

Abstract
Stem cell niches are special microenvironments that maintain stem cells and control their behavior to ensure tissue homeostasis and regeneration throughout life. The liver has a high regenerative capacity that involves stem/progenitor cells when the proliferation of hepatocytes is impaired. In recent years progress has been made in the identification of potential hepatic stem cell niches. There is evidence that hepatic progenitor cells can originate from niches in the canals of Hering; in addition, the space of Disse may also serve as a stem cell niche during fetal hematopoiesis and constitute a niche for stellate cells in adults.
Introduction
The stem cell niche hypothesis was developed in 1978 by Raymond Schofield (1), who proposed that stem cells reside within fixed compartments that promote and maintain their characteristics. These microenvironments or niches regulate the behavior of stem cells by preserving their quiescent state or controlling the balance between self-renewal and differentiation after activation. These mechanisms sustain the stem cell pool to ensure tissue homeostasis and regeneration throughout life. The niche is defined not only by the site where a stem cell is located, but also by the composition of the microenvironment of the surrounding cells. Signals from the environment and intrinsic programs are required to control stemness, proliferation, and differentiation of stem cells.
Research on stem cell microenvironments in different tissues indicates recurring motifs in the niche composition. The interaction of neighboring cells with stem cells is critical for the establishment of a stem cell niche, through both secreted signaling factors and direct cell-cell contacts (2). Endothelial cells and stromal cells of the bone marrow, for example, release CXCL12, also called stromal cell–derived factor 1 (3–5), which is the only ligand for CXCR4. The interaction of CXCL12 and CXCR4 is essential to initiate and maintain stem cell niches through the control of stem/progenitor cell migration. The recruitment of stem cells from the bone marrow is partly mediated by local downregulation of CXCL12, which facilitates their mobilization into the bloodstream (6). Other factors secreted by stem cell neighbors are also known to regulate stem cell behavior. The β-catenin–dependent or canonical WNT signaling pathway can preserve quiescence in hematopoietic stem cells and is essential to maintain stemness (7–9) but was also reported to initiate cell differentiation (10, 11). The noncanonical WNT ligand WNT5a, which mediates β-catenin–independent WNT signaling, was recently identified as an important factor of the hematopoietic stem cell niche to regulate quiescence by counteracting canonical WNT signaling (10). A possible role of hedgehog (Hh) signaling for the maintenance of quiescent stem cells in their niche is not established yet and requires further investigation. However, some evidence has been presented that Hh signaling determines the development of stem cells in their niche (12). Members of the TGF-β family are TGF-β, nodal, activin, and BMP. This family controls stem cell fate by affecting diverse cellular functions, such as growth arrest, apoptosis, migration, and differentiation. BMP4, for example, is required for hematopoiesis and expressed by cells of the microenvironment of hematopoietic stem cells (13). TGF-β signaling is essential to maintain immaturity and quiescence of melanocyte stem cells in their niches within the hair bulge (14). Contacts of stem cells with neighboring cells are also pivotal for stem cell niches, as adherens junctions via cadherins play a role in the control of asymmetric and symmetric stem cell divisions (15). One example is neuronal cadherin, which is expressed by mesenchymal stem cells (MSCs) and hematopoietic stem/progenitor cells to enable cellular interactions (16). In addition, signaling pathways such as Notch require a direct physical contact of signal-sending and signal-receiving cells. Notch1 receptor, for instance, was shown to be critical to maintaining neuronal proliferation of stem cells in their niche, whereas other Notch receptors are apparently required for further cell development (11, 17–19).
Stem cells are often found on basement membranes that typically underlie the epithelium or endothelium (20). The contact of stem cells with basement membrane proteins is mediated by adhesion molecules called integrins. A loss or alteration of integrin expression enables recruitment of stem cells (21). Furthermore, the contact of niche cells to the peripheral nervous system helps to control stem cell recruitment. For instance, hematopoietic stem cell recruitment from the bone marrow into the bloodstream can be initiated by the sympathetic nervous system via innervation of their niche (22). High local concentrations of norepinephrine, the neurotransmitter of the peripheral sympathetic nervous system, in the hematopoietic stem cell niche are mediated by granulocyte colony–stimulating factor, apparently by inhibiting norepinephrine reuptake by nerve terminals, leading to a high sympathetic tone and finally enhanced stem/progenitor cell mobilization (22, 23). Despite the importance of all niche elements mentioned above, the presence of a stem cell ultimately defines the stem cell niche. Therefore, the identification of stem cells is usually the first step in defining stem cell niches within tissues of interest. Stem cells have two general properties: (a) they are able to renew themselves by dividing for long periods of time, and (b) they are unspecialized cells that can generate highly specialized effector cells (24). These characteristics must be fulfilled by stem cell candidates and are normally tested by a full set of rigorous in vitro and in vivo techniques such as multiple surface marker analysis, cell clone expansion assays, colony formation assays, multipotency tests, transplantation assays, and cell lineage–tracing studies. Most of these test systems were originally evolved in the hematopoietic stem cell field, which has a long history of research starting in the late 19th century (25).
Examples of stem cell niches
Well-defined stem cell niches in vertebrates are located in the bone marrow, crypts of the small intestine, and the bulge of hair follicles (26–29). All of these tissues are characterized by substantial cell turnover, which facilitates the detection of stem cell activity due to their continuous supply of cells. Apart from molecular markers typically expressed by stem cells, their ability to retain 3H-thymidine or the thymidine analog BrdU during asymmetric cell proliferation is often used for their identification. When a stem cell divides asymmetrically, one daughter cell remains a stem cell and retains 50% of the DNA label, and the other daughter cell becomes a lineage-committed progenitor cell (transit-amplifying cell) that steadily dilutes the label during proliferation. Although this method is intriguing, it has several disadvantages. If progenitor cells stop their proliferation and finally differentiate, mature effector cells can also retain the DNA label for a long time (30), and quiescent stem cells that did not enter the cell cycle during the labeling period remain unmarked. Stem cells in tissues with less cell turnover are more difficult to detect. One example is the brain, where the presence and activity of neuronal stem cells have been recently demonstrated despite the long-held belief that the adult brain cannot generate new neurons (31). Indeed, neuronal stem cells are now thought to be involved in learning and memory formation (31). A subpopulation of astrocytes that express glial fibrillary acidic protein (GFAP) was identified as neuronal stem cells in the subventricular and subgranular zone of the brain in mammals (32, 33). Liver also has relatively low cell turnover rates. The lifespan of hepatocytes is about 150–450 days under normal conditions (34, 35), but the liver has a high regenerative capacity after injury, as discussed below.
Hepatic stem cell niches
Among adult mammalian organs the liver appears unique, in that normal cell turnover is managed by the proliferation of differentiated parenchymal and nonparenchymal cells. The proverbial rapid reconstitution of liver mass after intoxication or injury is realized by residual liver cells without obvious contribution of stem/progenitor cells (36, 37). Liver stem/progenitor cells, termed hepatic progenitor cells in humans (38) and oval cells in rodents (39, 40), emerge when hepatocyte proliferation is overwhelmed by persistent or severe liver injury. Oval cells, named for their morphological appearance, are bipotent precursor cells that can contribute to the formation of bile ducts (41) and to the regeneration of liver parenchyma as determined by transplantation studies (42–44). They represent a heterogeneous population of duct-forming or nonductular cells that show varying expression patterns (45), with characteristics of bile duct cells (41) and immature parenchymal cells (46). These precursor cells appear in the portal field of the liver (46) after persistent severe liver injury such as chronic viral hepatitis (47) or proliferative senescence of hepatocytes in steatohepatitis (48) and are thought to originate from the terminal branches of the intrahepatic biliary system, the canals of Hering (49).
Canals of Hering.
Substantial evidence for the existence of a stem cell niche in the canals of Hering, which are lined by small cholangiocytes and hepatocytes, has come from label-retention assays (50). In addition, moderate hepatic injury through sublethal acetaminophen intoxication revealed several locations for BrdU-retaining cells: in the canals of Hering, in intralobular bile ducts, in the periphery of bile ducts, and within the portal field, which contained small label-retaining hepatocytes (50) (Figure 1). Stem/progenitor cells that reside within the canals of Hering and bile ducts seem to generate these small hepatocytes, which retain the BrdU label for at least 2 months (50). This interpretation supports the earlier concept of the “streaming liver,” which states that new hepatocytes are continuously generated in the portal field and mature on their way to the central vein (51). It should be mentioned here that much controversy has surrounded this concept in the past, as discussed by others previously (35). In addition, cells without hepatobiliary markers (termed null cells) that are close to portal tracts are also label-retaining cells, but their identity and origin remains unclear (50) (Figure 1). Null cells might represent stem/progenitor cells from extrahepatic sites and may originate from circulating bone marrow cells (50, 52). Bone marrow–derived stem/progenitor cells that express thymus cell antigen-1 were already reported to contribute to liver regeneration (53).
Figure 1. Model of the hepatic stem cell niche in the canals of Hering.

Label-retaining cells (black nuclei) represent putative stem cells and are located in the canals of Hering as keratin-expressing cells. Also, cholangiocytes of intralobular bile ducts, small hepatocytes, and less-characterized null cells retain the BrdU label after acetaminophen treatment of mice (50). Coexpression of keratin 19 and albumin in small hepatocytes at the interface of cholangiocytes and hepatocytes in the canals of Hering indicate that hepatocytes are generated at this site by stem/progenitor cells (116). A continuous production of hepatocytes within the portal field is supported by SOX9 fate-mapping analysis (54). Small cholangiocytes lining the canals of Hering and ductules may represent cholangiocyte precursors, which potentially contribute to the cholangiocyte population of larger bile ducts (116) and could explain the presence of label-retaining cholangiocytes at this site (50).
Additional support for a stem cell niche within the portal field was provided by a lineage-tracing study that relied on the detection of sex determining region Y–box containing gene 9–expressing (SOX9-expressing) cells (54). In this transgenic mouse model, the SOX9 promoter controlled the expression of a Cre recombinase fused to mutated human estrogen receptor T2 (ERT2), which cleaved a stop codon following tamoxifen binding to enable the irreversible expression of the β-galactosidase (LacZ) reporter gene. This approach permanently labeled SOX9-expressing cells and their progeny. Time-resolved analysis of LacZ appearance by X-gal staining after tamoxifen exposure revealed that the reporter first appeared in bile duct cells, which include cells in the canals of Hering, and then in hepatocytes (54). Thus, bile duct cells seem to represent precursor cells of hepatocytes. The number of LacZ-labeled hepatocytes increased with time, and hepatocytes from the SOX9 lineage spread from the portal field to the central vein, reminiscent of the streaming liver hypothesis (51, 54). This observation was made in the normal mouse liver but was more pronounced after applying a methionine- and choline-deficient diet supplemented with ethionine (MCDE) (54), which is a chronic hepatic injury model associated with an oval cell response (55, 56). Interestingly, SOX9 fate mapping failed to label α-fetoprotein–expressing oval cells that appeared on the MCDE diet (54). A small subpopulation of oval cells seemed to derive from the SOX9-positive biliary duct cells (54), which further supports the concept of a stem cell niche associated with bile ducts. Given that precursor cells involved in the early organogenesis of the liver remained unlabeled by SOX9 fate mapping as observed with α-fetoprotein–positive oval cells in adult mice (54), it should be clarified whether SOX9 is simply not expressed or whether the reporter gene is silenced by epigenetic mechanisms in these cells; silencing of reporter genes is frequently observed in stem/progenitor cells from transgenic animal models (57–60).
Human hepatic stem cells are described as small epithelial cell adhesion molecule–expressing (EpCAM-expressing) cells that constitute approximately 0.5%–2.5% of the adult liver parenchyma. They lack α-fetoprotein synthesis but exhibit the expression of some cholangiocyte and hepatocyte markers (61). As the epithelium of small bile ducts expresses EpCAM in normal liver (62), it is possible that several cells represent stem/progenitor cells in the canals of Hering. The existence of primitive stem cells without hepatobiliary markers at this site is not established yet, and more stem cell–restricted molecular markers are necessary to elucidate this. Relatively few data on the surrounding environment of hepatic stem cells in the canals of Hering in normal liver are available thus far. Most studies have relied on murine liver injury models associated with an oval cell response to characterize possible niche elements. These studies revealed that one key element of the niche is the extracellular matrix component laminin, a protein that maintains the undifferentiated phenotype of oval cells and promotes their proliferation in vitro (63, 64). Indeed, a basement membrane is present in the canals of Hering (65). Potentially all cell types of the liver can interact with hepatic stem cells in the canals of Hering or with duct-forming oval cells after severe liver injury, including more mature cholangiocytes, hepatocytes, stellate cells, portal myofibroblasts, liver sinusoidal endothelial cells (LSECs), and cells of the immune system such as macrophages.
Several studies have investigated the signaling factors released or presented by neighboring cells that influence the behavior of oval cells. Hepatic stellate cells (also called Ito cells) and myofibroblasts are closely associated with oval cells during liver regeneration (11, 66) and release growth factors, chemokines, and cytokines that can potentially influence their survival, proliferation, migration, and differentiation (61, 67–74). HGF is one signaling factor released by stellate cells that can affect migration, proliferation, and differentiation of hepatic progenitor cells (67, 75, 76). Prevention of hepatic stellate cell activation through a diet supplemented with 2% L-cysteine is associated with a significantly decreased oval cell response in rats after inhibition of hepatocyte proliferation by 2-acetylaminofluorene (2AAF) and subsequent liver injury through partial hepatectomy (73), demonstrating the importance of activated stellate cells as supportive stromal cells for hepatic progenitor cells. Activation of canonical WNT signaling via β-catenin through the ligand WNT3a secreted by macrophages initiates the differentiation of oval cells into hepatocytes, whereas stimulation of Notch signaling in oval cells via the ligand jagged 1 from myofibroblasts favors their differentiation into cholangiocytes (11). This provides an interesting example of cell fate specification through the competing interaction of Notch and canonical WNT signaling. Thus, stellate cells, myofibroblasts, and macrophages obviously act as supporting cells to create a suitable microenvironment for expanding hepatic progenitor cells (11, 64, 67, 68, 73). Little information is available yet about possible influences of hepatocytes, mature cholangiocytes, and LSECs on hepatic progenitor cells. Initial evidence exists that injured or dying hepatocytes release the Hh ligands sonic Hh (Shh) and Indian Hh (Ihh), which can activate the Hh pathway in hepatic progenitor cells to promote their survival and expansion (77, 78). Afferent vagal innervation of hepatic bile ducts has been reported in one study (79), and nerves extend into the canals of Hering (65). Interestingly, the recruitment of hepatic progenitor cells is enhanced following inhibition of the sympathetic nervous system (80) or disconnection of the hepatic nerves during liver transplantation (81). The peripheral nervous system apparently controls the recruitment of progenitor cells in the canals of Hering.
Space of Disse.
Until hematopoiesis is established in the bone marrow postnatally, the liver is one of the main hematopoietic organs during fetal development. Sites of blood formation are visible through clusters of hematopoietic stem/progenitor cells within the sinusoids of fetal liver parenchyma (82). During fetal development, hematopoietic stem cells constitute 0.1%–0.2% of the liver cells (83). These niches may potentially be occupied again by hematopoietic stem/progenitor cells in adults when the bone marrow fails to provide an adequate environment in diseases such as osteomyelofibrosis (84). In addition to the canals of Hering, other areas of the liver can also transiently provide a niche for stem cells.
Similar to the bone marrow, the liver exhibits a discontinuous capillary network made by fenestrated LSECs. The space of Disse, named after the German anatomist and histologist Joseph Disse, is a unique perisinusoidal space within the liver parenchyma delineated by LSECs and parenchymal cells. Microvilli of parenchymal cells extend into this intercellular space to improve the uptake of blood plasma components. Hepatic stellate cells are vitamin A–storing cells that typically reside within the space of Disse. Stellate cells are mainly known as the primary collagen-producing cells following liver injury, which are responsible for fibrogenesis in chronic liver disease (85). In addition to a role for stellate cells in fibrogenesis, evidence exists that stellate cells have properties of stem cells (86–92). The expression of stem/progenitor cell–associated genes such as nestin in stellate cells of the liver and pancreas (86, 89, 92–94) and the potential of stellate cells to differentiate into hepatocyte-like cells in vitro (86, 92) indicate that stellate cells have stem cell characteristics. Moreover, stellate cells can grow as cell clones in culture and maintain their capability to differentiate, which suggests self-renewal potential (92). Further supporting this notion, hepatic stellate cells have the potential to develop into undifferentiated embryonal sarcoma cells in the liver (95). Fate-mapping analyses that relied on a GFAP-dependent reporter mouse model indicated that cells expressing this stellate cell marker contributed to liver regeneration through differentiation into hepatocytes and cholangiocytes after application of a MCDE diet (88). This contribution of stellate cells to liver regeneration was recently confirmed with pancreatic stellate cells, which were transplanted via tail vein injections into rats that underwent partial hepatectomy in the presence of 2AAF and substantially contributed to liver regeneration through differentiation into epithelial cell lineages (92). An important role of stellate cells in liver regeneration is also underlined by the fact that the CD95 ligand does not induce apoptosis in quiescent hepatic stellate cells despite expression of CD95/Fas in these cells (96, 97). In quiescent hepatic stellate cells, CD95 ligand even inactivates CD95 through tyrosine nitration and acts as a mitogen through Src-dependent shedding of the epidermal growth factor (96). Though some experimental evidence supports the idea that stellate cells function as stem/progenitor cells, the stem cell status of stellate cells and their contribution to tissue repair remains controversial in the field, given that stem/progenitor cell–based liver regeneration is generally seen as a process that involves keratin-expressing progenitor cells. Although the participation of mesodermal cells in epithelial tissue regeneration is under debate, evidence suggests that MSCs from bone marrow or adipose tissue can contribute to liver repair through differentiation (98–100). Since MSCs are reported to occur in all organs (101), it is important to elucidate a possible function of MSCs in maintaining hepatic stem cell niches as supportive stromal cells. Interestingly, desmin-expressing hepatic stellate cells are closely associated with hematopoietic sites in the fetal liver and are suggested to support hematopoiesis as liver-resident MSCs (102, 70, 91, 103). Similar to neuronal stem cells and bone marrow MSCs (33, 104), hepatic stellate cells seem to fulfill a dual role as supportive cells on the one hand and precursor cells on the other.
Quiescent stellate cells are maintained in the space of Disse. A basement membrane–like structure with reticular collagen IV and laminin is present in normal liver, and basement membrane proteins are known to preserve quiescence in stellate cells (105). Activated stellate cells show a phenotype reminiscent of myofibroblasts that is reversible, as recently observed in vivo (106, 107). Liver parenchymal cells sustain quiescence of stellate cells by the release of soluble factors that apparently include canonical WNT ligands when they are cultured together (72, 108). The chemokine CXCL12 can attract hepatic stellate cells via CXCR4 (71, 72), and stellate cells were experimentally shown to migrate to LSECs in response to CXCL12 (72). In addition, stellate cells start to release CXCL12 after activation (70–72) and can potentially attract other cells. Apart from signaling pathways that rely on soluble factors, Notch signaling requires cell contacts and may also be relevant for stellate cells. In the liver, signal-sending cells that present Notch ligands such as jagged 1 are hepatocytes and cholangiocytes (72, 109, 110), whereas quiescent hepatic stellate cells exhibit no jagged 1 protein synthesis (72). Jagged 1 appears during the activation of stellate cells (72), which further indicates that activated stellate cells can also provide a suitable environment for hepatic progenitor cells. Nerve endings are also found close to stellate cells in the liver (111). Hepatic stellate cells can directly respond to perivascular nerve stimulation through the release of the osmolyte myoinositol and exhibit Ca2+ transients in response to phenylephrine (112, 113). Their stimulation with norepinephrine leads to a rapid release of prostaglandins, which can activate glycogenolysis in neighboring liver parenchymal cells (114, 115), thereby increasing the local glucose concentration. Thus, stellate cells can integrate signals from distant cells or organs to affect the behavior of neighboring cells in their niche. These findings indicate that the microenvironment in the space of Disse controls the behavior of stellate cells and represents their niche (Figure 2).
Figure 2. Model of intercellular communication in the stellate cell niche of normal liver.
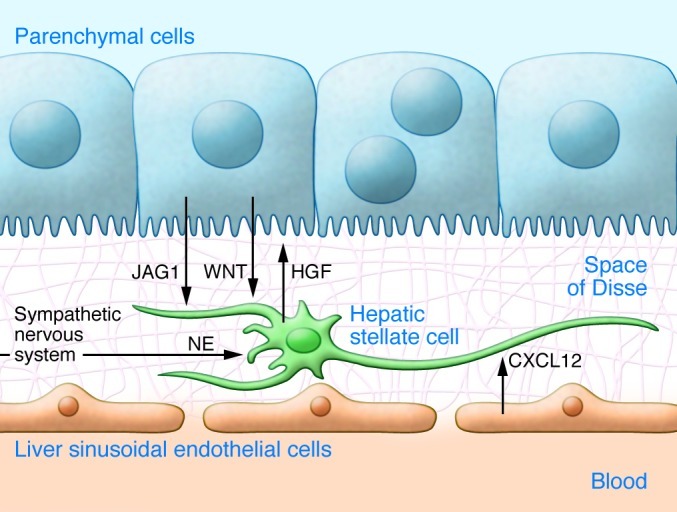
Hepatic stellate cells (HSCs) are located on basement membrane proteins such as laminin and collagen type IV (grid) between LSECs and parenchymal cells (PCs). HSCs are attracted to LSECs through CXCL12 and are associated with PCs, which express Notch ligands such as jagged 1 (JAG1) and release canonical WNT ligands to influence stellate cell development (72). Quiescent stellate cells in turn secrete HGF, which is probably involved in liver tissue homeostasis and can support hepatic progenitor cells (75, 76). HSCs receive signals from the sympathetic nervous system through norepinephrine (NE) release (112, 115).
Future research and perspectives
The identification and elucidation of hepatic stem cell niche composition and function in the normal and regenerating liver are pivotal to developing therapeutic strategies for patients with impaired liver regeneration, which is usually seen in chronic liver diseases. Moreover, proper in vitro expansion of stem/progenitor cells without loss of their developmental potential, as well as establishment of cell differentiation protocols for the generation of functional hepatocytes, are important for therapeutic cell transplantation. Substantial progress has been made in recent years in all of these areas. However, additional fundamental research using rigorous test systems, such as those established in the hematopoietic stem cell field, is required to fully characterize liver stem cells and to understand the architecture of the stem cell niches in the liver. It is not clear yet whether the expansion of ducts by proliferating hepatic progenitor cells during liver regeneration exactly mirrors the niche in the canals of Hering. Therefore, future studies should also investigate the stem cell niche in the normal liver as well. The selective inhibition of niche components by using knockout animals and specific inhibitors of receptors may provide new insights. Possible targets include niche elements such as cell-cell contacts (e.g., neuronal cadherin), signaling pathways (e.g., canonical/noncanonical WNT, Notch, TGF-β family, Hh), the CXCL12/CXCR4 axis, basement membrane proteins (e.g., laminin, collagen type IV), and integrins. Moreover, putative stem cells in the canals of Hering should be better defined by unique markers to enable their exact characterization and to clarify the presence of stem cells without hepatobiliary markers at this site. Homing of transplanted EpCAM+ hepatic stem cells in the canals of Hering has not yet been reported and would further support the concept of the stem cell niche. In contrast, homing of migrating hematopoietic stem cells can reoccur in the space of Disse of the adult liver under certain conditions. Additionally, transplanted stellate cells are capable of homing to the space of Disse (92). The first evidence is provided by in vitro experiments examining possible interactions of niche elements in the space of Disse (72), but these findings need to be confirmed in animal models as described above.
As general features of stem cell niches can be found in both the canals of Hering and the space of Disse, it is worth outlining the differences between these two sites. The canals of Hering appear to contain slow-cycling precursor cells in the normal liver (50, 54, 116), whereas the space of Disse normally contains quiescent stellate cells and hematopoietic stem/progenitor cells that only rarely divide. The direct exposure of hepatic stem cells to high concentrations of bile acids in the canals of Hering represents a prominent difference from the environment in the space of Disse. Bile acids act not only as emulgators of lipids but also as signaling molecules that can support liver regeneration (117). Another difference in the composition of these two niches is the presence of LSECs in the space of Disse and cholangiocytes in the canals of Hering, both of which can release CXCL12 to attract CXCR4-expressing cells (72, 118). Additional differences and their consequences may become apparent in further analyses. Future research should also address how hepatic stem cell niches are affected by chronic diseases and aging. Are niches altered or lost, and are these changes responsible for impaired liver regeneration? Variation in the composition and abundance of basement membrane proteins can be observed in liver diseases (119), which may also adversely affect the maintenance of stem cell niches. An age-related decline in stem cell niche integrity is known (120) and could lead to impaired self-renewal and inappropriate differentiation of stem cells in their niche. Indeed, further characterization of the liver stem cell niches as well as changes in liver diseases are challenges for future research that promise new insights into liver physiology.
Acknowledgments
The authors are grateful to the German Research Foundation (Deutsche Forschungsgemeinschaft) for the financial support through the Collaborative Research Center 575 (SFB 575) Experimental Hepatology and Collaborative Research Center 974 (SFB 974) Communication and Systems Relevance during Liver Injury and Regeneration.
Footnotes
Conflict of interest: The authors have declared that no conflict of interest exists.
Citation for this article:J Clin Invest. 2013;123(5):1874–1880. doi:10.1172/JCI66027.
References
- 1.Schofield R. The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells. 1978;4(1–2):7–25. [PubMed] [Google Scholar]
- 2.Morrison SJ, Spradling AC. Stem cells and niches: mechanisms that promote stem cell maintenance throughout life. Cell. 2008;132(4):598–611. doi: 10.1016/j.cell.2008.01.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Peled A, et al. The chemokine SDF-1 stimulates integrin-mediated arrest of CD34(+) cells on vascular endothelium under shear flow. J Clin Invest. 1999;104(9):1199–1211. doi: 10.1172/JCI7615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Imai K, et al. Selective secretion of chemoattractants for haemopoietic progenitor cells by bone marrow endothelial cells: a possible role in homing of haemopoietic progenitor cells to bone marrow. Br J Haematol. 1999;106(4):905–911. doi: 10.1046/j.1365-2141.1999.01644.x. [DOI] [PubMed] [Google Scholar]
- 5.Schajnovitz A, et al. CXCL12 secretion by bone marrow stromal cells is dependent on cell contact and mediated by connexin-43 and connexin-45 gap junctions. Nat Immunol. 2011;12(5):391–398. doi: 10.1038/ni.2017. [DOI] [PubMed] [Google Scholar]
- 6.Lapidot T, Dar A, Kollet O. How do stem cells find their way home? Blood. 2005;106(6):1901–1910. doi: 10.1182/blood-2005-04-1417. [DOI] [PubMed] [Google Scholar]
- 7.Fleming HE, et al. Wnt signaling in the niche enforces hematopoietic stem cell quiescence and is necessary to preserve self-renewal in vivo. Cell Stem Cell. 2008;2(3):274–283. doi: 10.1016/j.stem.2008.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Reya T, et al. A role for Wnt signalling in self-renewal of haematopoietic stem cells. Nature. 2003;423(6938):409–414. doi: 10.1038/nature01593. [DOI] [PubMed] [Google Scholar]
- 9.Sato N, Meijer L, Skaltsounis L, Greengard P, Brivanlou AH. Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat Med. 2004;10(1):55–63. doi: 10.1038/nm979. [DOI] [PubMed] [Google Scholar]
- 10.Sugimura R, et al. Noncanonical Wnt signaling maintains hematopoietic stem cells in the niche. Cell. 2012;150(2):351–365. doi: 10.1016/j.cell.2012.05.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Boulter L, et al. Macrophage-derived Wnt opposes Notch signaling to specify hepatic progenitor cell fate in chronic liver disease. Nat Med. 2012;18(4):572–579. doi: 10.1038/nm.2667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Brownell I, Guevara E, Bai CB, Loomis CA, Joyner AL. Nerve-derived sonic hedgehog defines a niche for hair follicle stem cells capable of becoming epidermal stem cells. Cell Stem Cell. 2011;8(5):552–565. doi: 10.1016/j.stem.2011.02.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Goldman DC, Bailey AS, Pfaffle DL, Al Masri A, Christian JL, Fleming WH. BMP4 regulates the hematopoietic stem cell niche. Blood. 2009;114(20):4393–4401. doi: 10.1182/blood-2009-02-206433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nishimura EK, et al. Key roles for transforming growth factor beta in melanocyte stem cell maintenance. Cell Stem Cell. 2010;6(2):130–140. doi: 10.1016/j.stem.2009.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Marthiens V, Kazanis I, Moss L, Long K, Ffrench-Constant C. Adhesion molecules in the stem cell niche--more than just staying in shape? J Cell Sci. 2010;123(pt 10):1613–1622. doi: 10.1242/jcs.054312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wein F, et al. N-cadherin is expressed on human hematopoietic progenitor cells and mediates interaction with human mesenchymal stromal cells. Stem Cell Res. 2010;4(2):129–139. doi: 10.1016/j.scr.2009.12.004. [DOI] [PubMed] [Google Scholar]
- 17.Nyfeler Y, et al. Jagged1 signals in the postnatal subventricular zone are required for neural stem cell self-renewal. EMBO J. 2005;24(19):3504–3515. doi: 10.1038/sj.emboj.7600816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Basak O, Giachino C, Fiorini E, Macdonald HR, Taylor V. Neurogenic subventricular zone stem/progenitor cells are Notch1-dependent in their active but not quiescent state. J Neurosci. 2012;32(16):5654–5666. doi: 10.1523/JNEUROSCI.0455-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Karanu FN, Yuefei L, Gallacher L, Sakano S, Bhatia M. Differential response of primitive human CD34– and CD34+ hematopoetic cells to the Notch ligand Jagged-1. . Leukemia. 2003;17(7):1366–1374. doi: 10.1038/sj.leu.2402973. [DOI] [PubMed] [Google Scholar]
- 20.Fuchs E, Tumbar T, Guasch G. Socializing with the neighbors: stem cells and their niche. Cell. 2004;116(6):769–778. doi: 10.1016/S0092-8674(04)00255-7. [DOI] [PubMed] [Google Scholar]
- 21.Watt FM, Hogan BL. Out of Eden: stem cells and their niches. Science. 2000;287(5457):1427–1430. doi: 10.1126/science.287.5457.1427. [DOI] [PubMed] [Google Scholar]
- 22.Katayama Y, et al. Signals from the sympathic nervous system regulate hematopoietic stem cell egress from the bone marrow. Cell. 2006;124(2):407–421. doi: 10.1016/j.cell.2005.10.041. [DOI] [PubMed] [Google Scholar]
- 23.Lucas D, et al. Norepinephrine reuptake inhibition promotes mobilization in mice: potential impact to rescue low stem cell yields. Blood. 2012;119(17):3962–3965. doi: 10.1182/blood-2011-07-367102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Weissman IL. Stem cells: units of development, units of regeneration, and units in evolution. Cell. 2000;100(1):157–168. doi: 10.1016/S0092-8674(00)81692-X. [DOI] [PubMed] [Google Scholar]
- 25.Ramalho-Santos M, Willenbring H. On the origin of the term “stem cell”. Cell Stem Cell. 2007;1(1):35–38. doi: 10.1016/j.stem.2007.05.013. [DOI] [PubMed] [Google Scholar]
- 26.Ehninger A, Trumpp A. The bone marrow stem cell niche grows up: mesenchymal stem cells and macrophages move in. J Exp Med. 2011;208(3):421–428. doi: 10.1084/jem.20110132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Barker N, et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449(7165):1003–1007. doi: 10.1038/nature06196. [DOI] [PubMed] [Google Scholar]
- 28.Taylor G, Lehrer MS, Jensen PJ, Sun TT, Lavker RM. Involvement of follicular stem cells in forming not only the follicle but also the epidermis. Cell. 2000;102(4):451–461. doi: 10.1016/S0092-8674(00)00050-7. [DOI] [PubMed] [Google Scholar]
- 29.Oshima H, Rochat A, Kedzia C, Kobayashi K, Barrandon Y. Morphogenesis and renewal of hair follicles from adult multipotent stem cells. Cell. 2001;104(2):233–245. doi: 10.1016/S0092-8674(01)00208-2. [DOI] [PubMed] [Google Scholar]
- 30. Goldman B. Magic marker myths.Nature Reports Stem Cells. Nature Web site. http://www.nature.com/stemcells/2008/0801/080131/full/stemcells.2008.26.html . Updated January 31, 2008. Accessed March 6, 2013.
- 31.Deng W, Aimone JB, Gage FH. New neurons and new memories: how does adult hippocampal neurogenesis affect learning and memory? Nat Rev Neurosci. 2010;11(5):339–350. doi: 10.1038/nrn2822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Doetsch F, Caille I, Lim DA, Garcia-Verdugo JM, Alvarez-Buylla A. Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell. 1999;97(6):703–716. doi: 10.1016/S0092-8674(00)80783-7. [DOI] [PubMed] [Google Scholar]
- 33.Doetsch F. A niche for adult neural stem cells. Curr Opin Genet Dev. 2003;13(5):543–550. doi: 10.1016/j.gde.2003.08.012. [DOI] [PubMed] [Google Scholar]
- 34.MacDonald RA. Lifespan of liver cells. Arch Intern Med. 1961;107:335–343. doi: 10.1001/archinte.1961.03620030023003. [DOI] [PubMed] [Google Scholar]
- 35.Bralet MP, Branchereau S, Brechot C, Ferry N. Cell lineage study in the liver using retroviral mediated gene transfer. Evidence against the streaming of hepatocytes in normal liver. Am J Pathol. 1994;144(5):896–905. [PMC free article] [PubMed] [Google Scholar]
- 36.Michalopoulos GK. Liver regeneration. J Cell Physiol. 2007;213(2):286–300. doi: 10.1002/jcp.21172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Riehle KJ, Dan YY, Campbell JS, Fausto N. New concepts in liver regeneration. J Gastroenterol Hepatol. 2011;26(suppl 1):203–212. doi: 10.1111/j.1440-1746.2010.06539.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.De Vos R, Desmet V. Ultrastructural characteristics of novel epithelial cell types identified in human pathologic liver specimens with chronic ductular reaction. Am J Pathol. 1992;140(6):1441–1450. [PMC free article] [PubMed] [Google Scholar]
- 39.Farber E. Similarities of the sequence of the early histological changes induced in the liver of the rat by 2-acetylaminofluorine, and 3-methyl-4-dimethylaminoazo- benzene. Cancer Res. 1956;16:142–148. [PubMed] [Google Scholar]
- 40.Wilson JW, Leduc EH. Role of cholangioles in restoration of the liver of the mouse after dietary injury. J Pathol Bacteriol. 1958;76(2):441–449. doi: 10.1002/path.1700760213. [DOI] [PubMed] [Google Scholar]
- 41.Lenzi R, et al. Histogenesis of bile duct-like cells proliferating during ethionine hepatocarcinogenesis: evidence for a biliary epithelial nature of oval cells. Lab Invest. 1992;66(3):390–402. [PubMed] [Google Scholar]
- 42.Faris RA, Hixson DC. Selective proliferation of chemically altered rat liver epithelial cells following hepatic transplantation. Transplantation. 1989;48(1):87–92. doi: 10.1097/00007890-198907000-00021. [DOI] [PubMed] [Google Scholar]
- 43.Yasui O, Miura N, Terada K, Kawarada Y, Koyama K, Sugiyama T. Isolation of oval cells from Long-Evans Cinnamon rats and their transformation into hepatocytes in vivo in the rat liver. Hepatology. 1997;25(2):329–334. doi: 10.1053/jhep.1997.v25.pm0009021943. [DOI] [PubMed] [Google Scholar]
- 44.Song S, et al. Ex vivo transduced liver progenitor cells as a platform for gene therapy in mice. Hepatology. 2004;40(4):918–924. doi: 10.1002/hep.20404. [DOI] [PubMed] [Google Scholar]
- 45.Koenig S, Probst I, Becker H, Krause P. Zonal hierarchy of differentiation markers and nestin expression during oval cell mediated rat liver regeneration. Histochem Cell Biol. 2006;126(6):723–734. doi: 10.1007/s00418-006-0204-3. [DOI] [PubMed] [Google Scholar]
- 46.Debeva MD, Shafritz DA. Activation, proliferation, and differentiation of progenitor cells into hepatocytes in the D-galactosamine model of liver regeneration. Am J Pathol. 1993;143(6):1606–1620. [PMC free article] [PubMed] [Google Scholar]
- 47.Libbrecht L, Desmet V, Van Damme B, Roskams T. Deep intralobular extension of human hepatic ‘progenitor cells’ correlates with parenchymal inflammation in chronic viral hepatitis: can ‘progenitor cells’ migrate? J Pathol. 2000;192(3):373–378. doi: 10.1002/1096-9896(2000)9999:9999<::AID-PATH700>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- 48.Yang S, et al. Oval cells compensate for damage and replicative senescence of mature hepatocytes in mice with fatty liver disease. Hepatology. 2004;39(2):403–411. doi: 10.1002/hep.20082. [DOI] [PubMed] [Google Scholar]
- 49.Theise ND, et al. The canals of Hering and hepatic stem cells in humans. Hepatology. 1999;30(6):1425–1433. doi: 10.1002/hep.510300614. [DOI] [PubMed] [Google Scholar]
- 50.Kuwahara R, Kofman AV, Landis CS, Swenson ES, Barendswaard E, Theise ND. The hepatic stem cell niche: identification by label-retaining cell assay. Hepatology. 2008;47(6):1994–2002. doi: 10.1002/hep.22218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zajicek G, Oren R, Weinreb M., Jr The streaming liver. Liver. 1985;5(6):293–300. doi: 10.1111/j.1600-0676.1985.tb00252.x. [DOI] [PubMed] [Google Scholar]
- 52.Sell S. Heterogeneity and plasticity of hepatocyte lineage cells. Hepatology. 2001;33(3):738–750. doi: 10.1053/jhep.2001.21900. [DOI] [PubMed] [Google Scholar]
- 53.Petersen BE, et al. Bone marrow as a potential source of hepatic oval cells. Science. 1999;284(5417):1168–1170. doi: 10.1126/science.284.5417.1168. [DOI] [PubMed] [Google Scholar]
- 54.Furuyama K, et al. Continuous cell supply from a Sox9-expressing progenitor zone in adult liver, exocrine pancreas and intestine. Nat Genet. 2011;43(1):34–41. doi: 10.1038/ng.722. [DOI] [PubMed] [Google Scholar]
- 55.Tee LB, Kirilak Y, Huang WH, Morgan RH, Yeoh GC. Differentiation of oval cells into duct-like cells in preneoplastic liver of rats placed on a choline-deficient diet supplemented with ethionine. Carcinogenesis. 1994;15(12):2747–2756. doi: 10.1093/carcin/15.12.2747. [DOI] [PubMed] [Google Scholar]
- 56.Akhurst B, et al. A modified choline-deficient, ethionine-supplemented diet protocol effectively induces oval cells in mouse liver. Hepatology. 2001;34(3):519–522. doi: 10.1053/jhep.2001.26751. [DOI] [PubMed] [Google Scholar]
- 57.Cherry SR, Biniszkiewicz D, van Parijs L, Baltimore D, Jaenisch R. Retroviral expression in embryonic stem cells and hematopoietic stem cells. Mol Cell Biol. 2000;20(20):7419–7426. doi: 10.1128/MCB.20.20.7419-7426.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.He J, Yang Q, Chang LJ. Dynamic DNA methylation and histone modifications contribute to lentiviral transgene silencing in murine embryonic carcinoma cells. J Virol. 2005;79(21):13497–13508. doi: 10.1128/JVI.79.21.13497-13508.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Harting MT, Jimenez F, Cox CS., Jr Isolation of mesenchymal stem cells (MSCs) from green fluorescent protein positive (GFP+) transgenic rodents: the grass is not always green(er). Stem Cells Dev. 2009;18(1):127–135. doi: 10.1089/scd.2008.0046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Li F, et al. Generation of a novel transgenic mouse model for bioluminescent monitoring of survivin gene activity in vivo at various pathophysiological processes: survivin expression overlaps with stem cell markers. Am J Pathol. 2010;176(4):1629–1638. doi: 10.2353/ajpath.2010.090414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Schmelzer E, et al. Human hepatic stem cells from fetal and postnatal donors. J Exp Med. 2007;204(8):1973–1987. doi: 10.1084/jem.20061603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.de Boer CJ, van Krieken JH, Janssen-van Rhijn CM, Litvinov SV. Expression of Ep-CAM in normal, regenerating, metaplastic, and neoplastic liver. J Pathol. 1999;188(2):201–206. doi: 10.1002/(SICI)1096-9896(199906)188:2<201::AID-PATH339>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- 63.Clayton E, Forbes SJ. The isolation and in vitro expansion of hepatic Sca-1 progenitor cells. Biochem Biophys Res Commun. 2009;381(4):549–553. doi: 10.1016/j.bbrc.2009.02.079. [DOI] [PubMed] [Google Scholar]
- 64.Lorenzini S, et al. Characterisation of a stereotypical cellular and extracellular adult liver progenitor cell niche in rodents and diseased human liver. Gut. 2010;59(5):645–654. doi: 10.1136/gut.2009.182345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Shibasaki S, Aoki T, Yoshimi T. Liver Innervation: And The Neural Control Of Hepatic Function. Electronmicroscopic studies on the innervation in human liver. In: Shimatzu T, ed. London, United Kingdom; John Libby & Company Ltd.; 1997:57–65. [Google Scholar]
- 66.Paku S, Schnur J, Nagy P, Thorgeirsson SS. Origin and structural evolution of the early proliferating oval cells in rat liver. Am J Pathol. 2001;158(4):1313–1323. doi: 10.1016/S0002-9440(10)64082-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Evarts RP, Hu Z, Fujio K, Marsden ER, Thorgeirsson SS. Activation of hepatic stem cell compartment in the rat: role of transforming growth factor alpha, hepatocyte growth factor, and acidic fibroblast growth factor in early proliferation. Cell Growth Differ. 1993;4(7):555–561. [PubMed] [Google Scholar]
- 68.Hu Z, Evarts RP, Fujio K, Marsden ER, Thorgeirsson SS. Expression of hepatocyte growth factor and c-met genes during hepatic differentiation and liver development in the rat. Am J Pathol. 1993;142(6):1823–1830. [PMC free article] [PubMed] [Google Scholar]
- 69.Terada R, et al. Stromal cell-derived factor-1 from biliary epithelial cells recruits CXCR4-positive cells: implications for inflammatory liver diseases. Lab Invest. 2003;83(5):665–672. doi: 10.1097/01.lab.0000067498.89585.06. [DOI] [PubMed] [Google Scholar]
- 70.Kubota H, Yao HL, Reid LM. Identification and characterization of vitamin A-storing cells in fetal liver: implications for functional importance of hepatic stellate cells in liver development and hematopoiesis. Stem Cells. 2007;25(9):2339–2349. doi: 10.1634/stemcells.2006-0316. [DOI] [PubMed] [Google Scholar]
- 71.Hong F, et al. Hepatic stellate cells express functional CXCR4: role in stromal cell-derived factor-1alpha-mediated stellate cell activation. Hepatology. 2009;49(6):2055–2067. doi: 10.1002/hep.22890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sawitza I, Kordes C, Reister S, Häussinger D. The niche of stellate cells within rat liver. Hepatology. 2009;50(5):1617–1624. doi: 10.1002/hep.23184. [DOI] [PubMed] [Google Scholar]
- 73.Pintilie DG, Shupe TD, Oh SH, Salganik SV, Darwiche H, Petersen BE. Hepatic stellate cells’ involvement in progenitor-mediated liver regeneration. Lab Invest. 2010;90(8):1199–1208. doi: 10.1038/labinvest.2010.88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Wang Y, et al. Paracrine signals from mesenchymal cell populations govern the expansion and differentiation of human hepatic stem cells to adult liver fates. Hepatology. 2010;52(4):1443–1454. doi: 10.1002/hep.23829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Schirmacher P, Geerts A, Peitrangelo A, Dienes HP, Rogler CE. Hepatocyte growth factor/hepatopoietin A is expressed in fat-storing cells from rat liver but not myofibroblast-like cells derived from fat-storing cells. Hepatology. 1991;15(1):5–11. doi: 10.1002/hep.1840150103. [DOI] [PubMed] [Google Scholar]
- 76.Ishikawa T, et al. Hepatocyte growth factor/c-met signaling is required for stem-cell-mediated liver regeneration in mice. Hepatology. 2012;55(4):1215–1226. doi: 10.1002/hep.24796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Sicklick JK, et al. Hedgehog signaling maintains resident hepatic progenitors throughout life. Am J Physiol Gastrointest Liver Physiol. 2006;290(5):G859–G870. doi: 10.1152/ajpgi.00456.2005. [DOI] [PubMed] [Google Scholar]
- 78.Jung Y, et al. Signals from dying hepatocytes trigger growth of liver progenitors. Gut. 2010;59(5):655–665. doi: 10.1136/gut.2009.204354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Berthoud HR, Kressel M, Neuhuber WL. An anterograde tracing study of the vagal innervation of rat liver, portal vein and biliary system. Anat Embryol (Berl). 1992;186(5):431–442. doi: 10.1007/BF00185458. [DOI] [PubMed] [Google Scholar]
- 80.Oben JA, Diehl AM. Sympathetic nervous system regulation of liver repair. Anat Rec A Discov Mol Cell Evol Biol. 2004;280(1):874–883. doi: 10.1002/ar.a.20081. [DOI] [PubMed] [Google Scholar]
- 81.Cassiman D, Libbrecht L, Sinelli N, Desmet V, Denef C, Roskams T. The vagal nerve stimulates activation of the hepatic progenitor cell compartment via muscarinic acetylcholine receptor type 3. Am J Pathol. 2002;161(2):521–530. doi: 10.1016/S0002-9440(10)64208-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ayres-Silva Jde P, Manso PP, Madeira MR, Pelajo-Machado M, Lenzi HL. Sequential morphological characteristics of murine fetal liver hematopoietic microenvironment in Swiss Webster mice. Cell Tissue Res. 2011;344(3):455–469. doi: 10.1007/s00441-011-1170-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Jordan CT, McKearn JP, Lemischka IR. Cellular and developmental properties of fetal hematopoietic stem cells. Cell. 1990;61(6):953–963. doi: 10.1016/0092-8674(90)90061-I. [DOI] [PubMed] [Google Scholar]
- 84.Kim CH. Homeostatic and pathogenic extramedullary hematopoiesis. J Blood Med. 2010;1:13–19. doi: 10.2147/JBM.S7224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Friedman SL, Roll FJ, Boyles J, Bissell DM. Hepatic lipocytes: the principal collagen-producing cells of normal rat liver. Proc Natl Acad Sci U S A. 1985;82(24):8681–8685. doi: 10.1073/pnas.82.24.8681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Kordes C, et al. CD133+ hepatic stellate cells are progenitor cells. . Biochem Biophys Res Commun. 2007;352(2):410–417. doi: 10.1016/j.bbrc.2006.11.029. [DOI] [PubMed] [Google Scholar]
- 87.Sicklick JK, et al. Evidence for epithelial-mesenchymal transitions in adult liver cells. Am J Physiol Gastrointest Liver Physiol. 2006;291(4):G575–G583. doi: 10.1152/ajpgi.00102.2006. [DOI] [PubMed] [Google Scholar]
- 88.Yang L, et al. Fate-mapping evidence that hepatic stellate cells are epithelial progenitors in adult mouse livers. Stem Cells. 2008;26(8):2104–2113. doi: 10.1634/stemcells.2008-0115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Covas DT, et al. Multipotent mesenchymal stromal cells obtained from diverse human tissues share functional properties and gene-expression profile with CD146+ perivascular cells and fibroblasts. . Exp Hematol. 2008;36(5):642–654. doi: 10.1016/j.exphem.2007.12.015. [DOI] [PubMed] [Google Scholar]
- 90.Mato E, Lucas M, Petriz J, Gomis R, Novials A. Identification of a pancreatic stellate cell population with properties of progenitor cells: new role for stellate cells in the pancreas. Biochem J. 2009;421(2):181–191. doi: 10.1042/BJ20081466. [DOI] [PubMed] [Google Scholar]
- 91.Castilho-Fernandes A, et al. Human hepatic stellate cell line (LX-2) exhibits characteristics of bone marrow-derived mesenchymal stem cells. Exp Mol Pathol. 2011;91(3):664–672. doi: 10.1016/j.yexmp.2011.09.002. [DOI] [PubMed] [Google Scholar]
- 92.Kordes C, Sawitza I, Götze S, Häussinger D. Stellate cells from rat pancreas are stem cells and can contribute to liver regeneration. PLoS One. 2012;7(12):e51878. doi: 10.1371/journal.pone.0051878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Niki T, et al. Class VI intermediate filament protein nestin is induced during activation of rat hepatic stellate cells. Hepatology. 1999;29(2):520–527. doi: 10.1002/hep.510290232. [DOI] [PubMed] [Google Scholar]
- 94.Lardon J, Rooman I, Bouwens L. Nestin expression in pancreatic stellate cells and angiogenic endothelial cells. Histochem Cell Biol. 2001;117(6):535–540. doi: 10.1007/s00418-002-0412-4. [DOI] [PubMed] [Google Scholar]
- 95.Tanaka S, Takasawa A, Fukasawa Y, Hasegawa T, Sawada N. An undifferentiated embryonal sarcoma of the liver containing adipophilin-positive vesicles in an adult with massive sinusoidal invasion. Int J Clin Exp Pathol. 2012;5(8):824–829. [PMC free article] [PubMed] [Google Scholar]
- 96.Reinehr R, Sommerfeld A, Häussinger D. CD95 ligand is a proliferative and antiapoptotic signal in quiescent hepatic stellate cells. Gastroenterology. 2008;134(5):1494–1506. doi: 10.1053/j.gastro.2008.02.021. [DOI] [PubMed] [Google Scholar]
- 97.Sommerfeld A, Reinehr R, Häussinger D. Bile acid-induced epidermal growth factor receptor activation in quiescent rat hepatic stellate cells can trigger both proliferation and apoptosis. J Biol Chem. 2009;284(33):22173–22183. doi: 10.1074/jbc.M109.005355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Sato Y, et al. Human mesenchymal stem cells xenografted directly to rat liver are differentiated into human hepatocytes without fusion. Blood. 2005;106(2):756–763. doi: 10.1182/blood-2005-02-0572. [DOI] [PubMed] [Google Scholar]
- 99.Chamberlain J, et al. Efficient generation of human hepatocytes by the intrahepatic delivery of clonal human mesenchymal stem cells in fetal sheep. Hepatology. 2007;46(6):1935–1945. doi: 10.1002/hep.21899. [DOI] [PubMed] [Google Scholar]
- 100.Aurich I, et al. Functional integration of hepatocytes derived from human mesenchymal stem cells into mouse livers. Gut. 2007;56(3):405–415. doi: 10.1136/gut.2005.090050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Crisan M, et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell. 2008;3(3):301–313. doi: 10.1016/j.stem.2008.07.003. [DOI] [PubMed] [Google Scholar]
- 102.Kiassov AP, et al. Desmin expressing nonhematopoietic liver cells during rat liver development: an immunohistochemical and morphometric study. Differentiation. 1995;59(4):253–258. doi: 10.1046/j.1432-0436.1995.5940253.x. [DOI] [PubMed] [Google Scholar]
- 103.Kordes C, Sawitza I, Götze S, Häussinger D. Hepatic stellate cells support hematopoiesis and are liver-resident mesenchymal stem cells. Cell Physiol Biochem. 2013;31:290–304. doi: 10.1159/000343368. [DOI] [PubMed] [Google Scholar]
- 104.Caplan AI, Dennis JE. Mesenchymal stem cells as trophic mediators. J Cell Biochem. 2006;98(5):1076–1084. doi: 10.1002/jcb.20886. [DOI] [PubMed] [Google Scholar]
- 105.Friedman SL, Roll FJ, Boyles J, Arenson DM, Bissel DM. Maintenance of differentiated phenotype of cultured rat hepatic lipocytes by basement membrane matrix. J Biol Chem. 1989;264(18):10756–10762. [PubMed] [Google Scholar]
- 106.Kisseleva T, et al. Myofibroblasts revert to an inactive phenotype during regression of liver fibrosis. Proc Natl Acad Sci U S A. 2012;109(24):9448–9453. doi: 10.1073/pnas.1201840109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Troeger JS, et al. Deactivation of hepatic stellate cells during liver fibrosis resolution in mice. Gastroenterology. 2012;143(4):1073–1083. doi: 10.1053/j.gastro.2012.06.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Kordes C, Sawitza I, Häussinger D. Canonical Wnt signaling maintains the quiescent stage of hepatic stellate cells. Biochem Biophys Res Commun. 2008;367(1):116–123. doi: 10.1016/j.bbrc.2007.12.085. [DOI] [PubMed] [Google Scholar]
- 109.Jensen CH, et al. Transit-amplifying ductular (oval) cells and their hepatocytic progeny are characterized by a novel and distinctive expression of delta-like protein/preadipocyte factor 1/fetal antigen 1. Am J Pathol. 2004;164(4):1347–1359. doi: 10.1016/S0002-9440(10)63221-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Köhler C, Bell AW, Bowen WC, Monga SP, Fleig W, Michalopoulos GK. Expression of Notch-1 and its ligand Jagged-1 in rat liver during liver regeneration. Hepatology. 2004;39(4):1056–1065. doi: 10.1002/hep.20156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.McCuskey RS. Anatomy of efferent hepatic nerves. Anat Rec A Discov Mol Cell Evol Biol. 2004;280(1):821–826. doi: 10.1002/ar.a.20087. [DOI] [PubMed] [Google Scholar]
- 112.vom Dahl S, Bode JG, Reinehr R, Mönnighoff I, Kubitz R, Häussinger D. Release of osmolytes from perfused rat liver on perivascular nerve stimulation: alpha-adrenergic control of osmolyte efflux from parenchymal and nonparenchymal liver cells. Hepatology. 1999;29(1):195–204. doi: 10.1002/hep.510290114. [DOI] [PubMed] [Google Scholar]
- 113.Reinehr RM, Kubitz R, Peters-Regehr T, Bode JG, Häussinger D. Activation of rat hepatic stellate cells in culture is associated with increased sensitivity to endothelin 1. Hepatology. 1998;28(6):1566–1577. doi: 10.1002/hep.510280617. [DOI] [PubMed] [Google Scholar]
- 114.Häussinger D, Stehle T, Gerok W, Sies H. Perivascular nerve stimulation and phenylephrine responses in rat liver. Metabolic effects, Ca2+ and K+ fluxes. . Eur J Biochem. 1987;163(1):197–203. doi: 10.1111/j.1432-1033.1987.tb10755.x. [DOI] [PubMed] [Google Scholar]
- 115.Athari A, Hänecke K, Jungermann K. Prostaglandin F2alpha and D2 release from primary Ito cell cultures after stimulation with noradrenaline and ATP but not adenosine. Hepatology. 1994;20(1 pt 1):142–148. doi: 10.1016/0270-9139(94)90146-5. [DOI] [PubMed] [Google Scholar]
- 116.Turner R, et al. Human hepatic stem cell and maturational liver lineage biology. Hepatology. 2011;53(3):1035–1045. doi: 10.1002/hep.24157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Huang W, et al. Nuclear receptor-dependent bile acid signaling is required for normal liver regeneration. Science. 2006;312(5771):233–236. doi: 10.1126/science.1121435. [DOI] [PubMed] [Google Scholar]
- 118.Terada R, et al. Stromal cell-derived factor-1 from biliary epithelial cells recruits CXCR4-positive cells: implications for inflammatory liver diseases. Lab Invest. 2003;83(5):665–672. doi: 10.1097/01.lab.0000067498.89585.06. [DOI] [PubMed] [Google Scholar]
- 119.Matsumoto S, et al. Immunohistochemical study on phenotypical changes of hepatocytes in liver disease with reference to extracellular matrix composition. Liver. 1999;19(1):32–38. doi: 10.1111/j.1478-3231.1999.tb00006.x. [DOI] [PubMed] [Google Scholar]
- 120.Mayack SR, Shadrach JL, Kim FS, Wagers AJ. Systemic signals regulate ageing and rejuvenation of blood stem cell niches. Nature. 2010;463(7280):495–500. doi: 10.1038/nature08749. [DOI] [PubMed] [Google Scholar]