

Abstract
In the clinical treatment of breast cancer, anti-mitotic cytotoxic agents are one of the most commonly employed chemotherapies, owing largely to their antiproliferative effects on the growth and survival of adherent cells in studies that model primary tumor growth. Importantly, the manner in which these chemotherapeutics impact the metastatic process remains unclear. Furthermore, since dissemination of tumor cells through the systemic circulation and lymphatics necessitates periods of detached survival, it is equally important to consider how circulating tumor cells respond to such compounds. To address this question, we exposed both nontumorigenic and tumor-derived epithelial cell lines to two anti-tumor compounds, jasplakinolide and paclitaxel (Taxol), in a series of attached and detached states. We report here that jasplakinolide promoted the extension of microtubule-based projections and microtentacle protrusions in adherent and suspended cells, respectively. These protrusions were specifically enriched by upregulation of a stable post-translationally modified form of α-tubulin, and this occurred prior to, and independently of any reductions in cellular viability. Microtubule-stabilization with Taxol significantly enhanced these effects. Additionally, Taxol promoted the attachment and spreading of suspended tumor cell populations on extracellular matrix. While the antiproliferative effects of these compounds are well-recognized and clinically valuable, our findings that microfilament and microtubule binding chemotherapeutics rapidly increase the mechanisms that promote endothelial adhesion of circulating tumor cells warrant caution to avoid inadvertently enhancing metastatic potential while targeting cell division.
Keywords: chemotherapeutics, Microtentacles, circulating tumor cells, metastasis
Introduction
The majority of patients who succumb to breast cancer die from complications of metastasis, a process by which cells escape a site of primary growth and colonize distant tissue [1]. Because the metastatic process is so complex, designing therapeutic strategies to suppress metastatic spread has been difficult [2]. Without the means to specifically block the dissemination of cells from a primary tumor, current chemotherapies operate primarily on the basis of systemic cytotoxicity directed against actively dividing cells. Because the cytoskeleton is a key regulator of cellular function and viability, some of the most clinically successful chemotherapies operate by directly targeting cytoskeletal filaments [3–6]. The most prominent examples of this approach are the taxanes, docetaxel and paclitaxel, a family of microtubule (MT) stabilizing agents which disrupt mitotic spindle dynamics and promote apoptosis via activation of the spindle checkpoint during M-phase [7–9]. Actin microfilament dynamics are also critical for many cellular processes, including cytokinesis, thus there has been significant interest in actin-binding compounds as novel chemotherapies. Studies have shown that the microfilament-nucleating compound jasplakinolide (jas), disrupts filamentous actin (F-actin) architecture in cultured cells, and has antiproliferative and cytotoxic effects on a range of tumor-derived cell lines via a caspase-dependent mechanism [10]. Additionally it has been shown that jas inhibits the growth of Lewis lung carcinoma, and murine xenografts in vivo [11], and such compounds are considered by many a viable alternative to traditional MT-binding chemotherapeutics [12].
Although the taxanes and actin-binding compounds such as jas have been utilized as tools to dissect various MT and actin-dependent cell-biological processes [13–15], their application to the study of cancer has largely been limited to a description of their antiproliferative effects over periods ranging from 24–72 hours [10, 11]. The vast majority of studies to-date have examined these effects exclusively with regard to adherent cell populations. As a result, the effects of these chemotherapeutics on suspended cell populations have been largely overlooked. Since dissemination of tumor cells through the lymphatics or vasculature requires release from extracellular matrix (ECM) and indefinite periods of suspension, it is equally important to consider the cytoskeletal dynamics of circulating tumor cells (CTCs) and their contributions to survival and metastatic spread. Interestingly, taxane treatment has been shown to increase the concentration of CTCs in the bloodstream more than 1000-fold when applied in advance of surgery [16]. Additionally, taxanes can worsen progression-free survival compared to anthracycline treatment when used as a monotherapy [17], though a mechanism to explain such findings has not been delineated. Our recent work describing microtubule-based microtentacles (McTNs) in detached and circulating breast tumor cells [18, 19], provides a mechanism by which cytoskeletally-directed chemotherapies could influence CTCs and highlights the need to investigate these effects.
Current methods for detecting CTCs in metastatic patients operate at extremely low optical resolution, and would completely overlook any potential chemotherapeutic enhancements of McTNs [16, 20]. Therefore, it is important to utilize high-resolution imaging techniques to better understand how these drugs impact the behavior of detached and circulating tumor cells. By extension of this reasoning, we chose to examine how human mammary epithelial cells respond, in both adherent and suspended states, to brief periods of exposure to paclitaxel (Taxol) and jasplakinolide (jas). We report here that jas disrupted F-actin, induced the formation of microtubule-rich projections in adherent mammary epithelial cells, and promoted the formation of McTNs in suspended cell populations. This effect was enhanced significantly by MT-stabilization with Taxol. Close examination of suspended cell morphology revealed that deterioration of F-actin architecture caused a compensatory rearrangement of cytoplasmic MTs, alterations in suspended cell morphology, and the formation of McTNs. In addition to enhancing these effects, Taxol promoted the attachment and spreading of suspended tumor cell populations on ECM. Our findings demonstrate that jas and Taxol significantly impact the morphology of suspended tumor cells, while MT-stabilization with Taxol promotes adhesive behavior. Therefore, while the antiproliferative effects of these compounds are well-recognized, our findings suggest that brief exposures of circulating tumor cells to microfilament and MT-binding chemotherapeutics may actually stimulate the extravasation of disseminated tumor cells, and therefore warrant caution and further understanding of how these compounds influence detached and circulating tumor cells.
Materials & Methods
Cell Culture
MCF-10A human MECs were cultured in DMEM/F12 (Gibco, CA) supplemented with 5% horse serum (Gibco, CA), epidermal growth factor (20ng/mL), hydrocortisone (500ng/mL), and insulin (5μg/mL). SkBr3 and MDA-436 human breast tumor-derived cell lines were obtained from American Type Culture Collection (Manassas, VA), and cultured in DMEM supplemented with 10% bovine calf serum. Growth media was supplemented with penicillin/streptomycin (100μg/mL) and L-glutamine (2 mmol/L). All cells were maintained in 5% CO2 at 37ºC.
Microtentacle Scoring and live cell imaging
Cells were labeled with fluorescent wheat germ agglutinin (WGA) Alexa Fluor® 594 conjugate for 20 minutes in culture conditions (Invitrogen, CA; 1:200), rinsed with PBS and incubated with Phenol Red-free DMEM (DMEM −PR) containing 500nM jasplakinolide (jas) (Calbiochem, CA), 5μM Latrunculin-A (LA) (BioMol, Plymouth Meeting, PA), 1.17 μM Taxol (tax) (Sigma chemical, MO), combinations thereof, or 0.0015% dimethylsulfoxide (DMSO) vehicle control for 1hr in culture conditions. Drug-containing media were aspirated, cells were trypsinized and fresh DMEM − PR suspensions + drug were blindly distributed in duplicate to Costar ultra low attachment plates (Corning, NY).
Single-cell microtentacle (McTN) scores were deemed positive for suspended cells displaying motile, flexible plasma membrane protrusions of length equal to or greater than the cell radius. 100 or more cells were blindly scored in each well, with a minimum of 200 counts per trial. Propidium iodide (PI) (Sigma; 1:3000) staining was accomplished by co-incubating WGA Alexa Fluor® 594 labeled cells in a PBS dilution containing PI for 20 minutes at culture conditions. Imaging was performed on an Olympus CKX41 inverted fluorescent microscope (Melville, NY) equipped with an Olympus F-view II 12-bit CCD digital camera and Olympus MicroSuite™ Five acquisition software and band pass filters for DAPI, GFP, and Texas red.
XTT viability assay
Cells were seeded in Costar clear bottom 96 well plates (Corning, NY) and incubated with the appropriate drug in DMEM −PR for 6 hrs at culture conditions, followed by the addition of activated XTT reagent to triplicate sample wells. Cells were incubated in the presence of XTT reagent for 3–4 hrs at culture conditions; absorbance was quantitated at 450nm in a BioTek™ plate spectrophotometer. Absolute absorbance values were corrected against blank cell-containing wells, and relative values were calculated against DMSO vehicle control wells.
Immunoblotting
Cells were incubated in DMEM−PR containing 500nM jas (1,3,6 hrs) 0.05% DMSO vehicle control (6 hrs), 1μg/mL TRAIL (2 hrs), 1μg/mL Taxol (1hr), or serum supplemented DMEM (growth-6 hrs). Cells were harvested in ice-cold RIPA lysis buffer ((50mM Tris-HCl (pH 8.0), 150mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, 1mM phenylmethylsulfonyl fluoride, 1% protease inhibitor cocktail (Sigma, P2714) for 30 minutes on ice. Lysates were centrifuged (14000rpm × 10 minutes) and protein concentration was measured using a Lowry-based assay (Bio-Rad, Hercules, CA). Total protein (20μ) was separated by SDS-PAGE on 10% polyacrylamide gels and then transferred to Immuno-Blot PVDF membranes (Bio-Rad, Hercules, CA). Membranes were blocked in 5% milk in TBS with 0.1% Tween 20 for 1hr at room temperature followed by an overnight incubation at 4°C in monoclonal anti α-tubulin clone DM1A (Sigma; 1:2000), β-Actin (Sigma; 1:2000), rabbit polyclonal anti-detyrosinated tubulin (Chemicon International, CA; 1:1000), or PARP (H-250, Santa Cruz Biotechnology; 1:1000). Secondary detection was performed with IgG conjugated horseradish peroxidase (1:5000; GE Healthcare) and blots were visualized with ECL plus chemiluminescent, detection kit. Signal was quantitated with an EpiChemi3 Imaging System (UVP, Inc.; Upland, CA) Hamamatsu CCD camera (Hamamatsu Photonics Hamamatsu City, Japan).
Immunofluorescence
5×104 cells were seeded on glass coverslips and incubated in growth media overnight. Cells were then incubated in DMEM− PR supplemented with appropriate drug or DMSO for 1 hr. Cells were fixed in 3.7% Formaldehyde for 10 minutes at room temperature, permeabilized (0.25% Triton X-100/PBS, 10 minutes), and blocked for 1 hr (PBS/5% BSA/0.5% NP40). Cells were incubated overnight at 4°C with mouse monoclonal anti α-tubulin tubulin clone DM1A (Sigma; 1:1000), or rabbit polyclonal anti-detyrosinated tubulin (Chemicon International, CA; 1:1000) in PBS. Secondary detection was via incubation with anti-mouse or anti-rabbit IgG Alexa Fluor®488 secondary antibodies (Molecular Probes, CA; 1:1000), with phalloidin 555 Alexa™ (Molecular Probes; 1:200), and Hoescht 33342 nuclear dye (Sigma; 1:5000) for 1 hr at room temperature. Imaging was performed on an Olympus CKX41 inverted fluorescent microscope (Melville, NY) with an Olympus F-view II 12-bit CCD digital camera, and Olympus MicroSuite™ Five acquisition software. Specimens for adherent confocal analysis were fixed in a 0.5% glutaraldehyde solution for 10 minutes at RT, permeabilized, blocked and labeled as described above. Analysis was performed on a Nikon Eclipse TE2000-E (Nikon Instruments, Inc., Melville, NY) with a Hamamatsu ORCA-ER CCD, Improvision spinning disc confocal unit, and an xyz motorized stage. Images analysis was performed in Volocity (ImproVision® Inc., Waltham, MA). Suspended cells were prepared for confocal microscopy by simultaneous fixation and permeabilization in a microtubule stabilization buffer (80mM PIPEs pH6.8, 1mM MgCl2, 5mM EGTA, 0.5%NP40) supplemented with 0.5% NP40 and 0.3% glutaraldehyde for 10 minutes at room temperature. Cells suspensions were centrifuged onto 0.5% polyethyleneimine coated coverslips at 300rpm for 6 minutes with a slow acceleration and deceleration, rinsed, blocked with 5%BSA for 10 minutes and stained with alpha tubulin (sigma DMA1; 1:1000) for 1 hour. Image stacks were acquired using a Zeiss LSM 510 META laser scanning confocal microscope, and image analysis and 3-dimensional rendering was performed in Imaris 6.1® (Bitplane® Inc., Saint Paul, MN).
ECM-attachment assay
Coverslips were coated with 50μg/mL Poly-L-lysine (Sigma chemical, MO) for 20 minutes at room temperature, washed 3 times with PBS, and inverted on an 80 μL droplet of 0.2% gelatin (Sigma, G2500) for 10 minutes at room temperature. Gelatin matrix was crosslinked by fixation with 0.5% glutaraldehyde in PBS for 10 minutes, and reactive groups were quenched on ice for 7 minutes in 10mg/mL sodium borohydride (USB, 16940-66-2) 1.5×105 cells were plated over the gelatin-coverslips in serum-free DMEM−PR in the presence of appropriate drug, and incubated at 37ºC, and fixed at 45 minutes (early timepoint), or 220 minutes (late-timepoint) in microtubule stabilization buffer (80mM PIPEs pH6.8, 1mM MgCl2, 5mM EGTA, 0.5%NP40) supplemented with 0.3% glutaraldehyde, or 3.7% formaldehyde in PBS, respectively, for 10 minutes at room temperature. Early timepoint coverslips were quenched again with NaBH4 exactly as before. Cells were permeabilized (0.25% Triton X-100/PBS, 10 minutes), blocked for 1 hr (PBS/5% BSA/0.5% NP40), and labeled as described above for alpha tubulin, phalloidin, and DNA. Quantification of cell attachment was performed using an a cell counting algorithm (Image J, Bethesda, MD) to quantitate nuclei contained within a precise area of 1.1mm2. Phalloidin staining permitted identification of round cells, which were totaled, and divided by total cell number to yield percentage rounded, from which percentage spread could be derived. Five images were analyzed for each treatment, and an unpaired one-tailed student’s t-test was performed to assess significant changes in value relative to vehicle control.
Results
Disrupting F-actin promotes organization of microtubule-rich projections in adherent cells
Intracellular microtubule-microfilament force balancing is an important feature of mammalian cells whereby contractility of the actin cytoskeleton is counterbalanced by expansion of microtubules; thus microtubule depolymerization causes cells to shrink due to unrestricted contraction of the actin cytoskeleton [21]. Given this counterbalance, we alternatively induced F-actin disruption and MT stabilization with actin and tubulin-binding chemotherapeutics and determined their effects on cytoplasmic microtubules. To achieve this, MCF-10A mammary epithelial cells were examined by immunofluorescence to visualize effects on F-actin and α-tubulin architecture after a 1 hr exposure to jas (500nM), Taxol (1.17μM), or a combination of jas and Taxol (Fig. 1). Taxol promoted robust polymerization of α-tubulin and the formation of perinuclear MT bundles (Fig. 1E-1H). Notably, these cytoplasmic MT bundles were contained exclusively within the cell body. Conversely, exposure to 500nM jas, a concentration predetermined to be disruptive by 1 hr in MCF-10As (Fig. S1), perturbed F-actin organization and induced MT-enriched projections that extended beyond the cellular periphery (Fig. 1I-1L). Taxol-induced MT stabilization dramatically augmented this effect, significantly enriching the MT density of these cellular projections (Fig. 1M-1P). In all cases where a MT projection was observed (inset panels), the appropriate channel overlay revealed the absence of F-actin. This reinforces the notion that perturbation of cortical actin structure promotes expansive MT polymerization. Similar results were obtained for SkBr3s and MDA-436 tumor-derived cell lines, indicating that these effects are common among mammary epithelial cells of varying tumorigenicity and metastatic potential (Figs S2 and S3).
Figure 1.
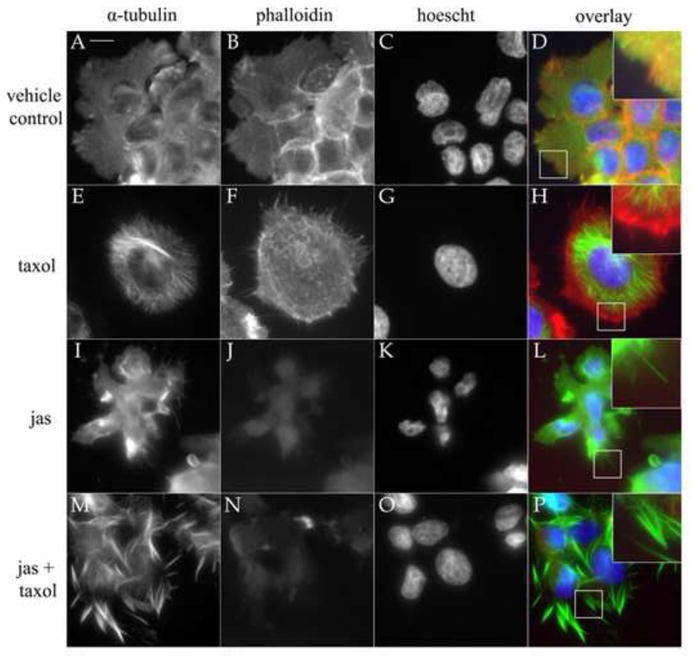
Filamentous-actin deterioration and MT-stabilization induce α-tubulin projections. To assess the effects of F-actin disruption on the presence and absence of stabilized tubulin, cells were treated with 0.15% DMSO (vehicle; A–D), 1μg/mL Taxol (tax; E–H), 500nM jasplakinolide (jas; I–L), and a combination of Taxol and jas (M-P). Cells were stained for actin with phalloidin (red), antibodies to α-tubulin (green), and Hoescht 33342 (blue) nuclear dye. Taxol reinforced microtubules and promoted the formation of MT bundles and astral bodies (E) that were contained by the microfilament cytoskeleton (F). Jas treatment collapsed F-actin (J), resulting in a loss of regular cell morphology and polymerization of thin microtubule extensions beyond cellular borders (L). Taxol augmented this effect when combined with jas (P). Inset overlay panels (2X mag) reveal the absence of F-actin in tubulin extensions (Scale bar is 10 microns).
The MT-rich extensions induced by F-actin disruption bore resemblance to McTN protrusions produced by suspended cells. To further compare their structural similarities, we examined MCF-10A cells for the presence of detyrosinated (glu)-tubulin, a specific post-translationally modified form of α-tubulin which concentrates in stable MTs [22] and is a major structural component of McTNs [19]. Figure 2 demonstrates that Taxol stimulated the formation of cytoplasmic detyrosinated microtubules (glu-MTs), in contrast to the diffuse staining pattern of the vehicle control treated MCF-10A cells. Small actin-rich microspikes are clearly visible in these cells (Fig. 2D–2F). Jas exposure induced the concentration of glu-tubulin into robust cytoplasmic filaments in a subset of cells in both SkBr3 and MDA-436 populations (Figs S4 and S5), while MCF-10As formed plasma membrane protrusions that were decorated with glu-tubulin puncta (Fig. 2G–2I). Taxol augmented this effect, increasing cytoplasmic levels of glu-tubulin and significantly enriching the concentration of glu-MTs in membrane projections (Fig. 2J–2L). This qualitative rise in glu-tubulin abundance was confirmed by western blot analysis, demonstrating that glu-tubulin levels accumulate over time in response to jas exposure, relative to vehicle control, and are greatly enhanced by Taxol (Fig. 3). These results demonstrate that these chemotherapeutic agents can induce McTN formation, as well as the extension of McTN-like protrusions in adherent cells, indicating that MT expansion is free to occur when the restraints of the cortical actin cytoskeleton are removed. This is significant in light of the fact that glu-tubulin is upregulated in invasive breast tumors [23] and because glu-tubulin rich McTNs have recently been implicated as potential determinants of metastatic success [19].
Figure 2.
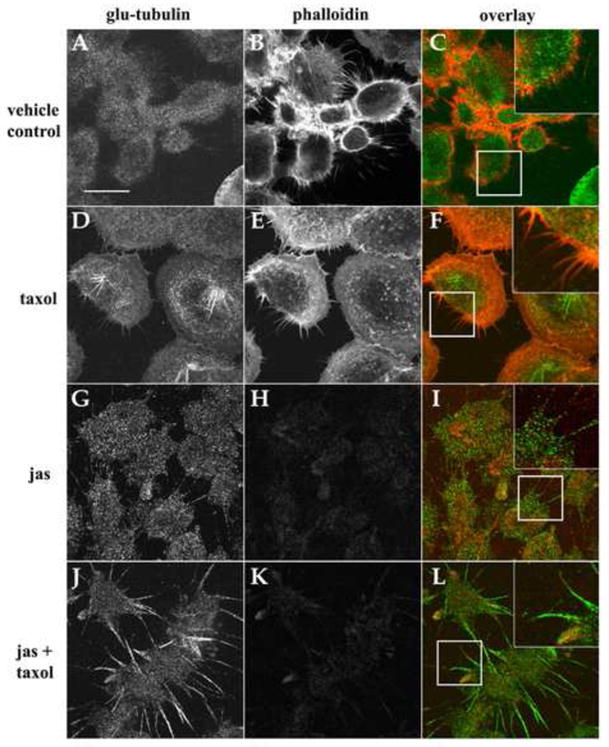
Glu-tubulin is enriched in MT-protrusions and upregulated in response to chemotherapeutic agents. Confocal microscopy of cells treated with 0.15% DMSO (vehicle; AC), 1μg/mL Taxol (tax; D–F), 500nM jasplakinolide (jas; G–I), and a combination of jas and Taxol (J–L). Relative to vehicle control cells (A) Taxol increased the abundance of glu-tubulin in all cells examined and caused de novo formation of glu-tubulin-enriched microtubules in the perinuclear region (D). Glu-tubulin was restricted to the cell body and small F-actin membrane spikes were apparent (F, inset). Jas disrupted F-actin (H) and increased glu-tubulin levels and protrusions from the cell body (G). These extensions were devoid of F-actin with particulate glu-tubulin staining (I). Taxol induced enrichment of glu-tubulin in the cellular periphery, and robust membrane protrusions (J). Overlay images confirm the absence of F-actin in protrusions (L, inset panels: 2x mag). (Scale bar is 10 microns).
Figure 3.
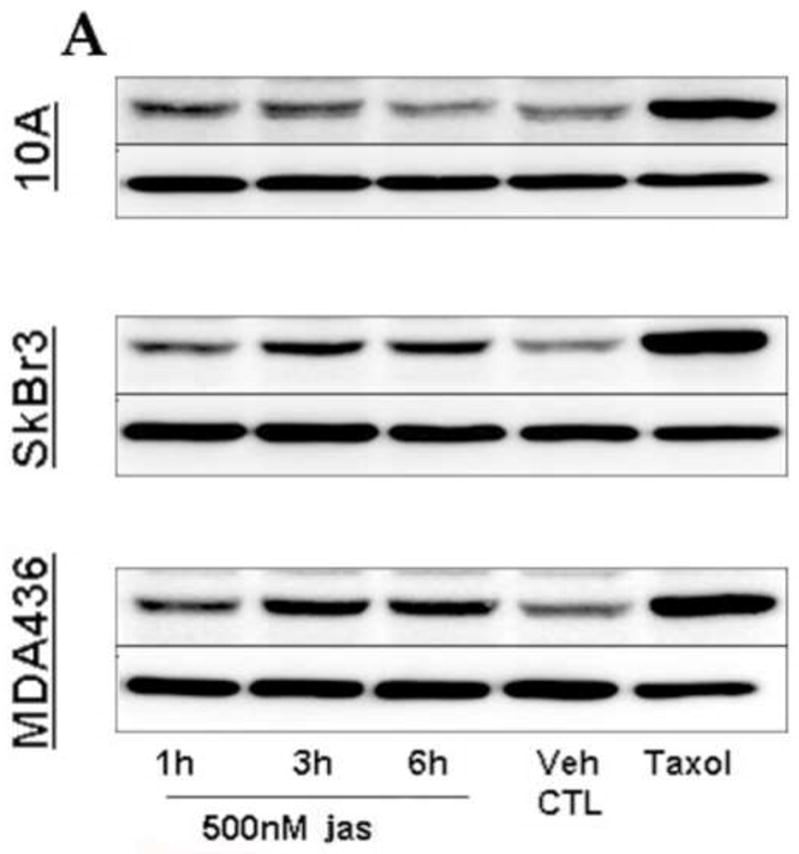
Jasplakinolide upregulates glu-tubulin. Cells were treated with jas for 1, 3, and 6 hrs, lysed, immuoblotted, and probed for glu-tubulin. MCF-10As showed an increase at out to 3 hrs, and declined to vehicle control (Veh CTL = 0.15% DMSO) levels by 6 hrs. SkBr3 and MDA-436 cells both exhibited significant accumulation of glu-tubulin by 3 hrs, and sustained elevated levels to 6 hrs. α-Tubulin levels indicate even loading of protein samples. Multiple independent lysate sets revealed a consistent trend of increased glu-tubulin (n=3, additional blots not shown).
Chemotherapeutic agents induce microtentacles independently of their cytotoxic effects
To test whether jasplakinolide and Taxol would induce the formation of McTNs in suspended cells, MCF-10A cells and two breast carcinoma tumor lines, SkBr3 and MDA-436, were treated with jas (500nM), Taxol (1.17μM) or combinations of jas and Taxol for 1 hr. Cells were then labeled with a membrane-targeted fluorescent compound, suspended, and examined for alterations in McTN formation. Blinded quantification revealed differing percentages of cells displaying McTNs in each population (Fig. 4A–4C). Latrunculin-A (LA, 5μM), an actin depolymerizing compound, served as positive control for McTN induction. In each case, jas induced significant increases in McTN formation relative to vehicle control conditions. This effect was most pronounced in the weakly invasive SkBr3 cell line, which has the lowest basal McTN frequency of all three cell lines examined, showing an increases of approximately 16-fold above control levels (Fig. 4C). All three cell lines exhibited further increases in McTN frequency upon exposure to co-treatment with jas and Taxol (P<0.05, t-test).
Figure 4.
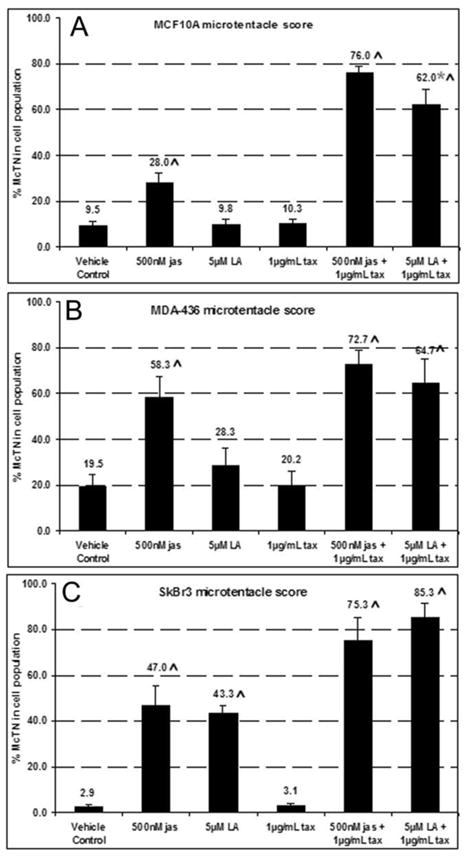
Chemotherapeutic agents enhance microtentacles in mammary epithelial cells. At least 100 live cells were blindly scored for the presence of microtentacle protrusions, in duplicate over a minimum of three independent trials (n ≥ 6). One hour treatment with 500nM jas increased McTN frequency in nontumorigenic MCF-10As (A), while LA made no significant difference. Both jas and LA induced dramatic increases in McTN score for MDA-436 (B) and SkBr3 (C) breast tumor lines. Taxol augmented these increases all cases, in combination with jas or LA (A–C). Mean values are plotted as percentage of total cells counted + standard error is represented. Significant cell clustering occurred in MCF-10As after suspension in LA + Taxol, so some trials were unable to be quantitated accurately, leading to an n=3 for this specific treatment, rather than an n=6 (*). Carats (^) indicate trials that were significantly increased over 0.15% DMSO vehicle control (Student’s t test, p < 0.01).
Taxol alone was insufficient to enhance McTN formation, but capable of acting synergistically with jas to increase McTN production. Thus it appears that F-actin disruption is necessary for an acute increase in the frequency of McTN extension, as suggested by the results obtained from the adherent-cell immunofluorescence experiments (Figs. 1 and 2). To further examine the structural changes induced by these chemotherapeutic agents in suspended cells, MCF-10A, SkBr3, and MDA-436 cells were exposed to jas, Taxol, or a combination of jas and Taxol for 1 hr, fixed in suspension and then centrifuged onto glass coverslips and analyzed with a laser scanning confocal microscope (Fig. 5). Figure 5 shows that these cells produce varying amounts of McTNs basally (Fig. 5, top row), ranging from very few protrusions per cell (SkBr3 and MCF-10A) to an abundance of tubulin-rich projections (MDA-436). Jas exposure resulted in the acquisition of more dynamic microtubule architecture and increased McTN frequency (Fig. 5, middle row). This effect was significantly enhanced for all cell lines by the addition of Taxol (Fig. 5, bottom row).
Figure 5.
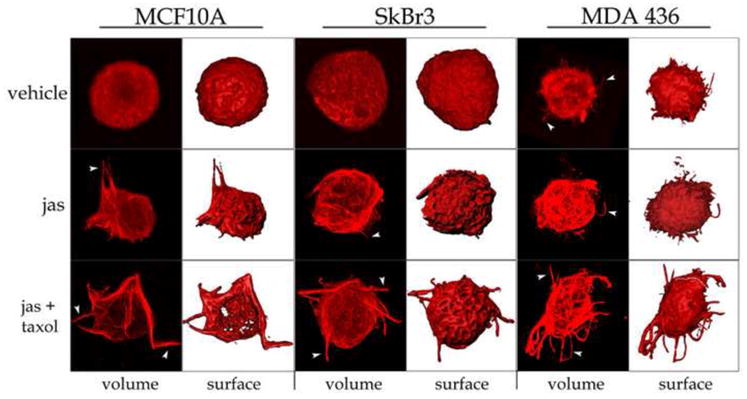
Chemotherapeutic agents stimulate MT polymerization and McTN formation. MCF-10A, SkBr3 and MDA-436 cells were treated with 0.15% DMSO vehicle control (top row), 500nM jas (middle row), or a combination of jas and Taxol (bottom row, 1μg/mL) for 1 hour. Cells were suspended for 15 minutes, fixed, and gently spun onto glass coverslips. Cells were then labeled with an antibody against α-tubulin and analyzed by confocal microscopy. Three-dimensional volumes are presented (volume) as well as surface renders to indicate changes in cellular architecture (surface). Treatment with jas or jas+Taxol increased the extension of microtubules beyond the cell periphery in microtentacle protrusions (arrowheads).
Jasplakinolide is known to inhibit proliferation of acute myeloid leukemia, carcinomas of the prostate, lung and breast in vitro, as well as murine xenografts in vivo [11]. Until now, reported experiments have primarily focused on the application of jas as an antiproliferative agent, which has been well-documented in tumor cells after treatment for longer than 24 hours [11, 24]. Conversely, our experiments utilized brief exposures (≤1hr) to study immediate cytoskeletal responses to jas. To determine if the McTNs we observed in response to jas were related to any potential cytotoxic effects of the drug within this 1 hr period, XTT cellular viability assays were performed on cells treated with a range of concentrations for 6 hrs. Figure 6A demonstrates that none of the cell lines exhibited a decrease in cellular viability within that period, at concentrations 20-fold above those used to induce McTN formation (inset panel shows no loss of viability relative to vehicle control at 10μM jas). Additionally, Figure 6B shows that cells with active McTNs excluded the nuclear dye Propidium iodide (PI). Collectively, these results indicate that McTNs are actively produced by viable breast tumor cells and therefore do not result from cytotoxic effects of jasplakinolide.
Figure 6.

Jasplakinolide does not incur cytotoxicity or apoptosis by 6 hrs. (A) MCF-10A, SkBr3, and MDA-436 populations were assayed by colorimetric detection of XTT tetrazolium reduction for alterations in viability after 6 hrs of incubation with 500nM jas. Each of the cell lines tested were viable at the experimental concentration of 500nM, remaining above a value of 1.00, relative to 0.15% DMSO control treated cells. This trend was observed for concentrations up to 10μM (inset, blue line denotes relative viability of 1.00). Bars represent mean + S.E.M for samples from three independent trials.. (B) Individual cells were tested for their ability to exclude propidium iodide (PI). Arrows specify McTN protrusions visible via fluorescent labeling with wheat germ agglutinin (WGA) (top row). PI stained nuclei of cells with compromised membrane integrity, indicated by arrowheads (bottom row). (C) Western blot analysis reveals no significant cleavage of ADP-ribose polymerase (ΔPARP) by 3 hrs in MCF-10As, and by 6 hrs of 500nM jas exposure in the tumor lines examined. TRAIL positively induced PARP cleavage (far right lane). SkBr3 cells exhibit decreased levels of β-actin at each time-point, and MCF-10As appear to upregulate β-actin over the experimental time course, while expression levels were unaltered in MDA-436s. α-tubulin levels indicate even loading of protein samples.
To specifically address the possibility that jas-induced McTN formation is linked to apoptosis, cells were assayed for cleavage of poly ADP-ribose polymerase (PARP) following jas exposure (Fig. 6C). TNF-related apoptosis inducing ligand (TRAIL) induced robust PARP cleavage (ΔPARP) by 2 hrs and served as a positive control for caspase-induced apoptosis. Minimal PARP cleavage was detectable in MCF-10As or SkBr3s at 3 hrs of jas exposure, while MDA-436s were resistant out to 6 hrs. Jas induced McTNs by 1 hr in all three cell lines (Figs. 4, 5) these data indicate that McTN induction occurs independently from apoptosis and impaired viability. Additionally, jas treatment reduced levels of β-actin in MCF-10As and SkBr3s (Fig. 6C). This trend inversely correlates with the degree of McTN induction described for each cell line, and reinforces a model where actin microfilaments can actively suppress McTN extension.
Tumor cell attachment to extracellular substrates occurs via a microtubule-dependent mechanism that is enhanced by Taxol
Within detached and circulating tumor cells, the microtubule cytoskeleton comprises the primary structural component of McTNs [18, 19], and has been shown to be required for initial adhesion to capillary walls in vivo [25]. Such findings support a two-step model of extravasation, involving an initial tubulin-dependent adhesion, followed by a secondary actin-driven penetration of the endothelial layer. Therefore we examined how Taxol exposure would impact the efficiency with which suspended tumor cells attach and settle onto an extracellular substrate. To make this determination, the highly metastatic MDA-436 cell line was selected for a series of quantitative microscopic measures of cellular attachment and spreading. 1.5×105 cells were plated over a cross-linked gelatin matrix in serum-free medium, in the presence of jas (500nM), Taxol (1.17μM), a combination of jas and Taxol, or the MT depolymerizing agent colchicine (125 μM) and incubated at 37ºC. At 45 minutes, the cells were fixed, immunostained for detection of α-tubulin and F-actin, labeled with a fluorescent nuclear dye, and imaged at low magnification. Figure 7 displays one of the 5 random fields of approximately 1mm2 that were captured for each drug treatment; an automated counting algorithm was then applied to each field in order to quantitate the total number of cells attached, and the fraction of that total that failed to spread (rounded cells). Relative to vehicle control conditions, Taxol induced rapid and efficient attachment of the suspended cells, and promoted extensive cell spreading (Fig.7E, F), relative to vehicle control (Fig. 7A, B). In contrast, jas slightly reduced the efficiency of attachment (Fig. 7C, D), while co-treatment with Taxol restored attachment to levels above that of vehicle control (Fig. 7G, H). Very few jas-treated cells were able to spread, as F-actin is required for the formation of focal contacts that permit the maintenance of a flattened, spread morphology [26, 27]. Depolymerization of MTs with colchicine significantly reduced the total number of cells that attached within 45 minutes, suggesting that MTs regulate initial stages of cell adhesion (Fig. 7I, J). Figure 8 represents the quantitative differences in total cell attachment and percentages of cell rounding and cell spreading, all of which were significant relative to vehicle control (p<0.003, t test). These results indicate that tumor cell adhesion to ECM is a two step process that depends initially on MTs for initial binding and F-actin for the formation of permanent contacts and the maintenance of a flattened morphology.
Figure 7.
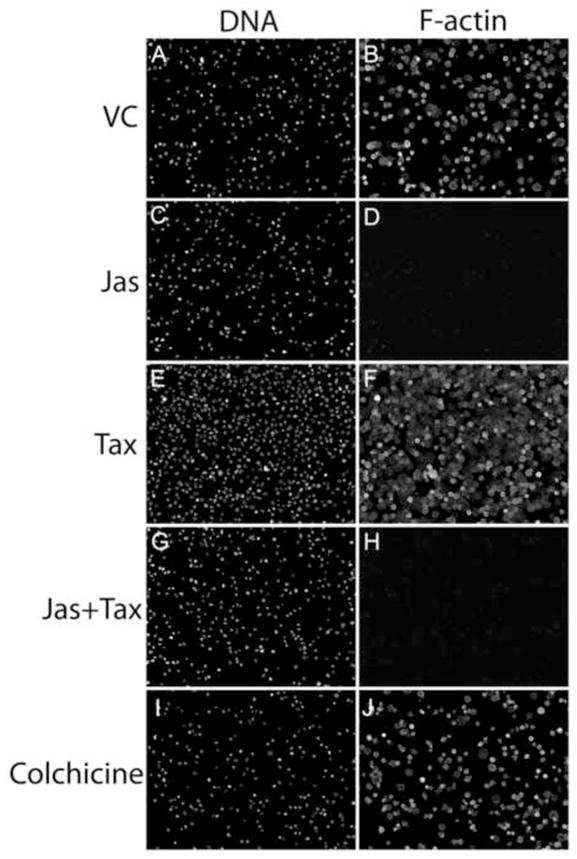
Taxol promotes tumor cell-ECM binding. Equivalent concentrations of MDA-436 cells were plated over gelatin-coated coverslips in serum-free media supplemented with 500nMjas (C and D), 1.17μMTaxol (E and F), jas + Taxol (G and H) or 125μM colchicine (I and J). After 45 minutes, unattached cells were removed by gentle aspiration and washing, and attached cells were fixed and labeled with a nuclear dye (A, C, E, G, I) and a fluorescent phalloidin conjugate (B, D, F, H, J). Images were captured at 10X magnification, revealing differences in total numbers of attached cells.
Figure 8.
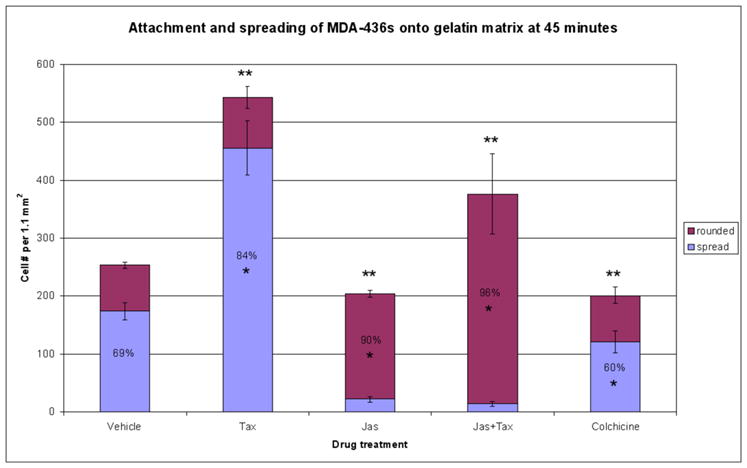
Taxol significantly enhances tumor cell adhesion and spreading. Five random fields captured at 10X magnification were collected for each drug treatment. Cell nuclei were counted to quantify the number of cells attached within an area of 1.1mm2. Within this area, the fraction of attached cells that remained rounded was also determined via a phalloidin stain, yielding percentages of cell spreading in the presence of each drug. Total cell attachment was inhibited by both jas and colchicine. Taxol was sufficient to significantly enhance adhesion, and restored this effect in jas-treated cells (** denotes significant change in total cell adhesion relative to vehicle control, p<0.001, n=5). Taxol alone increased the percentage of cells that assumed a spread morphology by 45 minutes, while treatments including jas or colchicine impeded cell spreading (* denotes significant percentage change relative to vehicle control, p<0.003, n=5). Percentage labels indicate majority values for each category (i.e. over 50% rounded or spread).
To assess whether the enhanced attachment and spreading of Taxol-treated cells would persist for periods longer than 45 minutes, MDA-436 cells were re-plated over cross-linked gelatin matrices and incubated in the absence of serum and presence of appropriate drug for 220 minutes, and then fixed. High-resolution confocal microscopy revealed that, relative to cells incubated under vehicle control conditions, Taxol continued to enhance cell spreading and the formation of cell-cell contacts (Fig. 9I–9L). In contrast, jas treated cells, though capable of attaching to the substrate, were unable to spread and remained rounded. In addition, jas induced the formation of discrete cytoplasmic MT bundles and extensions that projected away from the cell body (Fig. 9E–9H). While these structures were reminiscent of McTNs in suspended cells, the resemblance was even more striking in cells co-treated with jas and Taxol (Fig. 9M–9P). Interestingly, cell morphology under vehicle control conditions was variable, as the cells were unsynchronized and often appeared to be undergoing migratory behavior (Fig. 9A–9D). Colchicine did not significantly perturb actin-driven cell spreading (Fig. 9Q–9T). These data support a model in which MT-stabilization enhances initial adhesion of circulating cells, and further fortifies that attachment by promoting efficient spreading.
Figure 9.
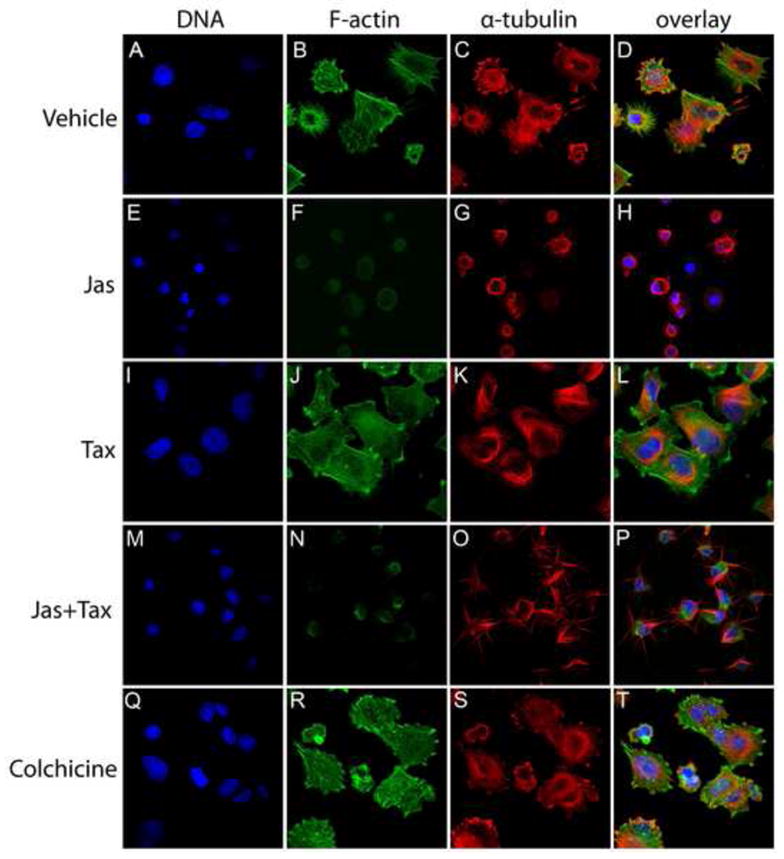
Taxol-treated tumor cells remain flattened and firmly attached beyond 3 hours. Cells were plated over a gelatin matrix in the presence of drug and allowed to settle for 220 minutes. Cells were then fixed and stained with Hoescht 33342 (DNA), phalloidin (F-actin), and an antibody to α-tubulin and analyzed with a laser scanning confocal microscope. Under vehicle control conditions, cell morphology was highly variable (A–D). F-actin was disrupted in jas-treated cells, and MT-rich projections could be seen extending from the cell body (E–H). Co-treatment with Taxol induced a specific enrichment of α-tubulin in the protrusions, relative to the cell body (M–P). Taxol alone stimulated a high percentage of the population to undergo extensive cell spreading (I–L), and some spreading was observed in the colchicine treatment, as F-actin was not perturbed in these cells (Q–T).
Discussion
Given the great clinical success of the docetaxel and paclitaxel in the treatment of breast cancer, among other cancer types [28], there has been significant interest in the development of compounds that directly modulate the stability microtubules and microfilaments [6, 8, 12]. Studies characterizing the efficacy of such compounds have focused primarily on their antiproliferative effects, and to a lesser extent on additional aspects of metastatic progression which are thought to occur in an anchorage-dependent manner, such as adherent cell motility [6–12, 14]. Such studies have focused principally on early steps of tumor progression that occur prior to release ECM release, transit through the circulation, extravasation, and colonization of secondary tissues [1], and therefore fail to address the manner in which these chemotherapies impact the behavior circulating tumor cells, or those that are not actively circulating, but disseminated and resisting apoptosis. This distinction is especially relevant in the clinical treatment of breast cancer, as tumor dormancy is a major contributor to poor patient prognosis [24, 29]. Alarmingly, surgical resection of a primary tumor increases the concentration of epithelial cells in systemic circulation by 1000-fold, and tumor cells can still be detected in the bloodstream of patients many years after entering what is clinically recognized as remission [16, 30]. Many of these patients continue to undergo regular adjuvant chemotherapy regimens [31], thus it is critically important to understand the specific mechanistic impact of such chemotherapies on circulating, and quiescent tumor cells. Adaptive taxane-resistance is a common phenomenon in the clinical treatment of many cancers [32], but this resistance is generally measured by a loss of cytotoxic potency. In addition, extended periods of dormant survival require tumor cells to be resistant to apoptotic stimuli, and are thus ineffectively targeted by cytotoxic chemotherapies, due to a combination of mitotic inactivity and apoptotic resistance [29, 33]. Circulating and dormant tumor cells may fail to die in response to MT and microfilament-binding chemotherapeutics, but the cytoskeletal behavior of these tumor cells is likely to be affected by continued exposure to these drugs.
Given this possibility, we chose to investigate the manner in which epithelial cells respond to chemotherapeutic disruption of microfilament architecture and dynamic MT instability. Exposure to nanomolar concentrations of jasplakinolide in vitro was sufficient to severely disrupt the microfilament order of MCF-10A mammary epithelial cells, and the two mammary carcinoma lines, SkBr3, and MDA-436, (Fig. S1). These concentrations were comparable to those used in studies of actin polymerization kinetics of rat fibroblasts [13] and approximately 4-fold lower than those used in studies of jas-induced apoptosis of various tumor-derived cell lines [10]. Short-term (≤ 1hr) exposures permitted us to observe the functional and structural changes in MT configuration exerted by the deterioration of F-actin, and extend these observations to suspended tumor cell populations. Our results reinforce earlier findings that the cortical actin cytoskeleton opposes extension of MT-based McTN protrusions, since Taxol-induced MT-stabilization was insufficient to enhance McTN frequency on its own. In contrast, Taxol significantly enhanced the frequency of McTNs in suspended cell populations, and the prevalence and apparent stability of adherent MT projections, when combined with microfilament disruption. A similar antagonism between MTs and microfilaments has been documented in rat and chick fibroblasts, wherein MT-depolymerization enhances contractility, and stress fiber formation [21]. Additionally, concomitant actin depolymerization and MAP-2 mediated microtubule stabilization is a natural phenomenon during neurite initiation [34], indicating that this antagonism is not restricted to epithelial cell types.
Glu-tubulin expression and localization was altered by jas treatment, and further enhanced by Taxol. Cytoplasmic Glu-MTs are more stable relative to tyrosinated-MTs [35], and high levels of tubulin detyrosination have been linked via an unknown mechanism to invasive breast tumors [23]. Additionally, increased levels of glu-tubulin are found in tumor cells of a high McTN frequency [18], thus the specific enrichment of glu-tubulin in McTNs may stabilize McTNs and thereby contribute to metastatic potential. Importantly, periods of up to 1 hr of combined exposure to jas and Taxol augmented this glu-MT enrichment, without impairing cellular viability or inducing apoptosis. Therefore, short exposures to jas and/or Taxol could ultimately promote adhesion and extravasation of suspended cells, especially since these metastatic enhancements might persist for long periods during prolonged exposure to chemotherapy-resistant CTCs. Korb et al. has provided in vivo observations of circulating HT-29 colon carcinoma cells in the hepatic capillary beds of living rats, and found that the initial adhesion of circulating colon carcinoma cells tumor cells was microtubule dependent, and enhanced by actin depolymerization [25]. As these adherent cells were unable to penetrate the basement membrane and migrate into the surrounding tissue, this finding outlines a two-step model for extravasation, wherein initial adhesion is tubulin-dependent and subsequent escape into surrounding tissue is driven by actin-mediated contractile forces. Our results support this model, and indicate that the initial phase of this adhesive transition depends upon microtentacles to facilitate endothelial or ECM contact. Therefore exposure of CTCs to MT-stabilizing agents such as Taxol could enhance this initial MT-dependent adhesion and promote extravasation into secondary sites.
In this study, Taxol served to enhance the adhesion and efficient cell spreading of MDA-436 tumor cells, relative to cells under vehicle control conditions. It appears that while cells normally transition to a flattened morphology upon attachment which is accompanied by the disappearance of MT protrusions, F-actin deterioration in suspension disrupts this transition allowing McTN-like protrusions to persist in an adherent state. Additionally, Taxol significantly enhanced the length, stability, and frequency of these protrusions in suspended cells. Furthermore, these structural changes stimulated the attachment of MDA-436s and persisted for the duration of the experiment. Though no apparent increase in McTN frequency was recorded in response to Taxol alone, there was a very significant increase in ECM adhesion, and degree of cell spreading after 45 minutes. This suggests that while Taxol is insufficient to stimulate the formation of additional McTNs, it stabilizes existing structures and therefore promotes ECM contact and adhesion via a MT-dependent mechanism. It is likely because of the fact that the highly invasive MDA-436 cell line produces high levels of McTNs basally, that this enhancement was so pronounced in these cells [19].
The cytoskeleton governs many aspects of epithelial cell function, including cell-cell and cell-ECM binding, and locomotion across extracellular substrates [3]. Deregulation of these processes contributes to the invasive potential of many carcinomas, and is usually accompanied by gross morphological changes to a migratory mesenchymal-like phenotype [36]. Specifically, structural disorganization of the actin cytoskeleton is a hallmark of malignant transformation [5, 37]. It has been shown that the degree of flexibility in the cortical actin network of a given cell line, correlates with both its metastatic potential [38, 39], and inherent McTN production [19]. Because microtubules break easily when encountering compressive force [40], it is believed that the actin cortex serves as a barrier that restricts the expansive polymerization of MTs. McTN formation, in turn, appears to be enhanced by relaxation of the cortical actin cytoskeleton, thus highly invasive tumor cells tend to produce more McTNs basally than their noninvasive counterparts [19]. Given our report that Taxol enhanced MT-dependent tumor cell adhesion, these findings highlight the importance of determining whether Taxane treatment may pose a comparatively higher risk to a subset of breast cancer patients with aggressive tumors, as the CTCs in such patients are likely to have increased flexibility in their actin cortices. In light of our recent observations that microtentacles were present in a high percentage of circulating tumor cells isolated from the blood of breast cancer patients with advanced metastatic disease (Balzer, EM and Chumsri, S unpublished data), the work described here highlights the importance of determining whether Taxane treatment can promote extravasation of disseminated tumor cells. So while stabilizing microtubules and disrupting actin filaments can each decrease primary tumor volume, it is important to consider the effects of these treatments on circulating tumor cells to avoid inadvertently enhancing the metastatic potential of disseminated tumor cells while targeting cell division.
Supplementary Material
Figure S1 - Jasplakinolide (jas) is a membrane-permeable microfilament nucleating compound that promotes microfilament polymerization and disrupts F-actin architecture. Phalloidin stain of 0.15% DMSO vehicle control (left panel) and 500nM jas (right panel) treated MCF-10A cells.
Figure S2 – F-actin deterioration in SkBr3s induces α-tubulin projections. Cells were treated with 0.15% DMSO (vehicle; A–D), 1μg/mL taxol (tax; E–H), 500nM jasplakinolide (jas; I–L), or jas + taxol (M–P). Cells were labeled with phalloidin (red), antibodies to α-tubulin (green), and Hoescht 33342 (blue) nuclear dye. Taxol promoted microtubule polymerization along the cellular periphery (E), contained within an intact microfilament cytoskeleton (H). Jas treatment collapsed F-actin (J), resulting in polymerization of thin microtubule extensions beyond cellular borders (L). Taxol augmented this effect (P). Inset overlay panels (2x mag) reveal the absence of F-actin in tubulin extensions (Scale bar is 10 microns).
Figure S3 – Filamentous-actin deterioration in MDA-436s induces α-tubulin projections. Cells were treated with 0.15% DMSO (vehicle; A–D), 1μg/mL taxol (tax; E–H), 500nM jasplakinolide (jas; I–L), and jas + taxol (M–P). Cells were labeled with phalloidin (red), antibodies to α-tubulin (green), and Hoescht 33342 (blue) nuclear dye. Taxol promoted robust polymerization along the cellular periphery (E) that was contained within an intact microfilament cytoskeleton (F). Jas treatment collapsed F-actin (J), resulting in polymerization of thin microtubule extensions beyond cellular borders (L). Taxol augmented this effect when combined with jas (P). Inset overlay panels (2x mag) reveal the absence of F-actin in tubulin extensions (Scale bar is 10 microns).
Figure S4 – SkBr3s reorganize and upregulate glu-tubulin in response to jas. Confocal analysis of cells treated with 0.15% DMSO (vehicle; A-C), 1μg/mL taxol (tax; D–F), 500nM jasplakinolide (jas; G–I), and a combination of jas+taxol (J–L). Taxol increased abundance of glu-tubulin, and induced the formation of discrete cytoplasmic filaments (D). Glu-tubulin was restricted by F-actin in cells with intact actin cytoskeletons (F, inset). Jas treated cells showed widespread loss of filamentous actin (H), in addition to robust glu-tubulin filaments within the cell body (G). Including taxol with jas caused peripheral redistribution of glu-tubulin, and membrane extensions (J). An overlay image confirms the absence of F-actin in extensions (L inset panels: 2x mag). (Scale bar is 10 microns).
Figure S5 – MDA-436s reorganize and upregulate glu-tubulin in response to jas. Confocal analysis of cells treated with 0.15% DMSO (vehicle; A–C), 1μg/mL taxol (tax; D–F), 500nM jasplakinolide (jas; G–I), and a combination of jas + tax (J–L). Taxol increased abundance of filamentous glu-tubulin along the cell periphery (D). Glu-tubulin was restricted by small F-actin membrane spikes in cells with intact actin cytoskeletons (F, inset). Jas induced widespread loss of filamentous actin (H), and the formation of cytoplasmic glu-tubulin filaments (G). Including taxol with jas caused redistribution of glu-tubulin to the cell periphery, and membrane extensions (J). Extensions were devoid of polymerized actin (L, inset panels: 2x mag). (Scale bar is 10 microns).
ABBV
- jas
jasplakinolide
- McTN
microtentacle
- tax
Taxol
- ECM
extracellular matrix
- CTC
circulating tumor cell
- F-actin
filamentous actin
- PR
Phenol Red
- Glu-tubulin
detyrosinated tubulin
Bibliography
- 1.Mehlen P, Puisieux A. Metastasis: a question of life or death. Nat Rev Cancer. 2006;6:449–58. doi: 10.1038/nrc1886. [DOI] [PubMed] [Google Scholar]
- 2.Chambers AF, Groom AC, MacDonald IC. Dissemination and growth of cancer cells in metastatic sites. Nat Rev Cancer. 2002;2:563–72. doi: 10.1038/nrc865. [DOI] [PubMed] [Google Scholar]
- 3.Calderwood DA, Shattil SJ, Ginsberg MH. Integrins and actin filaments: reciprocal regulation of cell adhesion and signaling. J Biol Chem. 2000;275:22607–10. doi: 10.1074/jbc.R900037199. [DOI] [PubMed] [Google Scholar]
- 4.Gupton SL, Gertler FB. Filopodia: the fingers that do the walking. Sci STKE. 2007:re5. doi: 10.1126/stke.4002007re5. [DOI] [PubMed] [Google Scholar]
- 5.Weber K, Lazarides E, Goldman RD, Vogel A, Pollack R. Localization and distribution of actin fibers in normal transformed and revertant cells. Cold Spring Harb Symp Quant Biol. 1975;39(Pt 1):363–9. doi: 10.1101/sqb.1974.039.01.047. [DOI] [PubMed] [Google Scholar]
- 6.Jordan MA, Wilson L. Microtubules and actin filaments: dynamic targets for cancer chemotherapy. Curr Opin Cell Biol. 1998;10:123–30. doi: 10.1016/s0955-0674(98)80095-1. [DOI] [PubMed] [Google Scholar]
- 7.Pellegrini F, Budman DR. Review: tubulin function, action of antitubulin drugs, and new drug development. Cancer Invest. 2005;23:264–73. doi: 10.1081/cnv-200055970. [DOI] [PubMed] [Google Scholar]
- 8.Jordan MA, Wilson L. Microtubules as a target for anticancer drugs. Nat Rev Cancer. 2004;4:253–65. doi: 10.1038/nrc1317. [DOI] [PubMed] [Google Scholar]
- 9.Tao W. The mitotic checkpoint in cancer therapy. Cell Cycle. 2005;4:1495–9. doi: 10.4161/cc.4.11.2130. [DOI] [PubMed] [Google Scholar]
- 10.Odaka C, Sanders ML, Crews P. Jasplakinolide induces apoptosis in various transformed cell lines by a caspase-3-like protease-dependent pathway. Clin Diagn Lab Immunol. 2000;7:947–52. doi: 10.1128/cdli.7.6.947-952.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Takeuchi H, Ara G, Sausville EA, Teicher B. Jasplakinolide: interaction with radiation and hyperthermia in human prostate carcinoma and Lewis lung carcinoma. Cancer Chemother Pharmacol. 1998;42:491–6. doi: 10.1007/s002800050850. [DOI] [PubMed] [Google Scholar]
- 12.Rao J, Li N. Microfilament actin remodeling as a potential target for cancer drug development. Curr Cancer Drug Targets. 2004;4:345–54. doi: 10.2174/1568009043332998. [DOI] [PubMed] [Google Scholar]
- 13.Bubb MR, Spector I, Beyer BB, Fosen KM. Effects of jasplakinolide on the kinetics of actin polymerization. An explanation for certain in vivo observations. J Biol Chem. 2000;275:5163–70. doi: 10.1074/jbc.275.7.5163. [DOI] [PubMed] [Google Scholar]
- 14.Cramer LP. Role of actin-filament disassembly in lamellipodium protrusion in motile cells revealed using the drug jasplakinolide. Curr Biol. 1999;9:1095–105. doi: 10.1016/s0960-9822(99)80478-3. [DOI] [PubMed] [Google Scholar]
- 15.Schiff PB, Horwitz SB. Taxol stabilizes microtubules in mouse fibroblast cells. Proc Natl Acad Sci U S A. 1980;77:1561–5. doi: 10.1073/pnas.77.3.1561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Camara O, Rengsberger M, Egbe A, Koch A, Gajda M, Hammer U, Jorke C, Rabenstein C, Untch M, Pachmann K. The relevance of circulating epithelial tumor cells (CETC) for therapy monitoring during neoadjuvant (primary systemic) chemotherapy in breast cancer. Ann Oncol. 2007;18:1484–92. doi: 10.1093/annonc/mdm206. [DOI] [PubMed] [Google Scholar]
- 17.Piccart-Gebhart MJ, Burzykowski T, Buyse M, Sledge G, Carmichael J, Luck HJ, Mackey JR, Nabholtz JM, Paridaens R, Biganzoli L, Jassem J, Bontenbal M, Bonneterre J, Chan S, Basaran GA, Therasse P. Taxanes alone or in combination with anthracyclines as first-line therapy of patients with metastatic breast cancer. J Clin Oncol. 2008;26:1980–6. doi: 10.1200/JCO.2007.10.8399. [DOI] [PubMed] [Google Scholar]
- 18.Whipple RA, Cheung AM, Martin SS. Detyrosinated microtubule protrusions in suspended mammary epithelial cells promote reattachment. Exp Cell Res. 2007;313:1326–36. doi: 10.1016/j.yexcr.2007.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Whipple RA, Balzer EM, Cho EH, Matrone MA, Yoon JR, Martin SS. Vimentin filaments support extension of tubulin-based microtentacles in detached breast tumor cells. Cancer Res. 2008;68:5678–88. doi: 10.1158/0008-5472.CAN-07-6589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Molnar B, Ladanyi A, Tanko L, Sreter L, Tulassay Z. Circulating tumor cell clusters in the peripheral blood of colorectal cancer patients. Clin Cancer Res. 2001;7:4080–5. [PubMed] [Google Scholar]
- 21.Danowski BA. Fibroblast contractility and actin organization are stimulated by microtubule inhibitors. J Cell Sci. 1989;93 ( Pt 2):255–66. doi: 10.1242/jcs.93.2.255. [DOI] [PubMed] [Google Scholar]
- 22.Kreitzer G, Liao G, Gundersen GG. Detyrosination of tubulin regulates the interaction of intermediate filaments with microtubules in vivo via a kinesin-dependent mechanism. Mol Biol Cell. 1999;10:1105–18. doi: 10.1091/mbc.10.4.1105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mialhe A, Lafanechere L, Treilleux I, Peloux N, Dumontet C, Bremond A, Panh MH, Payan R, Wehland J, Margolis RL, Job D. Tubulin detyrosination is a frequent occurrence in breast cancers of poor prognosis. Cancer Res. 2001;61:5024–7. [PubMed] [Google Scholar]
- 24.Naumov GN, Townson JL, MacDonald IC, Wilson SM, Bramwell VH, Groom AC, Chambers AF. Ineffectiveness of doxorubicin treatment on solitary dormant mammary carcinoma cells or late-developing metastases. Breast Cancer Res Treat. 2003;82:199–206. doi: 10.1023/B:BREA.0000004377.12288.3c. [DOI] [PubMed] [Google Scholar]
- 25.Korb T, Schluter K, Enns A, Spiegel HU, Senninger N, Nicolson GL, Haier J. Integrity of actin fibers and microtubules influences metastatic tumor cell adhesion. Exp Cell Res. 2004;299:236–47. doi: 10.1016/j.yexcr.2004.06.001. [DOI] [PubMed] [Google Scholar]
- 26.Seipel K, O’Brien SP, Iannotti E, Medley QG, Streuli M. Tara, a novel F-actin binding protein, associates with the Trio guanine nucleotide exchange factor and regulates actin cytoskeletal organization. J Cell Sci. 2001;114:389–99. doi: 10.1242/jcs.114.2.389. [DOI] [PubMed] [Google Scholar]
- 27.Woodring PJ, Litwack ED, O’Leary DD, Lucero GR, Wang JY, Hunter T. Modulation of the F-actin cytoskeleton by c-Abl tyrosine kinase in cell spreading and neurite extension. J Cell Biol. 2002;156:879–92. doi: 10.1083/jcb.200110014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Crown J, O’Leary M, Ooi WS. Docetaxel and paclitaxel in the treatment of breast cancer: a review of clinical experience. Oncologist. 2004;9(Suppl 2):24–32. doi: 10.1634/theoncologist.9-suppl_2-24. [DOI] [PubMed] [Google Scholar]
- 29.Brackstone M, Townson JL, Chambers AF. Tumour dormancy in breast cancer: an update. Breast Cancer Res. 2007;9:208. doi: 10.1186/bcr1677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pachmann K. Longtime recirculating tumor cells in breast cancer patients. Clin Cancer Res. 2005;11:5657. doi: 10.1158/1078-0432.CCR-05-0191. author reply 5657–8. [DOI] [PubMed] [Google Scholar]
- 31.Gralow J, Rugo H, Gradishar W, O’Shaughnessy JA, Jahanzeb M, Perez E, Tripathy D. Novel taxane formulations in the treatment of breast cancer: a thought leader discussion and consensus roundtable. Clin Breast Cancer. 2008;8:33–7. doi: 10.3816/CBC.2008.n.046. [DOI] [PubMed] [Google Scholar]
- 32.Montgomery RB, Guzman J, O’Rourke DM, Stahl WL. Expression of oncogenic epidermal growth factor receptor family kinases induces paclitaxel resistance and alters beta-tubulin isotype expression. J Biol Chem. 2000;275:17358–63. doi: 10.1074/jbc.M000966200. [DOI] [PubMed] [Google Scholar]
- 33.Martin SS, Ridgeway AG, Pinkas J, Lu Y, Reginato MJ, Koh EY, Michelman M, Daley GQ, Brugge JS, Leder P. A cytoskeleton-based functional genetic screen identifies Bcl-xL as an enhancer of metastasis, but not primary tumor growth. Oncogene. 2004;23:4641–5. doi: 10.1038/sj.onc.1207595. [DOI] [PubMed] [Google Scholar]
- 34.Dehmelt L, Smart FM, Ozer RS, Halpain S. The role of microtubule-associated protein 2c in the reorganization of microtubules and lamellipodia during neurite initiation. J Neurosci. 2003;23:9479–90. doi: 10.1523/JNEUROSCI.23-29-09479.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Infante AS, Stein MS, Zhai Y, Borisy GG, Gundersen GG. Detyrosinated (Glu) microtubules are stabilized by an ATP-sensitive plus-end cap. J Cell Sci. 2000;113 ( Pt 22):3907–19. doi: 10.1242/jcs.113.22.3907. [DOI] [PubMed] [Google Scholar]
- 36.Garber K. Epithelial-to-mesenchymal transition is important to metastasis, but questions remain. J Natl Cancer Inst. 2008;100:232–3. 239. doi: 10.1093/jnci/djn032. [DOI] [PubMed] [Google Scholar]
- 37.Friedman E, Verderame M, Lipkin M, Pollack R. Altered actin cytoskeletal patterns in two premalignant stages in human colon carcinoma development. Cancer Res. 1985;45:3236–42. [PubMed] [Google Scholar]
- 38.Guck J, Schinkinger S, Lincoln B, Wottawah F, Ebert S, Romeyke M, Lenz D, Erickson HM, Ananthakrishnan R, Mitchell D, Kas J, Ulvick S, Bilby C. Optical deformability as an inherent cell marker for testing malignant transformation and metastatic competence. Biophys J. 2005;88:3689–98. doi: 10.1529/biophysj.104.045476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Remmerbach TW, Wottawah F, Dietrich J, Lincoln B, Wittekind C, Guck J. Oral cancer diagnosis by mechanical phenotyping. Cancer Res. 2009;69:1728–32. doi: 10.1158/0008-5472.CAN-08-4073. [DOI] [PubMed] [Google Scholar]
- 40.Janmey PA. Mechanical properties of cytoskeletal polymers. Curr Opin Cell Biol. 1991;3:4–11. doi: 10.1016/0955-0674(91)90159-v. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1 - Jasplakinolide (jas) is a membrane-permeable microfilament nucleating compound that promotes microfilament polymerization and disrupts F-actin architecture. Phalloidin stain of 0.15% DMSO vehicle control (left panel) and 500nM jas (right panel) treated MCF-10A cells.
Figure S2 – F-actin deterioration in SkBr3s induces α-tubulin projections. Cells were treated with 0.15% DMSO (vehicle; A–D), 1μg/mL taxol (tax; E–H), 500nM jasplakinolide (jas; I–L), or jas + taxol (M–P). Cells were labeled with phalloidin (red), antibodies to α-tubulin (green), and Hoescht 33342 (blue) nuclear dye. Taxol promoted microtubule polymerization along the cellular periphery (E), contained within an intact microfilament cytoskeleton (H). Jas treatment collapsed F-actin (J), resulting in polymerization of thin microtubule extensions beyond cellular borders (L). Taxol augmented this effect (P). Inset overlay panels (2x mag) reveal the absence of F-actin in tubulin extensions (Scale bar is 10 microns).
Figure S3 – Filamentous-actin deterioration in MDA-436s induces α-tubulin projections. Cells were treated with 0.15% DMSO (vehicle; A–D), 1μg/mL taxol (tax; E–H), 500nM jasplakinolide (jas; I–L), and jas + taxol (M–P). Cells were labeled with phalloidin (red), antibodies to α-tubulin (green), and Hoescht 33342 (blue) nuclear dye. Taxol promoted robust polymerization along the cellular periphery (E) that was contained within an intact microfilament cytoskeleton (F). Jas treatment collapsed F-actin (J), resulting in polymerization of thin microtubule extensions beyond cellular borders (L). Taxol augmented this effect when combined with jas (P). Inset overlay panels (2x mag) reveal the absence of F-actin in tubulin extensions (Scale bar is 10 microns).
Figure S4 – SkBr3s reorganize and upregulate glu-tubulin in response to jas. Confocal analysis of cells treated with 0.15% DMSO (vehicle; A-C), 1μg/mL taxol (tax; D–F), 500nM jasplakinolide (jas; G–I), and a combination of jas+taxol (J–L). Taxol increased abundance of glu-tubulin, and induced the formation of discrete cytoplasmic filaments (D). Glu-tubulin was restricted by F-actin in cells with intact actin cytoskeletons (F, inset). Jas treated cells showed widespread loss of filamentous actin (H), in addition to robust glu-tubulin filaments within the cell body (G). Including taxol with jas caused peripheral redistribution of glu-tubulin, and membrane extensions (J). An overlay image confirms the absence of F-actin in extensions (L inset panels: 2x mag). (Scale bar is 10 microns).
Figure S5 – MDA-436s reorganize and upregulate glu-tubulin in response to jas. Confocal analysis of cells treated with 0.15% DMSO (vehicle; A–C), 1μg/mL taxol (tax; D–F), 500nM jasplakinolide (jas; G–I), and a combination of jas + tax (J–L). Taxol increased abundance of filamentous glu-tubulin along the cell periphery (D). Glu-tubulin was restricted by small F-actin membrane spikes in cells with intact actin cytoskeletons (F, inset). Jas induced widespread loss of filamentous actin (H), and the formation of cytoplasmic glu-tubulin filaments (G). Including taxol with jas caused redistribution of glu-tubulin to the cell periphery, and membrane extensions (J). Extensions were devoid of polymerized actin (L, inset panels: 2x mag). (Scale bar is 10 microns).