

Abstract
Complex systems ensure resilience through multiple controllers acting at rapid and slower timescales. The need for efficient information flow through complex systems encourages small-world network structures. On the basis of these principles, a group of regions associated with top-down control was examined. Functional magnetic resonance imaging showed that each region had a specific combination of control signals; resting-state functional connectivity grouped the regions into distinct ‘fronto-parietal’ and ‘cingulo-opercular’ components. The fronto-parietal component seems to initiate and adjust control; the cingulo-opercular component provides stable ‘set-maintenance’ over entire task epochs. Graph analysis showed dense local connections within components and weaker ‘long-range’ connections between components, suggesting a small-world architecture. The control systems of the brain seem to embody the principles of complex systems, encouraging resilient performance.
Introduction
The brain learns and adapts to environmental change, while showing resilience to local perturbation and damage. Complex adaptive systems seem to follow common organizational principles across many levels of scale, from subcellular components to social systems that resolve the tension between adaptability and resilience [1–4]. Here, we focus on two of these principles that illuminate the organization of the brain at the systems level: (i) The importance of multiple controlling variables, and (ii) the small-world architecture of efficient information-processing networks.
Complex biological and social systems are often driven by several separate control mechanisms with distinct functional properties [1]. Because the number of controlling variables is usually at least two, but fewer than ten, this principle has been named the ‘rule of hand’ [1]. The different controlling variables often affect the overall state of the system through distinct mechanisms that operate on separable temporal scales [5,6]. Systems with distinct rapid-acting and more slowly changing controlling variables can simultaneously be highly stable, yet flexible. For example, the ecological state of a forest can be rapidly affected by changes in the number of leaf-eating insects and more slowly by changes in the growth of large tree species. The presence of multiple control mechanisms also increases the resilience of a system to perturbation. For example, our sense of balance is supported in parallel by the vestibular system, the visual system and peripheral joint-position sensors. If any one of these three control variables is impaired, some level of balance is still maintained by the remaining mechanisms.
Information flow through complex networks of nodes can be made efficient by structuring the flow between the nodes in certain ways. Networks consisting of multiple densely connected clusters with small numbers of connections between clusters (i.e. small-world networks) are more efficient at information transmission (Figure 1) than are either randomly connected or highly regular lattice networks [2,3,7]. Such ‘small-world’ networks are ubiquitous. For example, the anatomical connections of the macaque visual system and the neuronal connections of Caenorhabditis elegans have both been described as small-world networks [2].
Figure 1.
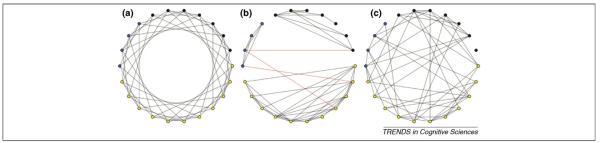
Schematic depicting regular, small-world and random graphs. Small-world graphs can be generated by replacing local connections in regular (lattice) graphs with longer-range short-cuts. (a) Sample regular graph (lattice) containing 22 nodes and 52 edges. In regular graphs, each node is only connected to the next n nodes around the ring in a regular pattern. Regular graphs have long Lp and high Cp. (b) Data-derived graph containing 22 nodes [regions of interest (ROIs)] and 52 edges (functional connections) that seems small-world-like. Densely intraconnected clusters (black, yellow and blue) are linked through long-range short cuts (highlighted in red). Small-world networks are ’clumpy’, as reflected by high Cp and much shorter Lp than regular graphs. This type of network organization enables faster information transfer between any pair of nodes. (c) Sample random graph containing 22 nodes and 52 edges. Random graphs have moderately short Lp and low Cp. Functional connectivity diagrams of actual brain regions are neither regular nor random but more small-world like.
Here, we propose that the human brain implements top-down control in ways consistent with the complex systems principles of using multiple controllers and small-world-like architecture. We have chosen the study of top-down control as our example [8,9] because it is a complex function, probably supported by sets of interrelated brain regions that configure downstream processing in accordance with conscious goals [10,11].
Earlier studies [12,13] most commonly ascribed top-down control to several prefrontal regions, mainly the dorsolateral prefrontal cortex (dlPFC) and dorsal anterior cingulate cortex/medial superior frontal cortex (dACC/msFC). By contrast, we argue that top-down control is not implemented by such a limited number of regions but rather by a larger collection of functionally related regions. Second, based on both functional studies and recently developed ‘functional connectivity’ methods [3,4,11,14–17] (Box 1), we make the case that these regions are organized into relatively separate networks. Further, we present evidence that these separate control networks function at different timescales, making different contributions to the adaptability and stability of top-down control, respectively. Lastly, we argue that the network structure of these regions develops and embodies efficient small-world information processing.
Box 1. Complex systems methods for studying top-down control.
Our proposed dual-networks model of top-down control is built on several different imaging methods and analysis tools.
Mixed blocked/event-related fMRI design
Mixed blocked/event-related fMRI designs [14] separately code for task epochs, cues and different types of trials. Hence, mixed designs enable the separate extraction of start-cue signals related to control initiation, sustained signals related to set-maintenance, and error-related signals. Error-related activations (error > correct activation) represent an easily obtainable surrogate marker for at least a subset of feedback signals.
Cross-studies analyses
Complex systems theory predicts that top-down control is probably supported by a fairly large number of brain regions. By combining fMRI activation data across a variety of tasks using meta-analytic techniques, such as voxel-wise conjunction and fixed-effects meta-analytic images, general control regions can be identified reliably [11].
Rs-fcMRI
Recently popularized rs-fcMRI measures low frequency correlations (usually < 0.1 Hz) in the blood oxygenation level-dependent (BOLD) signal at rest. Statistically reliable low-frequency BOLD signal correlations might, at least in part, reflect a history of Hebbian coactivation. However, the exact nature of the rs-fcMRI signal and its relationship to structural connections remain to be elucidated. It is our contention, that rs-fcMRI measures provide a robust measure of the long-term functional relatedness of brain regions [16].
Graph theory
Graph theory is a branch of mathematics designed for exploring network relationships. Graphs consist of nodes (in our case ROIs) and edges (BOLD correlations between ROIs). Graph theory enables the simultaneous display of large numbers of nodes and the connections between them. Recent work has generated a variety of graph metrics, such as Cp and characteristic Lp (see text) [3,4,57,61].
Spring embedding
Spring-embedding algorithms, such as the Kamada–Kawai method, optimize graph layout in 2D by identifying the lowest-energy state of a given graph [17]. Such algorithms consider connected nodes as being attracted towards each other, proportional to their Lp, whereas nodes that are weakly connected or unconnected are pushed apart.
Graph movies
Static graph displays are limited, in that they provide a single snapshot of a network at a specific connection strength threshold. This limitation can be overcome by combining multiple graphs at different correlation thresholds into a movie (see supplementary material, Movie 1).
Control implemented by a large set of distributed brain regions
Many single-unit and functional magnetic resonance imaging (fMRI) studies, showing that the dlPFC can maintain task-relevant information during the delay between a cue and a subsequent trial, have triggered intense focus on the lateral prefrontal cortex as a top-down controller [12,18]. However, a series of recent event-related, and mixed blocked/event-related human fMRI studies have shown that a large collection of lateral and medial frontal, prefrontal and parietal brain regions also have important roles in top-down control [11,19–26] (Figure 2).
Figure 2.

Distinct fronto-parietal and cingulo-opercular control networks. (a) The network structure of human control networks is displayed in a two-dimensional graph layout. Black lines indicate strong resting state functional connections between brain regions. The thickness of the lines indicates the relative connection strength (r). A spring-embedding algorithm (Net-Draw) was used to generate the 2D graph layout [17]. This algorithm treats each connection as a spring; thus, brain regions with similar patterns of connections are brought closer together in 2D space. This method arranges the nodes of a graph in ‘connection space’ rather than anatomical space. Regions sharing connections are placed close together, whereas minimally connected regions are spatially distant. For example, the left and right IPS have similar connectivity profiles and are therefore positioned closely adjacent in the network graph. For each region (circle), the central color indicates which network it belongs to (black = cingulo-opercular; blue = cerebellar and yellow = fronto-parietal). The outer color indicates the predominant control signal type of each region (red = set-maintenance; blue = error-related and yellow = start cue-related). At the displayed correlation threshold (r ≥ 0.15), the cingulo-opercular and fronto-parietal networks are not directly connected to each other but each network is connected to the cerebellar error-network through regions that also carry error information (the thalamus, dlPFC and IPL). This architecture suggests that both networks might be communicating error signals (or codes) to and from the cerebellum, in parallel. (b) Distinct cingulo-opercular (black) and fronto-parietal (yellow) control networks, in addition to cerebellar regions (blue circles) are shown on an inflated surface rendering of the human brain [55].
Other fMRI, single-unit, event-related potentials and transcranial magnetic stimulation studies support this broader view [27–30]. For example, several investigators have shown that brain regions, including the inferior parietal lobule (IPL) [21,22,31] and anterior prefrontal cortex (aPFC) [11,24,25,32,33], maintain task and context information. In addition, fMRI studies have shown that the anterior insula/frontal operculum (aI/fO) and dACC/msFC carry a variety of control signals [11,20,23,26–28,34–36].
Posner and Petersen’s [10] classic model of attentional control proposed that different control regions carry out distinct functions. Consistent with this idea, our cross-studies analyses of mixed blocked/event-related fMRI data, which examined three different types of control signals, showed that one set of control regions carried task set-maintenance signals [aI/fO, dACC/msFC, anterior prefrontal cortex (aPFC)], whereas other regions showed predominantly feedback and adjustment (the dlPFC and IPL), or control initiation activity [the intraparietal sulcus (IPS) and dorsal frontal cortex (dFC)] [11].
Control regions separate into fronto-parietal and cingulo-opercular networks
Cataloging functional differences between brain regions alone provides limited insight into how these regions relate to one another in information processing terms. Hence, recent studies have used resting state functional connectivity MRI (rs-fcMRI) to examine functional relationships between sets of regions [16]. Two recent studies employing rs-fcMRI, one using graph theory and hierarchical clustering [8] and the other using independent component analysis (ICA) [37], have shown that the control regions of the brain separate into distinct networks. Along with the identification of these networks using ICA [37], visualization of network structure across different correlation thresholds demonstrates that the separation is independent of specific graph definition thresholds (see supplementary material, Movie 1).
Here, we refer to the two distinct control networks as the ‘fronto-parietal’ and ‘cingulo-opercular’ networks. The fronto-parietal network consists of the dlPFC, IPL, dFC, IPS, precuneus and middle cingulate cortex (mCC). The cingulo-opercular network includes the aPFC, aI/fO, dACC/msFC and thalamus (Figure 2). Our functional connectivity analyses also showed that the cerebellum forms a separate but related cluster of regions that is interposed between the fronto-parietal and cingulo-opercular networks (Figure 2a; see supplementary material, Movie 1).
Distinct functions of control networks: adaptive control and set-maintenance
Mixed blocked/event-related fMRI designs can separate brain signals based on differences in their temporal profiles [14,38]. In our mixed blocked/event-related fMRI analyses [11], the fronto-parietal network contains signals that potentially initiate and adjust control on a trial-to-trial basis, whereas the cingulo-opercular network provides stable ‘set-maintenance’ over the entire task epoch.
Regions in the fronto-parietal control network respond to cues signifying task onset. In addition, they respond differentially to events that carry performance feedback information (e.g. correct versus error trials) [11]. The fronto-parietal control network seems to include parts of the dorsal attention network proposed by Corbetta and Shulman [39], and also additional regions commonly associated with top-down control in cue-delay-trial paradigms (the dlPFC and IPL) [12,18,31]. The fronto-parietal control network, then, seems to combine brain regions that initiate attentional control in response to cues with regions that process performance feedback on a trial-by-trial basis to adjust control settings [8,9,37].
In addition to the control initiation and adjustment signals also seen in the fronto-parietal network, the cingulo-opercular network carries set-maintenance activity that extends across the performance of many trials of a task. Hence, the hallmark feature of the cingulo-opercular network is set-maintenance activity that spans the entire task epoch [11]. Consistent with our data, several other fMRI studies have also associated the aPFC, dACC/msFC and aI/fO with important task control functions [24,27,33,36,40]. Several recent studies have implicated the dACC/msFC [36,41], in addition to the aI/fO and aPFC, in decision making [40]. It stands to reason that some of the same brain regions that safely maintain task goals are also involved in making (and/or monitoring) choices in accordance with those task goals.
In our mixed blocked/event-related fMRI studies [11], a set of cerebellar regions displayed only error-related activity, consistent with the view that the cerebellum processes error information in the service of performance optimization [42,43]. Interestingly, the cerebellar error regions were connected to the dlPFC and IPL in the fronto-parietal network, and to the thalamus in the cingulo-opercular network, all regions characterized by error-related (feedback and adjustment) activity [44] (Figure 2). These results suggest that the cerebellum sends error codes to both networks and/or that it receives error information from one or both of the control networks of the brain.
Dual-networks model of top-down control
The combination of studies outlined earlier [8,11,37,45] suggests that human behavior draws on two different types of top-down control (Figure 3). The fronto-parietal and cingulo-opercular systems both seem to maintain task-relevant information, but for different purposes and using different mechanisms [11,18,23,25,32,33,46–49]. Hence, we argue that more adaptive control (fronto-parietal) and stable set-maintenance (cingulo-opercular) rely on distinct types of ‘sustained’ activity.
Figure 3.
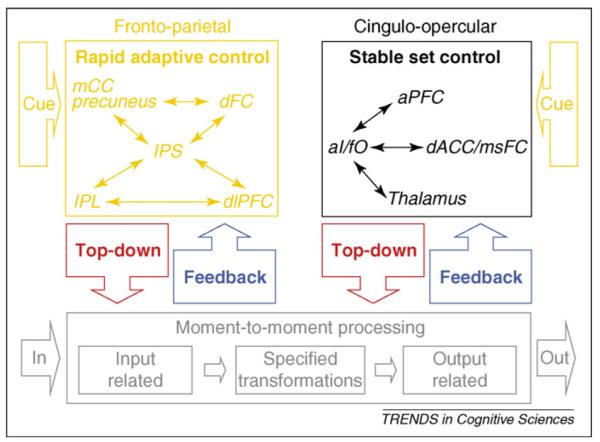
Model of proposed parallel rapid-adaptive and set-maintenance networks for human top-down control. Thin arrows schematize strong functional connections and boxed arrows schematize putative flow of information. The fronto-parietal and cingulo-opercular control networks might be organized in parallel. Both networks would thus interpret cues, implement top-down control and process bottom-up feedback. The fronto-parietal network might maintain task-relevant information in a more readily accessible form to adjust control rapidly. The cingulo-opercular network might stably maintain task sets across entire task epochs [9], perhaps in a less easily accessible, or remote, form. Adapted, with permission, from [9] Proceedings of the National Academy of Sciences. Copyright (2007) National Academy of Sciences, U.S.A.
The fronto-parietal network seems to actively maintain task-relevant information about one or a small number of trials, to implement control parameter adjustments more rapidly (Figure 3). We believe that cue-delay-target paradigms [12] detect this type of task control information as ‘sustained’ activity between a cue and the following target. The fronto-parietal network might also use error-related information to affect processing on the subsequent trial or for a limited number of subsequent trials.
Distinct from the maintenance activity seen between a cue and the following target in cue-delay-target paradigms, the cingulo-opercular network carries sustained activity that spans across the trials in a task epoch. The feedback signals received by the cingulo-opercular network, rather than causing immediate adjustments, might perhaps be integrated over many repetitions in a more protracted iterative fashion [36,50] (Figure 3).
Our dual-networks model of control (Figure 3) thus stands in contrast to several earlier models that ascribed top-down control functions primarily to the lateral PFC [18,49,51]. We argue that the lateral PFC does not constitute the lone top-down controller of the brain [18,51]. Instead, we contend that functionally and anatomically distinguishable regions of prefrontal (the dlPFC and aPFC), frontal (the dFC, dACC/msFC and aI/fO) and parietal cortex (the IPL, IPS and precuneus) contribute specific individual control functions [11] as nodes within two separate control networks [8,9,37,52]. Our proposed dual-networks model can account for the recent finding by Rossi et al. [53] that large lateral PFC lesions (apparently sparing the aPFC, dACC/msFC and, possibly, the aI/fO) severely impair the ability of a macaque to adjust behavior in response to frequent cue changes, whereas its ability to maintain set seems to be unaffected. By comparison, these results seem to be problematic for single-mechanism lateral PFC accounts of top-down control.
Furthermore, the finding that spatially adjacent prefrontal, frontal and parietal regions are separated into different clusters [9,11,37] seems inconsistent with the proposal that the PFC contains a single hierarchically organized mechanism that implements increasingly more abstract forms of control along a caudo-rostral gradient [13,49]. If the control system of the brain formed a single hierarchy, we would expect its functional connectivity to be more regular (Figure 1), such that each level was connected locally to the levels above and below.
The adaptive control and set-maintenance systems we propose are not only functionally, but also cytoarchitectonically distinct. Only the cingulo-opercular network contains von Economo neurons [54] – large layer V projection neurons found most abundantly in humans. This cytoarchitectonic dissociation suggests that the cingulo-opercular and fronto-parietal networks might have taken different evolutionary paths [54–56] (Box 2).
Box 2. Set-maintenance functions might have evolved later than adaptive control.
The presence of parallel top-down control networks in the human brain might be explained by the different evolutionary trajectories of the two networks. Evidence for this idea stems from cytoarchitectonic studies in different primate species, including humans.
Von Economo neurons are a class of layer V projection neurons fairly specific to hominids. They are increasingly more abundant in the more advanced species of apes (and some cetaceans), with highest abundance in humans.
Importantly, von Economo neurons are not uniformly distributed. The ACC and bilateral aI/fO carry particularly large numbers of von Economo neurons [54]. These data provide a cytological separation between the cingulo-opercular set-maintenance and fronto-parietal rapid-control networks.
The structural distinctiveness of the human dACC/msFC and aI/fO might be a reflection of the much greater reliance on stable set-maintenance in goal-directed behavior that is characteristic of hominoid primates, particularly humans. These data suggest that the recent evolution of the cingulo-opercular network in primates might have facilitated improved set-maintenance abilities, whereas the fronto-parietal control system took on its current role somewhat earlier.
Behavioral data also support the idea that human set-maintenance functions (assisted by the cingulo-opercular network) might be more developed. Stoet et al. [56] showed that humans are better at set-maintenance than macaques leading to less interference, but higher set-switching costs.
Please note that although we propose that control processes can be broken down into at least two basic types: rapid-adapting and set-maintenance, we do not feel that the control regions are interchangeable. We maintain that different nodes within each network carry out specialized processes that contribute to the overall functions of the networks.
Small-world control network architecture supports efficient information processing
Watts and Strogatz [2] highlighted that the connectional topology of many complex systems is neither completely regular (lattice) nor completely random. In a lattice, a given node or brain region is only locally connected to the next n nodes. Local connectivity is high and nodes are well clustered, but any signal traveling far across the network is significantly slowed because it has to cross too many nodes. One can think of a regular network as a system with only a local bus line that makes every stop (Figure 1).
By replacing some of the short-range links with long-range connections, Watts and Strogatz preserved local clustering, while also introducing between-cluster short cuts. Such a small-world network has a local bus line in addition to an express bus that directly connects different clusters without unnecessary stops.
These architectures can be described mathematically in terms of clustering coefficient (Cp) and characteristic path length (Lp). Cp describes the density of local clustering by measuring the fraction of neighbors of a given node that are also neighbors of each other, whereas Lp counts how many connections information has to traverse, on average, when traveling across the network.
Several investigators have suggested that the brain is organized in such a way that clusters of regions with similar functions segregate into networks, whereas sparse connections between networks enable the effective functional integration of processing streams [3,57–59]. Resting state connectivity studies [60], including our own [8,9], support this notion. Consistent with these observations, our graph-theoretical analyses of control networks revealed small-world organization, as indicated by relatively high Cp and small Lp [8].
Recent developmental rs-fcMRI data suggest that the structure of inter-regional networks in children is characterized by an abundance of local (short-range) connections. Over development, some local connections regress, whereas the strength of some long-range connections (short cuts) increase [9], mimicking the progression from a more lattice-like to a small-world structure [2] (Figure 1). In children aged between seven and nine years, the rs-fcMRI connectivity between the control regions of the brain correlates strongly with anatomical proximity. For example, in children, the dlPFC is more closely connected to the anatomically adjacent aPFC than to the IPL, even though the dlPFC and IPL are functionally more closely related in adults [9]. Consistent with the original description of small-world dynamics by Watts and Strogatz, the adult dual-networks control architecture emerges from a more regular organization [9] (Figure 1).
Conclusions and future directions
Evidence suggests that the principles of (i) multiple controlling variables and (ii) small-world connectivity hold true for the human brain, in particular for higher cognitive functions, such as top-down control.
In contradistinction to prior models, we argue that top-down control is driven by a fairly large collection of brain regions. These regions are distributed throughout the prefrontal, frontal and parietal cortex, in addition to the insula, cerebellum and thalamus.
Although previous models [18] treated control as a single continuously measurable entity, we believe top-down control in typical populations to be an amalgam of distinct processes, carried out by different brain regions, grouped into at least two separate control networks.
We propose that human behavior is, at the same time, flexible and highly stable because it can rely on at least two fairly parallel control networks: one optimized for rapid adaptive control (fronto-parietal) and the other for stable set-maintenance (cingulo-opercular). It stands to reason that the dual-networks control architecture is additionally advantageous because it increases the overall resilience of top-down control to damage or other perturbations. Although we have identified two control networks, it seems likely that additional controllers might exist, operating at other temporal and/or spatial scales.
In the future, our approach for studying top-down control networks should be expanded to include many more brain regions and functions. Similar to others who have advocated the elucidation of the human structural connectome [61], our final goal would be the visualization of all major functional relationships between all of the functional areas of the brain – a human functional connectome [61]. A human connectome based on both structural and functional connectivity data could then be annotated using activation studies that provide additional functional context. The development of the human connectome could be charted from birth to old age, and also examined in special populations [62], such as attention-deficit hyperactivity disorder [63], obsessive-compulsive disorder, Tourette syndrome, stroke [64] and traumatic brain injury patients, to better understand the systems-level pathophysiology of these important disorders.
Supplementary Material
Acknowledgements
We thank Francis M. Miezin and Steven M. Nelson for their suggestions and help with data analysis. We thank Marcus E. Raichle, Ronny A.T. Dosenbach, Jessica A. Church, Alecia C. Vogel and Yannic B.L. Dosenbach for helpful discussions. This work was supported by NIH grants NS41255 and NS46424 (S.E.P.), the John Merck Scholars Fund, the Burroughs-Wellcome Fund, the Dana Foundation (B.L.S.), the Ogle Family Fund (B.L.S.), a Washington University Chancellor’s Graduate Fellowship (to D.A.F.) and a United Negro College Fund/Merck Graduate Science Research Dissertation Fellowship (to D.A.F.).
Footnotes
Supplementary data Supplementary data associated with this article can be found, in the online version, at doi:10.1016/j.tics.2008. 01.001. Movie 1 can be viewed here, as well as the corresponding movie caption [65].
References
- 1.Gunderson LH, Holling CS. Panarchy: Understanding Transformations in Human and Natural Systems. Island Press; 2002. [Google Scholar]
- 2.Watts DJ, Strogatz SH. Collective dynamics of ‘small-world’ networks. Nature. 1998;393:440–442. doi: 10.1038/30918. [DOI] [PubMed] [Google Scholar]
- 3.Sporns O, et al. Organization, development and function of complex brain networks. Trends Cogn. Sci. 2004;8:418–425. doi: 10.1016/j.tics.2004.07.008. [DOI] [PubMed] [Google Scholar]
- 4.Sporns O, Zwi JD. The small world of the cerebral cortex. Neuroinformatics. 2004;2:145–162. doi: 10.1385/NI:2:2:145. [DOI] [PubMed] [Google Scholar]
- 5.Honey C, et al. Network structure of cerebral cortex shapes functional connectivity on multiple time scales. Proc. Natl. Acad. Sci. U. S. A. 2007 doi: 10.1073/pnas.0701519104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mantini D, et al. Electrophysiological signatures of resting state networks in the human brain. Proc. Natl. Acad. Sci. U. S. A. 2007;104:13170–13175. doi: 10.1073/pnas.0700668104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kearns M, et al. An experimental study of the coloring problem on human subject networks. Science. 2006;313:824–827. doi: 10.1126/science.1127207. [DOI] [PubMed] [Google Scholar]
- 8.Dosenbach NU, et al. Distinct brain networks for adaptive and stable task control in humans. Proc. Natl. Acad. Sci. U. S. A. 2007;104:11073–11078. doi: 10.1073/pnas.0704320104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Fair DA, et al. Development of distinct control networks through segregation and integration. Proc. Natl. Acad. Sci. U. S. A. 2007;104:13507–13512. doi: 10.1073/pnas.0705843104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Posner MI, Petersen SE. The attention system of the human brain. Annu. Rev. Neurosci. 1990;13:25–42. doi: 10.1146/annurev.ne.13.030190.000325. [DOI] [PubMed] [Google Scholar]
- 11.Dosenbach NU, et al. A core system for the implementation of task sets. Neuron. 2006;50:799–812. doi: 10.1016/j.neuron.2006.04.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.MacDonald AW, et al. Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control. Science. 2000;288:1835–1838. doi: 10.1126/science.288.5472.1835. [DOI] [PubMed] [Google Scholar]
- 13.Koechlin E, et al. The architecture of cognitive control in the human prefrontal cortex. Science. 2003;302:1181–1185. doi: 10.1126/science.1088545. [DOI] [PubMed] [Google Scholar]
- 14.Visscher KM, et al. Mixed blocked/event-related designs separate transient and sustained activity in fMRI. Neuroimage. 2003;19:1694–1708. doi: 10.1016/s1053-8119(03)00178-2. [DOI] [PubMed] [Google Scholar]
- 15.Fair DA, et al. A method for using blocked and event-related fMRI data to study ‘resting state’ functional connectivity. Neuroimage. 2007;35:396–405. doi: 10.1016/j.neuroimage.2006.11.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fox MD, Raichle ME. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 2007;8:700–711. doi: 10.1038/nrn2201. [DOI] [PubMed] [Google Scholar]
- 17.Kamada K, Kawai S. An algorithm for drawing general undirected graphs. Inf. Process. Lett. 1989;31:7–15. [Google Scholar]
- 18.Miller EK, Cohen JD. An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci. 2001;24:167–202. doi: 10.1146/annurev.neuro.24.1.167. [DOI] [PubMed] [Google Scholar]
- 19.Fan J, et al. The activation of attentional networks. Neuroimage. 2005;26:471–479. doi: 10.1016/j.neuroimage.2005.02.004. [DOI] [PubMed] [Google Scholar]
- 20.Cole MW, Schneider W. The cognitive control network: integrated cortical regions with dissociable functions. Neuroimage. 2007;37:343–360. doi: 10.1016/j.neuroimage.2007.03.071. [DOI] [PubMed] [Google Scholar]
- 21.Fincham JM, et al. Neural mechanisms of planning: a computational analysis using event-related fMRI. Proc. Natl. Acad. Sci. U. S. A. 2002;99:3346–3351. doi: 10.1073/pnas.052703399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fincham JM, Anderson JR. Distinct roles of the anterior cingulate and prefrontal cortex in the acquisition and performance of a cognitive skill. Proc. Natl. Acad. Sci. U. S. A. 2006 doi: 10.1073/pnas.0605493103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Crone EA, et al. Neural evidence for dissociable components of task-switching. Cereb. Cortex. 2006;16:475–486. doi: 10.1093/cercor/bhi127. [DOI] [PubMed] [Google Scholar]
- 24.Sakai K, Passingham RE. Prefrontal set activity predicts rule-specific neural processing during subsequent cognitive performance. J. Neurosci. 2006;26:1211–1218. doi: 10.1523/JNEUROSCI.3887-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Braver TS, et al. Neural mechanisms of transient and sustained cognitive control during task switching. Neuron. 2003;39:713–726. doi: 10.1016/s0896-6273(03)00466-5. [DOI] [PubMed] [Google Scholar]
- 26.Weissman DH, et al. Dorsal anterior cingulate cortex resolves conflict from distracting stimuli by boosting attention toward relevant events. Cereb. Cortex. 2005;15:229–237. doi: 10.1093/cercor/bhh125. [DOI] [PubMed] [Google Scholar]
- 27.Johnston K, et al. Top-down control-signal dynamics in anterior cingulate and prefrontal cortex neurons following task switching. Neuron. 2007;53:453–462. doi: 10.1016/j.neuron.2006.12.023. [DOI] [PubMed] [Google Scholar]
- 28.Taylor PC, et al. Subsecond changes in top down control exerted by human medial frontal cortex during conflict and action selection: a combined transcranial magnetic stimulation electroencephalography study. J. Neurosci. 2007;27:11343–11353. doi: 10.1523/JNEUROSCI.2877-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Crottaz-Herbette S, Menon V. Where and when the anterior cingulate cortex modulates attentional response: combined fMRI and ERP evidence. J. Cogn. Neurosci. 2006;18:766–780. doi: 10.1162/jocn.2006.18.5.766. [DOI] [PubMed] [Google Scholar]
- 30.Etkin A, et al. Resolving emotional conflict: a role for the rostral anterior cingulate cortex in modulating activity in the amygdala. Neuron. 2006;51:871–882. doi: 10.1016/j.neuron.2006.07.029. [DOI] [PubMed] [Google Scholar]
- 31.Liston C, et al. Anterior cingulate and posterior parietal cortices are sensitive to dissociable forms of conflict in a task-switching paradigm. Neuron. 2006;50:643–653. doi: 10.1016/j.neuron.2006.04.015. [DOI] [PubMed] [Google Scholar]
- 32.Koechlin E, et al. The role of the anterior prefrontal cortex in human cognition. Nature. 1999;399:148–151. doi: 10.1038/20178. [DOI] [PubMed] [Google Scholar]
- 33.Sakai K, Passingham RE. Prefrontal interactions reflect future task operations. Nat. Neurosci. 2003;6:75–81. doi: 10.1038/nn987. [DOI] [PubMed] [Google Scholar]
- 34.Dux PE, et al. Isolation of a central bottleneck of information processing with time-resolved FMRI. Neuron. 2006;52:1109–1120. doi: 10.1016/j.neuron.2006.11.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Forstmann BU, et al. Voluntary selection of task sets revealed by functional magnetic resonance imaging. J. Cogn. Neurosci. 2006;18:388–398. doi: 10.1162/089892906775990589. [DOI] [PubMed] [Google Scholar]
- 36.Rushworth MF, et al. Functional organization of the medial frontal cortex. Curr. Opin. Neurobiol. 2007;17:220–227. doi: 10.1016/j.conb.2007.03.001. [DOI] [PubMed] [Google Scholar]
- 37.Seeley WW, et al. Dissociable intrinsic connectivity networks for salience processing and executive control. J. Neurosci. 2007;27:2349–2356. doi: 10.1523/JNEUROSCI.5587-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Donaldson DI, et al. Dissociating state and item components of recognition memory using fMRI. Neuroimage. 2001;13:129–142. doi: 10.1006/nimg.2000.0664. [DOI] [PubMed] [Google Scholar]
- 39.Corbetta M, Shulman GL. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002;3:201–215. doi: 10.1038/nrn755. [DOI] [PubMed] [Google Scholar]
- 40.Ploran EJ, et al. Evidence accumulation and the moment of recognition: Dissociating perceptual recognition processes using fMRI. J. Neurosci. 2007;27:11912–11924. doi: 10.1523/JNEUROSCI.3522-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rushworth MF, et al. Action sets and decisions in the medial frontal cortex. Trends Cogn. Sci. 2004;8:410–417. doi: 10.1016/j.tics.2004.07.009. [DOI] [PubMed] [Google Scholar]
- 42.Fiez JA. Cerebellar contributions to cognition. Neuron. 1996;16:13–15. doi: 10.1016/s0896-6273(00)80018-5. [DOI] [PubMed] [Google Scholar]
- 43.Fiez JA, et al. Impaired nonmotor learning and error detection associated with cerebellar damage: a single-case study. Brain. 1992;115:155–178. doi: 10.1093/brain/115.1.155. [DOI] [PubMed] [Google Scholar]
- 44.Kelly RM, Strick PL. Cerebellar loops with motor cortex and prefrontal cortex of a nonhuman primate. J. Neurosci. 2003;23:8432–8444. doi: 10.1523/JNEUROSCI.23-23-08432.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Paxton JL, et al. Cognitive control, goal maintenance, and prefrontal function in healthy aging. Cereb. Cortex. 2007 doi: 10.1093/cercor/bhm135. DOI: 10.1093/cercor/bhm135 ( http://cercor.oxfordjournals.org/) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Curtis CE, D’Esposito M. Persistent activity in the prefrontal cortex during working memory. Trends Cogn. Sci. 2003;7:415–423. doi: 10.1016/s1364-6613(03)00197-9. [DOI] [PubMed] [Google Scholar]
- 47.Koechlin E, et al. Dissociating the role of the medial and lateral anterior prefrontal cortex in human planning. Proc. Natl. Acad. Sci. U. S. A. 2000;97:7651–7656. doi: 10.1073/pnas.130177397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Braver TS, Barch DM. Extracting core components of cognitive control. Trends Cogn. Sci. 2006;10:529–532. doi: 10.1016/j.tics.2006.10.006. [DOI] [PubMed] [Google Scholar]
- 49.Koechlin E, Summerfield C. An information theoretical approach to prefrontal executive function. Trends Cogn. Sci. 2007;11:229–235. doi: 10.1016/j.tics.2007.04.005. [DOI] [PubMed] [Google Scholar]
- 50.Amiez C, et al. Reward encoding in the monkey anterior cingulate cortex. Cereb. Cortex. 2006;16:1040–1055. doi: 10.1093/cercor/bhj046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Duncan J. An adaptive coding model of neural function in prefrontal cortex. Nat. Rev. Neurosci. 2001;2:820–829. doi: 10.1038/35097575. [DOI] [PubMed] [Google Scholar]
- 52.Clare Kelly AM, et al. Competition between functional brain networks mediates behavioral variability. Neuroimage. 2008;39:527–537. doi: 10.1016/j.neuroimage.2007.08.008. [DOI] [PubMed] [Google Scholar]
- 53.Rossi AF, et al. Top down attentional deficits in macaques with lesions of lateral prefrontal cortex. J. Neurosci. 2007;27:11306–11314. doi: 10.1523/JNEUROSCI.2939-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Nimchinsky EA, et al. A neuronal morphologic type unique to humans and great apes. Proc. Natl. Acad. Sci. U. S. A. 1999;96:5268–5273. doi: 10.1073/pnas.96.9.5268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Van Essen DC, Dierker DL. Surface-based and probabilistic atlases of primate cerebral cortex. Neuron. 2007;56:209–225. doi: 10.1016/j.neuron.2007.10.015. [DOI] [PubMed] [Google Scholar]
- 56.Stoet G, Snyder LH. Executive control and task-switching in monkeys. Neuropsychologia. 2003;41:1357–1364. doi: 10.1016/s0028-3932(03)00048-4. [DOI] [PubMed] [Google Scholar]
- 57.Sporns O, et al. Theoretical neuroanatomy and the connectivity of the cerebral cortex. Behav. Brain Res. 2002;135:69–74. doi: 10.1016/s0166-4328(02)00157-2. [DOI] [PubMed] [Google Scholar]
- 58.Achard S, et al. A resilient, low-frequency, small-world human brain functional network with highly connected association cortical hubs. J. Neurosci. 2006;26:63–72. doi: 10.1523/JNEUROSCI.3874-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ioannides AA. Dynamic functional connectivity. Curr. Opin. Neurobiol. 2007;17:161–170. doi: 10.1016/j.conb.2007.03.008. [DOI] [PubMed] [Google Scholar]
- 60.Bassett DS, et al. Adaptive reconfiguration of fractal small-world human brain functional networks. Proc. Natl. Acad. Sci. U. S. A. 2006 doi: 10.1073/pnas.0606005103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Sporns O, et al. The human connectome: a structural description of the human brain. PLoS Comput. Biol. 2005;1:e42. doi: 10.1371/journal.pcbi.0010042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.He BJ, et al. The role of impaired neuronal communication in neurological disorders. Curr. Opin. Neurol. 2007;20:655–660. doi: 10.1097/WCO.0b013e3282f1c720. [DOI] [PubMed] [Google Scholar]
- 63.Tian L, et al. Altered resting-state functional connectivity patterns of anterior cingulate cortex in adolescents with attention deficit hyperactivity disorder. Neurosci. Lett. 2006;400:39–43. doi: 10.1016/j.neulet.2006.02.022. [DOI] [PubMed] [Google Scholar]
- 64.He BJ, et al. Breakdown of functional connectivity in frontoparietal networks underlies behavioral deficits in spatial neglect. Neuron. 2007;53:905–918. doi: 10.1016/j.neuron.2007.02.013. [DOI] [PubMed] [Google Scholar]
- 65.Bender-deMoll S, McFarland DA. The art and science of dynamic network visualization. JoSS. 2006;7:2. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.