

Abstract
Recent expeditions have revealed high levels of biodiversity in the tropical deep-sea, yet little is known about the age or origin of this biodiversity, and large-scale molecular studies are still few in number. In this study, we had access to the largest number of solariellid gastropods ever collected for molecular studies, including many rare and unusual taxa. We used a Bayesian chronogram of these deep-sea gastropods (1) to test the hypothesis that deep-water communities arose onshore, (2) to determine whether Antarctica acted as a source of diversity for deep-water communities elsewhere and (3) to determine how factors like global climate change have affected evolution on the continental slope. We show that although fossil data suggest that solariellid gastropods likely arose in a shallow, tropical environment, interpretation of the molecular data is equivocal with respect to the origin of the group. On the other hand, the molecular data clearly show that Antarctic species sampled represent a recent invasion, rather than a relictual ancestral lineage. We also show that an abrupt period of global warming during the Palaeocene Eocene Thermal Maximum (PETM) leaves no molecular record of change in diversification rate in solariellids and that the group radiated before the PETM. Conversely, there is a substantial, although not significant increase in the rate of diversification of a major clade approximately 33.7 Mya, coinciding with a period of global cooling at the Eocene–Oligocene transition. Increased nutrients made available by contemporaneous changes to erosion, ocean circulation, tectonic events and upwelling may explain increased diversification, suggesting that food availability may have been a factor limiting exploitation of deep-sea habitats. Tectonic events that shaped diversification in reef-associated taxa and deep-water squat lobsters in central Indo-West Pacific were also probably important in the evolution of solariellids during the Oligo-Miocene.
Keywords: Biogeography, deep sea, Eocene–Oligocene transition, phylogeny
Introduction
The deep sea is the largest and most enigmatic of the Earth's ecosystems (Ramirez-Llodra et al. 2010). Covering almost two-thirds of the planet's surface, it was once thought to be devoid of life. Studies over the last hundred years, however, have shown that the deep sea is in fact rich in species, some with bizarre and novel adaptations to the challenges of living at great depth. This understudied but important marine environment is at risk from overexploitation and habitat destruction as a result of both fishing and mining ventures (e.g., Halfar and Fujita 2007; Van Dover 2011), and it is vital that we learn more about the diversity of its biota and their evolution before these habitats suffer further destruction.
Elucidating the factors driving diversification in the deep sea is of profound importance if we are to understand how deep-sea groups have evolved. Climate change has been shown to result in shifts in primary producers that also affect deep-sea community structure over periods of a few years (Ruhl and Smith 2004), over hundreds of thousands of years spanning the past four glacial–interglacial cycles (Yasuhara et al. 2009, 2012) and over millions of years (Smith and Stockley 2005). It is likely therefore that climate change over geological time has also played an important role in the evolution of deep-sea diversity (e.g., Lipps and Mitchell 1976; Gingerich 2006; Berger 2007).
The Palaeocene/Eocene boundary (∼55.8 Mya) was marked by a brief but intense global warming event, known as the Palaeocene-Eocene Thermal Maximum (PETM), which saw global temperatures rise by 5°C over 10,000 years. Sea surface temperatures rose between 5°C in the tropics and 9°C in the high latitudes, and bottom-water temperatures by 4–5°C over a period of about 10,000 years (Zachos et al. 2001, 2008). The event was also associated with a massive injection of 13C-depleted carbon into the ocean-atmosphere system (Dickens et al. 1995), resulting in shallowing of the calcite compensation depth (CCD) and acidification in the deep sea (Zachos et al. 2005). This in turn is thought to have contributed to the contemporaneous mass extinction of benthic Foraminifera, although most plankton survived (Tjalsma and Lohmann 1983; Zachos et al. 2005) and a transient diversification was observed in topical, planktonic foraminifera (Kelly et al. 1998). On land, this dramatic climate change resulted in a rapid increase in plant speciation and diversity in tropical habitats (Jaramillo et al. 2010), a turnover in large mammals in northern continents and possibly Africa (Gingerich 2006; Blois and Hadley 2009) and a rapid and transient northward migration of plants in North America (Wing et al. 2005). The PETM was followed by the Eocene Optimum, an extended period of very warm temperatures (Zachos et al. 2001, 2008).
Another dramatic change in global climate occurred at the Eocene–Oligocene transition (EOT), when there was a period of abrupt cooling lasting about 500 kyr between 33.5 and 34 Mya (Zachos et al. 2001, 2008; Pearson et al. 2008). Atmospheric and high latitude sea-surface temperatures cooled by ∼5°C (Kennett and Shackleton 1976; Liu et al. 2009) and both the thermocline and deep water of the Southern Ocean and Indian Ocean cooled by 2–3°C across the EOT (Dunkley Jones et al. 2008; Bohaty et al. 2012). The Earth shifted from “greenhouse” to “icehouse” conditions with large, permanent ice-sheets forming in Antarctica (Zachos et al. 1996; Lear et al. 2000). The growth of a continental-scale ice sheet in Antarctic is thought to have been a primary driver of changes to Antarctic circulation, which in turn caused increased latitudinal thermal gradients, increased thermohaline circulation, increased deep-basin ventilation, decreased deep-ocean acidity, a deepening of the CCD and intensified upwelling that coincided with periods of sea-level fluctuations (van Andel 1975; Coxall et al. 2005; Rea and Lyle 2005; Berger 2007; Miller et al. 2009). The onset of the Antarctic Circumpolar Current (ACC) also occurred concurrently with the EOT and tectonic events leading to the opening of both the Drake and Tasman Passages (Katz et al. 2011).
Intense chemical weathering of siliceous rocks at high latitudes is thought to have occurred prior to the EOT during the warm climate of the Eocene Optimum, releasing high concentrations of silica into the oceans (Lear et al. 2000). The ACC triggered mixing of deep-water layers around Antarctica, leading to an increase in silica and other nutrients in the deep sea (Lear et al. 2000; Berger 2007; Marx and Uhen 2010). The increase in nutrients is thought to have resulted in diversification of siliceous diatoms (Miller et al. 2009), which in turn is thought to have resulted in increased abundance of krill, leading to the diversification of whales in southern oceans (Berger 2007; Marx and Uhen 2010).
Little is known about the origin of deep-sea clades, but fossil evidence suggests that post-Palaeozoic order-level benthic marine taxa first appeared near or onshore, even if they are now found only in the deep sea (Jablonski et al. 1983; Jablonski and Bottjer 1991; Jablonski 2005). Iconic examples for this include primitive taxa such as monoplacophorans and stalked crinoids that dominated shallow marine environments during the Palaeozoic, but currently occur only in deep-sea refugia (Lemche 1957; Ameziane and Roux 1997; Aronson and Blake 2001; Kano et al. 2012). Conversely, the origins of lower level taxa such as families and genera do not always conform to this pattern (Jablonski 2005). Indeed, molecular evidence suggests radiations move from offshore to onshore (e.g., deep-sea corals, Lindner et al. 2008) as well as in the reverse direction (e.g., isopods, Raupach et al. 2012). Previous authors have suggested the shift in origin may have been due to deep-sea anoxic events that were frequent prior to the Cenozoic (Jacobs and Lindberg 1998); however, this is now debated (Jablonski 2005). An additional hypothesis is that early deep-sea radiations originated in shallow-water, but were displaced into deep-water as a result of pressure from predators or competitors (Vermeij 1987).
Another theory is that since the onset of glacial climates, Antarctica may have acted as a center of origin for deep-sea taxa, with Antarctic shelf taxa moving into deep-water as a result of climatic deterioration during glaciation periods and the subsequent loss of shallow-water habitat (Zinsmeister and Feldmann 1984; Crame 1993; Rogers 2000; Briggs 2003; Brandt et al. 2007; Strugnell et al. 2008). Range expansion of Antarctic marine organisms into the Southern Ocean followed the development of the Antarctic Circumpolar Current (ACC; ∼33.8 Mya) and the northward movement of Antarctic bottom water (20–5 Mya; Lawver and Gahagan 2003). The ACC connected shallow-water Antarctic fauna with deep-water in the Atlantic, Indian and Pacific Oceans contributing to the Cenozoic diversification in the Southern Ocean (Brey et al. 1996; Rogers 2000; Briggs 2003; Brandt et al. 2007; Strugnell et al. 2008; Clarke and Crame 2010).
We used a deep-sea radiation of vetigastropods as a tractable model to test these key hypotheses about origins and to determine the factors driving diversification on the continental slope. The marine gastropod family Solariellidae is a group of small (5 mm–2 cm) marine snails that occur globally, predominantly in deep-water, although some species occur as shallow as 5 m (Warén 1993). Specifically, our objectives were to use Bayesian inference to estimate species trees and divergence times, with palaeontological data informing calibration of key nodes in the tree. The resulting chronogram was used: (1) to test the hypothesis that deep-water communities arose from shallow-water ancestors; (2) to determine whether Antarctic shallow-water species represent new invasions or relictual ancestors of lineages that acted as a source of diversity for deep-water communities elsewhere; and (3) to examine the timing of diversification in order to determine the factors driving evolution in the deep sea. Two factors were of special interest: the two shifts in global temperature discussed above and tectonic activity in Southeast Asia. The latter has been shown to drive diversification in both shallow and deep-water groups in the Indo-West Pacific (Kohn 1990; Wilson and Rosen 1998; Williams 2007; Renema et al. 2008; Williams and Duda 2008; Bellwood et al. 2012; Cabezas et al. 2012) and we would expect to see a similar pattern in solariellids. We would predict that the PETM would have had little effect on deep-sea organisms over the depth distribution of solariellids, as they may have been less affected by dramatic temperature increases than intertidal and terrestrial organisms and most solariellid species occur above the CCD. Conversely, we would predict that events contemporaneous with the EOT might have led to increased diversification in Southern Ocean and Indo-West Pacific (IWP) solariellids, reflecting diversification patterns of other marine taxa in the Southern Ocean (e.g., Berger 2007; Miller et al. 2009; Marx and Uhen 2010).
Materials and Methods
Samples
Recently, MNHN deep-sea expeditions have obtained unprecedented collections of solariellids from New Caledonia, Vanuatu, Solomon Islands, Philippines, Norfolk Ridge, Chesterfield Bank, Papua New Guinea, Madagascar and Mozambique Channel, all of which were included in this study. Additional specimens from Japan, Antarctica, Norway, New Zealand, South Africa, and Australia were collected by the authors or loaned from other museums. Sequences were obtained from a total of 208 solariellid specimens and 25 outgroup taxa (Table 1 for solariellids, Table S1 for outgroup taxa). Based on recent revisions, our study has included all but two genera: the IWP genera Minolops (which may be synonymous with Spectamen; Marshall 1999) and one Atlantic genus, Microgaza. The choice of outgroup taxa was based on Williams (2012).
Table 1.
Solariellid specimens used in study, ordered by genera or clades identified in this study, along with details of sampling localities (expedition name, station number, detailed sampling locality, depth range of trawl or dredge and longitude and latitude of start point of dredge), registration numbers of voucher specimens, and EMBL accession numbers for sequences
| Species | Expedition/Station | Sample locality | Depth | Lat/Long | Reg | 28S | COI | 16S | 12S |
|---|---|---|---|---|---|---|---|---|---|
| Archiminolia 1 | SALOMON2/DW2301 | S Gatukai I., Solomon Islands | 267–329 m | 9°6.9′S, 158°20.6′E | MNHN 200718540 | HF586167 | HF586310 | HF586019 | HF585858 |
| Archiminolia 2 | EBISCO/CP2572 | N Bellona, New Caledonia | 324–330 m | 20°23′S, 158°45′E | MNHN 200718316 | HF586168 | HF586311 | HF586020 | HF585859 |
| Archiminolia 2 | EBISCO/DW2522 | S Banc Nova, New Caledonia | 310–318 m | 22°46′S, 159°21′E | MNHN 200718321 | HF586169 | HF586312 | HF586021 | HF585860 |
| Archiminolia 2 | EBISCO/DW2532 | N Banc Nova, New Caledonia | 350 m | 22°15′S, 159°27′E | MNHN 200734079 | HF586174 | HF586317 | HF586026 | HF585866 |
| Archiminolia 2 | NORFOLK2/DW2091 | Banc Antigonia, Norfolk Ridge | 600–896 m | 24°45′S, 168°06′E | MNHN 200718338 | HF586170 | HF586313 | HF586022 | HF585861 |
| Archiminolia 2 | NORFOLK2/DW2117 | Banc Kaimon Maru, Norfolk Ridge | 400 m | 23°24′S, 168°00′E | MNHN 200718339 | – | – | – | HF585862 |
| Archiminolia 2 | RV Tangaroa/2003020 | Norfolk Ridge, S of Norfolk I., Australia | 322–337 m | 29°41.8′S, 168°2.6′E | NMNZ M171105 | – | – | – | HF585867 |
| Archiminolia 2 | TERRASSES/DW3108 | Munida, Norfolk Ridge | 370–440 m | 23°01′S, 168°23′E | MNHN 20098803 | HF586171 | HF586314 | HF586023 | HF585863 |
| Archiminolia 2 | TERRASSES/DW3063 | Banc Antigonia, Norfolk Ridge | 430–480 m | 23°23′S, 168°00′E | MNHN 20098804 | HF586172 | HF586315 | HF586024 | HF585864 |
| Archiminolia 2 | TERRASSES/DW3107 | Munida, Norfolk Ridge | 380–440 m | 23°01′S, 168°23′E | MNHN 20098867 | HF586173 | HF586316 | HF586025 | HF585865 |
| Bathymophila 1 | EBISCO/CP2571 | N Bellona, New Caledonia | 298–309 m | 20°25′S, 158°45′E | MNHN 200718313 | HF586073 | HF586214 | HF585923 | HF585747 |
| Bathymophila 1 | EBISCO/DW2639 | N Lansdowne, New Caledonia | 289–294 m | 20°47′S, 161°01′E | MNHN 200734144 | HF586074 | HF586215 | HF585924 | HF585748 |
| Bathymophila 1 | EBISCO/DW2639 | S Lansdowne, New Caledonia | 289–294 m | 20°47′S, 161°01′E | MNHN 200734145 | HF586075 | HF586216 | HF585925 | HF585749 |
| Bathymophila 2 | CONCALIS/DW2993 | Grand Passage, New Caledonia | 700–730 m | 18°00′S, 163°02′E | MNHN 200735577 | HF586077 | HF586218 | HF585927 | HF585751 |
| Bathymophila 2 | CONCALIS/DW3023 | Grand Passage, New Caledonia | 285–300 m | 19°00′S, 163°26′E | MNHN 200735589 | HF586078 | – | HF585928 | HF585752 |
| Bathymophila 2 | EBISCO/DW2584 | Chesterfield, New Caledonia | 569–570 m | 19°38′S, 158°44′E | MNHN 200718323 | HE800722 | HE800623 | HE800762 | HE800673 |
| Bathymophila diadema | BIOPAPUA/CP3755 | Off Bougainville, Papua New Guinea | 662 m | 5°04′S, 154°29′E | MNHN 200915191 | HF586088 | HF586229 | HF585938 | HF585764 |
| Bathymophila diadema | CONCALIS/DW2983 | Grand Passage, New Caledonia | 367–430 m | 18°01′S, 163°02′E | MNHN 200735575 | HF586087 | HF586228 | HF585937 | HF585763 |
| Bathymophila diadema | EBISCO/CP2556 | W Bellona, New Caledonia | 741–791 m | 21°06′S, 158°32′E | MNHN 200718311 | HF586079 | HF586219 | HF585929 | HF585753 |
| Bathymophila diadema | EBISCO/CP2556 | W Bellona, New Caledonia | 741–791 m | 21°06′S, 158°32′E | MNHN 200718312 | HF586080 | HF586220 | HF585930 | HF585754 |
| Bathymophila diadema | EBISCO/CP2651 | SE Fairway, New Caledonia | 883–957 m | 21°29′S, 162°36′E | MNHN 200718319 | HF586081 | HF586221 | HF585931 | HF585755 |
| Bathymophila diadema | EBISCO/CP2651 | SE Fairway, New Caledonia | 883–957 m | 21°29′S, 162°36′E | MNHN 200718320 | HF586082 | HF586222 | HF585932 | HF585756 |
| Bathymophila diadema | EBISCO/DW2544 | W Bellona, New Caledonia | 650–723 m | 21°10′S, 158°39′E | MNHN 200718322 | HE800721 | HE800622 | HE800761 | HE800672 |
| Bathymophila diadema | SALOMON2/CP2249 | NW Vella, Lavella I., Solomon Islands | 782–884 m | 7°31.3′S, 156°17.7′E | MNHN 200718535 | – | HF586223 | – | HF585725 |
| Bathymophila diadema | SALOMON2/CP2249 | NW Vella, Lavella I., Solomon Islands | 782–884 m | 7°31.3′S, 156°17.7′E | MNHN 200913010 | – | – | – | HF585757 |
| Bathymophila diadema | SALOMON2/CP2249 | NW Vella, Lavella I., Solomon Islands | 782–884 m | 7°31.3′S, 156°17.7′E | MNHN 200913011 | – | HF586224 | – | HF585758 |
| Bathymophila diadema | TERRASSES/DW3040 | Mont J, Loyalty Ridge | 750–780 m | 23°58′S, 169°43′E | MNHN 20098802 | HF586083 | HF586225 | HF585933 | HF585760 |
| Bathymophila diadema | TERRASSES/DW3045 | Mont J, Loyalty Ridge | 660–710 m | 23°48′S, 169°46′E | MNHN 20098869 | HF586084 | HF586226 | HF585934 | HF585761 |
| Bathymophila diadema | TERRASSES/DW3045 | Mont J, Loyalty Ridge | 660–710 m | 23°48′S, 169°46′E | MNHN 20098871 | HF586085 | HF586227 | HF585935 | HF585762 |
| Bathymophila diadema | CONCALIS/DW2983 | Grand Passage, New Caledonia | 367–430 m | 18°01′S, 163°02′E | MNHN 200735574 | HF586086 | – | HF585936 | – |
| Bathymophila 4 | MIRIKY/CP3221 | Between Nosy-bé and Banc du Leven, Madagascar | 782 m | 12°47′S, 48°08′E | MNHN 20098762 | HF586089 | HF586230 | HF585939 | HF585765 |
| Bathymophila 4 | MIRIKY/CP3221 | Between Nosy-bé and Banc du Leven, Madagascar | 782 m | 12°47′S, 48°08′E | MNHN 20098763 | HF586090 | HF586231 | HF585940 | HF585766 |
| Bathymophila 4 | MIRIKY/CP3221 | Between Nosy-bé and Banc du Leven, Madagascar | 782 m | 12°47′S, 48°08′E | MNHN 20098764 | HF586091 | HF586232 | – | HF585767 |
| Bathymophila 4 | MIRIKY/CP3192 | Between Nosy-bé and Banc du Leven, Madagascar | 578–580 m | 12°26′S, 48°13′E | MNHN 20098769 | HF586092 | HF586233 | HF585941 | HF585768 |
| Bathymophila 4 | MIRIKY/CP3186 | Between Nosy-bé and Banc du Leven, Madagascar | 613–625 m | 12°34′S, 48°09′E | MNHN 20098770 | HF586093 | HF586234 | HF585942 | HF585769 |
| Bathymophila 4 | MIRIKY/CP3192 | Between Nosy-bé and Banc du Leven, Madagascar | 578–580 m | 12°26′S, 48°13′E | MNHN 20098771 | HF586094 | HF586235 | HF585943 | HF585770 |
| Bathymophila 4 | MIRIKY/CP3221 | Between Nosy-bé and Banc du Leven, Madagascar | 782 m | 12°47′S, 48°08′E | MNHN 20098772 | HF586095 | HF586236 | HF585944 | HF585771 |
| Bathymophila 4 | MIRIKY/CP3221 | Between Nosy-bé and Banc du Leven, Madagascar | 782 m | 12°47′S, 48°08′E | MNHN 20098773 | HF586096 | HF586237 | HF585945 | HF585772 |
| Bathymophila 5 | AURORA/CP2683 | Philippines | 1743–1754 m | 15°06′N, 123°04′E | MNHN 200718295 | HF586076 | HF586217 | HF585926 | HF585750 |
| Bathymophila 6 | CONCALIS/DW2990 | Grand Passage, New Caledonia | 650–700 m | 17°59′S, 163°03′E | MNHN 200735547 | HF586097 | HF586238 | HF585946 | HF585773 |
| Bathymophila 7 | EBISCO/CP2651 | SE Fairway, New Caledonia | 883–957 m | 21°29′S, 162°36′E | MNHN 200718317 | HF586098 | HF586239 | HF585947 | HF585774 |
| Bathymophila cf callomphala | CONCALIS/DW2980 | Grand Passage, New Caledonia | 574–660 m | 18°16′S, 162°57′E | MNHN 200735553 | HF586099 | HF586240 | HF585948 | HF585778 |
| Bathymophila 9 | CONCALIS/DW3023 | Grand Passage, New Caledonia | 285–300 m | 19°00′S, 163°26′E | MNHN 200735590 | HF586100 | HF586241 | HF585949 | HF585779 |
| Bathymophila 10 | BIOPAPUA/CP3724 | Vitiaz Straight, Papua New Guinea | 860–880 m | 05°59′S, 147°39′E | MNHN 200915182 | HF586101 | HF586242 | HF585950 | HF585781 |
| Bathymophila 11 | TARASOC/DW3369 | Niau, Tuamotu Archipelago | 412–520 m | 16°08′S, 146°24′W | MNHN 200915175 | HF586102 | HF586243 | HF585951 | HF585787 |
| Bathymophila 12 | BORDAU1/DW1469 | Fiji | 314–377 m | 19°40′S, 178°10′W | MNHN 200928741 | – | – | – | HF585775 |
| Bathymophila alabida | RV Karehoa/2000044 | S Kermadec Ridge, Rumble III volcano, New Zealand | 523 m | 35°43.4′S, 178°29.3′E | NMNZ M299686 | – | – | – | HF585776 |
| Bathymophila 14 | BORDAU1/DW1432 | Fiji | 477–493 m | 17°20′S, 178°44′W | MNHN NR | – | – | – | HF585777 |
| Bathymophila 15 | T/V Nagasaki-maru, N226/Dredge A | SW of Nagasaki, Kyushu I., Japan | 470–487 m | 32º 10′ N, 129º 30′ E | YK1383 | HF586103 | HF586244 | HF585952 | HF585782 |
| Bathymophila 16 | BOA1/CP2473 | Between Ambrim and Malekula, Vanuatu | 657–685 m | 16º 19′ S, 167º 47′ E | YK1385 | HF586104 | – | HF585953 | HF585783 |
| Bathymophila 17 | BENTHAUS/DW1951 | Lotus Bank, Austral Is. | 206–450 m | 23°49′S, 147°53′W | MNHM 20095062 | – | – | GQ160692 | – |
| Clade A sp 1 | BIOPAPUA/DW3688 | Seamount S of Manus I., Papua New Guinea | 402–640 m | 3°04′S, 147°32′E | MNHN 200915186 | HF586157 | HF586300 | – | HF585846 |
| Clade A sp 1 | BIOPAPUA/DW3687 | Seamount S of Manus I., Papua New Guinea | 305–579 m | 3°04′S, 147°32′E | MNHN 200915188 | HF586158 | HF586301 | HF586010 | HF585847 |
| Clade A sp 1 | BIOPAPUA/DW3687 | Seamount S of Manus I., Papua New Guinea | 305–579 m | 3°04′S, 147°32′E | MNHN 200915189 | HF586159 | HF586302 | HF586011 | HF585848 |
| Clade A tenorioi | PANGLAO2005/CP2394 | Off Balicasag I., Philippines | 470–566 m | 9°28.6′N, 123°40′E | MNHN 200718423 | – | HF586305 | HF586015 | HF585853 |
| Clade A tenorioi | PANGLAO2005/CP2394 | Off Balicasag I., Philippines | 470–566 m | 9°28.6′N, 123°40′E | MNHN 200718424 | HF586163 | HF586306 | HF586016 | HF585854 |
| Clade A tenorioi | PANGLAO2005/CP2394 | Off Balicasag I., Philippines | 470–566 m | 9°28.6′N, 123°40′E | MNHN 200718425 | HF586164 | HF586307 | – | HF585855 |
| Clade A tenorioi | PANGLAO2005/CP2394 | Off Balicasag I., Philippines | 470–566 m | 9°28.6′N, 123°40′E | MNHN 200718429 | HF586165 | HF586308 | HF586017 | HF585856 |
| Clade A tenorioi | PANGLAO2005/CP2399 | Bohol Sea, off Balicasag I., Philippines | 309–342 m | 9°31.7′N, 123°41.9′E | MNHN 200718394 | HF586166 | HF586309 | HF586018 | HF585843 |
| Clade A sp 3 | BIOPAPUA/CP3721 | Vitiaz Straight, Papua New Guinea | 542–554 m | 6°03′S, 147°37′E | MNHN 200915183 | HF586156 | HF586299 | HF586008 | HF585780 |
| Clade A sp 4 | NORFOLK2/DW2057 | Norfolk Ridge | 555–565 m | 24°40′S, 168°39′E | MNHN 200917849 | HF586160 | – | HF586013 | – |
| Clade A sp 4 | NORFOLK2/DW2057 | Norfolk Ridge | 555–565 m | 24°40′S, 168°39′E | MNHN 200917850 | HF586161 | HF586304 | HF586014 | HF585850 |
| Clade A sp 4 | NORFOLK2/DW2057 | Norfolk Ridge | 555–565 m | 24°40′S, 168°39′E | MNHN 200917851 | – | – | – | HF585851 |
| Clade A sp 4 | NORFOLK2/DW2057 | Norfolk Ridge | 555–565 m | 24°40′S, 168°39′E | MNHN 200917852 | HF586162 | – | – | HF585852 |
| Clade A sp 5 | AURORA/CP2695 | Philippines | 357–367 m | 14°46′N, 123°40′E | MNHN 200718282 | – | HF586303 | HF586012 | HF585849 |
| Clade A sp 6 | CSIRO RV “Southern Surveyor”/SS1005/012 | Perth Canyon, Western Australia | 479–484 m | 31.92°S, 115.02°E | WAM S25773 | – | HF586298 | – | HF585842 |
| Clade A sp 7 | NORFOLK1/DW1679 | Kaimon Maru Bank, Norfolk Ridge | 298–324 m | 24°43′S, 168°10′E | MNHN 200928739 | – | – | – | HF585844 |
| Clade A sp 7 | NORFOLK1/DW1691 | Eponge Bank, Norfolk Ridge | 509–513 m | 24°54′S, 168°22′E | MNHN 200928740 | – | – | HF586009 | HF585845 |
| Clade B iridescens | – | Off Shionomisaki, Wakayama Pref., Japan | 300 m | 33°24.8′N, 135°42′E | No voucher | EU530041 | – | – | – |
| Clade B sp 2 | MAINBAZA/CP3140 | Maputo transect, Mozambique Channel | 886–898 m | 23°33′S, 36°02′E | MNHN 20098739 | HE800720 | HE800621 | HE800760 | HE800671 |
| Clade B sp 2 | MAINBAZA/CP3140 | Maputo transect, Mozambique Channel | 886–898 m | 23°33′S, 36°02′E | MNHN 20098742 | HF586070 | HF586211 | HF585920 | HF585744 |
| Clade B sp 2 | MAINBAZA/CP3140 | Maputo transect, Mozambique Channel | 886–898 m | 23°33′S, 36°02′E | MNHN 20098744 | HF586071 | HF586212 | HF585921 | HF585745 |
| Clade B sp 3 | T/V Nagasaki-maru, N295/A-1 | SW of Nagasaki, Kyushu I., Japan | 498–503 m | 32º 09′ N, 129º 31′ E | YK1407 | HF586072 | HF586213 | HF585922 | HF585746 |
| Clade C sp. 1 | SALOMON1/CP1804 | Solomon Islands | 309–328 m | 9°32.0′S, 160°37.4′E | MNHN 200718507 | HF586057 | – | HF585906 | HF585857 |
| Clade C sp. 1 | SALOMON1/CP1804 | Solomon Islands | 309–328 m | 9°32.0′S, 160°37.4′E | MNHN 200913304 | – | – | – | HF585759 |
| Clade C sp. 1 | SALOMON1/CP1804 | Solomon Islands | 309–328 m | 9°32.0′S, 160°37.4′E | MNHN 200943074 | – | – | – | HF585724 |
| Clade C sp. 1 | SALOMON1 | Solomon Islands | – | – | MNHN NR | – | – | – | HF585727 |
| Clade C sp. 2 | BOA1/CP2466 | SE Malekula, Vanuatu | 786–800 m | 16°44′S, 167°59′E | MNHN 200718302 | – | – | HF585907 | HF585728 |
| Clade C sp. 2 | EBISCO/CP2651 | SE Fairway, New Caledonia | 883–957 m | 21°29′S, 162°36′E | MNHN 200718318 | HF586058 | – | HF585908 | HF585729 |
| Clade C sp. 2 | SALOMON2/CP2249 | NW Vella, Lavella I., Solomon Islands | 782–884 m | 7°31′S, 156°18′E | MNHN 200913008 | HF586059 | – | HF585909 | HF585730 |
| Clade C sp. 2 | SALOMON2/CP2249 | NW Vella, Lavella I., Solomon Islands | 782–884 m | 7°31′S, 156°18′E | MNHN 200913009 | – | – | – | HF585731 |
| Clade C sp. 3 | PANGLAO2005/CP2398 | Bohol Sea, off Balicasag I., Philippines | 713–731 m | 9°32.6′N, 123°40.5′E | MNHN 200718426 | HF586060 | HF586206 | HF585910 | HF585732 |
| Clade C sp. 4 | SALOMON2/DW2259 | Kolombangara I., Vella Gulf, Solomon Islands | 396–423 m | 8°03.7′S, 156°55.0′E | MNHN 200718539 | HF586061 | HF586207 | HF585911 | HF585733 |
| Clade C sp. 4 | SALOMON2/DW2259 | Kolombangara I., Vella Gulf, Solomon Islands | 396–423 m | 8°03.7′S, 156°55.0′E | MNHN 200943075 | – | – | – | HF585734 |
| Clade C sp. 5 | BIOPAPUA/DW3749 | Seamount off Bougainville, Papua New Guinea | 620–663 m | 5°39′S, 153°59′E | MNHN 200915184 | HF586064 | – | HF585914 | HF585738 |
| Clade C sp. 5 | BIOPAPUA/DW3749 | Seamount off Bougainville, Papua New Guinea | 620–663 m | 5°39′S, 153°59′E | MNHN 200915185 | HF586065 | – | HF585915 | HF585739 |
| Clade C sp. 5 | BIOPAPUA/CP3760 | Off Feni Is, Papua New Guinea | 613–660 m | 3°58′S, 153°43′E | MNHN 200915192 | HF586066 | HF586209 | HF585916 | HF585740 |
| Clade C sp. 5 | BIOPAPUA/CP3740 | Off Woodlark I., Papua New Guinea | 556–645 m | 9°12′S, 152°16′E | MNHN 200915193 | HF586067 | – | HF585917 | HF585741 |
| Clade C sp. 5 | SALOMON1/DW1772 | Solomon Islands | 570–756 m | 8°15.8′S, 160°40.4′E | MNHN 200718508 | HF586062 | – | HF585912 | HF585735 |
| Clade C sp. 5 | SALOMON2/CP2243 | W Vella, Lavella I., Solomon Islands | 518–527 m | 7°42.9′S, 156°27.3′E | MNHN 200718534 | – | – | – | HF585737 |
| Clade C sp. 5 | SALOMON2/CP2243 | W Vella, Lavella I., Solomon Islands | 518–527 m | 7°42.9′S, 156°27.3′E | MNHN 200943073 | HF586063 | HF586208 | HF585913 | HF585736 |
| Clade C sp. 6 | BIOPAPUA/CP3759 | Papua New Guinea | 287–352 m | 4°00′S, 153°36′E | MNHN 200915195 | HF586068 | – | HF585918 | HF585742 |
| Clade C sp. 7 | T/V Nagasaki-maru, N226/Dredge A | SW of Nagasaki, Kyushu I., Japan | 470–487 m | 32º 10′ N, 129º 30′ E | YK1384 | HF586069 | HF586210 | HF585919 | HF585743 |
| Clade C sp. 8 | EBISCO/CP2623 | Lansdowne, New Caledonia | 691–886 m | 20°06′S, 160°19′E | MNHN 200943077 | – | – | – | HF585722 |
| Clade C sp. 8 | TERRASSES/DW3041 | Loyalty Ridge, Mont J, New Caledonia | 800–840 m | 23°59′S, 169°44′E | MNHN 20098874 | HF586055 | – | HF585904 | HF585721 |
| Clade C sp. 8 | TERRASSES/DW3036 | Loyalty Ridge, Walpole, New Caledonia | 800 m | 22°41′S, 168°58′E | MNHN 20098876 | HF586056 | HF586205 | HF585905 | HF585723 |
| Clade C sp. 8 | TERRASSES/DW3045 | Loyalty Ridge, Mont J, New Caledonia | 660–710 m | 23°48′S, 169°46′E | MNHN 20098861 | HE800719 | HE800620 | HE800759 | HE800670 |
| Ilanga 1 | BIOPAPUA/DW3745 | Seamount off Bougainville, Papua New Guinea | 369–377 m | 5°33′S, 154°00′E | MNHN 200915190 | HF586107 | HF586246 | HF585957 | HF585789 |
| Ilanga 1 | BIOPAPUA/DW3745 | Seamount off Bougainville, Papua New Guinea | 369–377 m | 5°33′S, 154°00′E | MNHN 200915197 | HF586108 | HF586247 | HF585958 | HF585790 |
| Ilanga discus | MIRIKY/CP3188 | Between Nosy-bé and Banc du Leven, Madagascar | 298–301 m | 12°31′S, 48°22′E | MNHN 20098758 | HF586109 | HF586248 | HF585959 | HF585791 |
| Ilanga discus | MIRIKY/CP3188 | Between Nosy-bé and Banc du Leven, Madagascar | 298–301 m | 12°31′S, 48°22′E | MNHN 20098760 | HF586110 | HF586249 | HF585960 | HF585792 |
| Ilanga discus | MIRIKY/CP3188 | Between Nosy-bé and Banc du Leven, Madagascar | 298–301 m | 12°31′S, 48°22′E | MNHN 20098761 | HE800724 | HE800625 | HE800764 | HE800675 |
| Ilanga discus | MIRIKY/CP3188 | Between Nosy-bé and Banc du Leven, Madagascar | 298–301 m | 12°31′S, 48°22′E | MNHN 20098776 | HF586111 | HF586250 | HF585961 | HF585793 |
| Ilanga discus | MIRIKY/CP3188 | Between Nosy-bé and Banc du Leven, Madagascar | 298–301 m | 12°31′S, 48°22′E | MNHN 20098777 | HF586112 | HF586251 | HF585962 | HF585794 |
| Ilanga 3 | BOA1/CP2413 | Malo I., Vanuatu | 268–445 m | 15°42′S, 167°02′E | MNHN 200718301 | HF586114 | HF586253 | HF585964 | HF585796 |
| Ilanga 4 | T/V Nagasaki-maru, N295/AA | E of Fukue I., Goto Is, Kyushu, Japan | 235–238 m | 32º 30′ N, 129º 08′ E | YK1380 | HF586150 | HF586291 | HF586000 | HF585837 |
| Ilanga 4 | R/V Tansei-maru, KT-11-12/T5 | Off Cape Toi, Miyazaki, Kyushu I., Japan | 207–216 m | 31º 09′ N, 131º 26′ E | YK1485 | – | HF586292 | – | – |
| Ilanga 4 | TAIWAN2001/CP76 | Off Tashi, NE Coast of Taiwan | 115–170 m | 24°57′N, 122°02′E | MNHN 200718548 | – | – | – | HF585803 |
| Ilanga 5 | CONCALIS/CP2961 | Grand Passage, New Caledonia | 220–390 m | 19°04′S, 163°11′E | MNHN 200735552 | HF586122 | HF586263 | HF585973 | HF585806 |
| Ilanga 5 | CONCALIS/CP2961 | Grand Passage, New Caledonia | 220–390 m | 19°04′S, 163°11′E | MNHN 200735578 | HF586123 | HF586264 | HF585974 | HF585807 |
| Ilanga 5 | CONCALIS/CP2961 | Grand Passage, New Caledonia | 220–390 m | 19°04′S, 163°11′E | MNHN 200735579 | HF586124 | HF586265 | HF585975 | HF585808 |
| Ilanga 5 | CONCALIS/CP2961 | Grand Passage, New Caledonia | 220–390 m | 19°04′S, 163°11′E | MNHN 200735584 | HF586125 | HF586266 | HF585976 | HF585809 |
| Ilanga 6 | SANTO2006/AT112 | W Tutuba I., Vanuatu | 150–168 m | 15°33.5′S, 167°16.1′E | MNHN 200718446 | HF586127 | HF586268 | HF585978 | HF585812 |
| Ilanga 6 | SANTO2006/– | Vanuatu | No data | No data | MNHN 200718447 | HF586128 | HF586269 | HF585979 | HF585813 |
| Ilanga cf. norfolkensis | CONCALIS/CP3010 | Grand Passage, New Caledonia | 603 m | 18°46′S, 163°19′E | MNHN 200735580 | HF586131 | HF586272 | HF585982 | HF585816 |
| Ilanga cf. norfolkensis | CONCALIS/CP3010 | Grand Passage, New Caledonia | 603 m | 18°46′S, 163°19′E | MNHN 200735581 | HF586132 | HF586273 | HF585983 | HF585817 |
| Ilanga cf. norfolkensis | EBISCO/DW2603 | Chesterfield, New Caledonia | 568–570 m | 19°36′S, 158°43′E | MNHN 200718324 | HF586129 | HF586270 | HF585980 | HF585814 |
| Ilanga cf. norfolkensis | EBISCO/DW2603 | Chesterfield, New Caledonia | 568–570 m | 19°36′S, 158°43′E | MNHN 200718325 | HF586130 | HF586271 | HF585981 | HF585815 |
| Ilanga biradiatula | MAINBAZA/CP3135 | Maputo transect, Mozambique Channel | 480–503 m | 25°13′S, 35°18′E | MNHN 20098740 | HE800723 | HE800624 | HE800763 | HE800674 |
| Ilanga biradiatula | MAINBAZA/CP3135 | Maputo transect, Mozambique Channel | 480–503 m | 25°13′S, 35°18′E | MNHN 20098741 | HF586133 | HF586274 | HF585984 | HF585818 |
| Ilanga biradiatula | MAINBAZA/CP3135 | Maputo transect, Mozambique Channel | 480–503 m | 25°13′S, 35°18′E | MNHN 20098743 | HF586134 | HF586275 | – | HF585819 |
| Ilanga biradiatula | MIRIKY/CP3184 | Between Nosy-bé and Banc du Leven, Madagascar | 492–524 m | 12°40′S, 48°12′E | MNHN 20098759 | HF586135 | HF586276 | HF585985 | HF585820 |
| Ilanga 9 | EBISCO/CP2571 | N Bellona, New Caledonia | 298–309 m | 20°25′S, 158°45′E | MNHN 200718314 | HF586137 | HF586278 | HF585987 | HF585822 |
| Ilanga 9 | EBISCO/CP2571 | N Bellona, New Caledonia | 298–309 m | 20°25′S, 158°45′E | MNHN 200718315 | HF586138 | HF586279 | HF585988 | HF585823 |
| Ilanga 9 | EBISCO/DW2618 | Lansdowne, New Caledonia | 280–304 m | 20°06′S, 160°23′E | MNHN 200718326 | HF586139 | HF586280 | HF585989 | HF585824 |
| Ilanga 9 | EBISCO/DW2618 | Lansdowne, New Caledonia | 280–304 m | 20°06′S, 160°23′E | MNHN 200718327 | HF586140 | HF586281 | HF585990 | HF585825 |
| Ilanga 9 | EBISCO/DW2618 | Lansdowne, New Caledonia | 280–304 m | 20°06′S, 160°23′E | MNHN 200718328 | HE800725 | HE800626 | HE800765 | HE800676 |
| Ilanga 9 | EBISCO/CP2571 | N Bellona, New Caledonia | 298–309 m | 20°25′S, 158°45′E | MNHN 200734080 | HF586141 | HF586282 | HF585991 | HF585826 |
| Ilanga 10 | NORFOLK2/DW2135 | Norfolk Ridge, Banc Munida, New Caledonia | 295–330 m | 23°02′S, 168°21′E | MNHN 200718340 | – | – | – | HF585827 |
| Ilanga 10 | TERRASSES/CP3092 | SE Terrasses, New Caledonia | 360–380 m | 22°13′S, 167°12′E | MNHN 20098797 | – | – | – | HF585828 |
| Ilanga 10 | TERRASSES/CP3092 | SE Terrasses, New Caledonia | 360–380 m | 22°13′S, 167°12′E | MNHN 20098798 | HF586142 | HF586283 | HF585992 | HF585829 |
| Ilanga 10 | TERRASSES/CP3092 | SE Terrasses, New Caledonia | 360–380 m | 22°13′S, 167°12′E | MNHN 20098799 | HF586143 | HF586284 | HF585993 | HF585830 |
| Ilanga 10 | TERRASSES/CP3092 | SE Terrasses, New Caledonia | 360–380 m | 22°13′S, 167°12′E | MNHN 20098800 | HF586144 | HF586285 | HF585994 | HF585831 |
| Ilanga 10 | TERRASSES/CP3087 | SE Terrasses, New Caledonia | 380–400 m | 22°11′S, 167°12′E | MNHN 20098805 | HF586145 | HF586286 | HF585995 | HF585832 |
| Ilanga 10 | TERRASSES/CP3087 | SE Terrasses, New Caledonia | 380–400 m | 22°11′S, 167°12′E | MNHN 20098806 | HF586146 | HF586287 | HF585996 | HF585833 |
| Ilanga 10 | TERRASSES/CP3087 | SE Terrasses, New Caledonia | 380–400 m | 22°11′S, 167°12′E | MNHN 20098807 | HF586147 | HF586288 | HF585997 | HF585834 |
| Ilanga 10 | TERRASSES/CP3087 | SE Terrasses, New Caledonia | 380–400 m | 22°11′S, 167°12′E | MNHN 20098808 | HF586148 | HF586289 | HF585998 | HF585835 |
| Ilanga 10 | TERRASSES/DW3079 | SE Terrasses, Passe de la Sarcelle, New Caledonia | 300–420 m | 22°28′S, 167°29′E | MNHN 20098809 | HF586149 | HF586290 | HF585999 | HF585836 |
| Ilanga 11 | MAINBAZA/CP3143 | Maputo transect, Mozambique Channel | 264–277 m | 23°32′S, 35°46′E | MNHN 200915174 | HF586106 | – | HF585956 | HF585890 |
| Ilanga 12 | PANGLAO2004/T27 | Between Panglao I. and Pamilacan I., Philippines | 106–137 m | 9°33.4′N 123°51.0′E | MNHN 200718221 | – | – | – | HF585811 |
| Ilanga 12 | PANGLAO2004/T27 | Between Panglao I. and Pamilacan I., Philippines | 106–137 m | 9°33.4′N 123°51.0′E | MNHN 200913303 | HF586126 | HF586267 | HF585977 | HF585810 |
| Ilanga gotoi | PANGLAO2004/T31 | Between Panglao I. and Balicasag I., Philippines | 100–140 m | 9°33.0′N, 123°42.0′E | MNHN 200718349 | HF586136 | HF586277 | HF585986 | HF585821 |
| Ilanga laevissima | NMDP Africana/St A 18178 D | S of Tsitsikamma, W Cape, South Africa | 115 m | 34°25′S, 24°00′E | NMSA V3139 | – | – | HF586001 | – |
| Ilanga laevissima | NMDP Africana/St A 18994 D | Plettenberg Bay, S Cape, South Africa | 104 m | 34°19.5′S, 23°30′E | NMSA V4397 | HF586151 | HF586293 | HF586003 | HF585788 |
| Ilanga 15 | BIOPAPUA/CP3759 | Off Feni Is, Papua New Guinea | 287–352 m | 04°00′S, 153°36′E | MNHN 200915196 | HF586113 | HF586252 | HF585963 | HF585795 |
| Ilanga 16 | TERRASSES/DW3094 | SE Terrasses, New Caledonia | 250–300 m | 22°04′S, 167°03′E | MNHN 20098801 | HF586121 | HF586262 | HF585972 | HF585805 |
| Ilanga 17 | PANGLAO 2005/CP2393 | Bohol Sea, off Balicasag I., Philippines | 356–396 m | 9°30′N, 123°42′E | MNHN 200735011 | – | HF586260 | – | – |
| Ilanga 17 | PANGLAO2005/CP2332 | Bohol Sea, Maribojoc Bay, Philippines | 584–596 m | 9°38.2′N, 123°43.5′E | MNHN 200718416 | HF586115 | HF586254 | HF585965 | HF585797 |
| Ilanga 17 | PANGLAO2005/CP2331 | Bohol Sea, Maribojoc Bay, Philippines | 255–268 m | 9°39.2′N, 123°47.5′E | MNHN 200718417 | HF586116 | HF586255 | HF585966 | HF585798 |
| Ilanga 17 | PANGLAO2005/CP2340 | Bohol Sea, off Balicasag I., Philippines | 271–318 m | 9°29.4′N, 123°44.4′E | MNHN 200718418 | HF586117 | HF586256 | HF585967 | HF585799 |
| Ilanga 17 | PANGLAO2005/CP2340 | Bohol Sea, off Balicasag I., Philippines | 271–318 m | 9°29.4′N, 123°44.4′E | MNHN 200718419 | HF586118 | HF586257 | HF585968 | HF585800 |
| Ilanga 17 | PANGLAO2005/CP2340 | Bohol Sea, off Balicasag I., Philippines | 271–318 m | 9°29.4′N, 123°44.4′E | MNHN 200718420 | HF586119 | HF586258 | HF585969 | HF585801 |
| Ilanga 17 | PANGLAO2005/CP2344 | Bohol Sea, off Pamilacan I., Philippines | 128–142 m | 9°28.4′N, 123°50.1′E | MNHN 200718421 | HF586120 | HF586259 | HF585970 | HF585802 |
| Ilanga 17 | PANGLAO2005/CP2381 | Sill between Bohol and Sulu Seas, Dipolog Bay, Philippines | 259–280 m | 8°43′N, 123°19′E | MNHN 200735122 | – | HF586261 | HF585971 | HF585804 |
| Ilanga 18 | NMDP (Africana)/St A 18178 D | S of Tsitsikamma, W Cape, South Africa | 115 m | 34°25′S, 24°00′E | NMSA V3139 | – | – | HF586002 | – |
| Ilanga 19 | – | Off Shionomisaki, Wakayama Pref., Japan | 210 m | 33°25′N, 135°41.7′E | No voucher | EU530040 | EU530141 | – | – |
| Ilanga 20 | SALOMON2/CP2287 | E Rendova I., Solomon Islands | 253–255 m | 8°40.8′S, 157°24.6′E | MNHN 200718536 | – | – | – | HF585785 |
| Ilanga 20 | SALOMON2/CP2287 | E Rendova I., Solomon Islands | 253–255 m | 8°40.8′S, 157°24.6′E | MNHN 200913307 | – | – | HF585955 | HF585883 |
| Hazuregyra watanabei | R/V Wakataka-maru, Leg. 3/EF350 | Off Kinkazan, Miyagi, Honshu I., Japan | 350 m | 37º 59′ N, 141º 59′ E | YK1464 | HF586105 | HF586245 | HF585954 | HF585784 |
| “Machaeroplax” delicatus | R/V Tansei-maru, KT-11-12/T10-2 | Off Cape Toi, Miyazaki, Kyushu I., Japan | 1063–1082 m | 31º 07′ N, 131º 39′ E | YK1484 | HF586197 | HF586342 | HF586048 | HF585896 |
| Minolia nyssonus | – | Off Kanaya, Chiba, Honshu I., Japan | c. 150–200 m | 35º 11′ N, 139º 47′ E | YK1386 | – | HF586295 | – | – |
| Minolia nyssonus | T/V Seisui-maru, 96-05/D-4 | E of Daiozaki, Mie, Honshu I., Japan | 263 m | 34º 17′ N, 137º 10′ E | YK1355 | HF586152 | HF586294 | HF586004 | HF585838 |
| Minolia punctata | – | Off Misaki, Kanagawa, Honshu I., Japan | 80 m | 35º 09′ N, 139º 35′ E | YK1379 | HF586155 | HF586297 | HF586007 | HF585841 |
| Minolia sp. | – | Off Misaki, Kanagawa, Honshu I., Japan | 80 m | 35º 09′ N, 139º 35′ E | YK0205 | HF586154 | AB365226 | HF586006 | HF585840 |
| Minolia sp. | – | Off Zyogashima, Miura, Kanagawa Pref., Japan | – | – | No voucher | HF586153 | HF586296 | HF586005 | HF585839 |
| Solariella affinis | R/V “Harry Borthen” | Møre og Romsdal county, Vanylven, Rovdefjorden, NE of Kropperevet, Norway | 150–200 m | 62°11.45′N, 5°34′E | No voucher | – | – | – | HF585871 |
| Solariella affinis | R/V “Harry Borthen” | Møre og Romsdal county, Vanylven, Rovdefjorden, NE of Kropperevet, Norway | 150–200 m | 62°11.45′N, 5°34′E | NHMUK 20120233 | – | HF586321 | HF586029 | HF585872 |
| Solariella affinis | R/V “Harry Borthen” | Møre og Romsdal county, Vanylven, Rovdefjorden, NE of Kropperevet, Norway | 150–200 m | 62°11.45′N, 5°34′E | NHMUK 20120234 | – | – | – | HF585873 |
| Solariella segersi | PANGLAO2005/CP2344 | Bohol Sea, off Balicasag I., Philippines | 128–142 m | 9°28.4′N, 123°50.1′E | MNHN 200718422 | HF586177 | HF586322 | HF586030 | HF585875 |
| Solariella segersi | PANGLAO2005/CP2344 | Bohol Sea, off Balicasag I., Philippines | 128–142 m | 9°28.4′N, 123°50.1′E | No voucher | HF586178 | HF586323 | HF586031 | – |
| Solariella chodon | AURORA/CP2712 | Philippines | 139–140 m | 15°20′N, 121°30′E | MNHN 200718289 | HF586179 | HF586324 | HF586032 | – |
| Solariella chodon | PANGLAO2004/T26 | Boholi I., Cortes, Philippines | 123–135 m | 9°43.3′N, 123°48.8′E | MNHN 200718348 | HF586180 | HF586325 | HF586033 | – |
| Solariella 3 | TERRASSES/DW3109 | Munida, Norfolk Ridge, New Caledonia | 150–180 m | 23°01′S, 168°18′E | MNHN 20098857 | HF586182 | HF586327 | HF586035 | HF585877 |
| Solariella 3 | TERRASSES/DW3109 | Munida, Norfolk Ridge, New Caledonia | 150–180 m | 23°01′S, 168°18′E | MNHN 20098858 | HF586183 | HF586328 | – | HF585878 |
| Solariella 3 | TERRASSES/DW3109 | Munida, Norfolk Ridge, New Caledonia | 150–180 m | 23°01′S, 168°18′E | MNHN 20098859 | HF586184 | HF586329 | HF586036 | HF585879 |
| Solariella 3 | TERRASSES/DW3109 | Munida, Norfolk Ridge, New Caledonia | 150–180 m | 23°01′S, 168°18′E | MNHN 20098860 | HF586185 | HF586330 | HF586037 | HF585880 |
| Solariella 4 | MAINBAZA/CC3163 | Inhambane transect, Mozambique Channel | 406–410 m | 24°09′S, 35°42′E | MNHN 200915171 | HF586192 | – | HF586043 | HF585894 |
| Solariella 4 | MAINBAZA/CC3163 | Inhambane transect, Mozambique Channel | 406–410 m | 24°09′S, 35°42′E | MNHN 200915172 | – | – | – | HF585888 |
| Solariella 4 | MAINBAZA/CC3163 | Inhambane transect, Mozambique Channel | 406–410 m | 24°09′S, 35°42′E | MNHN 200915173 | – | – | – | HF585889 |
| Solariella 4 | MAINBAZA/CP3135 | Maputo transect, Mozambique Channel | 480–503 m | 25°13′S, 35°18′E | MNHN 20098745 | HF586191 | HF586337 | HF586042 | HF585887 |
| Solariella 4 | MAINBAZA/CP3135 | Maputo transect, Mozambique Channel | 480–503 m | 25°13′S, 35°18′E | MNHN 20098747 | HF586193 | – | HF586044 | HF585892 |
| Solariella dedonderorum | PANGLAO2005/DW2400 | Bohol Sea, off Balicasag I., Philippines | 111–115 m | 9°32.5′N, 123°41.8′E | MNHN 200718427 | HF586181 | HF586326 | HF586034 | HF585876 |
| Solariella 6 | SALOMON2/DW2169 | Russel I., W Bay, Solomon Islands | 100–200 m | 9°01.1′S, 159°5.7′E | MNHN 200718537 | – | HF586338 | – | HF585891 |
| Solariella 7 | BERYX/DW18 | New Caledonia | 250–270 m | 24°48′S, 168°09′E | MNHN NR | – | – | – | HF585874 |
| “Solariella”varicosa | R/V “Asterias” | Finnmark county, Varangerfjorden, SW of Vestre Jakobselv, Norway | 10–174 m | 70°4′N, 29°12′E | NHMUK 20120235 | – | – | – | HF585720 |
| Spectamen 1 | PANGLAO2004/T39 | W Pamilacan I., Cervera Shoal, Philippines | 100–138 m | 9°30.1′N, 123°50.4′E | MNHN 200718351 | HF586186 | HF586331 | HF586038 | HF585881 |
| Spectamen 2 | T/V Nagasaki-maru/N295, Dredge 1 | W of Takarajima I., Tokara Is, Japan | 183–184 m | 29º 25′ N, 127º 18′ E | YK1381 | HF586189 | HF586335 | HF586040 | HF585885 |
| Spectamen laevior | PANGLAO2005/CP2344 | Bohol Sea, off Balicasag I., Philippines | 128–142 m | 9°28.4′N, 123°50.1′E | MNHN 200718428 | HF586187 | HF586332 | HF586039 | HF585882 |
| Spectamen laevior | PANGLAO2005/CP2344 | Bohol Sea, off Balicasag I., Philippines | 128–142 m | 9°28.4′N, 123°50.1′E | MNHN 200913305 | – | HF586333 | – | HF585726 |
| Spectamen 4 | CSIRO RV “Southern Surveyor”/SS1005/042 | Off Bald I., Western Australia, Australia | 973–999 m | 35°16.11′S, 118°43.12′ | WAM S25789 | – | HF586318 | – | HF585868 |
| Spectamen 4 | CSIRO RV “Southern Surveyor”/SS1005/042 | Off Bald I., Western Australia, Australia | 973–999 m | 35°16.11′S, 118°43.12′E | WAM S25789 | HF586175 | HF586319 | HF586027 | HF585869 |
| Spectamen mutabilis | AURORA/CP2695 | Philippines | 357–367 m | 14°46′N, 123°40′E | MNHN 200718288 | HE800727 | HE800627 | HE800767 | HE800678 |
| Spectamen mutabilis | AURORA/CP2695 | Philippines | 357–367 m | 14°46′N, 123°40′E | MNHN 200928738 | HF586188 | HF586334 | – | HF585884 |
| Spectamen mutabilis | T/V Nagasaki-maru/N319, St. G3 | W of Kusagaki Is, Kyushu, Japan | 298–299 m | 30°39′N, 127°54′E | YK1462 | HF586190 | HF586336 | HF586041 | HF585886 |
| Spectamen philippensis | – | N Moreton I., Moreton Bay, Queensland, Australia | 31 m | 26°56.6′S, 153°24.2′E | NHMUK 20110452 | EU530042 | – | HE800766 | – |
| Spectamen philippensis | – | N Moreton I., Moreton Bay, Queensland, Australia | 31 m | 26°56.6′S, 153°24.2′E | NHMUK 20110452 | HF586176 | HF586320 | HF586028 | HF585870 |
| Spectamen philippensis | – | N Moreton I., Moreton Bay, Queensland, Australia | 31 m | 26°56.6′S, 153°24.2′E | NHMUK 20110452 | – | – | – | HE800677 |
| Suavatrochus sp | T/V Nagasaki-maru, N295/R-2(3) | W of Amami I., Japan | 704–730 m | 28 36′N, 127 04′E | YK1382 | HF586198 | HF586343 | HF586049 | HF585897 |
| Zetela 1 | MAINBAZA/CP3138 | Maputo transect, Mozambique Channel | 700–707 m | 25°13′S, 35°21′E | MNHN 20098748 | HF586195 | HF586341 | HF586047 | HF585895 |
| Zetela 1 | MAINBAZA/CP3138 | Maputo transect, Mozambique Channel | 700–707 m | 25°13′S, 35°21′E | MNHN 200915167 | HF586194 | HF586339 | HF586045 | HF585786 |
| Zetela 1 | MAINBAZA/CP3138 | Maputo transect, Mozambique Channel | 700–707 m | 25°13′S, 35°21′E | MNHN 200915169 | – | HF586340 | HF586046 | HF585893 |
| Zetela 2 | ANDEEP III/PS67/074-6-E | Eastern Weddell Sea, Antarctica | 1030 m | 71°18.35′S, 13°57.71′W | NHMUK 20120236 | HF586050 | HF586199 | HF585898 | HF585714 |
| Zetela 3 | BIOPEARL II/BIO6-AGT-2B | Amundsen Sea, Antarctica | 984–1000 m | 71°10′S, 109°53′W | NHMUK 20120237 | HF586052 | HF586201 | HF585900 | HF585716 |
| Zetela 3 | LAMPOS ANDEEP/150-1 | Burdwood Bank, Antarctica | 286–290 m | 54°30.22′S, 56°8.2′W | NHMUK 20120238 | HF586051 | HF586200 | HF585899 | HF585715 |
| Zetela 3 | LAMPOS ANDEEP/150-1 | Burdwood Bank, Antarctica | 286–290 m | 54°30.22′S, 56°8.2′W | No voucher | – | HF586202 | HF585901 | HF585717 |
| Zetela 3 | LAMPOS ANDEEP/150-1 | Burdwood Bank, Antarctica | 271–272 m | 54°1.36′S, 62°1.33′W | NHMUK 20120239 | HF586053 | HF586203 | HF585902 | HF585718 |
| Zetela 3 | LAMPOS ANDEEP/150-1 | Burdwood Bank, Antarctica | 286–290 m | 54°30.22′S, 56°8.2′W | NHMUK 20120240 | HF586054 | HF586204 | HF585903 | HF585719 |
| Zetela kopua | RV Tangaroa/2003209 | Seamount WNW of Three Kings Is, New Zealand | 1145–1185 m | 34°2.9′S, 171°8.2′E | NMNZ M160804 | HF586196 | – | – | – |
MNHN, Muséum National d'Histoire Naturelle, Paris; NHMUK, Natural History Museum, London; NMNZ, Museum of New Zealand Te Papa Tongarewa, Wellington; NMSA, KwaZulu-Natal Museum, South Africa; WAM, Western Australian Museum, Perth; YK, personal collection of Yasunori Kano; NR, not registered; GB, GenBank. Note new, corrected locality data for GenBank samples used in Williams et al. (2008). Details for outgroup taxa in Table S1.
Laboratory methods, sequence editing, and alignment
DNA was extracted from ethanol-preserved foot or mantle tissue (or in a few cases dried specimens) following the protocol described by Williams and Ozawa (2006). The amplification protocols described by Williams et al. (2010) were used to amplify portions of the nuclear 28S rRNA gene (28S: 1496 bp) and three mitochondrial genes: cytochrome oxidase subunit I (COI: 709 bp), 16S rRNA (16S: ∼610 bp) and 12S rRNA (12S: ∼685 bp). Sequence reactions were performed directly on purified PCR products using a BigDye Terminator v1.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA) and run on an Applied Biosystems 3730 DNA Analyser automated capillary sequencer. Sequencing and PCR primers are listed in Table S2. Sequences were edited using Sequencher (v. 4.8, Gene Codes Corporation, Ann Arbor, Michigan). A total of 670 sequences were analyzed in this study, of which 631 were new (EMBL accession numbers in Table 1).
Alignment of solariellid COI sequences was performed in MacClade (v 4.08 OSX; Maddison and Maddison 2003). Alignment of COI including outgroups required two insertions, each of a single amino acid for Liotiidae sequences (as previously noted by Kano 2008 and Williams 2012). Ribosomal genes were aligned using MAFFT (v 6.864; Katoh et al. 2002; online: http://mafft.cbrc.jp/alignment/server/). The G-INS-i option was used, which is recommended for sequences with global homology (Katoh et al. 2005), the gap opening penalty was set to 1 and the offset value was set at 0.1, as long gaps were not expected. Scoring matrix for nucleotide sequences were set to “1PAM/κ = 2” for 28S as sequences were very similar, but “20PAM/κ = 2” for mitochondrial ribosomal genes. Poorly aligned sites in rRNA alignments were identified using Gblocks Server (0.91b, Castresana 2000; http://molevol.cmima.csic.es/castresana/Gblocks_server.html) and removed from analyses. Parameters used in Gblocks allowed for smaller final blocks, gap positions within the final blocks and less strict flanking positions.
Species delimitation
We used the single-threshold, general mixed Yule-coalescent (GMYC) model as implemented by SPLITS (code written by T. Ezard, T. Fujisawa and T. Barraclough in R, v.2.10, http://cran-project) to identify species from sequence variation in mitochondrial genes. We used COI on its own, as COI is commonly used as a “barcoding” gene, but we also used concatenated sequences from all three mitochondrial genes as a previous study on low dispersal species has suggested that combined genes may be more informative than a single gene for species delimitation (Williams et al. 2011). We did not use 28S as the GMYC procedure provides a potential means of detecting species from single-locus sequence data (Monaghan et al. 2009). Instead, we examined the 28S sequences to determine whether any species shared identical genotypes.
Taxon sets differed between the two GMYC analyses. In the combined mitochondrial gene analysis, we used concatenated sequence from all mitochondrial genes including those specimens with two or three mitochondrial sequences. Where preliminary analyses showed sequences formed a tight cluster in independent gene trees, samples from each clade were limited to three specimens in the combined dataset. This dataset included some species that were missing COI data. All individuals with COI sequences were included in the single gene analysis. Eleven specimens were not included in either analysis because of missing data.
Ultrametric trees were produced for GMYC analyses using Bayesian inference as implemented in the program BEAST (v.1.7.1; Drummond and Rambaut 2007) with a relaxed lognormal clock, but without any fossil calibrations and a fixed mean rate of substitutions set to one. We used a constant coalescent prior, which is thought to be more conservative than a Yule prior for delimiting species (Monaghan et al. 2009). Where multiple genes were used, sequence variation was partitioned among genes and gene-specific nucleotide substitution model parameters were used, with each gene allowed to evolve at a different rate. Nucleotide substitution models used in preliminary analyses in BEAST were determined by MrModelTest using the hierarchical likelihood ratio test (v 2.1, J. Nylander, http://www.ebc.uu.se/systzoo/staff/nylander.html). Where multiple models were suggested, the simplest was chosen. The best models for each data set were determined to be HKY + I + G for 16S and 12S and GTR + I + G for COI. Analyses ran for 200,000,000 generations, sampling every 10,000 generations. The final species tree was a maximum clade credibility tree with median node heights based on 18,000 trees. Length of burnin was determined by examination of traces in Tracer (v. 1.5, Drummond and Rambaut 2007; available from http://beast.bio.ed.ac.uk/Tracer).
Phylogenetic reconstruction
Species trees using individual genes and concatenated sequences from all four genes were produced using Bayesian inference as implemented in MrBayes (v. 3.2.1, Huelsenbeck and Ronquist 2001). Nucleotide substitution models were those used in species delimitation analyses (plus 28S: GTR + G + I). The temperature was lowered to 0.15 to encourage swapping among chains and the propset command was used to increase the proposal probability of the topology parameter (individual gene datasets: ExtTBR(Tau,V); combined dataset: ExtTBR(Tau{all},V{all})) from 5% to 10%. These parameters were chosen based on previous studies of Trochoidea, which showed a large improvement in convergence time and effective sample size (ESS) values using these settings (Williams 2012). Analyses were run for 20,000,000 generations with a sample frequency of 1000. The first ten percent were discarded, so that 18,000 trees were accepted for each run. The datasets were analyzed in two independent runs, and the final tree was computed from the combination of accepted trees from each run (a total of 36,000 trees). Stationarity and convergence between the two runs were determined by examining the potential scale reduction factors (PSRF), standard deviation of split frequencies and by visual examination of.p files in Tracer (v. 1.5; available from http://beast.bio.ed.ac.uk/Tracer).
A chronogram, where branch length corresponds to time, was produced using Species Tree Ancestral Reconstruction (*BEAST). The *BEAST method co-estimates gene trees and a species tree and allows for the incorporation of multiple exemplars of each species and the independent evolution of each gene without fixing a single topology across loci (Heled and Drummond 2010). Two separate *BEAST analyses were undertaken to test how calibrations affect node ages. In one analysis we used an uncorrelated relaxed, lognormal clock with three calibrations based on fossil evidence. In the second, only one was used to date the root (see below for details). Eight independent *BEAST analyses ran between 322,000,000 and 500,000,000 generations with sample frequency of 10,000 for the three calibration analysis. Five independent runs were used in the single calibration analysis. In both cases, the Birth–Death tree prior was used for species-level analyses. Sequence variation was partitioned among genes and gene-specific nucleotide substitution model parameters were used, with each gene allowed to evolve at a different rate. Based on preliminary analyses, we simplified the nucleotide substitution models, using HKY + G + I for all genes, which resulted in improved ESS values. In the *BEAST analysis, we used only solariellid sequences, where each individual had sequence data for 28S and at least two mitochondrial gene sequences. Sequences were assigned to 68 species (not all species were included due to missing data) based on results from species delimitation tests and the number of individuals per species was limited to three to improve computation times. Tree topology was linked for the three mitochondrial genes, as the mitochondrial genome is inherited as a single locus. Default priors were used except for fossil calibrations and ucld.mean priors, which were changed to exponential.
As ages can vary between BEAST and *BEAST analyses (e.g., McCormack et al. 2011), we also ran two analyses with BEAST. As with the *BEAST, one had all three calibrations and one had only the root calibrated. The BEAST analyses ran for 100,000,000 generations with sampling every 10,000 generations. A Birth–Death prior with incomplete sampling was used, with each of the 68 included species represented by a single specimen. Sequences were concatenated and a single tree was produced for the four genes. Substitution models were the same as in *BEAST, but lognormal priors were used for ucld.mean priors.
The final *BEAST species trees and BEAST trees were maximum clade credibility trees with mean node heights based on the remaining trees after burnin of <13% trees in each run. Length of burnin was determined by examination of traces in Tracer.
Diversification
Plots of the log of the number of lineages against node height (“lineages through time”; LTT) were used to illustrate the rate of diversification using Laser (Rabosky 2006) in R (v. 2.15.0). We used the Constant Rate (CR) test with the gamma-statistic of Pybus and Harvey (2000) to determine whether the LTT plots were consistent with a constant net rate of diversification through time. Allowance was made for incomplete taxon sampling by drawing significance values from simulations using a Monte Carlo constant-rate Test (MCCR; Pybus and Harvey 2000) as implemented in Laser (in R). Sampling was incomplete in this study and it is not known exactly how many species are missing. For instance, species ranges are often quite small, so we assume that sampling in new areas would likely result in the discovery of new species. Moreover, only the IWP was intensively sampled and we are missing species from the Atlantic. To address this issue, we used a range of numbers for the total number of solariellids (100, 200, 300, 600, and 6000) that was likely to encompass the true number of species (we estimate the real number of species in Solariellidae is likely to be closer to 300 species than 100 or 6000).
Three alternative models of lineage accumulation were also used to test the distribution of speciation events over time using models described by Paradis (1997) implemented by the Analyses of Phylogenetics and Evolution package (APE; v. 3.0–5, in R). Model A assumes a constant rate of diversification over time and Model B assumes a gradual change in diversification over time and permits calculation of the parameter β. Values of β < 1 indicate that diversification is increasing, either as a result of increased rates of speciation or decreased rates of extinction, whereas values of β > 1 suggest that diversification is slowing down. Model C assumes that there are two distinct rates of diversification, each with its own rate of speciation before (δ1) and after (δ2) a defined point in time (Tc). We also used the relative cladogenesis test (Purvis et al. 1995) as implemented in R (Geiger package; Harmon et al. 2008) to identify nodes with a significantly increased rate of diversification.
Speciation rates for two genera (Ilanga and Bathymophila) were calculated using equations from Magallon and Sanderson (2000) as implemented in R (Geiger package). Bathymophila was chosen as a clade of interest because its species are distributed in intermediate to deep water. Species in this genus fall into two clades, and all species examined to date in one of these clades are sightless (see Discussion for details). Ilanga was chosen as a comparison to Bathymophila, because it is a shallow to intermediate depth clade and all species examined to date have pigmented eyes (Herbert 1987; this study). From literature reports, we know that at least 12 species of Ilanga (Herbert 1987) and six species of Bathymophila were not included in this study (Marshall 1999; Vilvens 2009; S. T. Williams and C. Vilvens, unpubl. data). Even so, the total number of species in either clade is not known, although Ilanga overall is likely better sampled than Bathymophila.
Fossil calibrations
We used three fossil records to calibrate the chronograms. In each case, the oldest recognizable member of a clade was used to date the node at the base of the crown group. The lower bound of the age range of a fossil gave the minimum age of the node, while the maximum was estimated as the lower bound of two stages older, which allows both for the uncertainty of the fossil age, and for the incompleteness of the fossil record.
The oldest recorded fossil we could unambiguously compare with Recent Solariellidae was “Solariella” montsecana from the Campanian of Torallola, Spain (Kiel and Bandel 2001). This species has axial ribs on the first teleoconch whorl and is quite similar to some specimens in Clade A, but it has axial ribs in the umbilicus and species sampled to date in Clade A do not, so it likely represents a separate genus. We used this fossil record to calibrate the age of the entire ingroup. The clade was constrained to be at least 71 Ma (95% interval: 71.4–89 Ma; mean in real space: 4.18, log stdev: 1, offset: 71).
The second calibration was based on Solariella sp. from the latest Oligocene part of the Lincoln Creek Formation in western Washington State, United States of America (Fig. 3, Kiel 2010). This species is similar to S. affinis so was used to date the crown of the clade including S. affinis, here referred to as Solariella. The Solariella clade was constrained to be at least 23 Ma (95% interval: 23.2–34 Ma; mean in real space: 2.555, log stdev: 1, offset: 23).
Figure 3.

Combined gene tree based on Bayesian inference using MrBayes for Solariellidae using four genes (28S, 16S, 12S and cytochrome oxidase subunit I [COI]), with outgroups removed for clarity (see Figure S3 for outgroup relationships). Support values are posterior probabilities (PP); branches with PP < 50% were collapsed, PP not shown for intraspecific relationships. See Table 1 for sample details. Monophyletic clades discussed in the text are indicated with a gray shaded box. Type species are in bold font. Note that species in Clade C are described by Vilvens and Williams (2013) and assigned to a new genus. Photos are of exemplar species from each clade: (a) Ilanga biradiatula; (b) “Solariella” varicosa; (c) Suavotrochus sp.; (d) “Machaeroplax” delicatus; (e) Clade C sp. 8; (f) Minolia sp.; (g) Clade B sp. 2; (h) Zetela 1; (i) Archiminolia 2; (j) Clade A sp. 5; (k) Solariella affinis; (l) Spectamen philippensis; (m) Bathymophila 7.
The third calibration was based on Zetela awamoana Laws 1939, from the Mount Harris Formation, South Island, New Zealand (Beu and Raine 2009); this fossil is from the Altonian stage of the New Zealand time scale, corresponding to the later half of the Burdigalian (early Miocene) of the international time scale (Hollis et al. 2010). Only one nominal species of Zetela, Z. kopua, was included in this study. Unfortunately sequence was obtained only from 28S for this specimen, so it was not included in the dated analyses. In the 28S tree, it was sister to an undescribed species from Madagascar (Mainbaza expedition) that based on shell characters we would assign to Lamellitrochus, which is a probable synonym of Zetela Marshall (1999). We therefore used the calibration to constrain the divergence age between this species (Zetela 1) and its sister taxa of two Antarctic species. The two Antarctic species were also tentatively assigned to Zetela on the basis of morphological similarity to Z. kopua and genetic similarity to Zetela 1. The node was constrained to be at least 16.7 Ma (Hollis et al. 2010) (95% interval: 16.7–27.9 Ma; mean in real space: 2.65, log stdev: 1, offset: 16.5).
Depth data
Depth data were only obtained for species and genera used in this study, as the assignment of species to genera is often uncertain. Data for each species were taken from collecting localities for each specimen used in this study (Table 1), and literature records where more detailed information was available for recognized species (Ilanga biradiatula, I. discus, Herbert 1987; “Solariella” varicosa, Warén 1993; “Archiminolia” alabida, “A.” diadema, Marshall 1999; Hazuregyra watanabei, Minolia nyssonus, “Machaeroplax” delicatus, Hasegawa 2009; Clade C spp. Vilvens and Williams 2013). Literature records were not used for Z. kopua or Solariella affinis as there are different “forms” that might represent different species (Warén 1993; Marshall 1999). Neither were they used for I. laevissima as the specimens identified in Herbert (1987) represent at least two species (genetic results from this study). Instead, museum collections at the NMSA were re-examined to find new depth data for I. laevissima s.s and Ilanga 18.
Most information from this study was based on dredge and trawl data and as such there is likely to be some degree of error, as depth data were not based on a point source. This effect was minimized by classifying depth range into one of three groups. Depth ranges were classified as “shallow” if species could be found in water <200 m (on the continental shelf); “deep” if species could be found in water >1050 m (bathyal zone); and “intermediate” if species were collected only in 200–1050 m (on the continental slope). Field observations have shown that “typical” deep-sea fauna (e.g., elasipod holothurians, stalked crinoids, hexactinellid sponges) can occur in the tropics as shallow as 150–180 m (Bouchet et al. 2008) justifying our choice of 200 m as a cut-off for shallow water taxa.
Depth ranges were plotted using Statistica (v.8; StatSoft Inc. 2008). The chronogram was used for ancestral character state reconstructions of depth using likelihood reconstruction methods and the Mk1 model in Mesquite (v. 2.75; Maddison and Maddison 2006, 2011). Only two states (shallow and intermediate + deep) were used in this analysis as only two specimens in the chronogram were collected from the bathyal zone.
Results
Species delimitation
A total of 71 evolutionary significant units (ESUs) were recognized as a result of GMYC analyses, with 65 entities being recognized in the GMYC_COI tree and 70 in the GMYC_mt-gene tree (Figures S1 and S2). Taxon sets differed between the two analyses, but where they overlapped, the results were completely congruent, except that two species in the GMYC_COI tree (Zetela 3 and Solariella chodon) were each recognized as two ESUs in the mt-gene tree. Using the equivalent of a 95% confidence interval, the total number of entities identified ranged from 62 to 70 in the GMYC_COI tree and 69 to 71 in the GMYC_mt-gene tree based on model substitutions at two log-likelihood units from the maximum (C.I.; Monaghan et al. 2009). If the lower C.I. is used to define species in the GMYC_COI tree three pairs of ESUs are combined (Ilanga 5 with Ilanga 16; Ilanga 4 with Ilanga 17; and Clade A sp. 5 with Clade A sp. 6). In the mt-gene tree using the lower C.I. limit, Zetela 3 and Solariella chodon are each recognized as single species, as in the GMYC_COI tree. As previous studies have shown that the number of species is probably overestimated in GMYC analyses of low-dispersal groups (Williams et al. 2011), we conservatively treat Zetela 3 and Solariella chodon each as a single species.
Eleven specimens were not included in either GMYC analysis because of missing data. These were each recognized as distinct species based on morphological differences and large genetic differences for the genes for which sequence was available (Archiminolia 3; Bathymophila alabida, Bathymophila 12, 14 and 17; Clade B iridescens; Ilanga 18 and 20; Solariella 7; “Solariella” varicosa; Zetela kopua).
Although slowly evolving, the nuclear 28S rRNA gene is sometimes useful for separating species and we found distinct genotypes for most species. The following pairs or groups shared an identical genotype: Ilanga 4, 17 and 19; Ilanga 5 and 16; Ilanga 11 and I. laevissima; Ilanga 1, I. biradiatula and I. cf. norfolkensis; Ilanga 3 and 15; Clade C sp. 5 and Clade C sp. 8; Spectamen 4 and S. mutabilis; and Minolia nyssonus, M. punctata and M. sp.
Overall, we recognized 82 species after GMYC analyses (Table 1) and used these species definitions in the *BEAST analysis. Examination of specimens used in genetic analyses confirmed that most putative species can be distinguished from their sister species morphologically by shell characters. Examination of the chronogram suggests that divergence times between two species pairs are very small (Ilanga 4 and 17, 1.22 Myr, HPD: 0.28–2.17; and Ilanga 5 and 16; 1.13 Myr, HPD: 0.2–2.03). These same pairs were combined into two single species in the COI tree, when using the lower confidence interval. Further work is needed to test their specific status. A third pair (Clade A sp. 5 and Clade A sp. 6) was not tested in this way as one of the putative species was not included in the dated analyses due to missing data. The status of these two species also needs further testing.
Phylogenetic analyses
We obtained well-resolved individual and combined gene trees using MrBayes (Figs. 1–3). In all analyses, average standard deviation of split frequencies approached zero, all parameter average PSRF values were ≤1.001 and minimum ESS values in combined runs exceeded 200 for all parameters. Visual examination of traces showed that all parameters converged between independent runs for each dataset.
Figure 1.

Single gene trees based on Bayesian inference using MrBayes for Solariellidae using mitochondrial genes (16S, cytochrome oxidase subunit I [COI]), with outgroups removed for clarity. Support values are posterior probabilities (PP); branches with PP < 50% were collapsed, PP not shown for intraspecific relationships. See Table 1 for sample details. Monophyletic clades discussed in the text are indicated with a gray shaded box, non-monophyletic groups with a gray outline box.
Figure 2.

Single gene trees based on Bayesian inference using MrBayes for Solariellidae using the 12S rRNA mitochondrial gene and the 28S rRNA nuclear gene with outgroups removed for clarity. Support values are posterior probabilities (PP); branches with PP < 50% were collapsed, PP not shown for intraspecific relationships. See Table 1 for sample details. Monophyletic clades discussed in the text are indicated with a gray shaded box, non-monophyletic groups with a gray outline box.
Ten clades corresponding to genera were recognized in this study. Only three species were not assigned to clades (Suavotrochus sp., “Machaeroplax” delicatus and “Solariella” varicosa). Three genera (Ilanga, Minolia, Spectamen) and one generic-level clade (Clade B) were recovered as monophyletic in all trees (Figs. 4). Clades A and C were monophyletic in at least two gene trees and the combined tree (Figs. 4). Archiminolia, Bathymophila and Solariella were not monophyletic as traditionally defined in any tree, but as re-defined in this study Solariella was monophyletic in all trees, Bathymophila in two gene trees and the combined gene tree and Archiminolia in all trees except 28S. Three species that we tentatively assigned to Zetela were monophyletic in the combined gene tree, but only Z. kopua and Zetela 1 formed a clade in the 28S tree (only sequence for 28S was available for Zetela kopua). The monotypic Hazuregyra was sister to Minolia in all analyses.
Figure 4.

(a) Evolution of global climate over the last 65 Myr. The graph shows a stacked deep-sea benthic foraminiferal oxygen-isotope curve. The δ18O temperature scale, on the right axis, applies only to the time preceding the onset of large-scale glaciation on Antarctica (about 35 million years ago). Modified from Figure 2 in Zachos et al. 2008. (b) Chronogram for Solariellidae, with branch lengths proportional to time (scale below in millions of years) based on the three-calibration *BEAST tree. Support values are posterior probabilities (PP, above branches); only values ≥50% are shown. Horizontal, light purple bars on nodes correspond to 95% highest posterior density (HPD) interval for node heights (ages). The 95% HPD is the shortest interval that contains 95% of the sampled values. Clades marked X and Y are discussed in the text. Wide vertical purple bars highlight time periods of interest. Clades with substantially increased rates of diversification are indicated with thickened, vertical black lines. Geographic and depth distributions of species are indicated by a colored box next to the species name (see Key for details). Nodes used to calibrate chronogram are marked with a black square: 1) ingroup calibration; 2) Solariella calibration; and 3) Zetela calibration.
Outgroup taxa formed well-supported clusters in individual and combined gene trees consistent with families and clades identified in previous studies (Williams 2012) (Figure S3).
Chronogram
Acceptable ESS values for the *BEAST analysis with three calibrations were obtained by combining 353,662 trees sampled from eight independent runs (ESS >150 for all parameters) and with one calibration by combining 215,331 trees sampled from five independent runs (ESS >200). All ESS values were greater than 200 for both BEAST runs.
The *BEAST tree with three fossil calibrations is shown in Figure 4. Other chronograms are not shown, as the four trees were almost identical in topology, with no well-supported branches (PP > 90%) in conflict. Ages were similar, but consistently younger in analyses with three calibrations rather than with one, both for BEAST and *BEAST (see Table 2 for summary of ages). Moreover, divergence time estimates in *BEAST analyses were generally older than BEAST estimates except for younger clades (particularly nodes <5 Myr). Support values were similar, but slightly lower in *BEAST analyses. Ages used in the Discussion are based on the *BEAST analysis using three calibrations. Relationships among some clades differed slightly between the MrBayes and *BEAST trees, but most of these differences were not well supported.
Table 2.
Estimated crown ages (and 95% highest posterior density interval) in millions of years for solariellid clades calculated in separate *BEAST or BEAST analyses
| Genus/Clade | *BEAST – 3 calibrations | *BEAST – 1 calibration | BEAST – 3 calibrations | BEAST – 1 calibration |
|---|---|---|---|---|
| Solariellidae | *73.08 Myr (71.09–76.6) | *73.77 Myr (71.07–78.91) | *72.61 Myr (71.05–75.01) | *72.83 Myr (71.75–75.73) |
| Archiminolia | 4.82 Myr (2.28–7.49) | 5.38 Myr (2.7–8.37) | 4.29 Myr (2.44–6.47) | 4.66 Myr (2.62–7.12) |
| Bathymophila | 20.46 Myr (14.19–26.86) | 22.94 Myr (16.11–29.54) | 18.91 Myr (14.16–24.28) | 20.5 Myr (14.37–27.13) |
| Clade C | 23.16 Myr (15.43–30.55) | 25.53 Myr (17.92–34.06) | 20.02 Myr (14.01–26.5) | 21.5 Myr (14.36–29.27) |
| Ilanga | 40.14 Myr (30.9–49.89) | 45.53 Myr (35.25–51.84) | 34.3 Myr (26.54–41.82) | 36.59 Myr (27.71–46.52) |
| Minolia | 6.89 Myr (3.97–9.83) | 7.49 Myr (4.25–10.79) | 6.12 Myr (3.93–8.51) | 6.57 Myr (4.16–9.38) |
| Clade A | 6.79 Myr (4–9.68) | 7.72 Myr (4.6–11.01) | 6.39 Myr (4.18–8.8) | 6.95 Myr (4.22–9.74) |
| Clade B | 13.28 Myr (7.67–18.63) | 14.58 Myr (8.38–20.84) | 11.67 Myr (7.27–16.21) | 12.57 Myr (7.39–17.94) |
| Solariella | *30.28 Myr (23.22–37.32) | 36.3 Myr (27.06–45.48) | *26.8 Myr (23.12–41.7) | 31.06 Myr (22.99–38.88) |
| Spectamen | 23.67 Myr (15.73–31.37) | 26.9 Myr (17.87–35.85) | 20.19 Myr (14.26–26.27) | 22.15 Myr (14.71–29.48) |
| Zetela | *18.31 Myr (16.55–21.17) | 20.3 Myr (13.71–26.91) | *17.89 Myr (16.58–20.2) | 18.93 Myr (12.76–25.49) |
| Antarctic Clade | 5.7 Myr (2.94–8.64) | 6.34 Myr (3.24–9.72) | 6.69 Myr (4.06–9.66) | 7.09 Myr (3.98–10.47) |
| Clade X | 34.33 Myr (26.15–42.84) | 37.55 Myr (28.46–47.42) | 31.26 Myr (24.5–38.85) | 33.68 Myr (24.4–44.17) |
| Clade Y | 53.27 Myr (43.03 –63.99) | 58.26 Myr (47.33–68.78) | 47.87 Myr (39.02 –57) | 52.72 Myr (41.65–63.21) |
| # unique clades | 25,892 | 22,574 | 142 | 141 |
| Highest log clade credibility | −5.98 | −5.60 | −5.37 | −5.00 |
Nodes used in calibrations marked with an asterisk.
Diversification
The LTT plot for the solariellid phylogeny was a straightline (not shown), which is the expectation under a constant birth–death model, where the slope equals speciation rate minus extinction rate (Harvey et al., 1994; Pybus and Harvey 2000). The MCCR test confirmed that the LTT plot did not differ significantly from a constant net rate of diversification over time (species sampled = 68; experimental γ = 0.206; number of replicates = 500) in a number of tests with an estimated total number of species ranging between 100 and 6000, thus showing that our result is robust even with the likelihood of missing taxa (range γ0.05 = −2.42 to −9.02; P range = 0.87 to 1).The survivorship analysis also suggested there was no significant difference between Model A (constant diversification) and Model B (gradual decrease in diversification over time; β = 1.09) (P = 0.35) or Model A and C (P = 0.58).
However, although the overall rate was constant, the relative cladogenesis test shows that one major clade, designated Clade X in the chronogram (Fig. 4) demonstrated a substantial, although not statistically significant increase in cladogenesis (P = 0.08). The most speciose subclade within this clade corresponds to the genus Bathymophila. Speciation rates were higher in Bathymophila than in the shallower-water genus Ilanga over a range of different estimates of total taxa assuming high levels of extinction and almost double when extinction was zero (Table 3).
Table 3.
Net diversification rate for two solariellid clades based on equations in Magallon and Sanderson (2000), with no extinction (ε = 0) or high extinction (ε = 0.9). N = estimated total number of species in clade, missing = number and percentage not included in this study
| Age (Myr) | ε = 0 | ε = 0.9 | N (missing) | |
|---|---|---|---|---|
| Bathymophila | 20.46 | 0.121 | 0.056 | 24 (7, 29%) |
| 0.138 | 0.069 | 34 (17, 50%) | ||
| 0.158 | 0.085 | 51 (34, 67%) | ||
| Ilanga | 40.14 | 0.076 | 0.039 | 42 (12, 30%) |
| 0.075 | 0.038 | 50 (20, 50%) | ||
| 0.085 | 0.047 | 60 (40, 67%) |
Depth data
Species were most common on the continental slope (200–1000 m), although the scarcity of both deeper-water species (>1050 m) and shallow species between 50 and 200 m may reflect sampling effort to some extent (Table 1). Combined sampling effort for all stations for the MNHN expeditions listed in this study was greatest in the 200–1050 m range with approximately 78% of stations occurring entirely within these limits (including stations where solariellids were not found). Approximately 17% of stations in these expeditions were all or partly in the range 50–200 m and 5% of stations were partly or entirely in depths >1050 m. Sampling in MNHN expeditions was intense in intertidal and subtidal waters, but in this study, only one solariellid species was found at less than 100 m at a tropical locality (Spectamen philippensis), suggesting that solariellids are rare in <50 m in warm, tropical waters, moving into very shallow water only in cooler water (e.g., Japan, Norway, South Africa). Even so, several genera were commonly collected from water defined as shallow for the purposes of this study (<200 m; Ilanga, Spectamen, Solariella and Minolia) (Table 1, Fig. 5). Only three species included in this study were collected at sites >1050 m (Bathymophila 5, “Machaeroplax” delicatus and Zetela kopua) (Table 1, Fig. 5), and few solariellids have been collected alive deeper than 2500 m.
Figure 5.
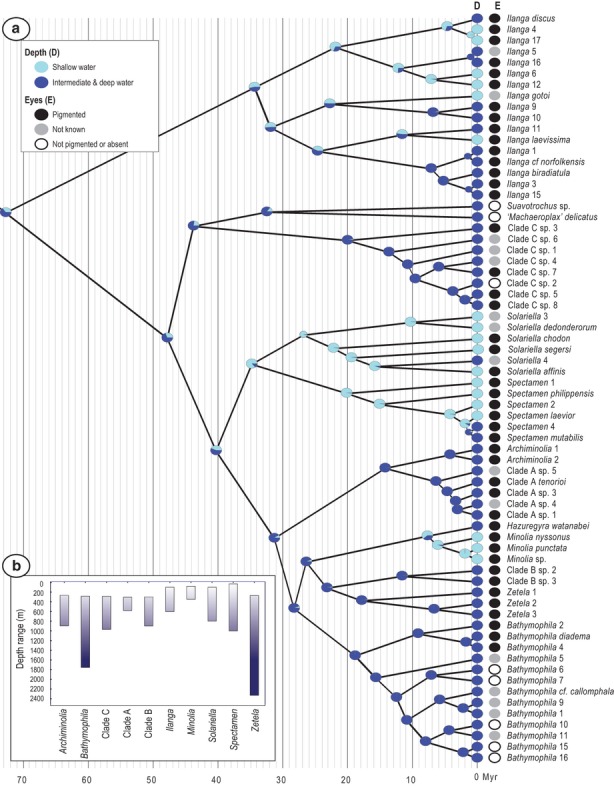
(a) Ancestral state reconstruction of depth distribution in Solariellidae. Ancestral states calculated using the Mk1 model in Mesquite. Pie charts show proportion of likelihood supporting either deep or shallow water habitat. Tree topology is based on the three-calibration *BEAST tree. The presence or absence of eyes is represented graphically next to taxon name. Note that only deep-water taxa are sightless. Light penetration in the ocean varies with latitude and distance from shore. The euphotic zone, where there is sufficient light for photosynthesis to occur, varies in depth, but may extend to around 200 m in the open ocean. (b) Depth ranges for genera and clades discussed in this study, including only species used in this study.
Discussion
Systematics
The family Solariellidae is in need of taxonomic revision, with species identification, assignment of species to genera and relationships among genera often uncertain despite several regional monographs that have advanced our understanding of the group (Quinn 1979, 1991; Herbert 1987; Marshall 1999; Vilvens 2009). For instance, of the total 82 species recognized in this study, probably more than two-thirds are either undescribed or have only been described in the last few years. The large number of new species in combination with their patchy distributions suggests that solariellids are extremely diverse and new species are likely to be found as sampling continues. It is important that the systematics of the group is resolved, and this will be addressed separately.
In this study, we note only that ten clades of generic rank were recognized. Contrary to expectation, shell characters could be used to distinguish between most of these clades. This is particularly useful, as many species have been described entirely on the basis of shell characters. Five clades correspond to known genera Ilanga, Spectamen, Minolia, Zetela and Clade C (currently being described by Vilvens and Williams 2013). Type species were included for all these genera except Zetela. A further two clades of possibly generic rank were identified (Clades A and B).
Clades were also identified that include species assigned to Solariella, Bathymophila and Archiminolia, although no type species were included. If the clades found in this study represent these genera, several species need generic reassignment. Regrettably, the type species of the nominotypical genus Solariella, S. maculata, cannot be included as it is a fossil species from the Pliocene. Several North Sea species, especially S. amabilis and S. affinis, are so similar to S. maculata, that they may confidently be considered to represent the genus and further members may yet be found off the coast of West Africa (Herbert 1987; Warén 1993; Marshall 1999). Our study did not include any West African taxa, but did include S. affinis. We tentatively assume that the clade including this species corresponds to Solariella sensu stricto.
The monotypic Hazuregyra was found to be sister and genetically similar to Minolia, and should perhaps be considered a synonym thereof. Three species Suavotrochus sp., “Machaeroplax” delicatus and “Solariella” varicosa do not cluster with known genera, but form a poorly supported clade together, sister to Clade C (Fig. 4). Their generic status needs further assessment, preferably including species from the Atlantic. In particular, Microgaza (not represented in our dataset) based on a species from the Gulf of Mexico, is conchologically similar to Ilanga and the name has been used for Indo-Pacific species, but it differs in radular morphology (Herbert 1987) and its affinities with Ilanga remain to be established.
Origin of deep-sea solariellids
It is not possible to give an accurate estimate of the proportion of total species sampled in this study (we show that there are likely many undescribed species), but species within a genus often share similar biogeographic and depth ranges and we include representatives of all but two currently accepted solariellid genera: the IWP genus Minolops (which may be synonymous with Spectamen; Marshall 1999) and one predominantly shallow-water Atlantic genus, Microgaza. Therefore, although any interpretation of the solariellid phylogeny we present here must be speculative, we can be fairly confident of most patterns observed in this study.
The oldest confirmed fossil solariellids are from shallow tropical settings of Late Cretaceous age (Hickman and McLean 1990; Kiel and Bandel 2001; Kiel et al. 2002). The first records from continental slope palaeodepths are of late Eocene and Oligocene age and were found in cool-temperate regions of both hemispheres (Maxwell 1992; Kiel 2010). The molecular data are not inconsistent with the fossil data, although there is no strong support for either shallow or deep-sea origin. In our tree, the genus Ilanga is sister to all other taxa sampled. Although this genus is sister to all other solariellids, ancestral state reconstruction suggests that a deep sea habitat is slightly more plausible for the common ancestor of solariellids (Fig. 5); however, our sampling of shallow water taxa is limited. We have included only 20 of the 32 Ilanga species recognized to date (other species listed by Herbert 1987). Of the total number of species, more than half (18) can be found in water <200 m and ten (not included in this study) have only been collected live in <100 m, suggesting that Ilanga is a tropical and temperate clade found predominantly in shallow or upper slope waters (50–300 m). The addition of these shallow-water species may change the result, as might the addition of the missing shallow-water genus Microgaza or any extinct genera. For instance, the oldest fossil identified is the shallow-water species “Solariella” montsecana from the Campanian of Torallola, Spain (Kiel and Bandel 2001). This species, although similar to species in Clade A, probably represents an extinct genus. Shallow, tropical origins of the group as suggested by the fossil data are consistent with patterns showing the tropics and areas with carbonate substrates have acted as cradles of diversity (Jablonski et al. 2006; Alfaro et al. 2007; Kiessling et al. 2010).
Conversely, it has also been suggested that solariellids from Antarctica might be more primitive than previously thought and an Antarctic origin was postulated for the family suggesting the few extant species from Antarctica represent relictual ancestors of lineages that acted as a source of diversity for deep-water communities elsewhere (Linse 2002). This study includes two out of eight recognized Antarctic and sub-Antarctic species, and these form a derived clade within the solariellid tree. The two Antarctic species diverged from their Indian Ocean sister species about 18 Mya (16.55–21.17) during a period of warmer climate (Zachos et al. 2001). This is consistent with the hypothesis that Antarctica acts as a sink for lineages immigrating during warmer periods (Clarke and Crame, 1992; Barnes et al. 2006; Göbbeler and Klussmann-Kolb 2009). The other six Antarctic species not included in this study have been assigned to Solariella (Linse 2002). If this assignment is correct, then an Antarctic origin is still unlikely for the family (although possible for the genus).
It has been suggested that deep-sea molluscs have arisen from multiple origins, but at the family and genus levels, the first members of the abyssal fauna to invade the deep sea probably did so in the relatively recent geologic past (Clarke 1962). This suggestion fits with our chronogram, which shows that invasions to the bathyal zone occurred only rarely and since the beginning of the Oligocene (given limited sampling). Invasions into intermediate depth water on the continental slope appear to have occurred more frequently.
The sister clade to Ilanga diverged around 53 Mya (HPD: 43.03–63.99; Clade Y, Fig. 4), with lineages in both shallow and deep-water (Figs. 5). Since then, there have been unambiguous invasions from shallow into deeper water (in Solariella and Spectamen) (Figs. 5). Other invasions are more difficult to interpret. For instance, most likely there was a single transition from deep to shallow water in Minolia about 8 or 9 Mya, but there may instead have been two transitions including one from deep to shallow water in the ancestral lineage (perhaps as much as 26–29 Mya) followed by a recent reversal to deep water again in Hazuregyra.
Pressure from predators or competitors is unlikely to have played an important role in the invasion of some lineages into deeper water, as specimens with repairs to their shells are frequent, suggesting that mechanical damage, possibly as a result of predation is also common in the deep sea. Equally, bathyal anoxic events probably played a limited role in preventing lineages from diversifying in deep-water in this group, as the radiation of extant taxa is Cenozoic and postdates the most widespread and frequent of these events (Jacobs and Lindberg 1998). Nearly all solariellid specimens from Antarctica collected in this study had highly corroded shells, although this was not evident for species collected at other sites. Arctic species can also show signs of corrosion (A. Waren, pers. obs.). The relevance of these observations, particularly in light of concerns about modern-day ocean acidification, cannot be determined without further work; however, one explanation may be related to the fact that carbon dioxide concentration increases at the poles as a result of decay of organic matter (e.g., Anderson et al. 2010) and over winter as there is virtually no photosynthesis.
The effect of Cenozoic global climate change on diversification
Climate change is known to be an important factor driving evolution (e.g., Lipps and Mitchell 1976; Berger 2007; Jaramillo et al. 2010). For instance, Vrijenhoek (2013) showed that the crown ages of dominant vent and seep taxa are younger than the PETM, and suggests that they may have radiated after the extinction of earlier lineages. Conversely, the crown age of Solariellidae predates the PETM and although the major solariellid clade sister to Ilanga diversified approximately 53 Mya (HPD: 43.03–63.99 Myr; Clade Y, Fig. 4), soon after the PETM (∼55.5 Mya), there is no other molecular evidence in this group for dramatic evolutionary response to this abrupt climate change. This may be because solariellids are most common on the continental slope in depths shallower than 2000 m, and as such were not probably affected by changes to the CCD or deep-sea anoxic events. It is, however, impossible to rule out that some bathyal lineages may have migrated permanently into shallower (continental slope) water or have gone extinct; testing these hypotheses would require detailed fossil evidence.
Another extremely abrupt transition in climate occurred 33.5 to 34 Mya spanning the Eocene–Oligocene boundary, when the Earth abruptly cooled and permanent continental-scale ice sheets first formed in Antarctica (Miller et al. 2009). According to our estimates, solariellid Clade X diversified within this period, approximately 34 Mya (HPD: 26.15–42.84), showing a substantial, although not statistically significant, increase in the rate of cladogenesis. Genera in Clade X are found predominantly in intermediate, slope water or bathyal depths, whereas its sister clade (Solariella + Spectamen) includes species that can be found in shallow shelf water. A greater diversification of slope rather than shelf species may have been due to a number of factors including an increase in nutrients on the continental slope, especially if food was a limiting factor for slope but not shelf habitats. Nannofossil evidence suggests that ocean productivity increased at intermediate depths (300–500 m) at the EOT in Tanzania, and in the equatorial Pacific and the Southern Ocean (Dunkley Jones et al. 2008; Lyle et al. 2008).
Increased productivity may have arisen as a result of erosion and release of nutrients from organic-rich, shallow, shelf deposits exposed during sea-level falls coincident with large-scale glaciation in Antarctica (Dunkley Jones et al. 2008). Increased ocean circulation at the EOT may also have enhanced production of Subantarctic Mode Water, which transports nutrients from Antarctica to the tropical Indian Ocean (Kiefer et al. 2006) and other tropical/subtropical regions (Dunkley Jones et al. 2008).
In further support of the idea that food may have been a limiting factor, a study on echinoids also showed that while generalist omnivores migrated into deep-water in low numbers over the last 200 Myr, specialist detritivores invaded the deep sea in large numbers between 55 and 75 Mya, probably as a result of increased organic carbon (Smith and Stockley 2005). Solariellids are also specialist deposit feeders; they use highly modified lips to sweep surface detritus into the mouth, and they have greatly shortened radula consistent with little mechanical wear and modified, bifid propodium and mesopodium to facilitate burrowing in soft sediment. Our estimates that the Recent Solariellidae radiated over the last 73 Myr are consistent with the pattern observed in echinoids.
Thus, food availability may have been a factor limiting exploitation of deep-sea habitats for some groups. Other factors, such as increased deep-basin ventilation, a decrease in deep-ocean acidity and a deepening of the CCD by more than 1000 m, which doubled the area of sea-floor subject to calcium carbonate deposition (Rea and Lyle 2005), may have opened up new ecological niches for some groups, allowing invasion of continental shelf and slope lineages into bathyal regions.
Other factors driving diversification in the deep sea
The increase in diversification in Clade X is due largely to its most speciose subclade, the genus Bathymophila. Several factors may have affected diversification in this genus. The Bathymophila clade diversified 20.46 Myr (HPD: 14.19–26.86), soon after the collision of the Australia and New Guinea plate with the southeast extremity of the Eurasian plate and the Philippines-Halmahera-New Guinea arc system ∼25 Mya (Hall 1998). This tectonic activity has been invoked as an important driver of speciation 20–25 Mya in shallow-water invertebrates (Kohn 1990; Wilson and Rosen 1998; Williams 2007; Renema et al. 2008; Williams and Duda 2008; Bellwood et al. 2012) and more recently in deep-water organisms (Cabezas et al. 2012) predominantly through the increased availability of new habitats and greater habitat complexity. Another possibility is that terrestrial run-off from the uplift of landmasses and concurrent volcanism provided additional food sources for deep-sea benthic fauna.
A different explanation might be suggested by a study that showed that speciation occurred more rapidly in deep-sea, eyeless clades of ostracods than shallow-water sighted clades (Syme and Oakley 2012). Eyes are unpigmented in several deep-slope and bathyal solariellids (e.g., “Machaeroplax” delicatus, Suavotrochus sp., Clade C sp. 2, Bathymophila 6, 7, 10, 15 and 16). In fact, all species corresponding to Marshall's (1999) concept of Bathymophila that have been examined have unpigmented eyes (this study; Marshall 1999), suggesting that it is a common condition for this clade. Like ostracods, diversification also appears to be higher in Bathymophila than other shallower, sighted clades. For example, speciation rates are up to double those in Ilanga, the most speciose clade sampled in this study with pigmented eyes and found in shallower water (Herbert 1987; Table 3). Key innovations are known to affect rates of diversification (Heard and Hauser 1995) and the loss of a character that no longer offers a selective advantage may also be viewed as an innovation (e.g., Jeffery 2005, 2009). Alternatively, the factor driving diversification in these groups may actually be the deep-water habitat, rather than the loss of eyes per se, as the two are often coupled.
Species ranges and biogeographic patterns
Solariellids have been shown to have exceptionally patchy distributions, suggesting highly specific ecological requirements (Marshall 1999). No species is known to be endemic to hot vents (Kiel 2006, 2010; Sellanes et al. 2008), although one species has been collected from cold seeps off Chile (Warén et al. 2011). Species used in this study were predominantly collected from soft sediment, were often rare, and more than half the species were found at only one station. In some cases, this probably reflects sampling effort; for example, many deeper-water species (>1050 m) were collected only once and sampling at these depths was more limited. Several species, however, were found at multiple stations within the IWP where sampling effort was concentrated. In every case, these stations with shared taxa were located within a single biogeographic zone (southwest Pacific, northwest Pacific, southwest Indian Ocean or southeast Indian Ocean), although one southeast Indian Ocean species (Clade A sp. 6) was genetically very similar to a species from northwest Pacific (Clade A. sp. 5). No species in this study are shared between southwest and northwest Pacific sites.
The division between southwest Pacific sites (including Eastern Australia, Papua New Guinea, Solomon Islands, Fiji, Vanuatu, Tuamotus, New Zealand) and northwest Pacific sites (including Japan, Taiwan and Philippines) has been observed at both the level of population structure in many highly dispersive, shallow-water species and in the distributions of some deep-sea species (e.g., Macaranas et al. 1992; McMillan and Palumbi 1995; Palumbi 1997; Williams and Benzie 1997; Planes and Fauvelot 2002; Barber et al. 2006; Imron et al. 2007; Magsino and Juinio-Meñez 2008; Lorion et al. 2010). The congruence of pattern is likely the result of the flow of equatorial currents in the Pacific. For high-dispersal, shallow-water species, surface currents act as a porous barrier by redirecting larvae and limiting direct gene flow between southern and northern Pacific sites. However, for deep-water groups with more modest dispersal potential, the eastward-flowing Equatorial Undercurrent, which flows most strongly at the thermocline (100–200 m) (Jewell 1995), is probably more important. The current is likely to have a strong influence on gene flow in solariellids as they have relatively short-lived lecithotrophic larvae (and sometimes brood larvae) (Herbert 1987; Marshall 1999). More highly dispersing species may find the Equatorial Undercurrent a porous barrier. For example, some moderately deep-water species of Bursa (a gastropod genus with teleplanic planktotrophic larvae) span the equator, occurring in both the Philippines and New Caledonia or the Philippines and the Solomon Islands (Castelin et al. 2012). A more profound barrier is likely the oxygen minimum zone below the Equatorial Undercurrent (300–400 m; Levitus 1982), which is most pronounced in the Eastern Pacific (Jewell 1995; Levin 2003). These factors combined probably serve as an effective barrier to dispersal of some deep-water taxa across the equator promoting allopatric speciation within biogeographic zones (Wilson 1999; Rogers 2000; McClain and Hardy 2010). The existence of several NW/SW Pacific species pairs supports this idea (Ilanga 1/I. cf. norfolkensis and Ilanga 3/15, Clade A sp. 1/3, Clade C sp. 4/7, Bathymophila 15/16).
Acknowledgments
We are especially grateful to P. Bouchet, who provided most the samples in this study and to B. Buge, P. Maestrati, N. Puillandre, V. Héros and P. Lozouet for co-ordinating the loans and providing information about the sampling localities. MNHN material used in this study was collected during expeditions arranged by P. Bouchet and acknowledged in Williams (2012) and the following MNHN expeditions: BERYX, BIOPAPUA, BOA 1, BORDEAU 1, CONCALIS, NORFOLK 1 and 2, SOLOMON 1 and TARASOC. We are grateful to T. Høisæter for samples from Norway, M. Ito and K. Sakuma for the specimen of Hazuregyra watanabei collected by RV Wakataka-maru (Tohoku National Fisheries Research Institute; Ninth Cruise, 2010, Leg. 3) and to chief scientist J. Hashimoto for specimens collected by T/V Nagasaki-maru. We also thank J. Llewellyn-Hughes and C. Griffin for operating automated sequencers at the NHM. The following people kindly arranged loans: J. Waterhouse and I. Loch (AM), G. Harasewych (SI), C. Whisson, S. Slack-Smith, and J. Fromont (WAM). Thanks to T. Nakano for sequences for Minolia sp., P. Kuklinski for analyses in Statistica and comments on the text, T. Ozawa for new locality data for GenBank specimens, J. Zachos for providing a copy of his figure (Fig. 4A), J. Pickering for building a systematic database, H. Taylor for photos of specimens in Figure 3 and G. Vermeij and R. Helwerda for helpful discussions. Two anonymous reviewers and M. Hart made comments that improved the manuscript. The BIOPEARL II expedition and KL are part of the BAS Polar Science for Planet Earth Programme funded by NERC.
Conflict of Interest
None declared.
Supporting Information
Additional Supporting Information may be found in the online version of this article:
Figure S1. Results of GMYC test on an ultrametric tree produced using BEAST for the cytochrome oxidase I sequences. (A) Tree with ESUs and species names marked on the right-hand side. Support for nodes are posterior probabilities, shown only for species-level relationships and PP > 50%. Branches in red indicate population structure. (B) Maximum likelihood plot showing peak congruent with threshold limit in B. (C) Lineage through time plot with red line showing the threshold level suggested by GMYC.
Figure S2. Results of GMYC test on an ultrametric tree produced using BEAST for concatenated sequences from all three mitochondrial genes. (A) Tree with ESUs and species names marked on the right-hand side. Support for nodes are posterior probabilities, shown only for species-level relationships and PP>50%. Branches in red indicate population structure. (B) Maximum likelihood plot showing peak congruent with threshold limit in B. (C) Lineage through time plot with red line showing the threshold level suggested by GMYC.
Figure S3. Combined gene tree based on Bayesian inference using MrBayes for Solariellidae using four genes (28S, 16S, 12S and COI), showing only relationship among outgroups and Solariellidae (solariellid clade is collapsed). Support values are posterior probabilities (PP); branches with PP < 50% were collapsed, PP not shown for intraspecific relationships. See Table S1 for outgroup sampling details. Families and relationships are consistent with those discussed in Williams (2012).
Table S1. Outgroup specimens used in study, ordered by family, along with details of sampling localities, registration numbers of voucher specimens, and EMBL accession numbers for sequences. MNHN, Muséum National d'Histoire Naturelle, Paris; NHMUK, Natural History Museum, London; and NSMT, National Museum of Nature and Science, Tokyo, Japan (NSMT). Photos of specimens are in Williams (2012) or available on MorphoBank online at http://www.morphobank.org/index.php/Projects/ProjectOverview/project_id/223.
Table S2. Forward (F) and reverse (R) PCR primers (also used in sequencing), and forward (FS) and reverse (RS) internal sequencing primers. Annealing temperatures and concentration of magnesium chloride (MgCl2) used in 50 μL polymerase chain reactions.
References
- Alfaro ME, Santini F, Brock CD. Do reefs drive diversification in marine teleosts? Evidence from the pufferfish and their allies (Order Tetraodontiformes) Evolution. 2007;61:2104–2126. doi: 10.1111/j.1558-5646.2007.00182.x. [DOI] [PubMed] [Google Scholar]
- Ameziane N, Roux M. Biodiversity and historical biogeography of stalked crinoids (Echinodermata) in the deep sea. Biodivers. Conserv. 1997;6:1557–1570. [Google Scholar]
- van Andel TH. Mesozoic/Cenozoic calcite compensation depth and global distribution of calcareous sediments. Earth Planet. Sci. Lett. 1975;26:187–194. [Google Scholar]
- Anderson LG, Tanhua T, Björk G, Hjalmarsson S, Jones EP, Jutterström S, et al. Arctic ocean shelf–basin interaction: an active continental shelf CO2 pump and its impact on the degree of calcium carbonate solubility. Deep Sea Res. 2010;57:869–879. [Google Scholar]
- Aronson RB, Blake DB. Global climate change and the origin of modern benthic communities in Antarctica. Am. Zool. 2001;41:27–39. [Google Scholar]
- Barber PH, Erdmann MV, Palumbi SR, Ayre D. Comparative phylogeography of three codistributed stomatopods: origin and timing of regional lineage diversification in the coral triangle. Evolution. 2006;60:1825–1839. [PubMed] [Google Scholar]
- Barnes DKA, Hodgson DA, Convey P, Allen CS, Clarke A. Incursion and excursion of Antarctic biota: past, present and future. Glob. Ecol. Biogeogr. 2006;15:121–142. [Google Scholar]
- Bellwood DR, Renema W, Rosen BR. Biodiversity hotspots, evolution and coral reef biogeography: a review. In: Gower D, Johnson KG, Rosen BR, Richardson J, Rüber L, Williams ST, editors. Biotic evolution and environmental change in Southeast Asia. London: Linnean Society; 2012. pp. 216–245. [Google Scholar]
- Berger WH. Cenozoic cooling, Antarctic nutrient pump, and the evolution of whales. Deep-Sea Res. 2007;54:2399–2421. [Google Scholar]
- Beu AG, Raine JI. Revised descriptions of New Zealand Cenozoic Mollusca from Beu and Maxwell (1990) GNS Sci. Misc. Ser. 2009;27 [Google Scholar]
- Blois JL, Hadley EA. Mammalian response to Cenozoic climate change. Annu. Rev. Earth Planet. Sci. 2009;37:181–208. [Google Scholar]
- Bohaty SM, Zachos JC, Delaney ML. Foraminiferal Mg/Ca evidence for Southern Ocean cooling across the Eocene–Oligocene transition. Earth Planet. Sci. Lett. 2012;317–318:251–261. [Google Scholar]
- Bouchet P, Héros V, Lozouet P, Maestrati P. A quarter-century of deep-sea malacological exploration in the South and West Pacific: where do we stand? How far to go? In: Héros V, Cowie RH, Bouchet P, editors. Tropical deep-sea Benthos 25. Mémoires du muséum national d'Histoire naturelle. Vol. 196. 2008. pp. 9–40. [Google Scholar]
- Brandt A, Gooday AJ, Brandão SN, Brix S, Brökeland W, Cedhagen T, et al. First insights into the biodiversity and biogeography of the Southern Ocean deep sea. Nature. 2007;447:307–311. doi: 10.1038/nature05827. [DOI] [PubMed] [Google Scholar]
- Brey T, Dahm C, Gorny M, Klages M, Stiller M, Arntz WE. Do Antarctic benthic invertebrates show an extended level of eurybathy? Antarct. Sci. 1996;8:3–6. [Google Scholar]
- Briggs JC. Marine centres of origin as evolutionary engines. J. Biogeogr. 2003;30:1–18. [Google Scholar]
- Cabezas P, Sanmartin I, Paulay G, Macpherson E, Machordom A. Deep under the sea: unraveling the evolutionary history of the deep-sea squat lobster Paramunida (Decapoda, Munididae) Evolution. 2012;66:1878–1896. doi: 10.1111/j.1558-5646.2011.01560.x. [DOI] [PubMed] [Google Scholar]
- Castelin M, Lorion J, Brisset J, Cruaud C, Maestrati P, Utge J, et al. Speciation patterns in gastropods with long-lived larvae from deep-sea seamounts. Mol. Ecol. 2012;21:4828–4853. doi: 10.1111/j.1365-294X.2012.05743.x. [DOI] [PubMed] [Google Scholar]
- Castresana J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000;17:540–552. doi: 10.1093/oxfordjournals.molbev.a026334. [DOI] [PubMed] [Google Scholar]
- Clarke AH. Annotated list and bibliography of the abyssal marine molluscs of the world. Natl. Mus. Can. Bull. 1962;181:1–114. i–vi. [Google Scholar]
- Clarke A, Crame JA. The Southern Ocean benthic fauna and climate change: a historical perspective. Phil. Trans. R. Soc. Lond. B. 1992;338:299–309. [Google Scholar]
- Clarke A, Crame JA. Evolutionary dynamics at high latitudes: speciation and extinction in polar marine faunas. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010;350:3655–3666. doi: 10.1098/rstb.2010.0270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coxall HK, Wilson PA, Pälike H, Lear CH, Backman J. Rapid stepwise onset of Antarctic glaciation and deeper calcite compensation in the Pacific Ocean. Nature. 2005;433:53–57. doi: 10.1038/nature03135. [DOI] [PubMed] [Google Scholar]
- Crame JA. Latitudinal range fluctuations in the marine realm through geological time. Trends Ecol. Evol. 1993;8:62–166. doi: 10.1016/0169-5347(93)90141-B. [DOI] [PubMed] [Google Scholar]
- Dickens GR, O'Neil JR, Rea DK, Owen RM. Dissociation of oceanic methane hydrate as a cause of the carbon isotope excursion at the end of the Paleocene. Paleoceanography. 1995;10:965–971. [Google Scholar]
- Drummond AJ, Rambaut A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007;7:214. doi: 10.1186/1471-2148-7-214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunkley Jones T, Brown PR, Pearson PN, Wade BS. Major shifts in calcareous phytoplankton assemblages through the Eocene-Oligocene transition of Tanzania and their implications for low-latitude primary production. Paleoceanography. 2008;23:PA4204. [Google Scholar]
- Gingerich PD. Environment and evolution through the Paleocene-Eocene thermal maximum. Trends Ecol. Evol. 2006;21:246–253. doi: 10.1016/j.tree.2006.03.006. [DOI] [PubMed] [Google Scholar]
- Göbbeler K, Klussmann-Kolb A. Out of Antarctica? – new insights into the phylogeny and biogeography of the Pleurobranchomorpha (Mollusca, Gastropoda) Mol. Phylogenet. Evol. 2009;55:996–1007. doi: 10.1016/j.ympev.2009.11.027. [DOI] [PubMed] [Google Scholar]
- Halfar J, Fujita RM. Danger of deep-sea mining. Science. 2007;316:987. doi: 10.1126/science.1138289. [DOI] [PubMed] [Google Scholar]
- Hall R. The plate tectonics of Cenozoic SE Asia and the distribution of land and sea. In: Hall R, Holloway JD, editors. Biogeography and geological evolution of SE Asia. Leiden: Backhuys Publishers; 1998. pp. 99–131. [Google Scholar]
- Harmon LJ, Weir J, Brock C, Glor RE, Challenger W. GEIGER: investigating evolutionary radiations. Bioinformatics. 2008;24:129–131. doi: 10.1093/bioinformatics/btm538. [DOI] [PubMed] [Google Scholar]
- Harvey PH, May RM, Nee S. Phylogenies without fossils. Evolution. 1994;48:523–529. doi: 10.1111/j.1558-5646.1994.tb01341.x. [DOI] [PubMed] [Google Scholar]
- Hasegawa K. Fujita T, editor. Upper bathyal gastropods of the Pacific coast of northern Honshu, Japan, chiefly collected by R/V Wakataka-maru. Natl. Mus. Nat. Sci. Monograph. 2009;39:225–383. Deep-sea fauna and pollutants off Pacific coast of northern Japan. [Google Scholar]
- Heard SB, Hauser DL. Key evolutionary innovations and their ecological mechanisms. Hist. Biol. 1995;10:151–173. [Google Scholar]
- Heled J, Drummond AJ. Bayesian inference of species trees from multilocus data. Mol. Biol. Evol. 2010;27:570–580. doi: 10.1093/molbev/msp274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herbert DG. Revision of the Solariellinae (Mollusca: Prosobranchia: Trochidae) in southern Africa. Ann. Natal Mus. 1987;28:283–382. [Google Scholar]
- Hickman CS, McLean JH. Systematic revision and suprageneric classification of trochacean gastropods. Nat. Hist. Mus. Los Ang. Co. Sci. Ser. 1990;35:1–169. [Google Scholar]
- Hollis CJ, Beu AG, Crampton JS, Jones AF. Calibration of the New Zealand Cretaceous-Cenozoic timescale to GTS2004. GNS Sci. Rep. 2010;43:1–20. [Google Scholar]
- Huelsenbeck JP, Ronquist F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics. 2001;17:754–755. doi: 10.1093/bioinformatics/17.8.754. [DOI] [PubMed] [Google Scholar]
- Imron BJeffrey, Hale P, Degnan BM, Degnan SM. Pleistocene isolation and recent gene flow in Haliotis asinina, an Indo-Pacific vetigastropod with limited dispersal capacity. Mol. Ecol. 2007;16:289–304. doi: 10.1111/j.1365-294X.2006.03141.x. [DOI] [PubMed] [Google Scholar]
- Jablonski D. Evolutionary innovations in the fossil record: the intersection of ecology, development, and macroevolution. J. Exp. Zool. 2005;304B:504–519. doi: 10.1002/jez.b.21075. [DOI] [PubMed] [Google Scholar]
- Jablonski D, Bottjer DJ. Environmental patterns in the origins of higher taxa: the post-Paleozoic fossil record. Science. 1991;252:1831–1833. doi: 10.1126/science.252.5014.1831. [DOI] [PubMed] [Google Scholar]
- Jablonski D, Sepkoski JJ, Bottjer DJ, Sheehan PM. Onshore-offshore patterns in the evolution of Phanerozoic shelf communities. Science. 1983;222:1123–1125. doi: 10.1126/science.222.4628.1123. [DOI] [PubMed] [Google Scholar]
- Jablonski D, Roy K, Valentine JW. Out of the tropics: evolutionary dynamics of the latitudinal diversity gradient. Science. 2006;314:102–106. doi: 10.1126/science.1130880. [DOI] [PubMed] [Google Scholar]
- Jacobs DK, Lindberg DR. Oxygen and evolutionary patterns in the sea: onshore/offshore trends and recent recruitment of deep-sea faunas. Proc. Natl. Acad. Sci. USA. 1998;95:9396–9401. doi: 10.1073/pnas.95.16.9396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaramillo CA, Ochoa D, Contreras L, Pagani M, Carvajal-Ortiz H, Pratt LM, et al. Effects of rapid global warming at the Paleocene-Eocene boundary on neotropical vegetation. Science. 2010;330:957–961. doi: 10.1126/science.1193833. [DOI] [PubMed] [Google Scholar]
- Jeffery WR. Adaptive evolution of eye degeneration in the Mexican blind cavefish. J. Hered. 2005;96:185–196. doi: 10.1093/jhered/esi028. [DOI] [PubMed] [Google Scholar]
- Jeffery WR. Regressive evolution in Astyanax cavefish annual review of genetics. Annu. Rev. Genet. 2009;3:25–47. doi: 10.1146/annurev-genet-102108-134216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jewell PW. Geologic consequences of globe-encircling equatorial currents. Geology. 1995;23:117–120. [Google Scholar]
- Kano Y. Vetigastropod phylogeny and a new concept of Seguenzioidea: independent evolution of copulatory organs in the deep-sea habitats. Zool. Scr. 2008;33:593–594. [Google Scholar]
- Kano Y, Kimura S, Kimura T, Warén A. Living Monoplacophora: morphological conservatism or recent diversification? Zool. Scr. 2012;41:471–488. [Google Scholar]
- Katoh K, Misawa K, Kuma K, Miyata T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002;30:3059–3066. doi: 10.1093/nar/gkf436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katoh K, Kuma K, Toh H, Miyata T. MAFFT version 5: improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005;33:511–518. doi: 10.1093/nar/gki198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katz ME, Cramer BS, Toggweiler JR, Esmay G, Liu C, Miller KG, et al. Impact of Antarctic Circumpolar Current development on Late Paleogene ocean structure. Science. 2011;332:1076–1079. doi: 10.1126/science.1202122. [DOI] [PubMed] [Google Scholar]
- Kelly DC, Bralower TJ, Zachos JC. Evolutionary consequences of the latest Paleocene thermal maximum for tropical planktonic foraminifera. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1998;141:139–161. [Google Scholar]
- Kennett JP, Shackleton NJ. Oxygen isotopic evidence for the development of the psychrosphere 38 Myr ago. Nature. 1976;260:513–515. [Google Scholar]
- Kiefer T, McCave I, Elderfield H. Antarctic control on tropical Indian Ocean sea surface temperatures and hydrography. Geophys. Res. Lett. 2006;33:L24612. [Google Scholar]
- Kiel S. New records and species of mollusks from Tertiary cold-seep carbonates in Washington State, USA. J. Paleontol. 2006;80:121–137. [Google Scholar]
- Kiel S. On the potential generality of depth-related ecologic structure in cold-seep communities: evidence from Cenozoic and Mesozoic examples. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010;295:245–257. [Google Scholar]
- Kiel S, Bandel K. Trochidae (Archaeogastropoda) from the Campanian of Torallola in northern Spain. Acta Geol. Pol. 2001;51:137–154. [Google Scholar]
- Kiel S, Bandel K, Perrilliat MC. New gastropods from the Maastrichtian of the Mexcala Formation in Guerrero, southern Mexico, part II: Archaeogastropoda, Neritimorpha and Heterostropha. Neues Jb. Geol. Paläontol. Abh. 2002;226:319–342. [Google Scholar]
- Kiessling W, Simpson C, Foote M. Reefs as cradles of evolution and sources of biodiversity in the Phanerozoic. Science. 2010;327:196–198. doi: 10.1126/science.1182241. [DOI] [PubMed] [Google Scholar]
- Kohn AJ. Tempo and mode of evolution in Conidae. Malacologia. 1990;32:55–67. [Google Scholar]
- Lawver LA, Gahagan LM. Evolution of Cenozoic seaways in the circum-Antarctic region. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003;198:11–37. [Google Scholar]
- Lear CH, Elderfield H, Wilson PA. Cenozoic deep-sea temperatures and global ice volumes from Mg/Cl in benthic forminiferal calcite. Science. 2000;287:269–272. doi: 10.1126/science.287.5451.269. [DOI] [PubMed] [Google Scholar]
- Lemche H. A new living deep-sea mollusc of the Cambro-Devonian class Monoplacophora. Nature. 1957;179:413–416. [Google Scholar]
- Levin LA. Oxygen minimum zone benthos: adaptation and community response to hypoxia. Oceanogr. Mar. Biol. Annu. Rev. 2003;41:1–45. [Google Scholar]
- Levitus S. Climatological atlas of the world ocean. Natl. Ocean. Atmos. Admin. Prof. Pap. 1982;13:172. [Google Scholar]
- Lindner A, Cairns SD, Cunningham CW. From offshore to onshore: multiple origins of shallow-water corals from deep-sea ancestors. PLoS ONE. 2008;3:e2429. doi: 10.1371/journal.pone.0002429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Linse K. The shelled Magellanic Mollusca: with special reference to biogeographic relations in the Southern Ocean. Theses Zool. 2002;74:1–251. [Google Scholar]
- Lipps JH, Mitchell E. Trophic model for the adaptive radiations and extinctions of pelagic marine mammals. Paleobiology. 1976;2:147–155. [Google Scholar]
- Liu Z, Pagani M, Zinniker D, Deconto R, Huber M, Brinkhuis H, et al. Global cooling during the Eocene-Oligocene climate transition. Science. 2009;323:1187–1190. doi: 10.1126/science.1166368. [DOI] [PubMed] [Google Scholar]
- Lorion J, Buge B, Cruaud C, Samadi S. New insights into diversity and evolution of deep-sea Mytilidae (Mollusca: Bivalvia) Mol. Phylogenet. Evol. 2010;57:71–83. doi: 10.1016/j.ympev.2010.05.027. [DOI] [PubMed] [Google Scholar]
- Lyle M, Barron J, Bralower TJ, Huber M, Lyle AO, Ravelo AC, et al. Pacific Ocean and Cenozoic evolution of climate. Rev. Geophys. 2008;46:RG2002. [Google Scholar]
- Macaranas JM, Ablan CA, Pante MJR, Benzie JAH, Williams ST. Genetic structure of giant clam (Tridacna derasa) populations from reefs in the Indo-Pacific. Mar. Biol. 1992;113:231–238. [Google Scholar]
- Maddison DR, Maddison WP. MacClade. Version 4.06 OSX. Sunderland, Massachusetts: Sinauer Associates; 2003. [Google Scholar]
- Maddison WP, Maddison DR. 2006. StochChar: a package of Mesquite modules for stochastic models of character evolution. Version 1.1.
- Maddison WP, Maddison DR. 2011. Mesquite: a modular system for evolutionary analysis. Version 2.75. Available at http://mesquiteproject.org.
- Magallon S, Sanderson MJ. Absolute diversification rates in angiosperm clades. Evolution. 2000;55:1762–1780. doi: 10.1111/j.0014-3820.2001.tb00826.x. [DOI] [PubMed] [Google Scholar]
- Magsino R, Juinio-Meñez M. The influence of contrasting life history traits and oceanic processes on genetic structuring of rabbitfish populations Siganus argeneus and Siganus fuscescens along the eastern Philippine coasts. Mar. Biol. 2008;154:519–532. [Google Scholar]
- Marshall BA. A revision of the recent Solariellinae (Gastropoda: Trochoidea) of the New Zealand region. Nautilus. 1999;113:4–42. [Google Scholar]
- Marx FG, Uhen MD. Climate, critters, and cetaceans: Cenozoic drivers of the evolution of modern whales. Science. 2010;327:993–996. doi: 10.1126/science.1185581. [DOI] [PubMed] [Google Scholar]
- Maxwell PA. Eocene mollusca from the vicinity of McCulloch's Bridge, Waihao River, South Canterbury, New Zealand: Paleoecology and systematics. New Zealand Geol. Surv. Paleontol. Bull. 1992;65:1–280. [Google Scholar]
- McClain CR, Hardy SM. The dynamics of biogeographic ranges in the deep-sea. Proc. Biol. Sci. 2010;277:3533–3546. doi: 10.1098/rspb.2010.1057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCormack JE, Heled J, Delaney KS, Peterson AT, Knowles LL. Calibrating divergence times on species trees versus gene trees: implications for speciation history of Aphelocoma jays. Evolution. 2011;65:184–202. doi: 10.1111/j.1558-5646.2010.01097.x. [DOI] [PubMed] [Google Scholar]
- McMillan WO, Palumbi SR. Concordant evolutionary patterns among Indo-West Pacific butterflyfishes. Proc. Biol. Sci. 1995;260:229–236. doi: 10.1098/rspb.1995.0085. [DOI] [PubMed] [Google Scholar]
- Miller KG, Wright JD, Katz ME, Wade BS, Browning JV, Cramer BS. Climate threshold at the Eocene–Oligocene transition: Antarctic ice sheet influence on ocean circulation. In: Koeberl C, Montanari A, et al., editors. The Late Eocene earth-hothouse, icehouse, and impacts: geological society of America special paper 452. 2009. pp. 1–10. Available at http://specialpapers.gsapubs.org/content/452. [Google Scholar]
- Monaghan MT, Wild R, Elliot M, Fujisawa T, Balke M, Inward DJG, et al. Accelerated species inventory on Madagascar using coalescent-based models of species delimitation. Syst. Biol. 2009;58:298–311. doi: 10.1093/sysbio/syp027. [DOI] [PubMed] [Google Scholar]
- Palumbi SR. Molecular biogeography of the Pacific. Coral Reefs. 1997;16:S47–S52. [Google Scholar]
- Paradis E. Assessing temporal variations in diversification rates from phylogenies: estimation and hypothesis testing. Proc. R. Soc. Lond. B. 1997;264:1141–1147. [Google Scholar]
- Pearson PN, McMillan IK, Wade BS, Dunkley Jones T, Coxall HK, Bown PR, et al. Extinction and environmental change across the Eocene-Oligocene boundary in Tanzania. Geology. 2008;36:179–182. [Google Scholar]
- Planes S, Fauvelot C. Isolation by distance and vicariance drive genetic structure of a coral reef fish in the Pacific Ocean. Evolution. 2002;56:378–399. doi: 10.1111/j.0014-3820.2002.tb01348.x. [DOI] [PubMed] [Google Scholar]
- Purvis A, Nee S, Harvey PH. Macroevolutionary inferences from primate phylogeny. Proc. Biol. Sci. 1995;260:329–333. doi: 10.1098/rspb.1995.0100. [DOI] [PubMed] [Google Scholar]
- Pybus OG, Harvey PH. Testing macro-evolutionary models using incomplete molecular phylogenies. Proc. R. Soc. Lond. B. 2000;267:2267–2272. doi: 10.1098/rspb.2000.1278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quinn JF. Biological results of the university of Miami deep-sea expeditions. 130. The systematics and zoogeography of the gastropod family Trochidae collected in the straits of florida and its approaches. Malacologia. 1979;19:1–62. [Google Scholar]
- Quinn JF. Lamellitrochus, a new genus of Solariellinae (Gastropoda: Trochidae) with descriptions of six new species from the Western Atlantic Ocean. Nautilus. 1991;105:81–91. [Google Scholar]
- Rabosky DL. LASER: a maximum likelihood toolkit for detecting temporal shifts in diversification rates from molecular phylogenies. Evol. Bioinform. 2006;2:247–250. [PMC free article] [PubMed] [Google Scholar]
- Ramirez-Llodra E, Brandt A, Danovaro R, Escobar B, De Mol E, German CR, et al. Deep, diverse and definitely different: unique attributes of the world's largest ecosystem. Biogeosciences. 2010;7:2851–2899. [Google Scholar]
- Raupach MJ, Mayer C, Malyutina M, Waegle J-W. Multiple origins of deep-sea Asellota (Crustacea: Isopoda) from shallow waters revealed by molecular data. Proc. Biol. Sci. 2012;276:799–808. doi: 10.1098/rspb.2008.1063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rea DK, Lyle MW. Paleogene calcite compensation depth in the eastern subtropical Pacific; answers and questions. Paleoceanography. 2005;20:PA1012. [Google Scholar]
- Renema W, Bellwood DR, Braga JC, Bromfield K, Hall R, Johnson KG, et al. Hopping hotspots: global shifts in marine biodiversity. Science. 2008;321:654–657. doi: 10.1126/science.1155674. [DOI] [PubMed] [Google Scholar]
- Rogers AD. The role of the oceanic oxygen minima in generating biodiversity in the deep sea. Deep-Sea Res. 2000;47:119–148. [Google Scholar]
- Ruhl HA, Smith KL., Jr Shifts in deep-sea community structure linked to climate and food supply. Science. 2004;305:513–515. doi: 10.1126/science.1099759. [DOI] [PubMed] [Google Scholar]
- Sellanes J, Quiroga E, Neira C. Megafauna community structure and trophic relationships at the recently discovered Concepción methane seep area, Chile, ∼36°S. ICES J. Mar. Sci. 2008;65:1102–1111. [Google Scholar]
- Smith AB, Stockley B. The geological history of deep-sea colonization by echinoids: roles of surface productivity and deep-water ventilation. Proc. Biol. Sci. 2005;272:865–869. doi: 10.1098/rspb.2004.2996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- StatSoft Inc. 2008. STATISTICA (data analysis software system). Ver. 8.0Available at http://www.statsoft.com. Accessed July 2012.
- Strugnell JM, Rogers AD, Prodöhl PA, Collins MA, Allcock AL. The thermohaline expressway: the Southern Ocean as a centre of origin for deep-sea octopuses. Cladistics. 2008;24:853–860. doi: 10.1111/j.1096-0031.2008.00234.x. [DOI] [PubMed] [Google Scholar]
- Syme AE, Oakley TD. Dispersal between shallow and abyssal seas and evolutionary loss and regain of compound eyes in cylindroleberidid ostracods: conflicting conclusions from different comparative methods. Syst. Biol. 2012;61:314–336. doi: 10.1093/sysbio/syr085. [DOI] [PubMed] [Google Scholar]
- Tjalsma RC, Lohmann GP. Paleocene-eocene bathyal and abyssal benthic foraminifera from the Atlantic Ocean. New York: The American Museum of Natural History; 1983. pp. 1–90. Micropaleontology, Spec. Publ., 4:i-iii. [Google Scholar]
- Van Dover CL. Tighten the regulations on deep-sea mining. Nature. 2011;470:31–33. doi: 10.1038/470031a. [DOI] [PubMed] [Google Scholar]
- Vermeij GJ. Evolution and escalation. An ecological history of life. Princeton, NJ: Princeton University Press; 1987. [Google Scholar]
- Vilvens C. New species and new records of Solariellidae (Gastropoda: Trochoidea) from Indonesia and Taiwan. Novapex. 2009;10:69–96. [Google Scholar]
- Vilvens C, Williams ST. New genus and new species of Solariellidae (Gastropoda: Trochoidea) from New Caledonia, Fiji, Vanuatu, Solomon Islands, Philippines, Papua New Guinea and French Polynesia. Trop. Deep-Sea Benthos. 2013;27 (in press) [Google Scholar]
- Vrijenhoek RC. On the instability and evolutionary age of deep-sea chemosynthetic communities. Deep-Sea Res. II. 2013 (in press) Available at http://dx.doi.org/10.1016/j.dsr2.2012.12.004(accessed January 3, 2013) [Google Scholar]
- Warén A. New and little known Mollusca from Iceland and Scandinavia. Part 2. Sarsia. 1993;78:159–201. [Google Scholar]
- Warén A, Nakano T, Sellanes J. A new species of Iothia (Gastropoda, Lepetidae) from Chilean methane seeps, with comments on the accompanying gastropod fauna. Nautilus. 2011;125:1–14. [Google Scholar]
- Williams ST. Origins and diversification of the Indo-West Pacific fauna: evolutionary history and biogeography of turban shells (Gastropoda, Turbinidae) Biol. J. Linn. Soc. 2007;92:573–592. [Google Scholar]
- Williams ST. Advances in molecular systematics of the gastropod superfamily Trochoidea. Zool. Scr. 2012;41:571–595. [Google Scholar]
- Williams ST, Benzie JAH. Indo-West Pacific patterns of genetic differentiation in the high dispersal starfish Linckia laevigata. Mol. Ecol. 1997;6:559–573. [Google Scholar]
- Williams ST, Duda TFJ. Did tectonic activity stimulate Oligo-Miocene speciation in the Indo-West Pacific? Evolution. 2008;62:1618–1634. doi: 10.1111/j.1558-5646.2008.00399.x. [DOI] [PubMed] [Google Scholar]
- Williams ST, Ozawa T. Molecular phylogeny suggests polyphyly of both the turban shells (family Turbinidae) and the superfamily Trochoidea (Mollusca: Vetigastropoda) Mol. Phylogenet. Evol. 2006;39:33–51. doi: 10.1016/j.ympev.2005.12.017. [DOI] [PubMed] [Google Scholar]
- Williams ST, Karube S, Ozawa T. Molecular systematics of Vetigastropoda: Trochidae, Turbinidae and Trochoidea redefined. Zool. Scr. 2008;37:483–506. [Google Scholar]
- Williams ST, Donald KM, Spencer HG, Nakano T. Molecular systematics of the marine gastropod families Trochidae and Calliostomatidae (Mollusca: Superfamily Trochoidea) Mol. Phylogenet. Evol. 2010;54:783–809. doi: 10.1016/j.ympev.2009.11.008. [DOI] [PubMed] [Google Scholar]
- Williams ST, Apte D, Ozawa T, Kagilis F, Nakano T. Speciation and dispersal along continental coastlines and island arcs in the Indo-West Pacific turbinid gastropod genus Lunella. Evolution. 2011;65:1752–1771. doi: 10.1111/j.1558-5646.2011.01255.x. [DOI] [PubMed] [Google Scholar]
- Wilson GDF. Some of the deep-sea fauna is ancient. Crustaceana. 1999;72:1019–1030. [Google Scholar]
- Wilson MEJ, Rosen BR. Implications of paucity of corals in the Paleogene of SE Asia: plate tectonics or centre of origin? In: Hall R, Holloway JD, editors. Biogeography and geological evolution of SE Asia. Leiden: Backhuys; 1998. pp. 165–195. [Google Scholar]
- Wing SL, Harrington GJ, Smith FA, Bloch JI, Boyer DM, Freeman KH. Transient floral change and rapid global warming at the Paleocene-Eocene boundary. Science. 2005;310:993–996. doi: 10.1126/science.1116913. [DOI] [PubMed] [Google Scholar]
- Yasuhara M, Hunt G, Cronin TM, Okahashi H. Temporal latitudinal-gradient dynamics and tropical instability of deep-sea species diversity. Proc. Natl. Acad. Sci. USA. 2009;106:21717–21720. doi: 10.1073/pnas.0910935106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasuhara M, Hunt G, Cronin TM, Hokanishi N, Kawahata H, Tsujimoto A, et al. Climatic forcing of Quaternary deep-sea benthic communities in the North Pacific Ocean. Paleobiology. 2012;38:162–179. [Google Scholar]
- Zachos JC, Quinn TM, Salamy K. High resolution (104 yr) deep-sea foraminiferal stable isotope time series. Paleoceanography. 1996;11:251–266. [Google Scholar]
- Zachos J, Pagani M, Sloan L, Thomas E, Billups K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science. 2001;292:686–693. doi: 10.1126/science.1059412. [DOI] [PubMed] [Google Scholar]
- Zachos JC, Röhl U, Schellenberg SA, Sluijs A, Hodell DA, Kelly DC, et al. Rapid acidification of the ocean during the Paleocene-Eocene thermal maximum. Science. 2005;308:1611–1615. doi: 10.1126/science.1109004. [DOI] [PubMed] [Google Scholar]
- Zachos JC, Dickens GR, Zeebe RE. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature. 2008;451:279–283. doi: 10.1038/nature06588. [DOI] [PubMed] [Google Scholar]
- Zinsmeister WJ, Feldmann RM. Cenozoic high latitude heterochroneity of southern hemisphere marine faunas. Science. 1984;224:281–283. doi: 10.1126/science.224.4646.281. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.