

Abstract
Classic mirror self-recognition mark tests involve familiarizing the subject with its mirror image, surreptitiously applying a mark on the subject’s eyebrow, nose, or ear, and measuring self-directed behaviors towards the mark. For many non-human primate species, however, direct gaze at the face constitutes an aggressive and threatening signal. It is therefore possible that monkeys fail the mark test because they do not closely inspect their faces in a mirror and hence they have no expectations about their physical appearance. In the current study we prevented 2 pig-tailed macaques (Macaca nemestrina) from seeing their own faces in a mirror and we adopted a modified version of the classic mark test in which monkeys were marked on the chest, a body region to which they normally have direct visual access but that in the current study was visible only via a mirror. Neither monkey tried to touch the mark on its chest, possibly due to a failure to understand the mirror as a reflective surface. To further the monkeys’ understanding of the mirror image, we trained them to reach for food using the mirror as the only source of information. After both monkeys had learned mirror-mediated reaching, we replicated the mark test. In this latter phase of the study, only 1 monkey scratched the red dye on the chest once. The results are consistent with other findings suggesting that monkeys are not capable of passing a mark test, and imply that face and body recognition rely on the same cognitive abilities.
Keywords: Mirror self-recognition, mark test, pigtail macaque, Macaca nemestrina
INTRODUCTION
The capacity of mirror self-recognition (MSR) is usually inferred when individuals pass the so-called mark test (Gallup 1970). In this test, subjects are first familiarized with their own mirror image. A mark is then surreptitiously placed on the subject’s eyebrow, nose, and/or ear. Subjects pass the test if they attempt to contact the mark after seeing themselves in the mirror. MSR has been studied in humans (e.g., Amsterdam 1972; Field and Adamiak 1990; Lewis and Brooks-Gunn 1979; see Parker Mitchell and Boccia 1994 for a review) by observing the reaction of young infants and toddlers to a mirror after having applied a red dye to their nose. The results show that, before the age of 18 months, few infants can use the reflected image to investigate the mark on the nose, and the majority does so only by two years of age. MSR has also been studied in non-human primates, showing that many chimpanzees (Pan troglodytes) and orangutans (Pongo pygmaeus) pass the mark test (Gallup 1970; de Veer et al. 2003; Suarez and Gallup 1981; Swartz et al. 1999) whereas many monkey species such as Macaca arctoides, Macaca mulatta, Macaca fascicularis (Gallup, 1970), and tufted capuchin monkeys (Cebus apella, Roma et al. 2007) do not. In contrast, as frequently described in these studies, monkeys display various social responses elicited by their mirror image. It seems, therefore, that monkeys lack the capacity of MSR.
Some studies, however, have hinted that monkeys do not simply treat their mirror image as if it were a stranger. Three studies, in particular, have suggested that even monkeys could have a certain capacity to distinguish their mirror image from “real” conspecifics. Eglash and Snowdon (1983) discovered that Pygmy marmosets (Cebuella pygmaea) display a rapid extinction of social threat and novelty responses to their own mirror image. In addition, monkeys subsequently used a mirror to locate otherwise unseen conspecifics and directed threat responses toward the real location of these animals rather than to their mirror image. De Waal and collaborators (de Waal et al. 2005) tested capuchins monkeys (Cebus apella) in three conditions where each subject was exposed to a familiar same sex partner, an unfamiliar same sex partner, and a mirror. Female capuchins showed more positive behaviors such as lip smacking, direct eye contact, swaying, and “bunny sitting” when placed in front of the mirror in comparison to an unfamiliar conspecific. Males, on the other hand, showed signs of distress or submission (e.g., squealing) and signs of high anxiety (e.g., glancing or curling up) almost exclusively towards the mirror. Reactions of both males and females suggest that capuchins distinguish between conspecifics and their own mirror images. Finally, Itakura (2001) reported that when confronted with an unfamiliar conspecific, the heart rate of a Japanese macaque (Macaca fuscata) first increased and then decreased whereas when exposed to a mirror, the heart rate decreased immediately. Taken together, these findings support the proposal by de Waal et al. (2005) that monkeys could have reached a level of self-other distinction intermediate between seeing their mirror image as other and recognizing it as self. If this hypothesis is correct, it might also explain why some studies have reported ambiguous occurrences of mark touching in monkeys. For example, Thompson and Boatright-Horowitz (1994) reinforced three adult pigtail macaques (Macaca nemestrina) for looking at their own mirror images, showing that in a subsequent mark test one monkey “touched her left knuckles to the small red mark on her brow, then in a continuous movement, rubbed the back of her left hand against the marked brow, inspected the back of her left hand, jaw-thrusted, and turned rapidly away from the mirror” (pp. 345). Another apparently positive result was reported by Boccia (1994), who tested 15 pigtail macaques with a lifetime of mirror exposure (up to 58,000 hours). In this study, one subject supposedly passed the test: “she looked in the mirror, swiped her hand across her face in the vicinity of the mark, and then looked her hand. The monkey performed this behavior 4 times in the test period with the mirror, examining her hand after each swipe” (pp. 354). Finally, Itakura (1987) reported mirror self recognition in one Japanese macaque (Macaca fuscata) who touched a mark placed under its nose three times in the presence of a mirror. While many remain unconvinced that the described behaviors constitute strong evidence of MSR, these results nonetheless highlight that monkeys can react to mirrors in ways that would not be expected if they were mistaking the mirror for a conspecific, and that monkeys might have a more sophisticated understanding of mirrors and mirror reflections than previously thought.
In order to understand why some primate species clearly pass and others appear to consistently fail the mark test, it may be helpful to more carefully analyze what abilities are involved in MSR. Mirrors reflect objects accurately and faithfully, which means that movements of a mirror image are contingent (albeit reversed) to one’s own movements. Hence, the first step to recognizing oneself appears to be an awareness of a kinesthetic-visual match between one’s own movements and the related somatosensory feedback of these movements (Mitchell 1993). In order to understand that the mirror image is not simply an object moving in synchrony with oneself, an individual also has to hold expectations about its own physical appearance, and then find a match between those expectations and the appearance of the mirror image. A mark test simply exploits a subject’s motivation to investigate a perceived deviation of one’s normal physical appearance.
Examples from the human literature illustrate that infants show evidence of kinesthetic-visual matching and feature matching even before they pass conventional mark tests. Meltzoff (1990, 1996) found that 14-month old infants recognize when they are being imitated, an achievement thought to be based on the match between one’s own movement (kinesthetic information) and the movements of the imitator (visual information). Bahrick et al (1996) presented infants with moving and still images of themselves and a peer, and found that infants as young as 5 months old appeared to prefer looking at the peer, suggesting that they recognized their own features. In a more recent study, Nielsen and collaborators (Nielsen et al. 2006) placed a bright sparkling sticker surreptitiously, and not directly visible, on the leg of 18–24 month old infants, and compared their attempts to retrieve the sticker (after having seen themselves in a mirror) with their performance on a conventional mark test (a sticker placed on the head). Toddlers showed similar performance on the leg recognition and face recognition test, but when the appearance of the toddlers’ legs was changed without the toddlers’ knowledge by surreptitiously slipping their legs into a new pair of loose-fitting sweatpants immediately before the test, performance dropped dramatically. These results suggest that in addition to kinesthetic-visual matching abilities, expectation about one’s visual appearance is crucial for successfully passing the mark test.
How developed are these two abilities in monkeys? Since the mark is traditionally placed on the subject’s head, we can see two potential problems that monkeys (but not human toddlers) might face. Paukner and collaborators discovered that pigtail macaques (Paukner et al. 2005) and capuchin monkeys (Paukner et al. 2009) are able to distinguish between an imitator and a non-imitator of their own actions, quite possibly on the basis of kinesthetic-visual matching. However, for many monkey species, direct gaze at the head or face is perceived as a threat. It therefore possible that monkeys avoid looking directly at their own faces when exposed to a mirror, preventing them from comparing somatosensory feedback and the mirror image’s movements, which may then result in a failure to establish kinesthetic-visual matching. Moreover, failure to attend to their own faces might also prevent monkeys from forming expectations about their appearance since their faces are never directly visible to them without a mirror. Monkeys that use mirrors to locate hidden food items (e.g. Anderson 1986; Itakura 1987) or see conspecifics through a mirror and direct threats at their real location (Eglash and Snowdon, 1983) may do so on the basis of having been able to visually match the mirror image to the real life objects or conspecifics. Taken together, the mark’s traditional location on the head, chosen as one of the few areas of the body that can only be seen through a mirror, may be particularly problematic to monkeys, and may prevent them from recognizing themselves in a mirror.
To overcome this problem, in the present series of experiments we attempted two novel modifications to the classic mark test. First, to avoid distraction from their own faces, we prevented monkeys from seeing their own head reflected in the mirror. This change might promote monkeys’ kinesthetic-visual matching abilities of e.g. their arm and hand movements. Other researches have prevented monkeys to see their gaze using angled mirrors (Anderson and Roeder 1989; Shillito et al 1999) but the profile of the head was still visible. Second, we placed a mark on the monkeys’ bodies at a location that under normal circumstances is directly visible to them but that they were prevented from seeing directly during the test. We hypothesized that monkeys should be familiar with the appearance of their own chests, e.g. through self-grooming of this area, and that they had formed an expectation about the appearance of their own chests. Assuming that monkeys can match the appearance of their chests from an upside-down perspective to the appearance of their chests in a mirror, we expected that they would recognize a deviation from their normal appearance when a red mark was placed in this area. The objective of the present study was to test whether monkeys would attempt to touch a mark on their chests and thereby pass a modified mark test.
EXPERIMENT 1
METHODS
Subjects
Subjects were two pig-tailed macaques (Macaca nemestrina), one male, Charlie, and one female, Patty. They were 7 and 8 years old, respectively, captive born, and housed at the Primate Section of the Department of Neuroscience, University of Parma. Both monkeys had been mother-reared until the age of 2–3 years. They were then singly housed (175 cm × 100 cm × 100 cm) with visual and auditory contact with other monkeys (Macaca nemestrina and Macaca mulatta). Monkeys were not food-deprived for the experiment and they received their daily food consisting of fresh fruits, vegetables, seeds and monkey chow after the experimental sessions. Patty had never been tested before with regards to MSR, whereas Charlie had participated in a classic mark test when he was 4 years old. In this previous test, Charlie had not attempted to touch a mark placed on his head. Before starting the test, both monkeys received training to come out of the cage using a leash, to sit in a primate chair, and to be touched and handled by the experimenters. At the time of the test, this procedure was well established for both monkeys and related to the experimental requirements of previous behavioural studies. All experimental protocols complied with the European law on the humane care and use of laboratory animals and were approved by the Veterinarian Animal Care and Use Committee of the University of Parma as well as the Italian Minister of Health.
Apparatus
All experiments took place in a room adjacent to the animal facilities, where monkeys had auditory but not visual contact with other animals. A mirror (60 cm × 50 cm) was hung on the wall approximately 70 cm distant from the subject and the monkey chair was placed approximately 45 cm above the floor level to prevent the monkeys from viewing their own faces in the mirror (see Figure 1a).
Fig 1.
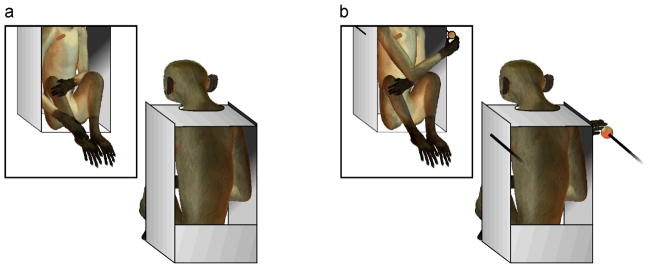
Before the mark test, monkeys were placed in front of the mirror to familiarize them with their reflected body image. Note that monkeys were prevented from seeing their own bodies directly, and that their faces and heads were not visible in the mirror. a) Red marks were applied to the monkeys’ chest (mark test 1 and 3). Subsequently, a second red mark was applied to the wrist (mark test 2 and 4). b) Experimental set-up to train monkeys to reach for food using the mirror image as a source of information.
Materials
A red carmine color Tombow ABT 845 (Tombow Pen & Pencil GmbH), acid and odor free, was used to create the mark on the body.
Procedure
Monkeys were tested at various times throughout the day depending on non-related experimental requirements. We first gave monkeys between 3 and 7 sessions of mirror exposure to allow them to familiarize themselves with the mirror and their own mirror image. At the start of an exposure session, monkeys were put in a primate chair which prevented vertical head movement. Monkeys were therefore not able to look directly down the front of their bodies (see Figure 1a). Charlie received one session per day for 7 days (2 sessions lasted 1 hour each, and 5 sessions lasted 30 min each), for a total of 4.5 hours. Patty received one session per day for 3 days (1 session lasted 1 hour each, and 2 sessions lasted 30 min each), for a total of 2 hours of mirror exposure.
After the mirror familiarization, the monkeys were marked on the chest (mark test 1). The red mark (6 cm × 2 cm) was positioned diagonally 2 cm over the right nipple. Monkeys were prevented from seeing the procedure by placing a panel under their chin and by touching their bodies with a finger and another capped color pen in several places to create a distraction. The arms were gently restrained during the procedure to permit the color to dry completely, and released after 2 min (this procedure also allowed us to avoid the use of an anesthetic, commonly used in non-human primate mark tests for safety reasons and to prevent subjects from seeing the application of the mark). Afterwards, as shown in Figure 1a, monkeys could see the red mark only by looking in the mirror. At the beginning of the test session, the mirror was covered with black fabric and monkeys were exposed to this non-reflective surface for 15 min. This procedure was carried out to ensure that the monkeys did not feel or smell the applied mark. The mirror was then revealed to both monkeys for 1 session, lasting 15 min. Following the pioneering study by Gallup and colleagues (Gallup at al. 1980), immediately after the first mark test, a second red mark (6 cm × 2 cm) was placed on one wrist (see Figure 1a). As before, the application of the wrist mark was not directly visible to the monkeys as the wrist was gently restrained and colored while touching the other wrist simultaneously. We let the color completely dry for two min before releasing the arms. The wrist mark was directly visible to the monkeys without the mirror. This second test (mark test 2) was carried out in order to confirm macaques’ interest in marks on their bodies. Both subjects received 1 test session, lasting 15 min. The red mark on the chest was thus visible for 30 min overall. At the end of the experiment, the marks were wiped off with a solution of water and alcohol. Following Povinelli and collaborators (Povinelli et al. 1997), we scored scratching, grooming and sniffing behaviors to the marked area (target) and to the contralateral, unmarked area of the body (off-target). We also scored other behaviors such as licking and fast rubbing directed to the red mark. To evaluate the monkeys’ mark touching behavior as evidence for MSR, we considered,, first, whether mark touching occurred more frequently in the presence of the mirror than in its absence; second, whether monkeys touched the mark while looking at the mirror, a behavior presumed to indicate mirror-guided touching (Gallup 1970).
All phases of the experiment were recorded with a digital camcorder (Canon MVX250i or MVX350i) positioned on the right side behind the monkey. All videos were analysed off-line.
RESULTS
Neither monkey showed any social response towards their mirror image (threat, lip smacking, pucker, fear grimace) during the initial familiarization sessions or the subsequent mark tests. Monkeys did not show any responses directed at the mark on their chests either during the first 15 min (while the monkeys were exposed to the covered mirror) or during the second 15 min (once the mirror was uncovered). After application of the wrist mark during mark test 2, Patty displayed 10 interactions with the marked wrist (target) (8 groom, 1 scratch, 1 lick) and only one with the unmarked wrist (off-target) (1 groom). Charlie displayed 5 interactions with the marked wrist (target) (1 groom, 1 sniff, 3 fast rub) but no interaction with the unmarked wrist (off-target). Both monkeys looked directly at the mark when touching their marked wrists without looking at the mirror.
DISCUSSION
During mark test 2, neither monkey attempted to touch the marks on their chests during any part of the test, although both monkeys focused on the marks on their wrists some times. Inspection of the wrist marks is in line with previous studies that placed a mark on a directly visible part of the body. For example, Boccia (1994) tested 15 pigtail macaques by placing a mark on their wrist and on their left brow and right ear, and found that 12 of them touched the mark on the wrist. Together with the present findings, these results confirm that pigtail macaques are interested, albeit modestly, in inspecting marks on their bodies when they can see them directly
Does the monkeys’ failure to touch the marks on their chests necessarily indicate that they do not recognize their own chests? Platt et al. (1991) suggested that true competence in mirror use requires three basic characteristics: a generalized concept of mirrors as a reflecting surface that provides contingencies of appearance and movement, mirror-mediated object localization and reaching, and instrumental behavior maintained by bringing the mirror or the self into appropriate position for locating and/or reaching for objects. One possibility is that our monkeys failed to show any interest in the marks on their chests because they did not sufficiently understand the mirror as a reflecting surface. To further the monkeys’ understanding of mirrors, we trained them to use the mirror in a mirror-mediated locating and reaching task.
EXPERIMENT 2
Several macaque species such as Macaca fascicularis (Anderson 1986), Macaca fuscata (Itakura 1987), Macaca tonkeana (Anderson 1986) and Macaca nemestrina (Boccia 1994) are capable of using the mirror instrumentally to locate hidden pieces of food. We trained our monkeys to reach for food using only the mirror as a source of information with the idea that a better understanding of the mirror image would allow the monkeys to obtain an improved awareness of their reflected image and, consequently, of their bodies.
METHODS
Subjects and apparatus were the same as in Experiment 1.
Materials
Two grey metal rods (30 cm long), handheld by the experimenter, were used during the training phase of the experiment.
Procedure
As in the previous experiment, monkeys were seated in a primate chair and placed in front of the mirror, which reflected the whole body but not the head. An experimenter sat approximately 40 cm behind the monkeys’ backs, with food items in place on the experimenter’s lap. We used fruits and vegetable such as oranges and fennel that overwhelmed the testing room with their fragrance, thus minimizing the possibility that monkeys used olfactory information to guide their behavior. During the baiting, both rods were withdrawn and placed close to the food, but only one of them was baited with a food item. Extending both arms, the experimenter then presented both rods simultaneously approximately 20 cm away from the sides of the primate chair, one on the left and one on the right side, for approximately 30s. Subjects could not see the food directly, but only through the reflection in the mirror (see Figure 1b). If monkeys reached towards the food item, they were allowed to take it and eat it. If they reached towards the non-baited rod, monkeys did not receive any food and the trial was ended immediately. We tested monkeys throughout the day dependent on non-related experimental requirements. Charlie received 9 training sessions, one session per day, each lasting approximately 45 min and including a different number of trials, from 80 to 149, depending on his performance. Patty received 6 training sessions, one per day, each lasting approximately 45 min and including between 26 and 132 trials. For each monkey the training finished when the percentage of correct trials remained above chance level for at least 3 consecutive sessions.
To ensure that monkeys were using visual information from the mirror, we also conducted some sessions in which we covered the mirror with black fabric and presented one baited and one unbaited rod at the side of their bodies and out of their field of vision. We scored any reaches that monkeys made towards either side of their bodies.
All phases of the experiment were recorded with a digital camcorder (Canon MVX250i or MVX350i) positioned on the right side behind the monkey. All videos were analysed off-line.
RESULTS
At the beginning of the training, both monkeys performed no better than chance (50% correct). However, both monkeys improved their performance as the training progressed (see Figure 2): both monkeys learned to reach for the correct side using only the information coming from the reflection of the mirror. When the mirror was covered, neither monkey attempted to grasp the food in any trial. Using a binomial test, we analyzed the probability of success across sessions. Charlie consistently selected the correct side significantly above chance after the seventh training session (p<0.005) while Patty consistently selected the correct side significantly above chance after the fourth training session (p<0.005). Neither monkey showed any social responses towards their mirror image.
Fig 2.

Percentage of correct reaching gestures towards the food across training sessions for Patty (grey line) and Charlie (black-dotted).
DISCUSSION
Both monkeys learned to use the mirror as source of information to guide their behavior, which indicates a capacity to recognize and grasp the offered food. Since monkeys performed no better than chance at the start of the training, it seems likely that they only had a limited understanding of the mirror. Learning to use the mirror to guide their behavior could indicate that they reached a more thorough understanding of the mirror’s reflective properties (see also Anderson 1986; Itakura 1987; Boccia 1994). If monkeys can reach for hidden food using the mirror as source of information without directly seeing the food, the association between objects in the mirror and objects in reality could extend to their own body and, in our particular case, to the marked chest. In other words, the training they received to reach for hidden food with the aid of the mirror might facilitate self-recognition.
EXPERIMENT 3
In order to test the hypothesis that monkeys failed to touch the marks on their chests during the first experiment due to an insufficient understanding of mirror properties, we repeated Experiment 1 after successful acquisition of mirror-guided reaching in Experiment 2.
METHODS
Subjects, apparatus and materials were the same as in Experiment 1.
Procedure
The procedure was the same as in Experiment 1. First, we colored part of monkey’s right chest with a red pen. Subsequently, monkeys were exposed to the mirror covered with a black fabric for 15 min. Then, the fabric was removed and we measured any self-directed response for 15 consecutive minutes of mirror exposure (mark test 3). Following mark test 3, we placed a second mark on one wrist which was directly visible to the monkeys without the mirror (mark test 4). This last test was carried out to ensure that monkeys remained interested in the marks on their bodies. As in previous experiments, monkeys were prevented from seeing their own face in the mirror reflection. All phases of the experiment were recorded with a digital camcorder (Canon MVX250i or MVX350i) positioned on the right side behind the monkey. All videos were analysed off-line.
RESULTS
During mark test 3, neither monkey showed any interactions with the red mark placed on its chest. During mark test 4, Patty interacted 8 times with the mark on her marked wrist (target) (3 groom, 2 scratch, 3 fast rub) and once with its contralateral part (off-target) (1 scratch). Interestingly, during this phase Patty also showed some behaviors directed at the red mark on her chest, although at a low frequency. In particular, at 8 minutes into the test, she scratched with the left hand the zone 1 cm under the mark on her chest and then, immediately after, she scratched exactly the red dye with the same hand. After these interactions, she also scratched the left and right sides of her body under the nipples, that is, outside both the target and the off-target areas, using both hands simultaneously. She then scratched again the body area located 1 cm under the red mark, one time, using the right hand. She never looked at her hands after these interactions, and she never touched exactly the chest region contralateral to the marked one (off-target). Due to the angle of the video recording, it is possible to ascertain that, when she touched the mark on the chest and for the duration of the interactions described, Patty was looking at the mirror. However, it is not possible to say with certainty whether she looked at the mark or elsewhere at her body. In contrast, before and after the described interactions, her gaze was not directed towards the mirror; she was in fact looking around the room, gazing left, right and above her head.
During mark test 4 Charlie showed 1 interaction with the red mark on the wrist (1 sniff) but no interactions with the mark on his chest. During all wrist interactions, monkeys’ gaze was fixed directly on the wrist marks, not via the mirror. Neither monkey showed any social response towards the mirror image.
DISCUSSION
After successful acquisition of mirror-guided reaching during Experiment 2, one monkey (Patty) touched the mark on her chest during mark test 4 in Experiment 3. Contact with the mark was fleeting, and the monkey did not inspect her hand after having touched the mark. Although her gaze was directed at the mirror, it is difficult to say if it was directly at the mark. There is therefore no strong evidence that this monkey recognized her chest in the mirror, and contact with the mark can be accounted for by chance. The other monkey, Charlie, failed to touch his chest mark but also showed overall low levels of interactions with the wrist mark, suggesting that the motivation of this monkey to examine marks on his body was low. Taken together, these results indicate that despite an understanding of mirrors as a reflective surface, monkeys nonetheless failed to react to marks on their bodies that are only visible via a mirror.
GENERAL DISCUSSION
In this study we assessed mirror self-recognition in monkeys by using a modified version of the mark test (Gallup 1970) in which we tried to overcome possible limits of the classic procedure that could prevent macaques from passing this test. Mirror self-recognition can be thought to rely on kinesthetic-visual matching abilities as well as being able to hold expectations about one’s appearance, matching these expectations to the mirror image, and reacting to violations of these expectations. We hypothesized that monkeys’ tendency to avoid looking at other monkeys’ faces directly (because direct gaze constitutes an aggressive and threatening signal) may prevent monkeys from establishing visual-kinesthetic matching with their mirror image. More importantly, lack of visual familiarity with the marked body part, typically the forehead and/or ear, may result in a lack of expectations of what this particular part of their bodies looks like. We thought to overcome these limitations by removing reflections of the monkeys’ heads as well as placing a mark on a part of their bodies with which they are familiar even outside of the mirror context.
In the first experiment (mark test 1), subjects did not react to the red mark placed on their chests. Since this failure could be due to a lack of understanding the reflective propriety of the mirror, we trained our monkeys to locate and grasp food using only the mirror as source of information (Experiment 2), to increase their comprehension of the reflective proprieties of the mirror. After the training, when a red mark was placed again on the chest (Experiment 3), one monkey (Patty) fleetingly touched the mark on her chest while looking towards the mirror. It is clear, however, that these data are quantitatively and qualitatively different from the descriptions of successful mark tests with great apes. Apes typically gaze carefully into the mirror while tactilely exploring the mark. At best, a few macaques have been reported to make a passing swipe at the mark and then to explore their hand (e.g. Boccia, 1994; Itakura, 1987; Thompson and Boatright-Horowitz, 1994). The monkey’s contact with the mark in the present study may be due to chance, although it is interesting to note that this monkey was also faster then the other monkey to learn using the mirror as a source of information to locate hidden food items.
The results of the present study are in line with previous results of mark tests in pigtail macaques, that is, despite removal of the reflection of their heads and placement of the mark on a body part that they should be familiar with, monkeys did not convincingly attempt to investigate the mark. Previous suggested explanations of these results, such as monkeys avoid directly looking at their own faces (Hauser et al., 1995), do not apply to this case. Interestingly, we never observed any social response directed at the mirror image by our monkeys, either during the familiarization phase or during the mark tests. Such responses are commonly reported when monkeys are first exposed to a mirror (e.g. Eglash and Snowdon 1983; Itakura 1987; Marchal and Anderson 1993). The fact that we did not observe social responses suggests two things: first, social responses by monkeys towards the mirror image may be primarily triggered by seeing their own face and gaze. As such, it is plausible that seeing their own face might be a salient stimulus that might more easily be mistaken for another individual. Second, monkeys in the present study did not appear to confuse their mirror image for another monkey; rather, as de Waal and colleagues (de Waal et al. 2005) have suggested, monkeys may represent an intermediate level of self recognition, not mistaking the reflection for a stranger, but not recognizing the reflection as self either.
Nielsen et al. (2006) suggested that passing a leg recognition test and a face recognition test requires essentially the same abilities, namely kinesthetic-visual matching as well as feature recognition. The present results appear to confirm this suggestion: monkeys fail to pass mark tests with marks placed on their faces or marks placed on other parts of their bodies. Mirror self-recognition is therefore not tied to recognition of facial features: face and body appear to be the same.
We have reason to believe that monkeys are capable of kinesthetic-visual matching as evidenced by their imitation recognition abilities (Paukner et al. 2005). Given their successful performance to retrieve food items, it appears that monkeys also understand mirrors as reflective surfaces. Why then do monkeys fail this test of self-recognition? The problem may lie with their ability to form expectations about their own appearance. Boccia (1994) pointed out that monkeys do not share with apes a keen interest in examining visual anomalies on their bodies (e.g. scratches or the presence of small visible objects, such as straw, etc.). While at this point it is difficult to say whether this lack of interest is the cause or the consequence of a failure to form expectations about their own appearance, it would appear to be an important factor to be investigated in future studies.
Gallup (1991) suggested moving away from mirror-image studies for the purpose of investigating self-recognition. One line of research that has received little attention in the literature, however, is macaques’ ability to use the mirror to find otherwise hidden food items. Even though it is known that macaques can use mirrors in this way (Anderson 1986), it is far less clear what the monkeys understand about mirrors in this context. That is, do monkeys understand that the reward in the mirror is the same reward that they will receive? Do they use the image of their own hand to guide their movement, or does the reflected food merely serve as a discriminative mark? Can monkeys distinguish between the image of their own hands and the image of a conspecific’s hand approaching the food? Video technology makes it feasible to test the effects of structural and temporal displacements of monkeys’ mirror/video-mediated behaviors, which might help to clarify monkeys’ understanding of mirrors. As shown by the recent study by Anderson et al. (2009), although self recognition was not found, monkeys appeared sensitive to the association between kinesthetic activity and external visual effect, discriminating between the real time image and the delayed image of themselves projected on the screen (see also Iriki et al, 2001). On the other hand, even human toddlers have difficulties with self-recognition tests when these are carried out using video recordings (e.g. Povinelli et al, 1996; Suddendorf, 1999), and some monkeys have been reported to threaten their own hand when projected on a TV screen (Menzel et al, 1985). Care should therefore be taken when using video or TV equipment for studying self-recognition. Indeed, a first step should be to investigate why both human toddlers and monkeys show such discrepancies in performance for what, to human adults, may be very similar media.
References
- Amsterdam BK. Mirror self-image reactions before age two. Dev Psychobiol. 1972;5:297–305. doi: 10.1002/dev.420050403. [DOI] [PubMed] [Google Scholar]
- Anderson JR. Mirror-mediated finding of hidden food by monkeys (Macaca tonkeana and Macaca fascicularis) J Comp Psychol. 1986;100:237–242. [Google Scholar]
- Anderson JR, Roeder JJ. Responses of capuchin monkeys (Cebus apella) to different conditions of mirror stimulation. Primates. 1989;30:581–587. [Google Scholar]
- Anderson JR, Kuroshima H, Paukner A, Fujita K. Capuchin monkeys (Cebus apella) respond to video image of themselves. Anim Cogn. 2009;12:55–62. doi: 10.1007/s10071-008-0170-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bahrick L, Moss L, Fadil C. Development of visual self-recognition in infancy. Ecol Psychol. 1996;8:189–208. [Google Scholar]
- Boccia ML. Mirror behavior in macaques. In: Parker ST, Mitchell RW, Boccia ML, editors. Self-awareness in animals and humans: developmental perspectives. Cambridge University Press; Cambridge: 1994. pp. 350–360. [Google Scholar]
- De Waal FBM, Dindo M, Freedman CA, Hall MJ. The monkey in the mirror: hardly a stranger. Proc Natl Acad Sci USA. 2005;102:11140–11147. doi: 10.1073/pnas.0503935102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Veer MW, Gallup GG, Jr, Theall LA, van den Bos R, Povinelli DJ. An 8-year longitudinal study of mirror self-recognition in chimpanzees (Pan troglodytes) Neuropsychologia. 2003;41:229–234. doi: 10.1016/s0028-3932(02)00153-7. [DOI] [PubMed] [Google Scholar]
- Eglash AR, Snowdon CT. Mirror-image responses in Pygmy marmoset (Cebuella pygmaea) Am J Primatol. 1983;5:211–219. doi: 10.1002/ajp.1350050305. [DOI] [PubMed] [Google Scholar]
- Field J, Adamiak J. Use of mirror reflection for self and non-self search during the second year. NZ J Psychol. 1990;19:58–62. [Google Scholar]
- Gallup GG., Jr Chimpanzees: Self-recognition. Science. 1970;167:86–87. doi: 10.1126/science.167.3914.86. [DOI] [PubMed] [Google Scholar]
- Gallup GG, Jr, Wallnau LB, Suarez SD. Failure to find self-recognition in mother-infant and infant-infant rhesus monkey pairs. Folia Primatol. 1980;33:210–219. doi: 10.1159/000155935. [DOI] [PubMed] [Google Scholar]
- Gallup GG., Jr . Toward a comparative psychology of self-awareness: species limitation and cognitive consequences. In: Goethals GR, Strauss J, editors. The self: an interdisciplinary approach. Springer Verlag; New York: 1991. pp. 121–135. [Google Scholar]
- Hauser MD, Kralik J, Botto-Mahan C, Garret M, Oser J. Self-recognition in primates: phylogeny and salience of species-typical features. Proc Natl Acad Sci USA. 1995;92:10811–10814. doi: 10.1073/pnas.92.23.10811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iriki A, Tanaka M, Obayashi S, Iwamura Y. Self-image in the video monitor coded by monkey intraparietal neurons. Neurosci Res. 2001;40:163–173. doi: 10.1016/s0168-0102(01)00225-5. [DOI] [PubMed] [Google Scholar]
- Itakura S. Mirror-guided behavior in Japanese macaques (Macaca fuscata fuscata) Primates. 1987;28:149–161. [Google Scholar]
- Itakura S. The level of self-knowledge in non-human primates: from the perspective of comparative cognitive science. In: Matsuzawa T, editor. Primate origin of human cognition and behavior. Springer; Tokyo: 2001. pp. 313–329. [Google Scholar]
- Lewis M, Brooks-Gunn J. Social cognition and the acquisition of self. Plenum; New York: 1979. [Google Scholar]
- Marchal P, Anderson JR. Mirror-image responses in Capuchin monkeys (Cebus apella): social responses and use of reflected environmental information. Folia Primatol. 1993;61:165–173. doi: 10.1159/000156745. [DOI] [PubMed] [Google Scholar]
- Meltzoff AN. Foundations for developing a concept of self: the role of imitation in relating self to other and the value of social mirroring, social modeling, and self practice in infancy. In: Cicchetti D, Beeghly M, editors. The self in transition: infancy to childhood. University of Chicago Press; Chicago: 1990. pp. 139–164. [Google Scholar]
- Meltzoff AN. The human infant as imitative generalist: a 20-year progress report on infant imitation with implications for comparative psychology. In: Heyes CM, Galef BG, editors. Social learning in animals: the roots of culture. Academic Press; London: 1996. pp. 347–370. [Google Scholar]
- Menzel EW, Jr, Savage-Rumbaugh ES, Lawson J. Chimpanzee (Pan troglodytes) spatial problem solving with the use of mirror and televised equivalent of mirrors. J Comp Psychol. 1985;99:211–217. [PubMed] [Google Scholar]
- Mitchell RW. Mental models of mirror-self-recognition: Two theories. New Ideas Psychol. 1993;11:295–325. [Google Scholar]
- Nielsen M, Suddendorf T, Slaughter V. Mirror self-recognition beyond the face. Child Dev. 2006;77:176–185. doi: 10.1111/j.1467-8624.2006.00863.x. [DOI] [PubMed] [Google Scholar]
- Parker ST, Mitchell RW, Boccia ML. Self-awareness in animals and humans. New York: Cambridge University Press; 1994. [Google Scholar]
- Paukner A, Anderson JR, Borelli E, Visalberghi E, Ferrari PF. Macaques (Macaca nemestrina) recognize when they are being imitated. Bio Lett. 2005;1:219–222. doi: 10.1098/rsbl.2004.0291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paukner A, Suomi S, Visalberghi E, Ferrari PF. Capuchin monkeys display affiliation toward humans who imitate them. Science. 2009;14:880–883. doi: 10.1126/science.1176269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Platt MM, Thompson RL, Boatright SL. Monkeys and mirrors. Questions of methodology. In: Fedigan LM, Asquith PJ, editors. The monkeys of Arashiyama: thirty-five years of research on Japan and the West. Suny Press; New York: 1991. pp. 274–290. [Google Scholar]
- Povinelli DJ, Landau KR, Perrilloux HK. Self-recognition in young children using delayed versus live feedback: evidence of a developmental asynchrony. Child Dev. 1996;67:1540–1554. [PubMed] [Google Scholar]
- Povinelli DJ, Gallup GG, Jr, Eddy TJ, Bierschwale DT, Engstrom MC, Perilloux HK, Toxopeus IB. Chimpanzees recognize themselves in mirrors. Anim Behav. 1997;53:1083–1088. [Google Scholar]
- Roma PG, Silberberg A, Huntsberry ME, Christensen CJ, Ruggiero AM, Suomi S. Mark test for mirror self-recognition in capuchin monkeys (Cebus apella) trained to touch marks. Am J Primatol. 2007;69:989–1000. doi: 10.1002/ajp.20404. [DOI] [PubMed] [Google Scholar]
- Shillito DJ, Gallup GG, Beck BB. Factors affecting mirror behavior in western lowland gorillas, Gorilla gorilla. Animl Behav. 1999;57:999–1004. doi: 10.1006/anbe.1998.1062. [DOI] [PubMed] [Google Scholar]
- Suarez SD, Gallup GG., Jr Self-recognition in chimpanzees and orangutans, but not gorillas. J Hum Evol. 1981;10:175–188. [Google Scholar]
- Suddendorf T. Children’s understanding of the relation between delayed video representation and current reality: a test for self-awareness? J Exp Child Psychol. 1999;72:157–176. doi: 10.1006/jecp.1998.2485. [DOI] [PubMed] [Google Scholar]
- Swartz KB, Sarauw D, Evans S. Comparative aspects of mirror self-recognition in great apes. In: Parker ST, Mitchell RW, Miles HL, editors. The mentalities of gorillas and orangutans in comparative perspective. Cambridge University Press; Cambridge: 1999. pp. 238–294. [Google Scholar]
- Thompson RL, Boatright-Horowitz SL. The question of mirror-mediated self recognition in apes and monkeys: some new results and reservation. In: Parker ST, Mitchell RW, Boccia ML, editors. Self-awareness in animals and humans: developmental perspectives. Cambridge University Press; Cambridge: 1994. pp. 330–349. [Google Scholar]