

Abstract
Cells communicate with neighboring cells and condition their local environment by secreting soluble factors into the extracellular space. These well-studied facets of cell biology are essential for the establishment and maintenance of physiological homeostasis. However, accumulating evidence has revealed that specific ligands, enzymes, and macromolecules are distributed into the extracellular space by virtue of their association with small vesicles, which are released by a variety of cell types. Although the biological significance of such vesicles was initially debated, purification and subsequent functional studies have shown that these extracellular vesicles are bioactive organelles carrying a wide range of protein and nucleic acid cargoes. In many cases these vesicles are laden with molecules that are involved in cell signaling, although other diverse functions are being revealed at a rapid pace. In this Perspective, we discuss recent developments in the understanding of the major pathways of extracellular vesicle biogenesis and how these vesicles contribute to the maintenance of physiological homeostasis.
INTRODUCTION
Cells play an active role in shaping their local environment by releasing factors that either affect neighboring cells or manipulate the biochemical properties of the extracellular milieu. Although soluble protein ligands have received the most experimental attention in this regard, a rapidly growing field of investigation suggests that these events are also mediated by small extracellular vesicles (ECVs). Indeed, ECVs are now known to affect processes ranging from immune signaling to angiogenesis to detoxification of bacterial products (Thery et al., 2009; Mause and Weber, 2010; Shifrin et al., 2012). In these cases and many others, ECVs facilitate the distribution of specific cargoes that mediate communication with or coercion of other cells or the conditioning of the extracellular environment. ECVs range in size from ∼50 to 500 nm and are typically enriched in specific proteins and lipids, which differentiates their composition from that of the plasma membrane (Mause and Weber, 2010). One challenge for investigators new to this field is the variable and confusing nomenclature that exists in the ECV literature; this is primarily a result of the many cellular sources, isolation procedures, and apparently distinct mechanisms of formation. For the purposes of this discussion, we group ECVs into two broad categories that are distinguished by their mechanism of formation: exosomes and ectosomes. Exosomes (40–100 nm) are likely produced through an exocytic pathway via the formation of multivesicular bodies (MVBs) and their subsequent fusion with the plasma membrane (Mathivanan et al., 2010). In contrast, ectosomes range from 100 to 500 nm in diameter and are released through outward budding of the plasma membrane. Other names for ectosomes one might encounter in the literature are “microvesicles,” “membrane particles,” “microparticles,” or “nanoparticles” (Bastida et al., 1984; Thery et al., 2001; Cocucci et al., 2009; Mathivanan et al., 2010; van der Pol et al., 2012). Both exosomes and ectosomes are formed such that the original plasma membrane topology is maintained, that is, vesicles are released into the extracellular space with membranes “right-side out” (van der Pol et al., 2012). Exosomes and ectosomes are produced by a vast array of cell types in a variety of physiological contexts, suggesting that they may contribute to many essential aspects of cell and tissue function, some of which will be discussed in more detail in what follows.
MECHANISMS OF ECV FORMATION
Exosome biogenesis
Although the molecular details underlying exosome production remained unclear for many years, recent models have converged on a late-endosomal, MVB-dependent pathway (de Gassart et al., 2004; Hurley and Odorizzi, 2012). How specific cargoes destined for exosomes sort into these membranes and how differentiated vesicles bud into the lumen of the MVB are still matters of debate. An initial lipid-centric model suggested that ceramide enrichment in endosomal membranes was a major driving force (Trajkovic et al., 2008). Because of the small size of its head group, ceramide may directly influence the shape of the membrane to promote bending and downstream budding of vesicles. This model also invokes ceramide in the formation of raft-like domains that presumably function to sort specific cargo molecules into exosomal membranes, although this point has not been tested directly. A second model that has gained support more recently suggests that exosome formation follows a mechanism that parallels the endolysomal degradation pathway, which relies on the function of endosomal sorting complex required for transport (ESCRT) machinery to generate MVBs (van Niel et al., 2006). Indeed, RNA interference silencing of the key ESCRT components tumor susceptibility gene 101 (TSG101, ESCRT-I), vacuolar protein sorting (VPS) 22 (ESCRT-II), charged multivesicular body protein (CHMP) 2A, CHMP4A/B/C (ESCRT-III), VPS4A/B, or ALIX (an ESCRT-III–interacting protein) significantly reduces exosome production in MCF-7 cells (Baietti et al., 2012). Other studies reported decreased exosome secretion from dendritic cells after silencing of the ESCRT-0 protein hepatocyte growth factor–regulated tyrosine kinase substrate (Hrs; Tamai et al., 2010). Cargoes destined for endolysomal degradation are selectively recruited to the limiting membrane of the MVB through a process that involves their ubiquitylation and subsequent ubiquitin-dependent interactions with ESCRT-0, -I, and -II (Raiborg and Stenmark, 2009). However, ALIX can bind directly to exosomal cargo molecules, which might represent a ubiquitin-independent recruitment mechanism and also provide a point of distinction between sorting into exosomes versus endolysosomes (Baietti et al., 2012; Hurley and Odorizzi, 2012). Although the extent of functional overlap between the ceramide-dependent and ESCRT-dependent pathways outlined here is unclear, the mechanistic details will likely become the focus of future studies.
After MVB formation, a subset of these organelles fuse with the plasma membrane and release their constituent exosomes (Thery et al., 2002). Although little is known about the direct regulation of MVB fusion with the plasma membrane, it appears that several Rab GTPases, including Rab5, Rab27, and Rab35, are involved in exosome secretion (Vanlandingham and Ceresa, 2009; Hsu et al., 2010; Ostrowski et al., 2010; Baietti et al., 2012). Further investigation will be needed to clarify the role of various Rab proteins in MVB fusion with the plasma membrane and exosome release.
Ectosome biogenesis
Although comparatively little is known about the details of ectosome formation, evidence suggests that ectosomes are produced not from an endomembrane system as for exosomes, but by direct budding from the plasma membrane (Thery et al., 2009). Despite this seemingly significant mechanistic distinction, proposed models of ectosome formation include elements that are common to the production of exosomes. For example, the topology of outward budding from the plasma membrane is equivalent to budding into the MVB lumen. Thus one might expect the ESCRT system to be involved in ectosome formation. Indeed, recent studies revealed that arrestin-domain containing-1 (ARRDC1) interacts with the ESCRT component TSG101 during ectosome shedding directly from the plasma membrane (Nabhan et al., 2012). Another parallel with exosome production is related to the significance of specific lipids in vesicle budding from the plasma membrane. ECVs of both types are enriched in cholesterol, ceramide, and other lipids implicated in raft formation. In glial cells, for example, production of ceramide is believed to promote membrane curvature during ectosome formation (Bianco et al., 2009).
An alternative pathway for ectosome formation might involve actin-based motors. In the context of the enterocyte brush border, the membrane-binding motor, myosin-1a (Myo1a), distributes along the length of the microvillus, where it exerts plus end (tipward)–directed force on the apical membrane (McConnell and Tyska, 2007). This activity leads to the accumulation of membrane at microvillar tips, which in turn drives the formation and release of ectosomes into the gut lumen (80–100 nm in diameter). Mice lacking Myo1a produce fewer ectosomes, and those that are released lack characteristic enrichment of cargo molecules, such as the host defense factor intestinal alkaline phosphatase (McConnell et al., 2009). Although the involvement of Myo1a in this system is likely an adaptation linked to the unique morphology of the microvillus, these studies suggest that manipulation of plasma membrane mechanics is of central importance to the mechanism of ectosome formation.
PHYSIOLOGICAL ROLES OF ECVS
ECVs are released from a variety of “donor” cell types and contribute to an array of physiological processes. In this section, our discussion of ECV function is organized according to physiological context; key examples and relevant references are highlighted in Table 1.
TABLE 1:
Physiological roles of ECVs.
| Category | Location/source | Function | Cargo | Referencea |
|---|---|---|---|---|
| Immunity | Enterocyte | Anti-inflammatory, antibacterial | Intestinal alkaline phosphatase | Shifrin et al. (2012) |
| Macrophage, mast cell, B cell, dendritic cell | Immune cell activation, antigen presentation, MHC transfer | IL-1β, TNF-α, HMGB1 | Thery et al. (2009); Lo Cicero and Raposo, (2013)* | |
| T-cell, polymorphonuclear cell | Immune modulation | FasL | van der Pol et al. (2012)* | |
| Microglial cell | Immunostimulation, increased neurotransmission | IL-1β | Turola et al. (2012)* | |
| Mono/lymphocyte | Promotion of Trypanosoma cruzi infection | TGF-β | Cestari et al. (2012) | |
| Syncytiotrophoblast | Immunosuppression | CD63, PLAP, TGF-β, FasL, ULBP, mRNAs, miRNAs | Mincheva-Nilsson and Baranov (2010)* | |
| Dendritic cell | TH1 response activation, natural killer activation, anti-inflammatory | MHC proteins, antigens, T-cell stimulatory molecules | Zitvogel et al. (1998), Thery et al. (2001), van der Pol et al. (2012)* | |
| Development | Stem cell | Tissue repair, plasticity, reprogramming | mRNA, miRNA | Camussi et al. (2011), Tetta et al. (2011)* |
| Cardiomyocyte in culture | Gene transfer, gene expression changes | mRNA, chromosomal DNA | Waldenstrom et al. (2012) | |
| Endothelial progenitor cell | Angiogenesis | mRNA | Mause and Weber (2010)* | |
| Lung tissue | Fate determination, marrow cell phenotype | mRNA | Aliotta et al. (2007, 2010, 2012*) | |
| Experimentally induced in HEK293 | Confer permissiveness for induced pluripotent stem cell generation | mCAT-1 | Mangeot et al. (2011) | |
| Neurological | Microglial cell | Immunostimulation, increased neurotransmission | IL-1β | Turola et al. (2012)* |
| Primary neuron, especially somatodendritic compartment | Synaptic transmission and plasticity, neurodegeneration | GluR2 | Schneider and Simons (2013) | |
| Bone | Epiphysis | Cartilage calcification | TNAP | Golub (2009, 2011)* |
| Blood/cardiovascular | Platelets | Thrombus formation, immune cell activation, angiogenesis/neovascularization | Tissue factor, chemokines, IL-1β, signaling lipids | von Hundelshausen and Weber (2007), Mause and Weber (2010), Owens and Mackman (2011), Rautou et al. (2011), Zwicker et al. (2011)* |
| Endothelial cell | Might promote atherosclerosis | Rautou et al. (2011)* | ||
| Smooth muscle cells (aorta) | Might promote atherosclerosis | Bobryshev et al. (2012) | ||
| Cancer | MDA-MB-231 | Increase tumor cell invasion | Amphiregulin | Higginbotham et al. (2011) |
| Most cancer cell types | Increased invasiveness, antitumor immunosuppression, possibly T-cell priming | Oncogenic sequences, single-strand DNA, retrotransposons, tumor antigens, cytokines | D'Souza-Schorey and Clancy (2012)*, Iero et al. (2008)* | |
| Glioma cell | Transfer of EGFRvIII | EGFRvIII | Al-Nedawi et al. (2008) |
HMGB1, high-mobility group protein B1; mCAT-1, murine cationic amino acid transporter 1; MHC, major histocompatibility complex; miRNA, micro RNA; PLAP, placental alkaline phosphatase;TGF-β, transforming growth factor β; TNAP, tissue-nonspecific alkaline phosphatase; TNF-α, tumor necrosis factor α; ULBP, UL16-binding protein.
aAsterisk indicates a review.
Development
Although relatively little is known about ECVs during vertebrate development, several reports provide clues as to how these vesicles might contribute to normal morphogenesis. ECVs are detected in the peripheral circulation of pregnant women, and ECV cargoes vary based on the phase of gestation (Abrahams et al., 2004; Frangsmyr et al., 2005; Atay et al., 2011; Clifton et al., 2012). Although it is unknown how ECVs act during gestation, studies have shown that proteins with established functions in development are found in ECVs. For example, β-catenin, Wnt, GSK3, and the Wnt cargo receptor Evi are components of exosomes purified from cultured human (HEK293, Caco-2) and Drosophila (S2,Kc167) cells (Chairoungdua et al., 2010; Taelman et al., 2010; Gross et al., 2012). The function of β-catenin– and Wnt-containing exosomes is not clear, but given the central role these proteins play in tissue and organ development (Petersen and Reddien, 2009), it is tempting to speculate that their function may be facilitated by distribution in ECVs. In addition, sonic hedgehog (SHH) is contained in ECVs from activating T lymphocytes, and these vesicles appear to function in promoting neovascularization (Soleti and Martinez, 2012). SHH has critical functions during embryogenesis, including patterning of the nervous system (Fuccillo et al., 2006) and development of the gastrointestinal system and other endoderm-derived organs (van den Brink, 2007). Thus, with high concentrations of the morphogen, it seems likely that SHH-containing ECVs could exert effects on developing organs and tissues. Taken together, the results of these studies are suggestive of developmental roles for ECVs, but future studies are warranted to clarify functional roles and molecular mechanisms.
Immunity
ECVs are shed from immune cells, with up-regulated production occurring upon stimulation by inflammatory signaling (van der Pol et al., 2012; Lo Cicero and Raposo, 2013). Conversely, infected cells can also release ECVs, which target immune cells and activate innate responses (Dreux et al., 2012). For example, hepatitis C–infected cells release viral RNA packaged inside exosomes (genetic material is a common ECV cargo), which are then taken up by plasmacytoid dendritic cells (Dreux et al., 2012). Moreover, ectosomes shed from intestinal epithelial cells are highly enriched in intestinal alkaline phosphatase and serve to both detoxify soluble bacterial lipopolysaccharide and inhibit bacterial colonization of the intestinal epithelial surface (Shifrin et al., 2012). Lumenal ECVs are shed constitutively, as they can be isolated from the small intestine of uninfected rodents and sterile cell culture models. However, ectosome production increases dramatically in the presence of bacterial pathogens (Shifrin et al., 2012). Similarly, ECV shedding from macrophages and microglial cells is dependent on activation of the ATP receptor and ligand-gated ion channel P2×7, which can be induced by exogenous ATP (e.g., as a result of cellular injury) or through normal ATP release into the extracellular environment of astrocyte–microglia cocultures (Bianco et al., 2005; Thomas and Salter, 2010). P2×7 can also be activated during sterile inflammation (Thomas and Salter, 2010). ATP released from astrocytes in culture is sufficient to stimulate ECV release from microglia (Bianco et al., 2005); these vesicles carry proinflammatory cytokines, including interleukin 1-β (IL1-β), and are capable of activating other macrophages and perhaps dendritic cells (Thomas and Salter, 2010). Activation of macrophages by ECVs suggests the initiation of a large-scale inflammatory response. However, it is possible that in some instances the targeted, local activation of ECV shedding through TLR4 signaling or other inflammatory mechanisms could actually prevent such a system-wide process. By releasing lumenal ECVs to combat bacterial attachment, for example, enterocytes could mitigate the spread of infection, thereby abrogating the need for mounting a systemic anti-inflammatory response (Shifrin et al., 2012). In the case of virally infected cells, host cells use ECVs to communicate with neighboring uninfected cells, which serves to initiate an immune response (Dreux et al., 2012). This allows an appropriate local-scale response without hyperactivating the immune system or further spreading the pathogen.
Bone formation
Matrix ECVs found in the epiphysis of long bones bear a strong resemblance to ECVs shed from enterocyte microvilli. Matrix ECVs are ectosomes released during the calcification process and are highly enriched in tissue-nonspecific alkaline phosphatase (Golub, 2009, 2011). This enzyme decreases the concentration of inorganic pyrophosphate, increases that of inorganic phosphate (Pi), and as a result promotes hydroxyapatite formation and bone mineralization (Golub, 2011). Although isolated from entirely different contexts, microvillus-derived ECVs and matrix ECVs share a common function in biochemically conditioning their local microenvironment. In the former case, dephosphorylation of lipopolysaccharide (LPS) by alkaline phosphatase significantly reduces LPS toxicity (Poelstra et al., 1997); in the latter, production of Pi is critical for proper assembly of normal bone. In both systems, however, the ability to produce ECVs with specific, enriched catalytic activity is needed for maintaining homeostasis.
Distribution of catalytic activity is not the only function of EVCs in bone; however, as Aliotta et al. (2007, 2010) discovered that marrow cell phenotype is altered by lung-derived ECVs through transfer of mRNA, aiding in recovery of lung tissue after injury. This suggests that ECVs may be involved in regulating stem cell fate decisions during tissue repair by regulating gene expression.
Cardiovascular function
In the cardiovascular system, circulating ECVs are released from virtually every cell type, including platelets, immune cells (van der Pol et al., 2012; Lo Cicero and Raposo, 2013), and endothelial and vascular smooth muscle cells. In some cases, these vesicles were shown to participate in the coagulation process (Cocucci et al., 2009), whereas other work revealed that cardiovascular ECVs can distribute HSP60 (Gupta and Knowlton, 2007). Waldenstrom et al. (2012) “transfected” fibroblasts with nucleic acid–containing ECVs isolated from adult mouse cardiomyocytes and demonstrated that genetic material from ECVs is transferred into target cells, where it induces changes in gene expression. Similarly, ECVs isolated from endothelial progenitor cells were able to stimulate vessel formation in culture and in an SCID mouse model, a process dependent on mRNA contained in the ECVs (Deregibus et al., 2007). In disease states, cardiovascular ECV production is upregulated, such as increased vesicle shedding observed in preatherosclerotic areas of the aorta (Bobryshev et al., 2012) and in cases of prothrombotic anemias (Zwicker et al., 2011). Bastida et al. (1984) described ECVs from isolated human glioblastoma cells that contain factors capable of activating clotting and platelet aggregation in vitro; this work is consistent with earlier findings of ECVs isolated from human breast cancer and guinea pig hepatocarcinoma cells (Dvorak et al., 1983). A notable feature of glioblastoma ECVs is their potent activity, showing up to 10-fold greater procoagulant activity compared with cellular membrane fractions. Similar to other systems, this finding demonstrates the utility of ECVs as distributors of a highly concentrated biological activity, allowing source cells to propagate the relevant signal in a targeted, efficient manner.
Nervous system function
In neural tissue, exosomes are released from many cell types (Turola et al., 2012), whereas ectosomes are shed by microglial cells (discussed earlier), neurons (Schiera et al., 2007), astrocytes (Bianco et al., 2005), and possibly oligodendrocytes (Turola et al., 2012; Verderio et al., 2012). Neuronal ECVs are enriched in fibroblast growth factor 2 (FGF2) and vascular endothelial growth factor (VEGF) and could function in regulating angiogenesis (Schiera et al., 2007). Astrocytic ECVs carry IL-1β, implicating the organelles in the mediation of immune responses, and an examination of oligodendrocyte-derived ECVs from patients with multiple sclerosis also suggests a role in proinflammatory signaling (Verderio et al., 2012). Faure et al. (2006) demonstrated the release of exosomes from neurons and astrocytes, which contained the GluR2 subunit of the glutamate receptor and other proteins. Although further work investigating the function of neuronal ECVs is needed, this and a subsequent study (Lachenal et al., 2011) suggest that neuronal-derived ECVs play a regulatory role in the CNS for neuronal plasticity, perhaps by recycling or disposing of various receptors or by transferring cargo proteins to receiving cells. ECVs were long believed to be a general cellular “trash disposal,” an idea that was mostly discarded as investigators discovered specific ECV functions. In the nervous system, however, removing cellular detritus is still considered a primary function of ECVs and one that may play a significant role in maintaining homeostasis by regulating receptors on the cell surface and removing pathology-inducing proteins targeted for degradation (Von Bartheld and Altick, 2011).
ECV FUNCTION IN DISEASE
Based on the broad contributions to normal physiology, it is perhaps not surprising that ECVs play roles in numerous diseases (atherosclerosis, coagulopathies, inflammation, infection and autoimmune disease; Thomas and Salter, 2010; Zwicker et al., 2011; Bobryshev et al., 2012; Dreux et al., 2012; Verderio et al., 2012). However, here we focus on their role in cancer.
A key finding that first implicated ECVs in cancer was the discovery that membrane vesicles shed from metastatic cells could induce lung metastasis (Poste and Nicolson, 1980). Early research on exosomes in the immune system (Chaput et al., 2004) led to their eventual identification as vesicles secreted from both mouse and human tumor cells (Wolfers et al., 2001; Andre et al., 2002). Although we do not fully understand the differences between ECV released from normal and cancer cells, recent work suggests that ECVs facilitate horizontal transfer of cargo and harness the activity of these cargoes to stimulate intracellular signaling pathways, promoting tumor progression (Muralidharan-Chari et al., 2010; Lee et al., 2011; Demory Beckler et al., 2013). Although we focus on protein cargo of ECVs in the subsequent discussion, it is becoming increasingly apparent that these vesicles contain DNA and RNA, which may also confer important proneoplastic effects (Valadi et al., 2007; Skog et al., 2008; Hong et al., 2009; Pisetsky et al., 2011).
Oncogenes represent an important class of active ECV cargoes that modulate recipient cells. Tissue transglutaminase (tTG)-containing ECVs purified from cancer cells induced NIH 3T3 cell survival and anchorage-independent growth, effects that were linked to transfer of tTG via ECVs. Moreover, exosomes purified from colon cancer cells transfer mutant G13D KRAS and induce anchorage-independent growth and colony formation in three-dimensional collagen matrix, further suggesting that oncogene-containing exosomes promote cancer-related phenotypes (Demory Beckler et al., 2013). In addition, treatment of mildly aggressive glioma cells with EGF receptor variant III (EGFRvIII)–containing ECVs increased observed levels of EGFRvIII in these cells, which may be partly responsible for the observed increases in VEGF production and anchorage-independent growth of recipient cells (Al-Nedawi et al., 2008). Not surprising, given the variety of cargo and roles in intercellular communication, it seems that in addition to promoting cancer phenotypes, ECVs also carry tumor suppressors. A recent study showed functional PTEN in exosomes purified from fibroblast cells and that treatment of cells with these exosomes reduced cellular proliferation (Putz et al., 2012). Combined, these data provide strong support for ECVs as vehicles for protein cargo transfer to recipient cells, allowing for promotion and suppression of tumor-related phenotypes.
In addition to inducing features of tumor initiation, data indicate a role for ECVs also in promoting angiogenesis, invasion, and metastasis. ECVs contain a wide variety of prometastatic cargo, including insulin-degrading enzyme, matrix metalloproteinases, tetraspanins, heat shock proteins, plasminogen, integrin family members, and growth factors (VEGF and FGF; Graner, 2011; D'Souza-Schorey and Clancy, 2012). Proangiogenic effects of ECVs include enhanced endothelial cell tubule formation and vascularization of in vivo Matrigel plugs (Hood et al., 2009; Mineo et al., 2012). Cathepsin B–containing ECVs and EGFR ligand–containing exosomes are two examples of ECVs that elicit recipient cell invasion in vitro (Giusti et al., 2008; Higginbotham et al., 2011). Luga et al., (2012) recently reported that exosomes secreted from fibroblasts enhance breast cancer cell motility and metastasis, a mechanism that was suggested to require exosome association with Wnt11 and activation of recipient breast cancer planar cell polarity proteins. This study provides additional support for the cross-talk of stroma and tumor and implicates stromal fibroblast ECVs in cancer progression. The identity of cargoes packaged in ECVs and their ability to induce invasion both support a role for these vesicles in promoting metastatic phenotypes.
Several key in vivo studies demonstrated the ability of ECVs to prime metastatic niches. Melanoma exosomes selectively home to and prime sentinel lymph nodes for melanoma cell metastasis (Hood et al., 2011). Similarly, pretreatment of mice with melanoma exosomes results in accelerated lung metastasis (Liu et al., 2010). Finally, it was recently reported that melanoma exosomes reprogram bone marrow–derived cells to a more provasculogenic phenotype that supports enhanced tumor growth and metastasis (Peinado et al., 2012). Taken together, these reports strongly indicate that ECVs act to enhance metastatic properties of recipient cells. Future studies are needed, however, to identify the mechanisms by which ECVs exert these actions.
COMMON THEMES AND CONCLUSIONS
Despite the diverse physiological contexts highlighted here, ECV functions generally partition into one of four categories: regulation of gene expression, induction of signaling, distribution of catalytic activity, and disposal of cellular “trash” (Figure 1). Even more broadly, ECVs serve to protect, concentrate, or remove cargo. Genetic material must be protected from degradation as it is transported from source to target cell. Ligands or receptors may need to be concentrated in order to efficiently activate a signaling cascade, whereas catalytic activity may be enhanced by concentrating an enzyme on the surface of an ECV. All of the studies highlighted here suggest that ECVs are highly adaptable cargo-carrying platforms.
FIGURE 1:
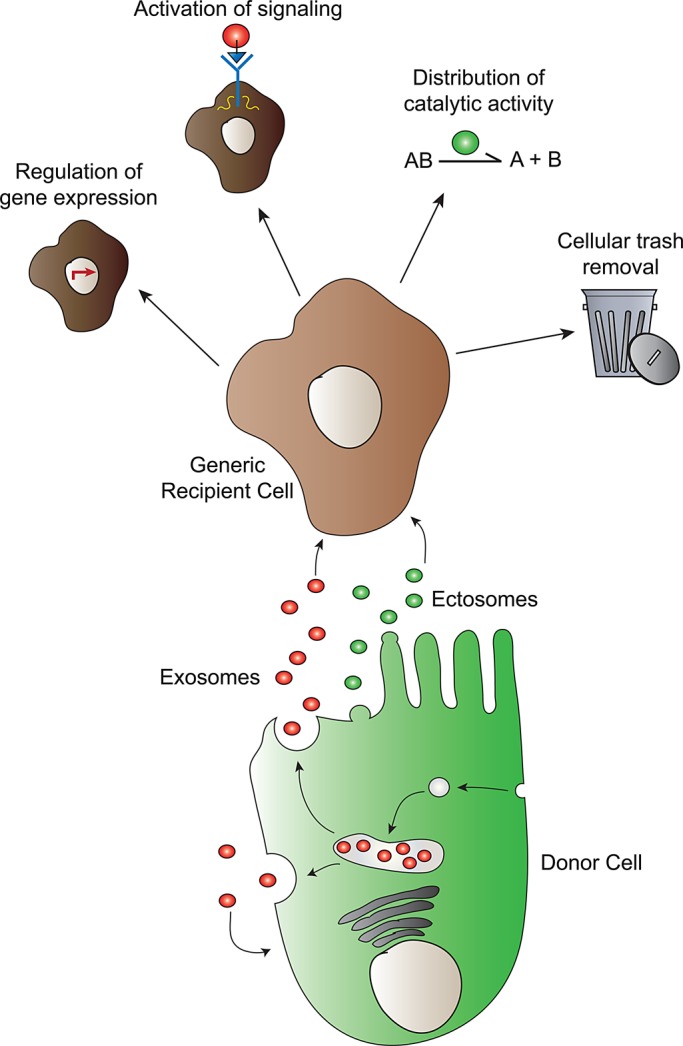
Extracellular vesicle biogenesis and function. Extracellular vesicles are formed via two known biogenesis pathways: exosomes via an endomembrane MVB intermediate, and ectosomes via direct budding from the plasma membrane. Once released into the extracellular environment, ECVs perform one of four general functions: 1) regulation of gene expression, 2) activation of cell signaling, 3) distribution of catalytic activity, and 4) cellular trash removal.
Given that discoveries from the last decade have demonstrated the near-ubiquity of ECVs, several new areas of investigation will be important for developing a full understanding of these organelles. More detailed studies of exosome and ectosome formation will provide much needed clarity into the distinctions between these two pathways. Moreover, very little is known about how ECVs are captured by recipient/target cells. Initial studies implicate specific lipids (e.g., phosphatidylserine) and integrins in this process, but the details are only beginning to come into focus (Deregibus et al., 2007; Iglesias et al., 2012). Once precise mechanisms of ECV tethering, fusion, and/or internalization with target cells have been established, understanding the transfer of enriched cargo molecules and downstream effects can be clarified.
Translational applications of ECV biology are in their infancy but include their use as therapeutic delivery vehicles and disease biomarkers. Recent studies showed that stem cell ECVs are able to reprogram diseased cells (Iglesias et al., 2012), and “custom” ECVs are capable of delivering experimental cargoes (Mangeot et al., 2011). Other potential ECV-based therapies, such as tissue repair and revascularization, are being investigated (Lee et al., 2012). ECVs also serve as attractive biomarkers of disease progression. In addition to blood and serum, ECVs can be purified from other biofluids, such as urine (Dear et al., 2013), and their cargo composition and rate of production are altered in various pathologies. However, a better understanding of ECV formation, uptake, and function is needed to expand current studies of ECVs as therapeutic delivery systems (Mangeot et al., 2011; Lee et al., 2012), tailor synthetic ECVs for treatment of disease, or use these organelles as biomarkers.
Acknowledgments
We thank members of the Tyska and Coffey laboratories for comments and advice. This work was supported by National Institutes of Health (NIH) Grants RO1-DK075555 (M.J.T.) and R56-DK095811 (M.J.T.), an American Heart Association Predoctoral Fellowship (D.A.S.), NIH Grants RO1-CA046413 and CA163563 (R.J.C.), a GI Special Program of Research Excellence P50-CA095103 (R.JC.), and NIH Grant R25-CA092043 (M.D.B.).
Abbreviations used:
- ECV
extracellular vesicle
- ESCRT
endosomal sorting complex required for transport
- LPS
lipopolysaccharide
- MVB
multivesicular body
- Myo1a
Myosin-1a
- PLAP
placental alkaline phosphatase
- SCID
severe combined immunodeficiency
- TNAP
tissue-nonspecific alkaline phosphatase
Footnotes
REFERENCES
- Abrahams VM, Straszewski-Chavez SL, Guller S, Mor G. First trimester trophoblast cells secrete Fas ligand which induces immune cell apoptosis. Mol Hum Reprod. 2004;10:55–63. doi: 10.1093/molehr/gah006. [DOI] [PubMed] [Google Scholar]
- Aliotta JM, Lee D, Puente N, Faradyan S, Sears EH, Amaral A, Goldberg L, Dooner MS, Pereira M, Quesenberry PJ. Progenitor/stem cell fate determination: interactive dynamics of cell cycle and microvesicles. Stem Cells Dev. 2012;21:1627–1638. doi: 10.1089/scd.2011.0550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aliotta JM, et al. Microvesicle entry into marrow cells mediates tissue-specific changes in mRNA by direct delivery of mRNA and induction of transcription. Exp Hematol. 2010;38:233–245. doi: 10.1016/j.exphem.2010.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aliotta JM, et al. Alteration of marrow cell gene expression, protein production, and engraftment into lung by lung-derived microvesicles: a novel mechanism for phenotype modulation. Stem Cells. 2007;25:2245–2256. doi: 10.1634/stemcells.2007-0128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al-Nedawi K, Meehan B, Micallef J, Lhotak V, May L, Guha A, Rak J. Intercellular transfer of the oncogenic receptor EGFRvIII by microvesicles derived from tumour cells. Nat Cell Biol. 2008;10:619–624. doi: 10.1038/ncb1725. [DOI] [PubMed] [Google Scholar]
- Andre F, et al. Malignant effusions and immunogenic tumour-derived exosomes. Lancet. 2002;360:295–305. doi: 10.1016/S0140-6736(02)09552-1. [DOI] [PubMed] [Google Scholar]
- Atay S, Gercel-Taylor C, Kesimer M, Taylor DD. Morphologic and proteomic characterization of exosomes released by cultured extravillous trophoblast cells. Exp Cell Res. 2011;317:1192–1202. doi: 10.1016/j.yexcr.2011.01.014. [DOI] [PubMed] [Google Scholar]
- Baietti MF, et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat Cell Biol. 2012;14:677–685. doi: 10.1038/ncb2502. [DOI] [PubMed] [Google Scholar]
- Bastida E, Ordinas A, Escolar G, Jamieson GA. Tissue factor in microvesicles shed from U87MG human glioblastoma cells induces coagulation, platelet aggregation, and thrombogenesis. Blood. 1984;64:177–184. [PubMed] [Google Scholar]
- Bianco F, et al. Acid sphingomyelinase activity triggers microparticle release from glial cells. EMBO J. 2009;28:1043–1054. doi: 10.1038/emboj.2009.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bianco F, Pravettoni E, Colombo A, Schenk U, Moller T, Matteoli M, Verderio C. Astrocyte-derived ATP induces vesicle shedding and IL-1 beta release from microglia. J Immunol. 2005;174:7268–7277. doi: 10.4049/jimmunol.174.11.7268. [DOI] [PubMed] [Google Scholar]
- Bobryshev YV, Killingsworth MC, Orekhov AN. Increased shedding of microvesicles from intimal smooth muscle cells in athero-prone areas of the human aorta: implications for understanding of the predisease stage. Pathobiology. 2012;80:24–31. doi: 10.1159/000339430. [DOI] [PubMed] [Google Scholar]
- Camussi G, Deregibus MC, Bruno S, Grange C, Fonsato V, Tetta C. Exosome/microvesicle-mediated epigenetic reprogramming of cells. Am J Cancer Res. 2011;1:98–110. [PMC free article] [PubMed] [Google Scholar]
- Cestari I, Ansa-Addo E, Deolindo P, Inal JM, Ramirez MI. Trypanosoma cruzi immune evasion mediated by host cell-derived microvesicles. J Immunol. 2012;188:1942–1952. doi: 10.4049/jimmunol.1102053. [DOI] [PubMed] [Google Scholar]
- Chairoungdua A, Smith DL, Pochard P, Hull M, Caplan MJ. Exosome release of beta-catenin: a novel mechanism that antagonizes Wnt signaling. J Cell Biol. 2010;190:1079–1091. doi: 10.1083/jcb.201002049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaput N, Taieb J, Schartz NE, Andre F, Angevin E, Zitvogel L. Exosome-based immunotherapy. Cancer Immunol Immunother. 2004;53:234–239. doi: 10.1007/s00262-003-0472-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clifton VL, Stark MJ, Osei-Kumah A, Hodyl NA. Review: the feto-placental unit, pregnancy pathology and impact on long term maternal health. Placenta. 2012;33(Suppl):S37–S41. doi: 10.1016/j.placenta.2011.11.005. [DOI] [PubMed] [Google Scholar]
- Cocucci E, Racchetti G, Meldolesi J. Shedding microvesicles: artefacts no more. Trends Cell Biol. 2009;19:43–51. doi: 10.1016/j.tcb.2008.11.003. [DOI] [PubMed] [Google Scholar]
- Dear JW, Street JM, Bailey MA. Urinary exosomes: a reservoir for biomarker discovery and potential mediators of intra-renal signaling. Proteomics. 2013 doi: 10.1002/pmic.201200285. DOI: 10.1002/pmic.201200285. [DOI] [PubMed] [Google Scholar]
- de Gassart A, Geminard C, Hoekstra D, Vidal M. Exosome secretion: the art of reutilizing nonrecycled proteins. Traffic. 2004;5:896–903. doi: 10.1111/j.1600-0854.2004.00223.x. [DOI] [PubMed] [Google Scholar]
- Demory Beckler M, Higginbotham JN, Franklin JL, Ham AJ, Halvey PJ, Imasuen IE, Whitwell C, Li M, Liebler DC, Coffey RJ. Proteomic analysis of exosomes from mutant KRAS colon cancer cells identifies intercellular transfer of mutant KRAS. Mol Cell Proteomics. 2013;12:343–355. doi: 10.1074/mcp.M112.022806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deregibus MC, Cantaluppi V, Calogero R, Lo Iacono M, Tetta C, Biancone L, Bruno S, Bussolati B, Camussi G. Endothelial progenitor cell derived microvesicles activate an angiogenic program in endothelial cells by a horizontal transfer of mRNA. Blood. 2007;110:2440–2448. doi: 10.1182/blood-2007-03-078709. [DOI] [PubMed] [Google Scholar]
- Dreux M, Garaigorta U, Boyd B, Decembre E, Chung J, Whitten-Bauer C, Wieland S, Chisari FV. Short-range exosomal transfer of viral RNA from infected cells to plasmacytoid dendritic cells triggers innate immunity. Cell Host Microbe. 2012;12:558–570. doi: 10.1016/j.chom.2012.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Souza-Schorey C, Clancy JW. Tumor-derived microvesicles: shedding light on novel microenvironment modulators and prospective cancer biomarkers. Genes Dev. 2012;26:1287–1299. doi: 10.1101/gad.192351.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dvorak HF, Van DeWater L, Bitzer AM, Dvorak AM, Anderson D, Harvey VS, Bach R, Davis GL, DeWolf W, Carvalho AC. Procoagulant activity associated with plasma membrane vesicles shed by cultured tumor cells. Cancer Res. 1983;43:4434–4442. [PubMed] [Google Scholar]
- Faure J, et al. Exosomes are released by cultured cortical neurones. Mol Cell Neurosci. 2006;31:642–648. doi: 10.1016/j.mcn.2005.12.003. [DOI] [PubMed] [Google Scholar]
- Frangsmyr L, Baranov V, Nagaeva O, Stendahl U, Kjellberg L, Mincheva-Nilsson L. Cytoplasmic microvesicular form of Fas ligand in human early placenta: switching the tissue immune privilege hypothesis from cellular to vesicular level. Mol Hum Reprod. 2005;11:35–41. doi: 10.1093/molehr/gah129. [DOI] [PubMed] [Google Scholar]
- Fuccillo M, Joyner AL, Fishell G. Morphogen to mitogen: the multiple roles of hedgehog signalling in vertebrate neural development. Nat Rev Neurosci. 2006;7:772–783. doi: 10.1038/nrn1990. [DOI] [PubMed] [Google Scholar]
- Giusti I, D'Ascenzo S, Millimaggi D, Taraboletti G, Carta G, Franceschini N, Pavan A, Dolo V. Cathepsin B mediates the pH-dependent proinvasive activity of tumor-shed microvesicles. Neoplasia. 2008;10:481–488. doi: 10.1593/neo.08178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golub EE. Role of matrix vesicles in biomineralization. Biochim Biophys Acta. 2009;1790:1592–1598. doi: 10.1016/j.bbagen.2009.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golub EE. Biomineralization and matrix vesicles in biology and pathology. Semin Immunopathol. 2011;33:409–417. doi: 10.1007/s00281-010-0230-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graner MW. Brain tumor exosomes and microvesicles: pleiotropic effects from tiny cellular surrogates. In: Garami M, editor. Molecular Targets of CNS Tumors. New York:: InTech; 2011. pp. 43–78. [Google Scholar]
- Gross JC, Chaudhary V, Bartscherer K, Boutros M. Active Wnt proteins are secreted on exosomes. Nat Cell Biol. 2012;14:1036–1045. doi: 10.1038/ncb2574. [DOI] [PubMed] [Google Scholar]
- Gupta S, Knowlton AA. HSP60 trafficking in adult cardiac myocytes: role of the exosomal pathway. Am J Physiol Heart Circ Physiol. 2007;292:H3052–H3056. doi: 10.1152/ajpheart.01355.2006. [DOI] [PubMed] [Google Scholar]
- Higginbotham JN, et al. Amphiregulin exosomes increase cancer cell invasion. Curr Biol. 2011;21:779–786. doi: 10.1016/j.cub.2011.03.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hong BS, et al. Colorectal cancer cell-derived microvesicles are enriched in cell cycle-related mRNAs that promote proliferation of endothelial cells. BMC Genomics. 2009;10:556. doi: 10.1186/1471-2164-10-556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hood JL, Pan H, Lanza GM, Wickline SA. Paracrine induction of endothelium by tumor exosomes. Lab Invest. 2009;89:1317–1328. doi: 10.1038/labinvest.2009.94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hood JL, San RS, Wickline SA. Exosomes released by melanoma cells prepare sentinel lymph nodes for tumor metastasis. Cancer Res. 2011;71:3792–3801. doi: 10.1158/0008-5472.CAN-10-4455. [DOI] [PubMed] [Google Scholar]
- Hsu C, et al. Regulation of exosome secretion by Rab35 and its GTPase-activating proteins TBC1D10A-C. J Cell Biol. 2010;189:223–232. doi: 10.1083/jcb.200911018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hurley JH, Odorizzi G. Get on the exosome bus with ALIX. Nat Cell Biol. 2012;14:654–655. doi: 10.1038/ncb2530. [DOI] [PubMed] [Google Scholar]
- Iero M, Valenti R, Huber V, Filipazzi P, Parmiani G, Fais S, Rivoltini L. Tumour-released exosomes and their implications in cancer immunity. Cell Death Differ. 2008;15:80–88. doi: 10.1038/sj.cdd.4402237. [DOI] [PubMed] [Google Scholar]
- Iglesias DM, et al. Stem cell microvesicles transfer cystinosin to human cystinotic cells and reduce cystine accumulation in vitro. PloS One. 2012;7:e42840. doi: 10.1371/journal.pone.0042840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lachenal G, Pernet-Gallay K, Chivet M, Hemming FJ, Belly A, Bodon G, Blot B, Haase G, Goldberg Y, Sadoul R. Release of exosomes from differentiated neurons and its regulation by synaptic glutamatergic activity. Mol Cell Neurosci. 2011;46:409–418. doi: 10.1016/j.mcn.2010.11.004. [DOI] [PubMed] [Google Scholar]
- Lee TH, D'Asti E, Magnus N, Al-Nedawi K, Meehan B, Rak J. Microvesicles as mediators of intercellular communication in cancer—the emerging science of cellular “debris.”. Semin Immunopathol. 2011;33:455–467. doi: 10.1007/s00281-011-0250-3. [DOI] [PubMed] [Google Scholar]
- Lee Y, El Andaloussi S, Wood MJ. Exosomes and microvesicles: extracellular vesicles for genetic information transfer and gene therapy. Hum Mol Genet. 2012;21:R125–R134. doi: 10.1093/hmg/dds317. [DOI] [PubMed] [Google Scholar]
- Liu Y, Xiang X, Zhuang X, Zhang S, Liu C, Cheng Z, Michalek S, Grizzle W, Zhang HG. Contribution of MyD88 to the tumor exosome-mediated induction of myeloid derived suppressor cells. Am J Pathol. 2010;176:2490–2499. doi: 10.2353/ajpath.2010.090777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lo Cicero A, Raposo G. Emerging concepts of tumor exosome–mediated cell-cell communication. In: Zhang H-G, editor. The Cell Biology of Exosomes: Historical and Perspectives. New York: Springer; 2013. pp. 1–32. [Google Scholar]
- Luga V, et al. Exosomes mediate stromal mobilization of autocrine Wnt-PCP signaling in breast cancer cell migration. Cell. 2012;151:1542–1556. doi: 10.1016/j.cell.2012.11.024. [DOI] [PubMed] [Google Scholar]
- Mangeot PE, Dollet S, Girard M, Ciancia C, Joly S, Peschanski M, Lotteau V. Protein transfer into human cells by VSV-G-induced nanovesicles. Mol Ther. 2011;19:1656–1666. doi: 10.1038/mt.2011.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mathivanan S, Ji H, Simpson RJ. Exosomes: extracellular organelles important in intercellular communication. J Proteomics. 2010;73:1907–1920. doi: 10.1016/j.jprot.2010.06.006. [DOI] [PubMed] [Google Scholar]
- Mause SF, Weber C. Microparticles: protagonists of a novel communication network for intercellular information exchange. Circ Res. 2010;107:1047–1057. doi: 10.1161/CIRCRESAHA.110.226456. [DOI] [PubMed] [Google Scholar]
- McConnell RE, Higginbotham JN, Shifrin DA, Jr, Tabb DL, Coffey RJ, Tyska MJ. The enterocyte microvillus is a vesicle-generating organelle. J Cell Biol. 2009;185:1285–1298. doi: 10.1083/jcb.200902147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McConnell RE, Tyska MJ. Myosin-1a powers the sliding of apical membrane along microvillar actin bundles. J Cell Biol. 2007;177:671–681. doi: 10.1083/jcb.200701144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mincheva-Nilsson L, Baranov V. The role of placental exosomes in reproduction. Am J Reprod Immunol. 2010;63:520–533. doi: 10.1111/j.1600-0897.2010.00822.x. [DOI] [PubMed] [Google Scholar]
- Mineo M, Garfield SH, Taverna S, Flugy A, De Leo G, Alessandro R, Kohn EC. Exosomes released by K562 chronic myeloid leukemia cells promote angiogenesis in a Src-dependent fashion. Angiogenesis. 2012;15:33–45. doi: 10.1007/s10456-011-9241-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muralidharan-Chari V, Clancy JW, Sedgwick A, D'Souza-Schorey C. Microvesicles: mediators of extracellular communication during cancer progression. J Cell Sci. 2010;123:1603–1611. doi: 10.1242/jcs.064386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nabhan JF, Hu R, Oh RS, Cohen SN, Lu Q. Formation and release of arrestin domain-containing protein 1-mediated microvesicles (ARMMs) at plasma membrane by recruitment of TSG101 protein. PNAS. 2012;109:4146–4151. doi: 10.1073/pnas.1200448109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ostrowski M, et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol. 2010;12:19–30. doi: 10.1038/ncb2000. supp 11–13. [DOI] [PubMed] [Google Scholar]
- Owens AP, Mackman N. Microparticles in hemostasis and thrombosis. Circ Res. 2011;108:1284–1297. doi: 10.1161/CIRCRESAHA.110.233056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peinado H, et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat Med. 2012;18:883–891. doi: 10.1038/nm.2753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petersen CP, Reddien PW. Wnt signaling and the polarity of the primary body axis. Cell. 2009;139:1056–1068. doi: 10.1016/j.cell.2009.11.035. [DOI] [PubMed] [Google Scholar]
- Pisetsky DS, Gauley J, Ullal AJ. Microparticles as a source of extracellular DNA. Immunol Res. 2011;49:227–234. doi: 10.1007/s12026-010-8184-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poelstra K, Bakker WW, Klok PA, Kamps JA, Hardonk MJ, Meijer DK. Dephosphorylation of endotoxin by alkaline phosphatase in vivo. Am J Pathol. 1997;151:1163–1169. [PMC free article] [PubMed] [Google Scholar]
- Poste G, Nicolson GL. Arrest and metastasis of blood-borne tumor cells are modified by fusion of plasma membrane vesicles from highly metastatic cells. Proc Natl Acad Sci USA. 1980;77:399–403. doi: 10.1073/pnas.77.1.399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Putz U, Howitt J, Doan A, Goh CP, Low LH, Silke J, Tan SS. The tumor suppressor PTEN is exported in exosomes and has phosphatase activity in recipient cells. Sci Signal. 2012;5:ra70. doi: 10.1126/scisignal.2003084. [DOI] [PubMed] [Google Scholar]
- Raiborg C, Stenmark H. The ESCRT machinery in endosomal sorting of ubiquitylated membrane proteins. Nature. 2009;458:445–452. doi: 10.1038/nature07961. [DOI] [PubMed] [Google Scholar]
- Rautou PE, Vion AC, Amabile N, Chironi G, Simon A, Tedgui A, Boulanger CM. Microparticles, vascular function, and atherothrombosis. Circ Res. 2011;109:593–606. doi: 10.1161/CIRCRESAHA.110.233163. [DOI] [PubMed] [Google Scholar]
- Schiera G, Proia P, Alberti C, Mineo M, Savettieri G, Di Liegro I. Neurons produce FGF2 and VEGF and secrete them at least in part by shedding extracellular vesicles. J Cell Mol Med. 2007;11:1384–1394. doi: 10.1111/j.1582-4934.2007.00100.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider A, Simons M. Exosomes: vesicular carriers for intercellular communication in neurodegenerative disorders. Cell Tissue Res. 2013;352:33–47. doi: 10.1007/s00441-012-1428-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shifrin DA, Jr, McConnell RE, Nambiar R, Higginbotham JN, Coffey RJ, Tyska MJ. Enterocyte microvillus-derived vesicles detoxify bacterial products and regulate epithelial-microbial interactions. Curr Biol. 2012;22:627–631. doi: 10.1016/j.cub.2012.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skog J, Wurdinger T, van Rijn S, Meijer DH, Gainche L, Sena-Esteves M, Curry WT, Jr, Carter BS, Krichevsky AM, Breakefield XO. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat Cell Biol. 2008;10:1470–1476. doi: 10.1038/ncb1800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soleti R, Martinez MC. Sonic Hedgehog on microparticles and neovascularization. Vitam Horm. 2012;88:395–438. doi: 10.1016/B978-0-12-394622-5.00018-3. [DOI] [PubMed] [Google Scholar]
- Taelman VF, Dobrowolski R, Plouhinec JL, Fuentealba LC, Vorwald PP, Gumper J, Sabatini DD, De Robertis EM. Wnt signaling requires sequestration of glycogen synthase kinase 3 inside multivesicular endosomes. Cell. 2010;143:1136–1148. doi: 10.1016/j.cell.2010.11.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamai K, et al. Exosome secretion of dendritic cells is regulated by Hrs, an ESCRT-0 protein. Biochem Biophys Res Commun. 2010;399:384–390. doi: 10.1016/j.bbrc.2010.07.083. [DOI] [PubMed] [Google Scholar]
- Tetta C, Bruno S, Fonsato V, Deregibus MC, Camussi G. The role of microvesicles in tissue repair. Organogenesis. 2011;7:105–115. doi: 10.4161/org.7.2.15782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thery C, Boussac M, Veron P, Ricciardi-Castagnoli P, Raposo G, Garin J, Amigorena S. Proteomic analysis of dendritic cell-derived exosomes: a secreted subcellular compartment distinct from apoptotic vesicles. J Immunol. 2001;166:7309–7318. doi: 10.4049/jimmunol.166.12.7309. [DOI] [PubMed] [Google Scholar]
- Thery C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses. Nat Rev Immunol. 2009;9:581–593. doi: 10.1038/nri2567. [DOI] [PubMed] [Google Scholar]
- Thery C, Zitvogel L, Amigorena S. Exosomes: composition, biogenesis and function. Nat Rev Immunol. 2002;2:569–579. doi: 10.1038/nri855. [DOI] [PubMed] [Google Scholar]
- Thomas LM, Salter RD. Activation of macrophages by P2×7-induced microvesicles from myeloid cells is mediated by phospholipids and is partially dependent on TLR4. J Immunol. 2010;185:3740–3749. doi: 10.4049/jimmunol.1001231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trajkovic K, Hsu C, Chiantia S, Rajendran L, Wenzel D, Wieland F, Schwille P, Brugger B, Simons M. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 2008;319:1244–1247. doi: 10.1126/science.1153124. [DOI] [PubMed] [Google Scholar]
- Turola E, Furlan R, Bianco F, Matteoli M, Verderio C. Microglial microvesicle secretion and intercellular signaling. Front Physiol. 2012;3:149. doi: 10.3389/fphys.2012.00149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valadi H, Ekstrom K, Bossios A, Sjostrand M, Lee JJ, Lotvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9:654–659. doi: 10.1038/ncb1596. [DOI] [PubMed] [Google Scholar]
- van den Brink GR. Hedgehog signaling in development and homeostasis of the gastrointestinal tract. Physiol Rev. 2007;87:1343–1375. doi: 10.1152/physrev.00054.2006. [DOI] [PubMed] [Google Scholar]
- van der Pol E, Boing AN, Harrison P, Sturk A, Nieuwland R. Classification, functions, and clinical relevance of extracellular vesicles. Pharmacol Rev. 2012;64:676–705. doi: 10.1124/pr.112.005983. [DOI] [PubMed] [Google Scholar]
- van Niel G, Porto-Carreiro I, Simoes S, Raposo G. Exosomes: a common pathway for a specialized function. J Biochem. 2006;140:13–21. doi: 10.1093/jb/mvj128. [DOI] [PubMed] [Google Scholar]
- Vanlandingham PA, Ceresa BP. Rab7 regulates late endocytic trafficking downstream of multivesicular body biogenesis and cargo sequestration. J Biol Chem. 2009;284:12110–12124. doi: 10.1074/jbc.M809277200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verderio C, et al. Myeloid microvesicles are a marker and therapeutic target for neuroinflammation. Ann Neurol. 2012;72:610–624. doi: 10.1002/ana.23627. [DOI] [PubMed] [Google Scholar]
- Von Bartheld CS, Altick AL. Multivesicular bodies in neurons: distribution, protein content, and trafficking functions. Prog Neurobiol. 2011;93:313–340. doi: 10.1016/j.pneurobio.2011.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Hundelshausen P, Weber C. Platelets as immune cells: bridging inflammation and cardiovascular disease. Circ Res. 2007;100:27–40. doi: 10.1161/01.RES.0000252802.25497.b7. [DOI] [PubMed] [Google Scholar]
- Waldenstrom A, Genneback N, Hellman U, Ronquist G. Cardiomyocyte microvesicles contain DNA/RNA and convey biological messages to target cells. PloS One. 2012;7:e34653. doi: 10.1371/journal.pone.0034653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolfers J, et al. Tumor-derived exosomes are a source of shared tumor rejection antigens for CTL cross-priming. Nat Med. 2001;7:297–303. doi: 10.1038/85438. [DOI] [PubMed] [Google Scholar]
- Zitvogel L, Regnault A, Lozier A, Wolfers J, Flament C, Tenza D, Ricciardi-Castagnoli P, Raposo G, Amigorena S. Eradication of established murine tumors using a novel cell-free vaccine: dendritic cell-derived exosomes. Nat Med. 1998;4:594–600. doi: 10.1038/nm0598-594. [DOI] [PubMed] [Google Scholar]
- Zwicker JI, Trenor CC, Furie BC, Furie B. Tissue factor-bearing microparticles and thrombus formation. Arterioscler Thromb Vasc Biol. 2011;31:728–733. doi: 10.1161/ATVBAHA.109.200964. [DOI] [PMC free article] [PubMed] [Google Scholar]