YA and p300 with the proximal promoter of the human EZH2 gene in human EOC cells, and this association correlates with increased levels of acetylated histone H3.
Figure 3. The association of NF-YA and its co-activator p300 with the human EZH2 gene promoter is enhanced in human EOC cells compared with normal HOSE cells.
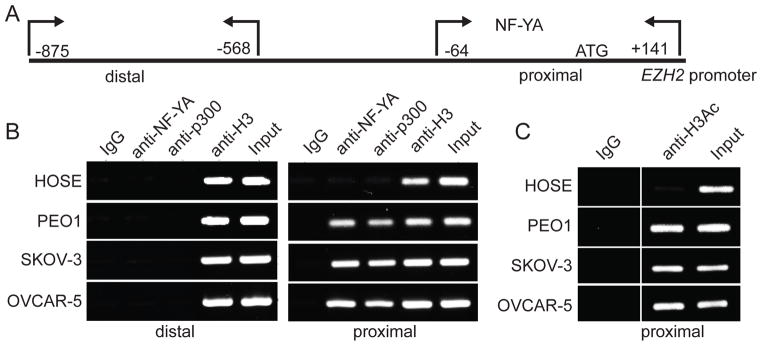
(A) Diagram of the human EZH2 gene promoter with indicated positions of primers used for proximal (−64/+141) and distal (−875/−568) regions of the human EZH2 gene promoter in ChIP analysis. (B) Normal HOSE cells or indicated EOC cell lines were subjected to ChIP analysis using antibodies specific to NF-YA or p300, respectively. An isotype matched IgG was used as a negative control, and antibody against core histone H3 was used as a positive control. After ChIP analysis, the distal and proximal regions of the human EZH2 gene promoter were subjected to PCR amplification using primers detailed in materials and methods and as diagramed in (A). (C) Same as (B) but using an antibody against acetylated histone H3.
NF-YA knockdown suppresses EZH2 expression and inhibits the growth of human EOC cells in vitro and in a xenograft EOC model
We next sought to determine the effects of NF-YA knockdown on expression of EZH2 in human EOC cells. To this end, we used three individual shRNA to the human NF-YA gene (shNF-YA). The knockdown efficacy of shNF-YAs was examined by immunoblotting (Figure 4A and S6A–B) and two shNF-YAs (#1 and #2) were found to efficiently reduce NF-YA levels, while a third (#3) was largely ineffective and used as a non-silencing negative control. Notably, shNF-YA that efficiently knocked down NF-YA also suppressed the expression of EZH2 (Figure 4A and S6A–B). Consistently, the levels of H3K27Me3, the product of EZH2’s methyltransferase activity, were also decreased by shNF-YA that knocks down NF-YA (Figure 4A and S6A–B). In addition, similar observations were made in lung cancer cell line H1299 and breast cancer cell line MCF7, suggesting that this is not unique in EOC cells (Figure S5). We observed the same effects using two individual shNF-YAs (Figure 4A and S6A–B), suggesting that this is not due to off-target effects. In contrast, levels of H3K9Me3, which are generated by different histone methyltransferases such as Suv39H1 and SETDB1 (31), were not affected by NF-YA knockdown (Figure 4A). As a negative control, the shNF-YA #3 that does not efficiently knockdown NF-YA also failed to overtly decrease the EZH2 and H3K27Me3 levels in human EOC cells (Figure 4A).
Figure 4. NF-YA knockdown suppresses the expression of EZH2 and inhibits the growth of human EOC cells in vitro and in a xenograft EOC mouse model.

(A) SKOV3 EOC cells were infected with lentivirus encoding the indicated shNF-YAs or control. Drug-selected cells were examined for expression of NF-YA, EZH2, H3K27Me3 and H3K9Me3 by immunoblotting. Expression of core histone H3 was used as a loading control. (B) Same as (A), but equal number of drug-selected cells was seeded and counted at indicated time points. * p < 0.05 compared with either control or shNF-YA #3. Mean of three independent experiments with SD and linear regression. (C) Same as (A), but equal number of drug-selected cells were assayed for anchorage-independent growth in soft agar. Mean of three independent experiments with SD. * p < 0.05 compared with either control or shNF-YA #3. (D) Same as (A), but equal number of the indicated cells were subcutaneously injected in immunocompromised nude mice (n = 5). Four weeks post injection, tumors were removed from mice. (E) Quantification of (D), the size of tumors was measured. Error bars represent SD. * p < 0.05 compared with either control or shNF-YA #3.
Expression of EZH2 positively correlates with expression of markers of cell proliferation (5) and NF-YA knockdown suppresses the expression of EZH2 (Figure 4A). Thus, we wanted to determine the effects of NF-YA knockdown on the growth of human EOC cells. Compared with controls, two individual shNF-YAs that efficiently knocked down NF-YA suppressed both the anchorage-dependent and -independent growth of human EOC cells (Figure 4B–C). In contrast, one shNF-YA (#3) that does not efficiently knockdown NF-YA failed to suppress the growth of human EOC cells (Figure 4B–C). Similar effects of NF-YA knockdown on cell growth were observed in multiple EOC cells (Figure 4 and S6), demonstrating that this is not cell line specific. Based on these results, we conclude that NF-YA knockdown suppresses the growth of human EOC cells in vitro in both an anchorage-dependent and -independent conditions.
Next, we examined the effects of NF-YA knockdown on the growth of human EOC cells in vivo in a xenograft mouse model. We subcutaneously injected control or shNF-YA-expressing human EOC cells into the immunocompromised (athymic nude) mice. Consistent with our in vitro findings, we found that compared with controls, shNF-YAs that efficiently knocked down NF-YA expression suppressed the growth of xenografted human EOC cells (Figure 4D–E). In contrast, the shNF-YA #3 that does not efficiently knockdown NF-YA also failed to significantly suppress the growth of implanted human EOC cells (Figure 4D–E). Thus, we conclude that NF-YA knockdown suppresses the growth of human EOC cells in vivo in a xenograft EOC mouse model.
NF-YA knockdown triggers apoptosis of human EOC cells
We next determined the mechanism by which NF-YA knockdown suppresses the growth of human EOC cells. Since NF-YA regulates EZH2 expression (Figure 2) and knockdown of EZH2 induces apoptosis of EOC cells (5), we examined the effects of NF-YA on apoptosis of human EOC cells. We observed a significant increase of apoptosis markers in NF-YA knockdown cells. These markers include an increase in Annexin V detection as measured by the Guava Nexin assay (Figure 5A–B) and an increase of cleaved caspase3 and cleaved PARP p85 (Figure 5C). HRK, a pro-apoptotic gene, has recently been demonstrated as an H3K27Me3 target gene and plays a key role in mediating apoptosis induced by decreasing H3K27Me3 levels (32). Since NF-YA knockdown suppresses EZH2 expression and decreasing H3K27Me3 levels, we sought to determine the effects of NF-YA knockdown on the expression of HRK. Indeed, HRK expression is significantly upregulated in NF-YA knockdown cells (Figure 5D). Together, we conclude that NF-YA knockdown induces apoptosis of human EOC cells.
Figure 5. NF-YA knockdown induces apoptosis of human EOC cells.
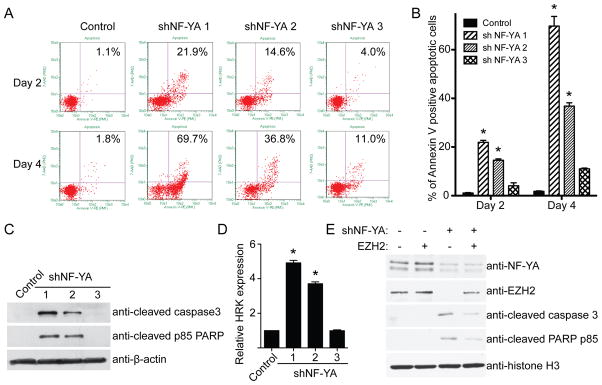
(A) SKOV3 EOC cells were infected with lentivirus encoding the indicated shNF-YAs or control. Guava Nexin assay was performed at the indicated time points. (B) Quantification of (A). Mean of three independent experiments with SD. * p < 0.05 compared with either control or shNF-YA #3. (C) Same as (A) but examined for expression of apoptosis markers, cleaved caspase3 and cleave PARP p85 by immunoblotting. (D) Same as (A) but examined for HRK gene expression by qRT-PCR. * p < 0.05 compared with either control or shNF-YA #3. (E) SKOV3 EOC cells were engineered to express a Tet-inducible EZH2. These cells were infected with a lentivirus shNF-YA with or without simultaneous induction of EZH2 expression. Drug-selected cells were examined for expression of NF-YA, EZH2 and indicated apoptosis markers by immunoblotting. Expression of core histone H3 was used a loading control.
Since NF-YA knockdown suppresses the expression of EZH2, we examined whether downregulation of EZH2 contributes to apoptosis induced by NF-YA knockdown. Toward this goal, we ectopically expressed an inducible EZH2 in EOC cells. We examined the expression of markers of apoptosis in cells expressing shNF-YA with or without inducible EZH2 expression. Compared with shNF-YA only cells, in which EZH2 is downregulated, ectopic EZH2 induction partially rescued the apoptosis induced by NF-YA knockdown as evidenced by decreased levels of apoptotic markers such as cleaved PARP p85 and cleaved caspase 3 (Figure 5E). Together, we conclude that apoptosis induced by NF-YA knockdown is, at least in part, due to suppression of EZH2 expression.
Discussion
We demonstrated that ectopic NF-YA expression is sufficient to drive EZH2 upregulation in normal HOSE cells (Figure 2B). Interestingly, we observed cell death in normal HOSE cells with NF-YA ectopic expression (data not shown), which prevented us from further analyzing these cells. Consistent with our observation, a previous report showed that ectopic NF-YA promotes p53-mediated apoptosis in normal mouse embryonic fibroblasts and inactivation of p53 partially suppresses the apoptosis induced by NF-YA expression (33). Indeed, we also found that p53 is activated upon NF-YA expression in normal HOSE cells (data not shown). These data suggest that p53 inactivation is necessary for the survival of NF-YA expressing cells. Notably, p53 is often inactivated in EOCs. For example, p53 is inactivated in virtually all high-grade serous EOC (7). Thus, it is possible that inactivation of p53 may cooperate with NF-YA in EOC cells. In addition, recent evidence suggests that a proportion of high-grade serous EOC may arise from the distal fallopian tube epithelial cells (34, 35). Thus, it will be interesting to examine whether ectopic NF-YA expression is sufficient to drive EZH2 expression in fallopian tube epithelial cells once these cells become more accessible.
We showed that NF-YA is upregulated in EOCs. However, TCGA gene copy-number analysis indicates that NF-YA gene (6p21.3) amplification occurs rare in EOCs (~1% specimens show >4 copy of NF-YA gene; http://cancergenome.nih.gov/). This suggests that additional mechanisms contribute to NF-YA upregulation in human EOCs. In the future, we will elucidate additional mechanisms that contribute to NF-YA upregulation in EOCs. In addition, it will critical to further validate the correlation between NF-YA expression and survival of EOC patients in independent cohorts.
We discovered that the expression of NF-YA and EZH2 positively correlates in human EOCs (Figure 2C–E), further supporting the premise that NF-YA plays a key role in regulating EZH2 expression. However, the Spearman’s coefficient r was 0.56 for EOC only and 0.64 for EOC plus normal HOSE cells (Figure 2E). This result is consistent with the idea that other factors, such as E2F and Elk-1 (Figure 1B) (6, 16), also play a role in the expression of EZH2.
We showed that EZH2 is a direct NF-YA target gene (Figure 3). EZH2 knockdown induces apoptosis in human EOC cells (5). Consistently, knockdown of NF-YA also triggers apoptosis of human EOC cells (Figure 5). However, ectopic EZH2 expression can only partially rescue the apoptosis induced by NF-YA knockdown and these cells will ultimately undergo apoptosis (Figure 5E and data not shown). These results suggest that other NF-YA target genes also contribute to the apoptosis induced by NF-YA knockdown. Indeed, the anti-apoptotic genes such as Bcl-xl and Bcl2 are also directly controlled by NF-Y transcription factor (36). Further studies are warranted to identify additional NF-YA target genes that contribute to the apoptosis of human EOC cells induced by NF-YA knockdown. In addition, these findings indicate that inhibiting NF-YA could be an alternative strategy for suppressing EZH2 in EOC cells and NF-YA represents a putative target for developing novel EOC therapeutics. Thus, it will be interesting to examine inhibitors of NF-YA transcription factor such as GWL-78 (37) on the malignant phenotypes of human EOC cells.
In summary, we show that two NF-Y binding CCAAT sites within the proximal region of the human EZH2 gene promoter play a key role in regulating EZH2 expression in human EOC cells. NF-YA, the regulatory subunit of NF-Y transcription factor, is upregulated in human EOC cells compared with normal HOSE cells. In addition, ectopic NF-YA drives EZH2 expression in normal HOSE cells. The expression of NF-YA positively correlates with the expression of EZH2 in primary EOCs and a high level of NF-YA expression predicts shorter overall survival in EOC patients. Consistently, NF-YA knockdown suppresses EZH2 expression and inhibits the growth of human EOC cells in vitro and in a xenograft mouse model. NF-YA knockdown triggers apoptosis of human EOC cells and downregulation of EZH2, at least in part, contributes to apoptosis induced by NF-YA knockdown in these cells. These results establish that NF-YA underlies EZH2 upregulation and is essential for proliferation of human EOC cells. We believe that these studies provided important insights into the biology of EOC and identified NF-YA as a potential target for intervention of EOC.
Supplementary Material
Acknowledgments
Grant Support: This work was supported by the National Cancer Institute of the National Institutes of Health (R01CA163377 to R.Z.) and, in part, by a DOD ovarian cancer academy award (OC093420 to R.Z.). Support of Core Facilities used in this study was provided by Cancer Center Support Grant (CCSG) CA010815 to The Wistar Institute.
Footnotes
Disclosure of potential conflict of interest: None.
Cited References
- 1.American Cancer Society. Cancer Facts & Figures 2012. Atlanta: American Cancer Society; 2012. [Google Scholar]
- 2.Arulkumaran S, Regan L, Farquharson DIM. Obstetrics and gynaecology. Oxford: Oxford University Press; 2011. [Google Scholar]
- 3.Shih Ie M, Kurman RJ. Ovarian tumorigenesis: a proposed model based on morphological and molecular genetic analysis. Am J Pathol. 2004;164:1511–8. doi: 10.1016/s0002-9440(10)63708-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Cao R, Wang L, Wang H, Xia L, Erdjument-Bromage H, Tempst P, et al. Role of histone H3 lysine 27 methylation in Polycomb-group silencing. Science. 2002;298:1039–43. doi: 10.1126/science.1076997. [DOI] [PubMed] [Google Scholar]
- 5.Li H, Cai Q, Godwin AK, Zhang R. Enhancer of zeste homolog 2 promotes the proliferation and invasion of epithelial ovarian cancer cells. Molecular cancer research: MCR. 2010;8:1610–8. doi: 10.1158/1541-7786.MCR-10-0398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bracken AP, Pasini D, Capra M, Prosperini E, Colli E, Helin K. EZH2 is downstream of the pRB-E2F pathway, essential for proliferation and amplified in cancer. EMBO J. 2003;22:5323–35. doi: 10.1093/emboj/cdg542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.The Cancer Genome Atlas Research Network. Integrated genomic analyses of ovarian carcinoma. Nature. 2011;474:609–15. doi: 10.1038/nature10166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mantovani R. The molecular biology of the CCAAT-binding factor NF-Y. Gene. 1999;239:15–27. doi: 10.1016/s0378-1119(99)00368-6. [DOI] [PubMed] [Google Scholar]
- 9.Marziali G, Perrotti E, Ilari R, Coccia EM, Mantovani R, Testa U, et al. The activity of the CCAAT-box binding factor NF-Y is modulated through the regulated expression of its A subunit during monocyte to macrophage differentiation: regulation of tissue-specific genes through a ubiquitous transcription factor. Blood. 1999;93:519–26. [PubMed] [Google Scholar]
- 10.Farina A, Manni I, Fontemaggi G, Tiainen M, Cenciarelli C, Bellorini M, et al. Down-regulation of cyclin B1 gene transcription in terminally differentiated skeletal muscle cells is associated with loss of functional CCAAT-binding NF-Y complex. Oncogene. 1999;18:2818–27. doi: 10.1038/sj.onc.1202472. [DOI] [PubMed] [Google Scholar]
- 11.Bolognese F, Wasner M, Dohna CL, Gurtner A, Ronchi A, Muller H, et al. The cyclin B2 promoter depends on NF-Y, a trimer whose CCAAT-binding activity is cell-cycle regulated. Oncogene. 1999;18:1845–53. doi: 10.1038/sj.onc.1202494. [DOI] [PubMed] [Google Scholar]
- 12.Li XY, Hooft van Huijsduijnen R, Mantovani R, Benoist C, Mathis D. Intron-exon organization of the NF-Y genes. Tissue-specific splicing modifies an activation domain. J Biol Chem. 1992;267:8984–90. [PubMed] [Google Scholar]
- 13.Gurtner A, Fuschi P, Magi F, Colussi C, Gaetano C, Dobbelstein M, et al. NF-Y dependent epigenetic modifications discriminate between proliferating and postmitotic tissue. PloS one. 2008;3:e2047. doi: 10.1371/journal.pone.0002047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Li Q, Herrler M, Landsberger N, Kaludov N, Ogryzko VV, Nakatani Y, et al. Xenopus NF-Y pre-sets chromatin to potentiate p300 and acetylation-responsive transcription from the Xenopus hsp70 promoter in vivo. EMBO J. 1998;17:6300–15. doi: 10.1093/emboj/17.21.6300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Yamanaka K, Mizuarai S, Eguchi T, Itadani H, Hirai H, Kotani H. Expression levels of NF-Y target genes changed by CDKN1B correlate with clinical prognosis in multiple cancers. Genomics. 2009;94:219–27. doi: 10.1016/j.ygeno.2009.06.003. [DOI] [PubMed] [Google Scholar]
- 16.Fujii S, Tokita K, Wada N, Ito K, Yamauchi C, Ito Y, et al. MEK-ERK pathway regulates EZH2 overexpression in association with aggressive breast cancer subtypes. Oncogene. 2011;30:4118–28. doi: 10.1038/onc.2011.118. [DOI] [PubMed] [Google Scholar]
- 17.Mantovani R. A survey of 178 NF-Y binding CCAAT boxes. Nucleic Acids Res. 1998;26:1135–43. doi: 10.1093/nar/26.5.1135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Liu J, Yang G, Thompson-Lanza JA, Glassman A, Hayes K, Patterson A, et al. A genetically defined model for human ovarian cancer. Cancer Res. 2004;64:1655–63. doi: 10.1158/0008-5472.can-03-3380. [DOI] [PubMed] [Google Scholar]
- 19.Lu C, Han HD, Mangala LS, Ali-Fehmi R, Newton CS, Ozbun L, et al. Regulation of tumor angiogenesis by EZH2. Cancer Cell. 2010;18:185–97. doi: 10.1016/j.ccr.2010.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mok SC, Bonome T, Vathipadiekal V, Bell A, Johnson ME, Wong KK, et al. A gene signature predictive for outcome in advanced ovarian cancer identifies a survival factor: microfibril-associated glycoprotein 2. Cancer Cell. 2009;16:521–32. doi: 10.1016/j.ccr.2009.10.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kurman RJ, Shih Ie M. The origin and pathogenesis of epithelial ovarian cancer: a proposed unifying theory. Am J Surg Pathol. 2010;34:433–43. doi: 10.1097/PAS.0b013e3181cf3d79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Levanon K, Crum C, Drapkin R. New insights into the pathogenesis of serous ovarian cancer and its clinical impact. J Clin Oncol. 2008;26:5284–93. doi: 10.1200/JCO.2008.18.1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tone AA, Begley H, Sharma M, Murphy J, Rosen B, Brown TJ, et al. Gene expression profiles of luteal phase fallopian tube epithelium from BRCA mutation carriers resemble high-grade serous carcinoma. Clinical cancer research: an official journal of the American Association for Cancer Research. 2008;14:4067–78. doi: 10.1158/1078-0432.CCR-07-4959. [DOI] [PubMed] [Google Scholar]
- 24.Hughes R, Kristiansen M, Lassot I, Desagher S, Mantovani R, Ham J. NF-Y is essential for expression of the proapoptotic bim gene in sympathetic neurons. Cell Death Differ. 2010 doi: 10.1038/cdd.2010.166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Salsi V, Caretti G, Wasner M, Reinhard W, Haugwitz U, Engeland K, et al. Interactions between p300 and multiple NF-Y trimers govern cyclin B2 promoter function. The Journal of biological chemistry. 2003;278:6642–50. doi: 10.1074/jbc.M210065200. [DOI] [PubMed] [Google Scholar]
- 26.Kramer A, Carstens CP, Wasserman WW, Fahl WE. CBP/cycA, a CCAAT-binding protein necessary for adhesion-dependent cyclin A transcription, consists of NF-Y and a novel Mr 115,000 subunit. Cancer Res. 1997;57:5117–21. [PubMed] [Google Scholar]
- 27.Kramer A, Carstens CP, Fahl WE. A novel CCAAT-binding protein necessary for adhesion-dependent cyclin A transcription at the G1/S boundary is sequestered by a retinoblastoma-like protein in G0. The Journal of biological chemistry. 1996;271:6579–82. doi: 10.1074/jbc.271.12.6579. [DOI] [PubMed] [Google Scholar]
- 28.Testa A, Donati G, Yan P, Romani F, Huang TH, Vigano MA, et al. Chromatin immunoprecipitation (ChIP) on chip experiments uncover a widespread distribution of NF-Y binding CCAAT sites outside of core promoters. The Journal of biological chemistry. 2005;280:13606–15. doi: 10.1074/jbc.M414039200. [DOI] [PubMed] [Google Scholar]
- 29.Di Agostino S, Strano S, Emiliozzi V, Zerbini V, Mottolese M, Sacchi A, et al. Gain of function of mutant p53: the mutant p53/NF-Y protein complex reveals an aberrant transcriptional mechanism of cell cycle regulation. Cancer Cell. 2006;10:191–202. doi: 10.1016/j.ccr.2006.08.013. [DOI] [PubMed] [Google Scholar]
- 30.Caretti G, Salsi V, Vecchi C, Imbriano C, Mantovani R. Dynamic recruitment of NF-Y and histone acetyltransferases on cell-cycle promoters. J Biol Chem. 2003;278:30435–40. doi: 10.1074/jbc.M304606200. [DOI] [PubMed] [Google Scholar]
- 31.Jenuwein T. The epigenetic magic of histone lysine methylation. FEBS J. 2006;273:3121–35. doi: 10.1111/j.1742-4658.2006.05343.x. [DOI] [PubMed] [Google Scholar]
- 32.Li H, Cai Q, Wu H, Vathipadiekal V, Dobbin ZC, Li T, et al. SUZ12 Promotes Human Epithelial Ovarian Cancer by Suppressing Apoptosis via Silencing HRK. Molecular cancer research: MCR. 2012 doi: 10.1158/1541-7786.MCR-12-0335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gurtner A, Fuschi P, Martelli F, Manni I, Artuso S, Simonte G, et al. Transcription factor NF-Y induces apoptosis in cells expressing wild-type p53 through E2F1 upregulation and p53 activation. Cancer Res. 2010;70:9711–20. doi: 10.1158/0008-5472.CAN-10-0721. [DOI] [PubMed] [Google Scholar]
- 34.Kurman RJ, Shih Ie M. The origin and pathogenesis of epithelial ovarian cancer: a proposed unifying theory. Am J Surg Pathol. 2010;34:433–43. doi: 10.1097/PAS.0b013e3181cf3d79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Levanon K, Crum C, Drapkin R. New insights into the pathogenesis of serous ovarian cancer and its clinical impact. J Clin Oncol. 2008;26:5284–93. doi: 10.1200/JCO.2008.18.1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Benatti P, Basile V, Merico D, Fantoni LI, Tagliafico E, Imbriano C. A balance between NF-Y and p53 governs the pro- and anti-apoptotic transcriptional response. Nucleic Acids Res. 2008;36:1415–28. doi: 10.1093/nar/gkm1046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kotecha M, Kluza J, Wells G, O’Hare CC, Forni C, Mantovani R, et al. Inhibition of DNA binding of the NF-Y transcription factor by the pyrrolobenzodiazepine-polyamide conjugate GWL-78. Mol Cancer Ther. 2008;7:1319–28. doi: 10.1158/1535-7163.MCT-07-0475. [DOI] [PubMed] [Google Scholar]
- 38.Hu Q, Bhattacharya C, Maity SN. CCAAT binding factor (CBF) binding mediates cell cycle activation of topoisomerase IIalpha. Conventional CBF activation domains are not required. J Biol Chem. 2002;277:37191–200. doi: 10.1074/jbc.M205985200. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.