

SUMMARY
Highly social animals provide alternative aging models in which vastly different lifespan patterns are flexible, and linked to social caste. Research in these species aims to reveal how environment, including social cues, can shape the transition between short-lived and extremely long-lived phenotypes with negligible senescence. Among honey bee workers, short to intermediate lifespans are typical for summer castes, while the winter caste can live up to 10 times longer. For summer castes, experimental interventions could predictably accelerate, slow or revert functional senescence. In contrast, little is known about the partic ular conditions under which periods of negligible senescence in winter castes can be disrupted or sustained. We asked how manipulation of social environment in colonies with long-lived winter bees might alter the pace of functional senescence, measured as learning performance, as well as of cellular senescence, measured as lipofuscin accumulation. We show that behavioral senescence becomes rapidly detectable when the winter state is disrupted, and changes in social task behaviors and social environment (brood) are induced. Likewise, we found that cellular senescence was induced by such social intervention. However, cellular senescence showed marked regional differences, suggesting that particular brain regions age slower than others. Finally, by preventing post-winter colonies from brood rearing, behavioral senescence became undetectable, even after transition to the usually short-lived phenotypes had occurred. We envision that social regulation of negligible functional senescence and highly dynamic accumulation of a universal symptom of cellular aging (lipofuscin) offers rewarding perspectives to target proximate mechanisms of slowed aging.
KEY WORDS: senescence, brain, learning, lipofuscin
INTRODUCTION
In most species, lifespan variation between individuals is comparably modest; however, in some species, normally short-lived individuals can develop into extremely long-lived phenotypes (Münch and Amdam, 2010; Tatar and Yin, 2001). The transition from phenotypes with rapid to phenotypes with slowed or negligible aging (Finch and Austad, 2001) is not normally explained by genetic differences. Rather, different aging patterns emerge in response to environmental changes. Many studies of flexible aging – including the present work – therefore aim to reveal how environmental cues interact with physiology to produce highly different lifespan outcomes.
Honey bees, ants and mole rats are exemplars of highly social animals with complex colony structures that are maintained through social interactions between different castes (social phenotypes). Vast lifespan differences in these animal models can be linked to caste-specific behaviors or colony demography (social environment). In the honey bee, one egg-laying female, the queen, is assisted by thousands of essentially sterile female workers (Seeley, 1995; Winston, 1980). The life histories of these workers – the subjects of this study – are plastic (Amdam, 2010; Amdam and Omholt, 2002; Keller and Jemielity, 2006; Lee, 2003; Münch and Amdam, 2010; Parker, 2010). Among them, winter or diutinus bees are the longest-lived workers, and can survive for almost a year (Fluri, 1990; Maurizio, 1954). Winter bees develop when brood load ceases and larval signals (brood pheromone) are absent from the colony (Fluri et al., 1982; Maurizio, 1950; Smedal et al., 2009).
Unlike long-lived phenotypes of some other invertebrates (Tatar et al., 2001; Tatar and Yin, 2001), winter bees are not dormant. Fueled on stored honey, winter bees produce heat to keep the core nest temperature at 26–28°C (Omholt, 1987), and can retain excessive internal protein stores through most of the winter. These stores might help increase longevity, and provide the resource for brood rearing in spring, which requires considerable amounts of protein and fat (Amdam and Omholt, 2002; Fluri et al., 1977). Consequently, worker bees born in summer take several days to mature, before tending the brood. During maturation they build up protein and lipid stores in their abdominal fat body (functionally homologous to liver and white adipose tissue) and the hypopharyngeal (HP) glands in the head (Crailsheim, 1998; Fluri et al., 1982). Once developed, these glands synthesize a proteinacious jelly that is fed to other colony members, including the brood. Nurse bees can survive for several months, but appear to eventually lose the ability to produce jelly (Crailsheim, 1991; Haydak, 1957). Long before such senescence occurs, however, most summer workers transition into the forager caste. Foragers have reduced internal stores of lipid and protein, including shrunken HP glands, and do not participate in brood rearing. These bees collect pollen and nectar outside the hive, and die within days (averaging 7–10 days) of initiating foraging tasks. Together, the different survival capacities of winter, nurse and forager bees correlate with specific social roles and food-related behaviors, which in turn are associated with the amount of nutrient resources that each caste embodies.
An established model to explain the flexible lifespan patterns of worker honey bees is that vitellogenin, a yolk precursor protein, provides a survival advantage to nurse bees and winter bees, hence the castes with intermediate to high levels of this protein. Vitellogenin can act as a behavioral, endocrine and immune modulator, and appears to have considerable antioxidant potential in both workers and queen bees (Amdam et al., 2004; Corona et al., 2007; Nelson et al., 2007; Seehuus et al., 2006b). Further studies focused on systems including metabolism, as well as insulin signaling, and identified age-related abundance changes for molecules involved in neuronal signaling and growth (Ament et al., 2008; Roberts and Elekonich, 2005; Wolschin and Amdam, 2007; Wolschin et al., 2009). In contrast, little is known about biomarkers of cellular senescence, which often can provide a more direct indication of deterioration than protein expression (Hsieh and Hsu, 2011; Seehuus et al., 2006a; Tolfsen et al., 2011).
While different survival capacities between animals in natural environments are not necessarily linked to aging (Nussey et al., 2008), a growing number of studies confirm that the temporal worker castes in honey bees differ in functional senescence – commonly defined as an age-related decline in functional status (Comfort, 1956; Grotewiel et al., 2005). Accelerated behavioral senescence within only 2 weeks of foraging was first demonstrated for olfactory learning (Behrends et al., 2007), the most well-established laboratory learning paradigm in honey bees (Bitterman et al., 1983). This rapid senescence has been corroborated by studies on tactile learning performance (Scheiner and Amdam, 2009), as well as on flight performance (Vance et al., 2009) and home finding in the field (Münch et al., 2010). The behavioral decline of foragers is paralleled by a sharp mortality increase after approximately 10 days of foraging (Dukas, 2008). Experimental restriction of daily flight periods further suggests that metabolically demanding flight activity is causally linked to a shortened lifespan (Neukirch, 1982), and to the age-related decline of learning function in foragers (Tolfsen et al., 2011). Few studies so far have explored the aging dynamics of winter bees (Behrends and Scheiner, 2010; Lindauer, 1963; Seehuus et al., 2006a), but the results suggest that organismal integrity, learning and memory are not compromised despite high chronological age.
Transforming phenotypes with different life expectancies into one another is a most intriguing aspect in experimental gerontology, and may reveal how aging is modulated. Such manipulations are readily achieved in honey bees, because transitions between worker castes are induced by changing the social demography of the colony. For example, precocious transitions to the short-lived forager stage are induced by removing foragers from colonies (Huang and Robinson, 1996). Conversely, reversal from short-lived foragers to usually longer-lived nurse bees is achieved by depleting colonies of nurse bees (Robinson et al., 1992). Finally, the longest-lived winter bee stage is induced by preventing brood rearing (Fluri et al., 1982; Maurizio, 1954). Here we aim to understand the conditions under which extreme longevity can develop or be disrupted through manipulation of environmental and social cues in colonies with long-lived winter bees. Using a classical olfactory learning paradigm we show that disruption and prolongation of the winter bee stage can accelerate and slow behavioral senescence, respectively. By comparing groups with accelerated and slowed behavioral senescence we asked how cellular senescence depends on worker phenotype and tissue identity. To this end, we quantified accumulation of lipofuscin, undegradable lipid-protein granules that are normally linked to chronological age and that are associated with functional decline as well as increased mortality (Double et al., 2008; Fonseca et al., 2005a; Hsieh and Hsu, 2011). Our results suggest that cellular senescence is not homogenous among tissues with different functions, and that lipofuscin accumulation is highly dynamic after the winter bee stage is disrupted.
MATERIALS AND METHODS
Subjects
Experiments on honey bees (Apis mellifera carnica Pollmann) were conducted during winter 2008 to 2009 at the Norwegian University of Life Sciences in Ås, Norway. Snow and average temperatures below 0°C (−3.35, −6.15, −7.0 and −0.5°C for December, January, February and March for 2008–2009) prevented bees from foraging or making orientation flights from December through March, with bees forming persistent winter clusters. Typical for A. m. carnica, brood rearing was found to be absent or negligible between October and March. This provided a rather homogeneous demographical distribution in outdoor colonies with a vast majority of winter bees born before October, being at least 6 months old by the end of March.
Accordingly, groups of high chronological age were obtained from outdoor colonies (old winter bees), and from post-winter colonies (old foragers), in which emergence of young bees was monitored (Experiments 1 and 2) or prevented (Experiment 3). A controlled emergence of young individuals from post-winter colonies provided a young, non-senesced control, which was made identifiable by paint marks.
The different worker types were obtained as follows: old winter bees were directly collected from winter clusters in outdoor hives. Old foragers from post-winter flight-room colonies (see below) were identified by a color tag they received when they were first observed to collect pollen or nectar at feeders (Experiment 2). Old foragers from post-winter outdoor colonies were identified by a color tag they received when returning from first foraging flights in natural habitats (Experiment 3). Choosing different colors that specify the marking date allowed us to determine the minimum number of days an individual spent foraging (‘foraging age’). Young nest bees, the non-senesced control, were obtained by collecting combs with capped brood from post-winter colonies. Upon emergence in an incubator, they received a color tag, were re-introduced into host colonies and were sampled after 9–12 days.
All worker groups were obtained from sister queen colonies with approximately 5000–7000 individuals each. To control for effects of hive origin and day-to-day variation, worker phenotypes were represented by individuals from at least two colony sources (replicate of hive), and comparisons included specimens that were collected and tested on several days (replicate of collection day).
Experimental design
Experiment 1
For a longitudinal comparison of winter bees, specimens were collected and tested for behavioral performance in early, mid and late winter, i.e. October, January and March, respectively. To evaluate whether learning performance of old winter bees is compromised as compared with chronologically young individuals, the young control was included during initial testing in early winter. These control bees were introduced, allowed to mature and were collected from the same colony replicates from which winter bees were collected. Because learning performance may vary due to different motivational state, we controlled for similar gustatory responsiveness during all measurements in winter (see Behavioral tests, below).
Experiment 2
A cross-sectional design was chosen to simultaneously test three different worker types in mid winter. Old winter bees were collected from outdoor hives, and were contrasted with old forager bees as well as the young control, both collected from indoor post-winter colonies. To induce foraging activity, increased egg-laying and brood care activity in the post-winter colonies, a flight room was designed following general technical principles described elsewhere (Pernal and Currie, 2001). In brief, two compartments with dimensions of 4.6×2.0×2.0 m (length × width × height) each hosted one colony replicate. The day:night cycle was set to 12 h:12 h with 25 and 17°C as the maximum and minimum temperatures, respectively. Eighteen banks with light fixtures, each containing one daylight and one UV source, were installed (Philips TL-D ActiViva Natural, Philips TL-D ActiViva UV, Royal Philips Electronics, Amsterdam, The Netherlands). Sunrise and sunset were simulated by switching the light fixtures on and off in a successive manner over 1.5 h. The relative humidity was between 50 and 60%. Feeders with grounded pollen granulate and 30% sucrose (H2O) were offered to induce foraging. To further stimulate brood rearing we additionally provided the colonies directly with dough made by blending pollen and sugar water. Established foraging and brood-rearing patterns were typically observed 7–10 days after colonies had been transferred into the flight room. Hives were maintained for not longer than 2 months in the flight room before being replaced.
Experiment 3
We tested how disrupting brood care affects aging when the post-winter stage naturally appears in April. Frequent monitoring for brood load allowed us to determine when post-winter colonies established a regular brood care pattern. Two colony replicates were allowed to rear brood (brood-right). Another two colony replicates were prevented from brood care (broodless) by caging the queen and removing all young brood. In addition, all capped brood was removed to prevent shifts in the age distribution by hatching, young individuals. In the brood-right colonies, bees were regularly marked in order to ensure constant availability of young controls (age: 9–12 days) and of foragers with a confirmed foraging duration of at least 14 days. Controls and foragers from the brood-right colonies were contrasted with old foragers and old nest bees from broodless hives. Only the latter groups, collected from the broodless hives, had a history of being in winter state, because all brood was removed before massive emergence of young brood occurred.
Behavioral tests
Testing gustatory response to a sucrose reward before learning provides a control for the motivational state of an individual. Individuals were collected, anesthetized by chilling at 4°C, and restrained in holders as described previously (Münch et al., 2010); gustatory response scores (GRS) were measured according to Behrends et al. (Behrends et al., 2007). In brief, each individual was presented with sucrose concentrations in an ascending order: 0, 0.1, 0.3, 1, 3, 10 and 30% sucrose in H2O. In each trial bees were monitored for a proboscis extension response (PER, ‘feeding response’) upon touching the sucrose droplet that was presented to their antennae. The GRS specifies the number of trials in which bees showed a positive PER, and varies from 0 (never responding) to 7 (PER to all sucrose concentrations).
Learning scores (LS) specify the individual learning performance during olfactory conditioning of the PER (Bitterman et al., 1983). During six acquisition trials bees were trained to associate an odor (carnation oil as the conditioned stimulus, CS) with a sucrose reward (30% sucrose in H2O as the unconditioned stimulus, US). The odor was delivered through a 10 ml syringe, loaded with filter paper containing 2 μl of carnation oil. Bees were placed in front of an exhaust fan, and were presented with the odor for 5 s. During the last 2 s of odor presentation the sucrose US was applied to the antennae. Following extension of the proboscis bees were allowed to feed on the sucrose (1 ml of 30% sucrose in H2O). A bee scored positive for showing a learned response when she extended the proboscis (PER+) within the time window before US (sucrose) application. The LS specifies the total number of acquisition trials in which bees scored positive. LS values range from 5 (good learners) to 0 (no expression of the learned response). Bees that responded spontaneously to the odor (CS) before the first CS–US pairing (first trial) were discarded so that only bees that showed an acquired response were included in further experiments (Behrends et al., 2007).
Sample preparation for lipofuscin and HP gland size measurements
The different phenotypes were collected in a manner similar to that described for Experiment 2, with the only exception that sampling took place in late winter, and individuals in the old groups (winter and forager bees) were more than 5 months old. After dissection, brain and HP gland tissue samples were fixed overnight at 4°C in 4% paraformaldehyde in phosphate-buffered saline (PBS; pH 7.2), and then washed for 3×10 min in PBS. For long-term storage the samples were incubated overnight in 30% sucrose in PBS, and then covered with Tissue-Tek O.C.T. (Miles Laboratories, Elkhart, IN, USA), snap-frozen in liquid nitrogen and stored at −80°C. To prepare for imaging, thawed and rinsed brain samples were embedded in 5% low melting point agarose (Sigma-Aldrich Chemie, Steinheim, Germany). Brains were then sectioned in a frontal plane with a vibrating blade microtome (40 μm; Leica VT 1000S, Leica Biosystems, Nussloch, Germany). Before mounting, brain sections and HP gland samples were cleared by an overnight incubation in 30% glycerol in PBS, followed by 2 h incubation in 50% glycerol in PBS.
Image acquisition, analyses of lipofuscin and HP gland size
Lipofuscin was identified by its characteristic autofluorescence (maximum emission=530–650 nm) and its granular appearance (Double et al., 2008; Sheehy, 2002). Dual channel image stacks were acquired in sequential acquisition mode on a Leica TCS SP5 Laser Scanning Microscope (LSCM, Leica Microsystems, Wetzlar, Germany). Of the two channels used, one was set to longer wavelength spectra (excitation=514, 561 nm; emission=570–650 nm), and revealed lipofuscin-like fluorescence, but also non-lipofuscin structures with mostly tubular shapes (trachea, ducts). The additional second channel at shorter wavelength spectra (excitation=405 nm; emission=410–450 nm) only revealed the tubular structures, but no lipofuscin. Hence, the superposition of images from the two channels could be used to facilitate the identification of lipofuscin (only present in one channel) and its separation from structures other than lipofuscin (present in both channels; see Results). Images were taken with a 40× oil immersion objective (numerical aperture 0.75–1.25, zoom 2.5, voxel size 0.2×0.2×1 μm3). The dimensions of all single image stacks were 155×155×11 μm. Laser power and detector sensitivity were kept constant for all sample sets to limit technical variability. To further minimize the impact of technical variation between different scanning sessions (days), we always included individuals from all three focus groups in a single session.
Image stacks were processed in ImageJ (v1.44i; US National Institutes of Health, Bethesda, MD, USA). For lipofuscin measurements in HP glands, six regions of interests (ROIs) with volumes of 78×155×11 μm were inspected per individual. A Gaussian filter with constant kernel size for each data set was applied to attenuate high-frequency noise in the maximum intensity projections, and the two image channels were merged. To reduce subjective bias, the observer was blind to test phenotype identity, and was required to follow fixed rules for selecting the lipofuscin granules that were to be measured. Specifically, the observer was forced to first select the most left lipofuscin granule of each image frame. Successively, a maximum number of 20 lines was measured, each connecting a preceding particle with its nearest neighbor to the right. The last 10 lipofuscin granules of each set were outlined to assess lipofuscin granule size (area in μm2). Each individual tissue was represented by multiple ROIs to minimize selection bias: N=6 regions for lipofuscin in glands, N=6 for lipofuscin in the brain's calyx and N=4 for lipofuscin in the smaller pars intercerebralis. Comparable ROIs in gland tissue were chosen by only selecting sections through the center of acinii with the main secretory duct clearly visible in the microscope. While the pars intercerebralis is a relatively small-sized structure, we limited the selection of calyx subregions to more anterior sections with a depth of ca. 100–250 μm. Together, our analyses are based on the following total numbers of individually measured lipofuscin granules: N=1181/2051 (area/length) in HP glands, N=1495/2658 in the brain's calyx and N=1021/2019 in the brain's pars intercerebralis.
For all tissue sites we first evaluated the applicability of two parameters that could potentially characterize lipofuscin accumulation; these were the size of individual lipofuscin granules, and the nearest neighbor distances between adjacent granules (‘granule density’). In contrast to social phenotype transitions, it is well known that lipofuscin accumulates with chronological age (Fonseca et al., 2005a). This prompted us to evaluate the two parameters by comparing the groups with large differences in chronological age. Analyzing granule size could reliably identify differences between young controls and old worker types (winter and flight-room forager bees; see Table 1). In contrast, our analyses of granule density measures were found to be less sensitive to differences in chronological age. Specifically, tests based on lipofuscin granule distances could not reliably detect age-related differences for the tissues of interest (supplementary material Table S1). For detecting tissue- and phenotype-dependent lipofuscin accumulation we therefore focused on granule size measurements.
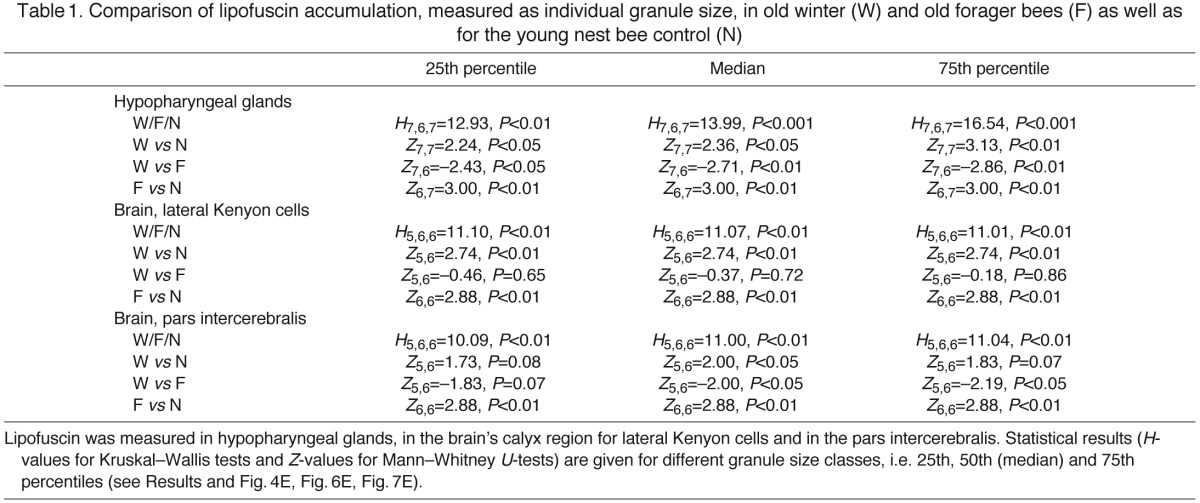
Table 1.
Comparison of lipofuscin accumulation, measured as individual granule size, in old winter (W) and old forager bees (F) as well as for the young nest bee control (N)
To measure the size of acinii in HP glands, images were obtained using a 20× oil immersion objective (numerical aperture 0.75; voxel size 0.8×0.8×2 μm). Images were viewed as maximum intensity projections representing volumes with a dimension of 775×775×12 μm. A total of 30 acinii taken from six different image locations was measured for each individual. The area of individual acinii was reconstructed by outlining undamaged acinii, which were not masked by other structures.
Statistical analysis
Statistical data were calculated using Statistica 6.0 (StatSoft, Tulsa, OK, USA). The non-parametric Kruskal–Wallis (KW) and Mann–Whitney U-tests (MWU) were applied to analyze behavioral scores (Münch et al., 2010) as well as to analyze differences in individual percentile values for lipofuscin accumulation.
RESULTS
Manipulation of behavioral senescence
Experiment 1 – no detection of behavioral senescence in bees throughout winter
First, we established that behavioral senescence was not detectable in our experimental population of old winter bees from October through March. Similar to a previous study (Behrends and Scheiner, 2010), no functional decline was detected within the observation period (H66,50,27=1.52, P=0.47, KW; Fig. 1A,B). Further, as demonstrated for the first sample point in early winter, the performance of winter bees was not poorer than the performance of the young nest bee control (Z66,68=0.63, P=0.53, MWU; Fig. 1A). Together, the longitudinal comparison between winter bees at different time points and the comparison to the young control in early winter suggest that winter bees never performed poorer than the young, non-senesced controls.
Fig. 1.

No symptom of behavioral senescence, measured as learning performance, was detected throughout 6 months of winter. (A,B) Olfactory learning performance was tested in October (early winter), January (mid winter) and March (late winter). No difference in learning score was detectable between winter bees at these time points (P=0.47, KW). Also, the learning score of young controls (young nest bees, N) was similar to that of winter bees, as established for the initial time point (early winter, P=0.53, MWU). For each learning trial the percentage of individuals with a positive response to the conditioned stimulus (PER+) is shown. (C) Likewise, gustatory responsiveness, critical for learning performance, was similar among winter stages (P=0.23, KW), and when compared with the young nest bee control (N, P=0.99, MWU). (D) Experimental colonies in late winter. (E) An indoor flight room was used to induce summer traits in former wintering colonies (cf. Fig. 2). In A and C, medians as well as upper and lower quartiles are shown for learning score and gustatory responsiveness, respectively.
An effect of colony replicate on learning performance was not detected, established by comparing individuals from three colony replicates during initial testing in early winter (winter bees, H20,22,24=0.50, P=0.78; young nest bee control, H27,22,19=2.32, P=0.31, KW). In principle, differences in responsiveness to the sucrose reward could have disguised actual differences in learning ability; for example, a higher responsiveness could have compensated for reduced learning ability in older groups (Scheiner et al., 2004). However, gustatory responsiveness was similar among all winter stages (H66,50,27=2.97, P=0.23) as well as in the young control (nest bees, Z66,68=0.01, P=0.99; Fig. 1C). Responsiveness, therefore, can be excluded as a confounding factor.
Experiment 2 – detection of behavioral senescence after summer traits were induced in mid winter
In summer, an age-related functional decline is predictably detected in bees only 2 weeks after changing to foraging tasks (see Introduction). We tested whether such rapid functional decline can also be induced in post-winter colonies that were moved into a flight room to artificially induce foraging and brood rearing (Fig. 1E). When brood load and foraging was high, we contrasted bees that had foraged for a minimum of 14 days in the flight room (old foragers) with the two groups that we had confirmed were not functionally compromised, i.e. old winter bees from outside locations (Fig. 1D) and young controls.
We show that induction of foraging was associated with a reduced learning performance compared with old winter bees and the young control (H50,56,45=17.52, P<0.001, KW; see graph for post hoc MWU tests; Fig. 2A,B). Similar to what we found previously for regular summer conditions (Münch et al., 2010), Fig. 2C indicates a more heterogeneous distribution of performance values for old foragers, as compared with less variable performance distributions in young controls and winter bees. A general impairment of learning function by the artificial environment in the flight room is unlikely, as good-performing young controls were housed together with the poor-performing foragers in the flight room. Neither replicate colony nor replicate day affected learning performance in the old focal groups of winter and forager bees (for colony effects: winter bees, Z25,25=0.13, P=0.90; forager bees, Z21,24=0.72, P=0.47; but Z24,32=2.42, P=0.02 for nest bees; for effect of day: winter bees, Z31,19=0.07, P=0.94; forager bees, Z23,22=1.17, P=0.22; nest bees, Z20,36=0.34, P=0.73; all MWU). Last, poor learning performance in foraging bees is not explained by reduced gustatory responsiveness, as all groups scored similarly high when tested for responsiveness to the reward (H50,45,56=2.42, P=0.30, KW; Fig. 2D).
Fig. 2.

Behavioral senescence became detectable when typical summer traits with brood rearing and foraging were induced in mid winter. (A,B) Two weeks after previous winter bees established a foraging pattern (F) in the flight room (Fig. 1E), their learning performance was contrasted with old winter bees (W) from colonies that remained outside, as well with young nest bees (N) that were born in the flight room. Old foragers (F) showed a reduced learning performance compared with the young control (N) and old winter bees (W, P<0.001; MWU). Learning curves are shown in B. (C) The responsiveness control was similar in all groups (P=0.30). In A and D, medians as well as upper and lower quartiles are shown for learning score and gustatory responsiveness. Significance levels of MWU post hoc tests for this and the remaining figures are depicted as follows: *P<0.05; **P<0.01; ***P<0.001.
Experiment 3 – the effect of brood rearing on behavioral senescence
In the former experiment, winter colonies were manipulated by providing artificial post-winter conditions, to which the colonies responded with increased brood rearing and foraging, whereby behavioral decline became detectable. Conversely, we now tested whether direct suppression of brood rearing in spring might counteract senescence. To this end we contrasted old worker types in post-winter colonies that were made broodless with reference groups in brood-right colonies (Fig. 3; see Experiment 3 in Materials and methods).
Fig. 3.

In spring, behavioral senescence was not detectable in colonies that were prevented from brood rearing. (A,B) After outwintering, two colonies were allowed to establish a brood-rearing pattern that is typical for summer bees (F, foragers; N, young nest bees). In contrast, two post-winter colonies were prevented from brood care by caging the queen and removing all brood rearing, while the former winter bees were separated into old foragers (FW) and old nest bees (W). In foragers, declined learning performance was only detected in colonies with brood (F) as compared with foragers from colonies that were made broodless (FW; P<0.001). Learning curves for all groups are shown in B. (C) A lower gustatory responsiveness was only detected for the young nest bee control (P<0.05). Thus, gustatory responsiveness cannot explain the apparent lack of behavioral senescence in old groups (W, FW) from the broodless colonies compared with foragers from the brood-right colonies (for further statistical details, see Results). In A and C, medians as well as upper and lower quartiles are shown for learning score and gustatory responsiveness.
While after 14 days of foraging the vast majority of foragers typically dies (Dukas, 2008; Wolschin et al., 2009), we did not observe such decrease in old foragers of broodless hives. Similarly, we did not observe a pronounced increase of poor-learning individuals 14 days after foraging was first confirmed for these individuals in the broodless colonies (data not shown). Several samplings confirmed this observation, of which Fig. 3 shows behavioral data for the time point when at least at least 70 days had passed after old foragers were returning from first foraging flights. A reduced learning performance was only detectable in foragers of brood-right colonies after 14 days of foraging (F in Fig. 3A,B), as compared with learning performances of old foragers (FW) and old nest bees (W) from the broodless colonies, and the young control from the brood-right colony (N in Fig. 3A,B; H35,34,43,33=33.03, P<0.001, KW; see graph for post hoc MWU).
Comparisons between the two replicate days and two replicate colonies did not reveal significant replicate effects that would explain the apparent lack of behavioral senescence in old foragers (F in Fig. 3A,B) from broodless hives (for colony effects: Z18,15=0.11, P=0.91; for effect of day: Z18,15=0.69, P=0.49; both MWU). We observed a slightly higher gustatory responsiveness in bees from the broodless hives compared with the young control, but not compared with the poor-performing foragers of the brood-right colonies (H35,34,43,33=11.24, P<0.05, see graph for post hoc MWU; Fig. 3C). Therefore, gustatory responsiveness also does not explain the disparate learning performance between foragers in the brood-right and broodless colonies.
Taken together, behavioral senescence only became detectable when brood rearing and foraging were induced in post-winter colonies (Experiment 2), but not when former winter hives were prevented from brood rearing (broodless colonies in Experiment 3.).
Inducible cellular senescence differs between tissues
We next asked whether inducible decline patterns are also manifested at the level of cellular senescence. We therefore contrasted lipofuscin accumulation in the young controls with levels in old winter bees, and in old foragers after the post-winter state was induced in the flight room (see Experiment 2).
Lipofuscin accumulation in HP glands
We first assessed cellular senescence in HP glands, as these are major storage sites for metabolites and may shape organismal aging through gain or loss of vital resources (Lee, 2003; Münch and Amdam, 2010). Representative micrographs in Fig. 4A–C show different levels of lipofuscin accumulation for the three worker castes: few, small granules for young controls (Fig. 4A), several medium-sized granules for old winter bees (Fig. 4B), and many medium-sized to large lipofuscin granules for old flight-room foragers (Fig. 4C). Phenotype dependent differences between worker castes are further substantiated by data on granule size distribution for all tested individuals (Fig. 4D). These differences become most apparent when comparing the larger granule size classes of each individual, e.g. granules that are larger than the 75th percentile value of the individual size distribution curves. To test for differences between worker types, every individual in each phenotype group was represented by the 25th, 50th (median) and 75th percentile value of the individual size distribution curves (Fig. 4E, cf. Fig. 4D). This was done, because age-specific cellular senescence can develop heterogeneously among cells (Herndon et al., 2002), and accumulation of aging biomarkers therefore might be detectable only when considering cells that accumulate the largest granules. However, for HP glands we detected significant differences between age groups and worker castes, independent of whether the analyses focused on smallest, median or largest granule classes per individual (25%, 50% and 75% of maximum granule size in Fig. 4E; see Table 1 for statistics). Taken together, young controls showed the lowest, and old winter bees the most intermediate, lipofuscin accumulation. Yet lipofuscin accumulation was significantly increased shortly after old bees transformed into foragers.
Fig. 4.

Lipofuscin-like (LF) fluorescent granules (‘age pigments’) are largest in the hypopharyngeal (HP) glands of forager bees. In contrast, winter bees of similar chronological age represent an intermediate stage of LF accumulation. (A–C) Projection views of image stacks showing LF fluorescence (dark gray to black, depicted with arrows). Representative examples for the young nest bee control (A), and for old winter (B) and old forager bees (C). For better identification of LF signals, images were taken in dual channel acquisition mode, i.e. at shorter and longer wavelengths simultaneously (see Materials and methods). Of these, the signals recorded at shorter wavelengths were used as a control, and depict non-specific autofluorescence (green) but not the LF-specific signal with autofluorescence at longer wavelengths (grayscale, arrows). (D) Distributions of LF granule sizes are shown for all individuals separately, with size percentiles sorted from smallest to largest. Note that extremely large LF granules were frequently observed in foragers (green lines, clipped at 150 μm2), as compared with winter bees (blue circles) and young control (gray lines). The dashed vertical lines depict quartiles and median values. (E) We assessed differences between the three phenotypes for small, median and large LF size classes (25th, 50th and 75th percentiles, see D). For all size classes, winter bees (W) showed intermediate LF sizes, significantly smaller than in foragers (F) but larger than in the young nest bee controls (N). Medians and quartiles for each phenotype are displayed. For statistical details, see Table 1. Scale bar in A for A–C, 20 μm.
Increased lipofuscin size can theoretically result both from a higher accumulation of cellular turnover products and from contracting gland tissue that seemingly compresses granules to larger aggregates. However, different accumulation of lipofuscin in winter bees and young controls cannot be explained by differences in gland size, as our data did not show significant differences in gland area measurements between these groups (HW=7,F=6,N=7=12.74, P<0.01, KW; ZW=7,N=7=−1.21, P=0.22, MWU; Fig. 5). Only for old foragers, for which we detected significant gland size differences compared with young controls (ZF=6,N=7=−3.00, P<0.01, MWU) and winter bees (ZW=7,F=6=3.00, P<0.01, MWU), might the high lipofuscin accumulation levels be partly explained by contracted gland tissue.
Fig. 5.

Acinii (‘units’) of HP glands are similar in size in old winter bees (W) and young controls (nest bees, N), but are significantly reduced in bees 14 days after foraging was initiated in the flight room (foragers, F). (A–C) Projection views of image stacks showing HP glands with single acinii (asterisks) and central ducts (arrow). Representative examples for young control (A), winter (B) and forager bees (C). The inset in A depicts reconstructed acinii as used for size measurements (N=28–30 area values per individual). (D) Acinii size distributions separately for all individuals, with size percentiles sorted from smallest to largest and the median value depicted by a vertical dashed line. Note that in contrast to foragers (green lines), the acinii size distributions differ considerably between individuals, both in young control (gray lines) and winter bees (blue circles). (E) Acinii size was significantly reduced in old foragers (F) compared with young nest bees (N; HW=7,F=6,N=7=12.74, P<0.01, KW; ZF=6,N=7=−3.00, P<0.01, MWU) and old winter bees (W; ZF=6,W=7=−3.00, P<0.01, MWU). Acinii size was not different between winter and young nest bees (ZW=7,N=7=−1.21, P=0.22, MWU). Medians and quartiles for all three phenotypes are displayed. For statistical details, see Table 1. Scale bar in C for A–C, 200 μm.
Lipofuscin accumulation in the calyx region of the brain
Cellular senescence in brain cells may directly compromise behavioral function, including learning function (see Results, Manipulation of behavioral senescence). Therefore, we assessed lipofuscin accumulation in lateral Kenyon cells of the mushroom bodies' calyx region, a major learning center of the honey bee brain (Fig. 6A–C; for complete statistical data, see Table 1). We found that calyx neurons in both winter bees and flight-room foragers had significantly larger lipofuscin granule sizes than those of the young controls. This was true for all three size classes assessed per individual (25th, 50th and 75th percentile size classes, Fig. 6D,E; W vs N and F vs N, Table 1). These data suggest that lipofuscin in the brain's calyx region accumulates with chronological age. In contrast, the caste transition from old winter to old forager bees was not associated with increased lipofuscin granule sizes (25th, 50th and 75th percentile size classes, Fig. 6D,E; W vs F, in Table 1).
Fig. 6.

Lipofuscin (LF) accumulation in the calyx region of the brain was similar in winter and forager bees. (A–C) Projection views of image stacks showing LF fluorescence (arrows) in the lateral calyx region of the mushroom bodies. Representative examples for young control (A), old winter (B) and old forager bees (C; for color scheme and age details, see Fig. 4). (D) Distributions of LF granule sizes for individual winter bees (blue circles), foragers (green lines) and young controls (gray lines). The dashed vertical lines depict quartiles and median values. (E) Potential group differences between the three phenotypes were assessed for small, median and large LF size classes, i.e. for the 25th, 50th and 75th percentile value of each individual (see D). For all size classes, old winter bees (W) were different from young nest bee controls (N), but not from old foragers (F). Medians and quartiles for each phenotype are displayed. For statistical details, see Table 1. Scale bar in A for A–C, 20 μm.
Lipofuscin accumulation in the brain's pars intercerebralis
Last, we quantified lipofuscin accumulation in neuronal somata of the pars intercerebralis, an area that includes many neuropeptidergic neurons (Fig. 7A–C; statistical data in Table 1). In contrast to the calyx region, we found that lipofuscin accumulation in the pars intercerebralis had increased with the transition from winter to the forager caste (Fig. 7D,E; W vs F, Table 1). This was true for the larger size classes that were found per individual (50th and 75th percentiles, Fig. 7D,E). Similarly, foragers had significantly larger lipofuscin granules than the chronologically much younger controls (F vs N, Table 1). In contrast, despite the large chronological age difference between winter bees and young controls, an increased lipofuscin accumulation was not clear, and was only detectable in one out of three size classes that were assessed per individual (50th percentile, Fig. 7D,E; F vs N in Table 1).
Fig. 7.

Lipofuscin (LF) accumulation in midline areas of the central brain (pars intercerebralis) was increased after old winter bees changed to foraging. (A–C) Projection views of image stacks showing LF fluorescence (arrows) in the pars intercebralis, a major neurosecretory center. Representative examples for young control (A), old winter (B) and old forager bees (C; for color scheme and age details, see Fig. 4). (D) Distributions of LF granule sizes for individual bees. Note the differences between forager bees (green lines), winter bees (blue circles) and young control bees (gray lines) in the class with largest LF granules (≥75th percentile). (E) Group differences between old winter (W), old forager (F) and young nest control bees (N) were evaluated for small, median and large LF size classes (25th, 50th and 75th percentile values of each individual; see D). Except for the smallest class (25th percentile), old foragers showed significantly larger LF granules than old winter bees. In addition, LF granules of winter bees were not different from young controls in two out of three size classes (25th and 50th; see Table 1). Medians and quartiles for each phenotype are shown. For statistical details, see Table 1. Scale bar in A for A–C, 20 μm.
DISCUSSION
Here we establish how accelerated and slowed senescence in bees can emerge in response to environmental changes. Behavioral senescence was assessed for olfactory learning performance, and remained undetectable in old winter bees throughout winter (Fig. 1). We detected behavioral decline shortly after old winter bees were moved into a flight room to induce post-winter traits with changed social task behaviors (the transition to foraging) and changed social environment (increased brood load; Fig. 2). Similar to behavioral decline, changes in cellular senescence, measured as lipofuscin accumulation, also became rapidly detectable after disrupting the winter state and transition to forager tasks (Figs 4, 7). In contrast to the lack of behavioral senescence, levels of cellular senescence were typically higher in old winter bees than in young controls (Figs 4, 7). We found that cellular senescence was tissue dependent, and most prominent in an organ that serves brood nourishment and metabolite storage (HP glands; Fig. 4A–C) compared with areas in the brain (Fig. 6A–C, Fig. 7A–C). Finally, by preventing post-winter colonies from brood rearing, behavioral senescence was not detectable even after transition to the forager state had occurred (Fig. 3). Overall, our data suggest that rapid age-related decline can be induced by changing social task behavior in winter bees. However, this link is contingent on social environment, as reduced brood load can counteract senescence even after old winter bees had changed to foraging tasks.
Our data on negligible behavioral senescence during normal winter conditions (Figs 1, 2) conform with a previous report that studied two different learning behaviors, olfactory and tactile acquisition (Behrends and Scheiner, 2010). In our study and in the previous studies on summer bees (Behrends et al., 2007; Scheiner and Amdam, 2009), groups with declined learning performance did not show changes in a task that depends on antennal function, GRS. Hence, these data do not indicate that severe ‘wear and tear’ effects in the antennae of foragers would contribute to declined learning performance. Rather, declined learning performance could be due to changes in the central brain, as shown earlier for synaptic protein levels (Wolschin et al., 2009). Further, the performance decline 14 days after foraging was initiated in the flight room (Fig. 2) is in accord with previous work on bees under natural summer conditions (Behrends et al., 2007; Münch et al., 2010; Scheiner and Amdam, 2009; Tolfsen et al., 2011). Significant decline after approximately 2 weeks of foraging both in summer and in post-winter bees makes it unlikely that a recent history of being in a winter state confers additional survival benefits over summer workers. Rather, extensive flight performance may pose perils (Magwere et al., 2006; Sohal, 1976; Williams et al., 2008) that efficiently level out physiological differences in worker bees with very different life histories, i.e. between comparably young summer and old post-winter foragers. However, the time point at which senescence becomes detectable after foraging had been initiated is not static, but can be delayed by restricting the daily foraging time (Tolfsen et al., 2011). It is therefore plausible that lowered flight activity can partly explain the lack of behavioral senescence in foragers from post-winter colonies that were made broodless (Fig. 3). In agreement with this assertion are observations on reduced foraging intensity in broodless hives (Free, 1967; Huang and Otis, 1989).
In conclusion, we propose that removal of brood and likely the lack of brood pheromone (Smedal et al., 2009) may protect from behavioral senescence through suppression of flight activity. Hence, our senescence measurements suggest that lifespan extension as previously observed for hives under broodless conditions (Fluri et al., 1982; Maurizio, 1950) can be linked to extended periods of negligible senescence. Other traits, such as thermoregulatory responses, can be associated with brood care and the long-lived winter state (Omholt, 1987). Reduced thermoregulation in broodless colonies may therefore have affected lifespan and aging in nest bees, but is unlikely to have affected lifespan in the forager caste (Fig. 3). The complex interaction of brood load, foraging age and foraging intensity may also account for an apparent discrepancy between our study and a previous study on winter bees by Behrends and Scheiner (Behrends and Scheiner, 2010). While the former authors could not establish a significant difference between indoor foragers and outdoor winter bees, our work (Fig. 2) and the previous report are not directly comparable. Behrends and Scheiner did not determine the minimum foraging age, and also did not directly address transitional changes in mid winter, but studied specimens in early autumn (Behrends and Scheiner, 2010). These seasonal differences might explain the low brood load they observed, contrasting our observations of intense egg laying and brood care in flight-room colonies, which again may entail different foraging intensities and hence senescence levels.
In contrast to behavioral decline, our data on cellular senescence suggest that both social factors and chronological age affect lipofuscin levels, and that effects differ between tissues. Specifically, we found that social caste transition from old winter to old forager bees was associated with increased lipofuscin levels, but only for HP glands (Fig. 4) and the brain's pars intercerebralis (Fig. 7) – not for the mushroom bodies' calyx region (Fig. 6). Strong lipofuscin accumulation in HP glands of old foragers was associated with a significant reduction in gland size (Fig. 5). This is in line with documented atrophic events that lead to a marked degeneration in HP glands, when nest bees change to foraging tasks (Fluri et al., 1982; Haydak, 1957; Maurizio, 1954). Atrophy of these glands is complex, and involves different modes of cell deaths with typical apoptosis but also necrosis symptoms (Silva de Moraes and Bowen, 2000). To our knowledge, it is not known how prevalences of programmed cell death and senescence-like events change when foragers age. For HP glands it is therefore not clear to what extent increased lipofuscin accumulation may also result from processes not typically considered as senescence.
For brain tissue, the comparison of Figs 6 and 7 reveals regional differences between old winter bees and old foragers. This implies that after caste transition lipofuscin accumulation rates are accelerated in the pars intercerebralis, a neurosecretory center. Although during normal aging, significant accumulation in the brain's pars intercerebralis is also documented for another insect, the African migratory locust (Locusta migratoria) (Fonseca et al., 2005a). Accumulation rates in this area were shown to reflect death rates, and thus increased frailty among individuals. Similarly, our data on functional decline in old foragers (Fig. 2) are reflected by increased lipofuscin accumulation in the pars intercerebralis (Fig. 7). In the calyx region, however, lipofuscin levels remained unchanged in old foragers, suggesting that cellular senescence after caste transition is absent or slower than in the pars intercerebralis. In accord, by using mass spectrometry and quantitative anatomy, we recently demonstrated that the protein and synapse matrices of the calyx were not changed in aging summer foragers. This is despite protein level changes detected for other areas of the central brain (Wolschin et al., 2009), and the behavioral decline typical for this worker caste (Behrends et al., 2007; Dukas, 2008; Münch et al., 2010). Cumulative evidence therefore suggests that the calyx, a major learning center, might show structural and molecular exceptions from accelerated aging patterns that are observed in other brain regions. Notably, previous studies that focused on calyx maturation rather than aging documented a persistent volumetric growth and neuronal branching (Durst et al., 1994; Fahrbach et al., 2003; Farris et al., 2001), as well as experience-dependent changes of the synaptic architecture (Groh and Meinertzhagen, 2010; Hourcade et al., 2010; Krofczik et al., 2008). Whether such patterns of persistent plasticity and growth can protect from rapid cellular senescence in the calyx is an intriguing question that is to be addressed by future studies.
Apart from phenotype-dependent differences in lipofuscin accumulation, we also detected increased levels of lipofuscin in both old groups (winter and forager bees) compared with the young control (Table 1, Figs 4, 6, 7). That old winter bees with negligible behavioral senescence (Fig. 1) still showed increased lipofuscin levels might indicate that behavioral senescence is preceded by cellular senescence. However, comparable studies also measured moderate lipofuscin levels throughout early adult life, followed by a marked increase towards the end of life, which is linked to functional decline and increased mortality (Double et al., 2008; Fonseca et al., 2005a). Similarly, long lifespan and lack of detectable behavioral decline in winter bees (Figs 1, 2) suggest that intermediate lipofuscin levels in old winter bees are not associated with harmful cellular deterioration.
On the cellular and molecular levels, diverse factors are proposed to influence senescence; among them are lipofuscin and excessive oxidative stress (Holliday, 2006). Lipofuscin, therefore, is not only considered a consequence of aging but may itself spur cellular, and specifically neuronal, degeneration (Sulzer et al., 2008). Studies in mammalian models suggest that lipofuscin accumulation can deplete the autophagic capacity of a cell (Terman, 2006), leading to incomplete turnover of cellular organelles, in particular mitochondria. Thereby dysfunctional – aged – mitochondria can no longer be replaced, and afflicted cells will deteriorate (Brunk and Terman, 2002). While accumulation rates of lipofuscin in vertebrate models are comparably slow, and are tied to chronological age, our study suggests that lipofuscin accumulation in the bee model can be highly dynamic, decoupled from chronological age, and is amenable for manipulation. Comparatively rapid, age-related accumulation rates over only few weeks are corroborated by recent study in honey bee fatbody cells (Hsieh and Hsu, 2011). In contrast, dynamic modulation, which in our study is associated with social phenotype transitions, is not well documented. Possible modulation was also reported for other invertebrates (decapodes), where lipofuscin accrual was found to be reversed after surgical treatments (Fonseca et al., 2005b). In conclusion, with a highly dynamic accumulation of lipofuscin (e.g. Fig. 3) and the potential to revert aging phenotypes (Amdam et al., 2005; Huang and Robinson, 1996), the honey bee offers rewarding perspectives to study how a most universal manifestation of cellular aging can be slowed or reversed.
ACKNOWLEDGEMENTS
We thank the two anonymous reviewers and the editor for helpful feedback on our manuscript.
LIST OF ABBREVIATIONS
- CS
conditioned stimulus
- GRS
gustatory responsiveness
- HP
hypopharyngeal
- KW
Kruskal–Wallis analysis of variance
- LS
learning score
- MWU
Mann–Whitney U-test
- PER
proboscis extension response
- US
unconditioned stimulus
FOOTNOTES
Supplementary material available online at http://jeb.biologists.org/cgi/content/full/216/9/1638/DC1
COMPETING INTERESTS
No competing interests declared.
FUNDING
G.V.A. and D.M. were supported by the Norwegian Research Council (180504, 191699, 213976/F20); G.V.A. received support from the PEW Charitable Trust, the National Institute on Aging (PO1AG22500).
REFERENCES
- Amdam G. V. (2010). Social context, stress, and plasticity of aging. Aging Cell 10, 18-27 [DOI] [PubMed] [Google Scholar]
- Amdam G. V., Omholt S. W. (2002). The regulatory anatomy of honeybee lifespan. J. Theor. Biol. 216, 209-228 [DOI] [PubMed] [Google Scholar]
- Amdam G. V., Simões Z. L., Hagen A., Norberg K., Schrøder K., Mikkelsen O., Kirkwood T. B., Omholt S. W. (2004). Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Exp. Gerontol. 39, 767-773 [DOI] [PubMed] [Google Scholar]
- Amdam G. V., Aase A. L., Seehuus S. C., Kim Fondrk M., Norberg K., Hartfelder K. (2005). Social reversal of immunosenescence in honey bee workers. Exp. Gerontol. 40, 939-947 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ament S. A., Corona M., Pollock H. S., Robinson G. E. (2008). Insulin signaling is involved in the regulation of worker division of labor in honey bee colonies. Proc. Natl. Acad. Sci. USA 105, 4226-4231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrends A., Scheiner R. (2010). Learning at old age: a study on winter bees. Front. Behav. Neurosci. 4, 15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrends A., Scheiner R., Baker N., Amdam G. V. (2007). Cognitive aging is linked to social role in honey bees (Apis mellifera). Exp. Gerontol. 42, 1146-1153 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bitterman M. E., Menzel R., Fietz A., Schäfer S. (1983). Classical conditioning of proboscis extension in honeybees (Apis mellifera). J. Comp. Psychol. 97, 107-119 [PubMed] [Google Scholar]
- Brunk U. T., Terman A. (2002). The mitochondrial-lysosomal axis theory of aging: accumulation of damaged mitochondria as a result of imperfect autophagocytosis. Eur. J. Biochem. 269, 1996-2002 [DOI] [PubMed] [Google Scholar]
- Comfort A. (1956). The Biology of Senescence. London: Routledge and Paul; [Google Scholar]
- Corona M., Velarde R. A., Remolina S., Moran-Lauter A., Wang Y., Hughes K. A., Robinson G. E. (2007). Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity. Proc. Natl. Acad. Sci. USA 104, 7128-7133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crailsheim K. (1991). Interadult feeding of jelly in honeybee (Apis mellifera L.) colonies. J. Comp. Physiol. B 161, 55-60 [Google Scholar]
- Crailsheim K. (1998). Trophallactic interactions in the adult honeybee (Apis mellifera L.). Apidologie 29, 97-112 [Google Scholar]
- Double K. L., Dedov V. N., Fedorow H., Kettle E., Halliday G. M., Garner B., Brunk U. T. (2008). The comparative biology of neuromelanin and lipofuscin in the human brain. Cell. Mol. Life Sci. 65, 1669-1682 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dukas R. (2008). Mortality rates of honey bees in the wild. Insect. Soc. 55, 252-255 [Google Scholar]
- Durst C., Eichmüller S., Menzel R. (1994). Development and experience lead to increased volume of subcompartments of the honeybee mushroom body. Behav. Neural Biol. 62, 259-263 [DOI] [PubMed] [Google Scholar]
- Fahrbach S. E., Farris S. M., Sullivan J. P., Robinson G. E. (2003). Limits on volume changes in the mushroom bodies of the honey bee brain. J. Neurobiol. 57, 141-151 [DOI] [PubMed] [Google Scholar]
- Farris S. M., Robinson G. E., Fahrbach S. E. (2001). Experience- and age-related outgrowth of intrinsic neurons in the mushroom bodies of the adult worker honeybee. J. Neurosci. 21, 6395-6404 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finch C. E., Austad S. N. (2001). History and prospects: symposium on organisms with slow aging. Exp. Gerontol. 36, 593-597 [DOI] [PubMed] [Google Scholar]
- Fluri P. (1990). How long do worker honeybees live? Schweizerische Bienen-Zeitung 113, 620-625 [Google Scholar]
- Fluri P., Wille H., Gerig L., Lüscher M. (1977). Juvenile hormone, vitellogenin and haemocyte composition in winter worker honeybees (Apis mellifera). Experientia 33, 1240-1241 [Google Scholar]
- Fluri P., Lüscher M., Wille H., Gerig L. (1982). Changes in weight of the pharyngeal gland and haemolymph titres of juvenile hormone, protein and vitellogenin in worker honey bees. J. Insect Physiol. 28, 61-68 [Google Scholar]
- Fonseca D. B., Brancato C. L., Prior A. E., Shelton P. M., Sheehy M. R. (2005a). Death rates reflect accumulating brain damage in arthropods. Proc. Biol. Sci. 272, 1941-1947 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fonseca D. B., Sheehy M. R., Blackman N., Shelton P. M., Prior A. E. (2005b). Reversal of a hallmark of brain ageing: lipofuscin accumulation. Neurobiol. Aging 26, 69-76 [DOI] [PubMed] [Google Scholar]
- Free J. B. (1967). Factors determining the collection of pollen by honeybee foragers. Anim. Behav. 15, 134-144 [DOI] [PubMed] [Google Scholar]
- Groh C., Meinertzhagen I. A. (2010). Brain plasticity in Diptera and Hymenoptera. Front. Biosci. S2, 268-288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grotewiel M. S., Martin I., Bhandari P., Cook-Wiens E. (2005). Functional senescence in Drosophila melanogaster. Ageing Res. Rev. 4, 372-397 [DOI] [PubMed] [Google Scholar]
- Haydak M. H. (1957). Changes with age in the appearance of some internal organs of the honeybee. Bee World 38, 197-207 [Google Scholar]
- Herndon L. A., Schmeissner P. J., Dudaronek J. M., Brown P. A., Listner K. M., Sakano Y., Paupard M. C., Hall D. H., Driscoll M. (2002). Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans. Nature 419, 808-814 [DOI] [PubMed] [Google Scholar]
- Holliday R. (2006). Aging is no longer an unsolved problem in biology. Ann. New York. Acad. Sci. 1067, 1-9 [DOI] [PubMed] [Google Scholar]
- Hourcade B., Muenz T. S., Sandoz J. C., Rössler W., Devaud J. M. (2010). Long-term memory leads to synaptic reorganization in the mushroom bodies: a memory trace in the insect brain? J. Neurosci. 30, 6461-6465 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsieh Y. S., Hsu C. Y. (2011). Honeybee trophocytes and fat cells as target cells for cellular senescence studies. Exp. Gerontol. 46, 233-240 [DOI] [PubMed] [Google Scholar]
- Huang Z. Y., Otis G. W. (1989). Factors determining hypopharyngeal gland activity of worker honey bees (Apis mellifera L.). Insectes Soc. 36, 264-276 [Google Scholar]
- Huang Z.-Y., Robinson G. E. (1996). Regulation of honey bee division of labor by colony age demography. Behav. Ecol. Sociobiol. 39, 147-158 [Google Scholar]
- Keller L., Jemielity S. (2006). Social insects as a model to study the molecular basis of ageing. Exp. Gerontol. 41, 553-556 [DOI] [PubMed] [Google Scholar]
- Krofczik S., Khojasteh U., de Ibarra N. H., Menzel R. (2008). Adaptation of microglomerular complexes in the honeybee mushroom body lip to manipulations of behavioral maturation and sensory experience. Dev. Neurobiol. 68, 1007-1017 [DOI] [PubMed] [Google Scholar]
- Lee R. D. (2003). Rethinking the evolutionary theory of aging: transfers, not births, shape senescence in social species. Proc. Natl. Acad. Sci. USA 100, 9637-9642 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindauer M. (1963). Allgemeine Sinnesphysiologie. Orientierung im Raum. Fortschr. Zool. 16, 58-140 [PubMed] [Google Scholar]
- Magwere T., Pamplona R., Miwa S., Martinez-Diaz P., Portero-Otin M., Brand M. D., Partridge L. (2006). Flight activity, mortality rates, and lipoxidative damage in Drosophila. J. Gerontol. A Biol. Sci. Med. Sci. 61, 136-145 [DOI] [PubMed] [Google Scholar]
- Maurizio A. (1950). The influence of pollen feeding and brood rearing on the length of life and physiological condition of the honeybee preliminary report. Bee World 31, 9-12 [Google Scholar]
- Maurizio A. (1954). Pollenernahrung und lebensvorgange bei der honigbiene (Apis mellifera L.). Landwirtsch. Jahrb. Schweiz. 245, 115-182 [Google Scholar]
- Münch D., Amdam G. V. (2010). The curious case of aging plasticity in honey bees. FEBS Lett. 584, 2496-2503 [DOI] [PubMed] [Google Scholar]
- Münch D., Baker N., Kreibich C. D., Bråten A. T., Amdam G. V. (2010). In the laboratory and during free-flight: old honey bees reveal learning and extinction deficits that mirror mammalian functional decline. PLoS ONE 5, e13504 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson C. M., Ihle K. E., Fondrk M. K., Page R. E., Amdam G. V. (2007). The gene vitellogenin has multiple coordinating effects on social organization. PLoS Biol. 5, e62 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neukirch A. (1982). Dependence of the life span of the honeybee (Apis mellifica) upon flight performance and energy consumption. J. Comp. Physiol. 146, 35-40 [Google Scholar]
- Nussey D. H., Coulson T., Festa-Bianchet M., Gaillard J. M. (2008). Measuring senescence in wild animal populations: towards a longitudinal approach. Funct. Ecol. 22, 393-406 [Google Scholar]
- Omholt S. W. (1987). Thermoregulation in the winter cluster of the honeybee, Apis mellifera. J. Theor. Biol. 128, 219-231 [Google Scholar]
- Parker J. D. (2010). What are social insects telling us about aging? Myrmecological News 13, 103-110 [Google Scholar]
- Pernal S. F., Currie R. W. (2001). Improved flight and rearing room design for honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 94, 793-805 [DOI] [PubMed] [Google Scholar]
- Roberts S. P., Elekonich M. M. (2005). Muscle biochemistry and the ontogeny of flight capacity during behavioral development in the honey bee, Apis mellifera. J. Exp. Biol. 208, 4193-4198 [DOI] [PubMed] [Google Scholar]
- Robinson G. E., Page R. E., Strambi C., Strambi A. (1992). Colony integration in honey bees: mechanisms of behavioral reversion. Ethology 90, 336-348 [Google Scholar]
- Scheiner R., Amdam G. V. (2009). Impaired tactile learning is related to social role in honeybees. J. Exp. Biol. 212, 994-1002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheiner R., Page R. E., Erber J. (2004). Sucrose responsiveness and behavioral plasticity in honey bee (Apis mellifera). Apidologie 35, 133-142 [Google Scholar]
- Seehuus S. C., Krekling T., Amdam G. V. (2006a). Cellular senescence in honey bee brain is largely independent of chronological age. Exp. Gerontol. 41, 1117-1125 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seehuus S. C., Norberg K., Gimsa U., Krekling T., Amdam G. V. (2006b). Reproductive protein protects functionally sterile honey bee workers from oxidative stress. Proc. Natl. Acad. Sci. USA 103, 962-967 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seeley T. D. (1995). The Wisdom of the Hive. Cambridge, MA: Harvard University Press; [Google Scholar]
- Sheehy M. R. (2002). A flow-cytometric method for quantification of neurolipofuscin and comparison with existing histological and biochemical approaches. Arch. Gerontol. Geriatr. 34, 233-248 [DOI] [PubMed] [Google Scholar]
- Silva de Moraes R. L. M., Bowen I. D. (2000). Modes of cell death in the hypopharyngeal gland of the honey bee (Apis mellifera L). Cell Biol. Int. 24, 737-743 [DOI] [PubMed] [Google Scholar]
- Smedal B., Brynem M., Kreibich C. D., Amdam G. V. (2009). Brood pheromone suppresses physiology of extreme longevity in honeybees (Apis mellifera). J. Exp. Biol. 212, 3795-3801 [DOI] [PubMed] [Google Scholar]
- Sohal R. S. (1976). Aging changes in insect flight muscle. Gerontology 22, 317-333 [DOI] [PubMed] [Google Scholar]
- Sulzer D., Mosharov E., Talloczy Z., Zucca F. A., Simon J. D., Zecca L. (2008). Neuronal pigmented autophagic vacuoles: lipofuscin, neuromelanin, and ceroid as macroautophagic responses during aging and disease. J. Neurochem. 106, 24-36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tatar M., Yin C. (2001). Slow aging during insect reproductive diapause: why butterflies, grasshoppers and flies are like worms. Exp. Gerontol. 36, 723-738 [DOI] [PubMed] [Google Scholar]
- Tatar M., Chien S. A., Priest N. K. (2001). Negligible senescence during reproductive dormancy in Drosophila melanogaster. Am. Nat. 158, 248-258 [DOI] [PubMed] [Google Scholar]
- Terman A. (2006). Catabolic insufficiency and aging. Ann. New York Acad. Sci. 1067, 27-36 [DOI] [PubMed] [Google Scholar]
- Tolfsen C. C., Baker N., Kreibich C., Amdam G. V. (2011). Flight restriction prevents associative learning deficits but not changes in brain protein-adduct formation during honeybee ageing. J. Exp. Biol. 214, 1322-1332 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vance J. T., Williams J. B., Elekonich M. M., Roberts S. P. (2009). The effects of age and behavioral development on honey bee (Apis mellifera) flight performance. J. Exp. Biol. 212, 2604-2611 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams J. B., Roberts S. P., Elekonich M. M. (2008). Age and natural metabolically-intensive behavior affect oxidative stress and antioxidant mechanisms. Exp. Gerontol. 43, 538-549 [DOI] [PubMed] [Google Scholar]
- Winston M. L. (1980). Seasonal patterns of brood rearing and worker longevity in colonies of the Africanized honey bee (Hymenoptera: Apidae) in South America. J. Kans. Entomol. Soc. 53, 157-165 [Google Scholar]
- Wolschin F., Amdam G. V. (2007). Comparative proteomics reveal characteristics of life-history transitions in a social insect. Proteome Sci. 5, 10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolschin F., Münch D., Amdam G. V. (2009). Structural and proteomic analyses reveal regional brain differences during honeybee aging. J. Exp. Biol. 212, 4027-4032 [DOI] [PMC free article] [PubMed] [Google Scholar]