

Abstract
The embryonic chick is a widely used model for the study of peripheral and central ganglion cell projections. In the auditory system, selective labeling of auditory axons within the VIIIth cranial nerve would enhance the study of central auditory circuit development. This approach is challenging because multiple sensory organs of the inner ear contribute to the VIIIth nerve 1. Moreover, markers that reliably distinguish auditory versus vestibular groups of axons within the avian VIIIth nerve have yet to be identified. Auditory and vestibular pathways cannot be distinguished functionally in early embryos, as sensory-evoked responses are not present before the circuits are formed. Centrally projecting VIIIth nerve axons have been traced in some studies, but auditory axon labeling was accompanied by labeling from other VIIIth nerve components 2,3. Here, we describe a method for anterograde tracing from the acoustic ganglion to selectively label auditory axons within the developing VIIIth nerve. First, after partial dissection of the anterior cephalic region of an 8-day chick embryo immersed in oxygenated artificial cerebrospinal fluid, the cochlear duct is identified by anatomical landmarks. Next, a fine pulled glass micropipette is positioned to inject a small amount of rhodamine dextran amine into the duct and adjacent deep region where the acoustic ganglion cells are located. Within thirty minutes following the injection, auditory axons are traced centrally into the hindbrain and can later be visualized following histologic preparation. This method provides a useful tool for developmental studies of peripheral to central auditory circuit formation.
Keywords: Neurobiology, Issue 73, Neuroscience, Behavior, Developmental Biology, Anatomy, Biomedical Engineering, Surgery, Development, Inner Ear, Cochlea, Auditory, Chick, Axon Tracing, VIIIth Cranial Nerve, nerve, ganglion, fiber, cochlear duct, basilar papilla, embryo, microinjection, animal model
Protocol
1. Prepare the Following Dissection Tools and Reagents
Artificial cerebrospinal fluid (aCSF; 130 mM NaCl, 3 mM KCl, 1.2 mM KH2PO4, 20 mM NaHCO3, 3 mM HEPES, 10 mM Glucose, 2 mM CaCl2, 1.3 mM MgSO4) continuously infused with 95% O2 / 5% CO2 at room temperature. For infusion, fill to 2/3 a 500 ml wide-mouth Nalgene jar with a hole drilled in the lid. Tank will be attached by tubing to a glass stem bubbler, which penetrates the aCSF through the hole in the jar lid. Adjust pressure to obtain a constant stream of bubbles that reach about 1/3 of the liquid but are not forceful enough to cause splashing over the rim. Submerge 5 ml glass vials (up to 6) into the Nalgene jar to separate individual samples from each other and from the gas flow turbulence. ACSF should be oxygenated for at least 20 min prior to any tissue incubation.
Small silicone dissection dish (35 mm x 10 mm Petri coated using Sylgard Elastomere Kit) with two dissection pins placed into small hexagonal polystyrene (47 mm bottom) weigh boat.
Two fine-tip forceps, one curved-tip forceps, a 50 ml beaker, and a container for tissue waste.
Pulled glass micropipette (1.2 OD / 0.9 ID) broken to approximately 20-50 μm opening at the tip and filled with rhodamine dextran amine (RDA, MW = 3,000, Invitrogen) in a 6.25% solution with 0.4% Triton X-100 in PBS. Attach by fine tubing to picospritzer and stabilize to micromanipulator.
2. Micro-dissection to Reveal Basilar Papilla at E8 4,5
With curved forceps, cut E8 embryo just below the brainstem, placing head directly onto silicone-coated dissection dish within weigh boat.
Using dissection pins, position the head with a ventral view so the ear on the side of interest is slightly visible, the head tilted slightly dorsally and pointing 10-30 degrees away from center (Figure 2A). Immediately secure the head with pins. If bilateral tracing is desired, center the head at 0 degrees, or a full ventral view so both sides can be accessed. Similarly, for a left-sided injection, secure the head at -10 to -30 degrees from center so the left side is accessible.
Using the 50 ml beaker, transfer 30 ml of oxygenated aCSF into the dish, fully submerging the tissue.
Gently grasp and peel down the lower jaw using fine-tip forceps, then remove the lower eyelid on the side of interest.
Remove the overlying vascular tissue, including any remaining palate and gullet. These soft tissues should peel away revealing the smooth surface of developing chondrocranium with the distinctive vertebral arteries at its surface. By this point, the characteristic white otoconial mass should be visible.
Carefully dissect the skin around the external meatus, cartilage of the jaw joint, and the middle ear while leaving the inner ear intact. Use the forceps to gently cut away these cartilaginous structures.
Remove vertebral arteries and pooling blood using a gentle sweeping motion with the fine forceps tips. Blood can obscure view and coagulation may plug the injection pipette opening.
The cochlear duct with adjacent sensory epithelium (basilar papilla) and acoustic ganglion lies directly beneath the chondrocranium 6 and can be visualized as an elongated finger-like projection extending from the inner ear ventrally, with the distal-most tip (apex) near the anterior midline 5. There may be a small pooling of blood (red) from the dissection and/or otolithic calcifications (white) at the apex of the cochlear duct (Figure 2B) that can aid in visualization. At the apex is the lagenar macula, a sensory patch thought to be of vestibular function 7,8.
3. Selective Labeling of CN VIII Auditory Fibers
Place the injection pipette near the embryo surface at the cochlear duct and basilar papilla region.
Slowly lower the micropipette to first puncture the cranium. This cranial region is thinner than surrounding areas and requires less force to penetrate. The micropipette should now be inside the cochlear duct. Optional: a small injection of tracing dye can be made here to aid in visualizing the basilar papilla.
Without repositioning the micropipette, continue lowering until it just breaches the deep border of the cochlear duct (Figure 1). Perform an injection here and repeat in multiple regions along the length of the basilar papilla (Figures 2C, 2D), excluding the most proximal tip where the vestibular lagena is located. With the picospritzer set between 10-30 psi, several injections are made. The volume of injected dye fluid varies, and depends on the extent of labeling desired. Each injection should result in a fluorescent dye-labeled spot of approximately 200 - 400 μm diameter (Figure 2D).
Once the injection is complete, use forceps to transect the sample above the brainstem and use a transfer pipette to place the caudal portion into a small jar within the container of aCSF continually being perfused with 95% O2 / 5% CO2.
For each embryo, allow 20-30 min for dye transfer, depending on the distance of central tracing desired.
Remove tissue and prepare for histological sectioning. In the example shown here, tissue is immersed in 4% paraformaldehyde (in 1X PBS) overnight, 30% sucrose (in 1X PBS) overnight.
4. Counterstaining and Analysis
Counterstaining is useful to help orient the observer to anatomical location as well as identify particular regions of interest. An effective counterstain in this preparation is neurofilament immunolabeling (Figures 1, 3B, 3E), which strongly labels axons and allows for demarcation of axonal tracts in the CNS.
Analysis should be performed throughout the labeled region, as the projections can extend several hundred microns or more along the rostrocaudal axis. Some examples of basic analyses are: targeting error rates, timing and/or changes in divergence of projection patterns. RDA-labeled axons are visualized in the peripheral and central nervous system with high resolution (Figures 3A, 3D).
Representative Results
The components of the VIIIth nerve and the anatomy of the nerve itself are complex and convoluted (Figures 1, 3). By selectively tracing fibers arising from acoustic ganglion cells, segments of the VIIIth nerve as well as primary auditory afferents within the brainstem can be cleanly traced and distinguished from their vestibular counterparts (Figures 2, 3). Likewise, this technique could be used to study peripheral projections of the acoustic ganglion cells (Figure 3G), or modified to study projections arising from individual vestibular organs. Because axons are cleanly traced along their entire length, accurate quantitative analyses can be performed distinguishing auditory from vestibular projections during development, including precise timing and patterns of initial innervation, targeting error rates, etc. Additionally, unlike the previous approach of auditory fiber labeling, this approach is performed on non-fixed, living tissue and thus can be used simultaneous to other in vitro experimentation.
Though this technique is suitable for embryos from E6-E9, best results commence at E8 when auditory axons have innervated their primary central targets 3. In contrast, vestibular projections arrive in the brainstem earlier 9,10 and thus would serve as better candidates for earlier tracing, though technically challenging due to the small size and relative immaturity of the vestibular apparatus at this age. Histological sections should be examined to confirm the location of dye injection, as the apical-most region of the basilar papilla contains a small sensory patch thought to be of vestibular function and if labeled could potentially contaminate the results.
Once the investigator is familiar with the anatomy, three of the most commonly encountered problems are as follows:
The cochlear duct is filled with tracing dye, but ganglion cell axons are not labeled. This problem likely indicates that the injection was not performed deep to the cochlear duct, where the ganglion cells and nerve fibers reside. This problem may be corrected by ensuring that the injection micropipette first punctures the chondrocranium, then is gently advanced so as to just puncture through the other side of the duct. Alternatively, the injection pipette may have been correctly positioned but delivered insufficient quantities of dye. Increasing injection pressure or number of injections may also overcome this issue.
The nerve is inadvertently transected as a consequence of excessive pulling on the cartilaginous labyrinth during dissection. This difficulty can be avoided by using fine dissection techniques and limited dissection of the middle ear.
The injection is too extensive and inadvertently labels other sensory ganglion cells or fibers arising from other nerve components. This problem can be minimized by optimizing injection protocols, and may require reducing the injection pressure or the number of pulses, or shifting the location of injections.
Two examples of potential analyses are:
Divergence of projection patterns: determines the extent of labeled axons along a particular axis and how they might be affected following experimentation. Rostrocaudal divergence can be described as:
(Number sections containing labeled axons) x (Thickness of each section) This measure may be corrected for the extent of dye label as a proportion of the basilar papilla, or in counterstained tissue, for the proportion of VIIIth nerve axons labeled. This is a useful tool for analyzing topographic projections, as the labeled axons can be cleanly traced to their terminal fields in the hindbrain. For measurements of tonotopy, dye injections should be limited to the region of interest along the basilar papilla.
Targeting errors: determines the frequency of mistargeted axons for every predetermined region of analyzed tissue (can be per section, per embryo, etc.). Perturbations may result in axons following an abnormal projection path or terminating in an inappropriate region. When quantifying errors, numbers should be normalized to account for difference in injection amounts between embryos. Smaller injections are more likely to result in the ability to score single axons. A basic targeting error analysis can be described as:
![]()
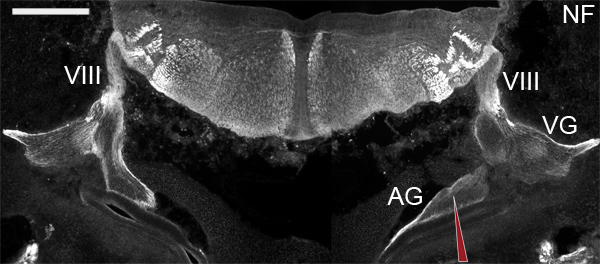 Figure 1. Coronal view of E6 brainstem for simplification with bilateral acoustic ganglia intact. Schematized red pipette illustrates accurate targeting of acoustic ganglion cells from a dorsolateral entry point. Sections are immunolabeled with anti-neurofilament to highlight axonal projections. Vestibular organs are rostral to sections shown. Region of RDA injection for selective auditory fiber tracing demonstrated with red pipette illustration. VIII = vestibulocochlear nerve, AG = acoustic ganglion, VG = vestibular ganglion. Dorsal is up. Scale bar = 300 μm.
Figure 1. Coronal view of E6 brainstem for simplification with bilateral acoustic ganglia intact. Schematized red pipette illustrates accurate targeting of acoustic ganglion cells from a dorsolateral entry point. Sections are immunolabeled with anti-neurofilament to highlight axonal projections. Vestibular organs are rostral to sections shown. Region of RDA injection for selective auditory fiber tracing demonstrated with red pipette illustration. VIII = vestibulocochlear nerve, AG = acoustic ganglion, VG = vestibular ganglion. Dorsal is up. Scale bar = 300 μm.
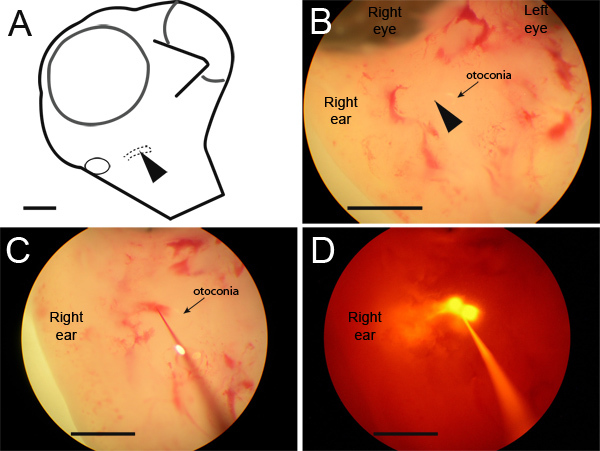 Figure 2. Microdissection and injection of target region.(A) Appropriate position of embryo on dissection dish, ventral view with head tilted dorsally and turned slightly to the side to allow better access to the right side. Right ear is shown and underlying basilar papilla is indicated with a dashed line. Arrowhead in (A) and (B) illustrates initial position of pipette insertion (B) Following dissection, a white otoconial mass is visible, demarcating the apical border of the basilar papilla (arrow). (C,D) First injection should provide an outline of the cochlear duct. (D) Subsequent injections should be made just deep to the cochlear duct and basilar papilla into the acoustic ganglion region. RDA fluorescence is useful in enhancing injection depth perception and each individual injection should create a fluorescent dye-labeled spot of approximately 200 - 400 μm diameter as seen here. Scale bar = 2 mm.
Figure 2. Microdissection and injection of target region.(A) Appropriate position of embryo on dissection dish, ventral view with head tilted dorsally and turned slightly to the side to allow better access to the right side. Right ear is shown and underlying basilar papilla is indicated with a dashed line. Arrowhead in (A) and (B) illustrates initial position of pipette insertion (B) Following dissection, a white otoconial mass is visible, demarcating the apical border of the basilar papilla (arrow). (C,D) First injection should provide an outline of the cochlear duct. (D) Subsequent injections should be made just deep to the cochlear duct and basilar papilla into the acoustic ganglion region. RDA fluorescence is useful in enhancing injection depth perception and each individual injection should create a fluorescent dye-labeled spot of approximately 200 - 400 μm diameter as seen here. Scale bar = 2 mm.
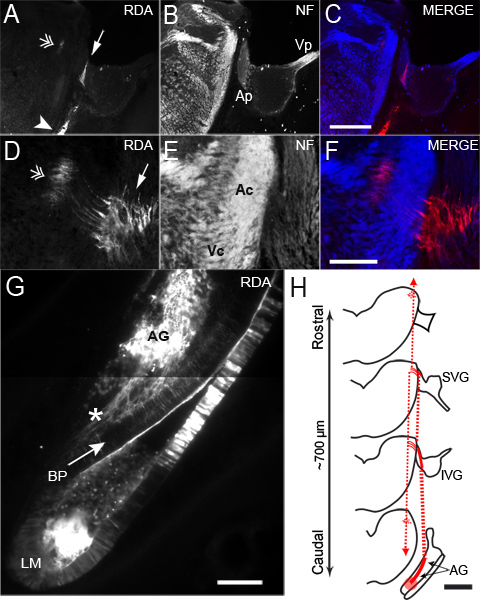 Figure 3. Representative images of selective auditory fiber tracing in an E8 embryo.(A-C) Low power and (D-G) high power 25 μm coronal sections collected from the same embryo at different planes. (A) RDA labeled axons can be traced from injection site (arrowhead), through the cranial nerve (arrow) and caudally toward the primary auditory nuclei targets (double arrowhead). (B) Immunolabeling with anti-neurofilament antibody highlights all axonal projections and the RDA-traced fibers from (A) are within the auditory projection pathway. Overlay of separate (A) and (B) channels shown as a color merge in (C). (D-F) High power image of the same embryo at a more caudal position demonstrates (A) high resolution of fibers at the nerve entry point (arrow) and along the central pathway (double arrowhead). (E) Neurofilament stain of all axons allows demarcation of PNS-CNS as well as putative auditory versus vestibular projections. Ap and Vp indicate respective auditory and vestibular components peripheral to the nerve entry point, whereas Ac and Vc indicate respective auditory and vestibular projections central to the nerve entry point. Overlay of separate (D) and (E) channels shown as a color merge in (F). (G) Histologic examination of the acoustic ganglion reveals location of labeled acoustic ganglion cells from the tracing shown in (A-F) and their peripheral projections (asterisk) are traced retrogradely to the basilar papilla. (H) Illustration of a 700 μm tracing pathway expected with auditory-specific VIII labeling at E8. Injection site (red circle) is caudal to nerve entry point, and central projections travel rostral and caudally once in the hindbrain. Vp = peripheral vestibular, Ap = peripheral auditory, Vc = central vestibular, Ac = central auditory, AG = acoustic ganglion, BP = basilar papilla, LM = lagenar macula, SVG = superior vestibular ganglion, IVG = inferior vestibular ganglion. Scale bar = 300 μm for A-C, 100 μm for D-F, 100 μm in G and 500 μm in H.
Figure 3. Representative images of selective auditory fiber tracing in an E8 embryo.(A-C) Low power and (D-G) high power 25 μm coronal sections collected from the same embryo at different planes. (A) RDA labeled axons can be traced from injection site (arrowhead), through the cranial nerve (arrow) and caudally toward the primary auditory nuclei targets (double arrowhead). (B) Immunolabeling with anti-neurofilament antibody highlights all axonal projections and the RDA-traced fibers from (A) are within the auditory projection pathway. Overlay of separate (A) and (B) channels shown as a color merge in (C). (D-F) High power image of the same embryo at a more caudal position demonstrates (A) high resolution of fibers at the nerve entry point (arrow) and along the central pathway (double arrowhead). (E) Neurofilament stain of all axons allows demarcation of PNS-CNS as well as putative auditory versus vestibular projections. Ap and Vp indicate respective auditory and vestibular components peripheral to the nerve entry point, whereas Ac and Vc indicate respective auditory and vestibular projections central to the nerve entry point. Overlay of separate (D) and (E) channels shown as a color merge in (F). (G) Histologic examination of the acoustic ganglion reveals location of labeled acoustic ganglion cells from the tracing shown in (A-F) and their peripheral projections (asterisk) are traced retrogradely to the basilar papilla. (H) Illustration of a 700 μm tracing pathway expected with auditory-specific VIII labeling at E8. Injection site (red circle) is caudal to nerve entry point, and central projections travel rostral and caudally once in the hindbrain. Vp = peripheral vestibular, Ap = peripheral auditory, Vc = central vestibular, Ac = central auditory, AG = acoustic ganglion, BP = basilar papilla, LM = lagenar macula, SVG = superior vestibular ganglion, IVG = inferior vestibular ganglion. Scale bar = 300 μm for A-C, 100 μm for D-F, 100 μm in G and 500 μm in H.
Discussion
Studies of the early development of the VIIIth nerve have been limited in part because of the difficulty in identifying embryonic axons arising from multiple distinct ganglia. Several studies have explored the molecular signals guiding auditory and vestibular sensory cell and ganglion cell fates during early development, 5,11,12 but the processes regulating central innervation have yet to be determined. Reports of acoustic ganglion cell projections typically describe peripheral processes to sensory epithelium 13-15, whereas less is known about the central processes projecting to primary nuclei. Studies of centrally-projecting VIII subdivisions during embryogenesis are often electrophysiological, such as optical recordings in embryonic chick hindbrain 16 or calcium imaging in a mammalian tissue slice 17 but cannot be performed prior to completion of neural circuits. The use of immunological markers to label auditory axons in the chick VIIIth nerve is not currently feasible. While distinct groups of transcription factors have been identified that correlate with auditory versus vestibular neurons18-23, the expression of these factors is excluded from axons. In mice, differences have been found between spiral ganglia and vestibular ganglia in gene expression, including some promising candidates that could differentiate axon growth among modalities.23 Further study of these gene products may yield important tools for both mammalian and avian studies. With the methods described here, auditory fibers in living embryonic tissue with an intact auditory circuit can be selectively traced to their central targets.
The chicken embryo is an especially useful vertebrate system for developmental biologists and is able to overcome some of the limitations found in mammalian systems. Regulation of early inner ear patterning and morphogenesis in birds shows remarkable similarity to mammals 1,12, while chick embryos are larger and more easily accessible than rodent embryos at early stages when the peripheral auditory system is being formed. The method presented here for selective tracing of auditory fibers facilitates early experimentation on the central projections of the developing inner ear.
We developed this protocol in an effort to differentially identify central auditory projections of the VIIIth nerve, but likewise it can be modified to selectively label fibers from other sensory organs of the inner ear. In contrast to previously described methods, this procedure is performed on living tissue with the entire circuit intact. A similar method performed was used to describe timing of VIII auditory afferent innervation during avian embryogenesis; however, vestibular fibers were also inadvertently labeled.3 Here we provide a method to clearly identify region of origin and termination, as well as high single fiber resolution of select centrally-projecting auditory afferents during vertebrate embryogenesis.
Disclosures
No conflicts of interest are declared.
Acknowledgments
The authors wish to thank Dr. Candace Hsieh for suggestions and assistance with imaging techniques and Dr. Doris Wu for expertise on chick inner ear anatomy during early embryogenesis. This work was supported by NSF IOS-0642346, NIH T32-DC010775, NIH T32-GM008620, NIH R01-DC010796, and DOE GAANN P200A120165.
References
- Groves AK, Fekete DM. Shaping sound in space: the regulation of inner ear patterning. Development. 2012;139:245–257. doi: 10.1242/dev.067074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pflieger JF, Cabana T. The vestibular primary afferents and the vestibulospinal projections in the developing and adult opossum, Monodelphis domestica. Anatomy and Embryology. 1996;194:75–88. doi: 10.1007/BF00196317. [DOI] [PubMed] [Google Scholar]
- Molea D, Rubel EW. Timing and topography of nucleus magnocellularis innervation by the cochlear ganglion. The Journal of Comparative Neurology. 2003;466:577–591. doi: 10.1002/cne.10896. [DOI] [PubMed] [Google Scholar]
- Bissonnette JP, Fekete DM. Standard atlas of the gross anatomy of the developing inner ear of the chicken. The Journal of Comparative Neurology. 1996;368:620–630. doi: 10.1002/(SICI)1096-9861(19960513)368:4<620::AID-CNE12>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- Brigande JV, Kiernan AE, Gao X, Iten LE, Fekete DM. Molecular genetics of pattern formation in the inner ear: do compartment boundaries play a role. Proceedings of the National Academy of Sciences of the United States of America. 1073;97:11700–11706. doi: 10.1073/pnas.97.22.11700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bellairs R, Osmond M. The atlas of chick development. 2nd Edn. Elsevier; 2005. [Google Scholar]
- Manley GA, Haeseler C, Brix J. Innervation patterns and spontaneous activity of afferent fibres to the lagenar macula and apical basilar papilla of the chick's cochlea. Hearing Research. 1991;56:211–226. doi: 10.1016/0378-5955(91)90172-6. [DOI] [PubMed] [Google Scholar]
- Code RA. Efferent neurons to the macular lagena in the embryonic chick. Hearing Research. 1995;82:26–30. doi: 10.1016/0378-5955(94)00159-n. [DOI] [PubMed] [Google Scholar]
- Maklad A, Fritzsch B. Development of vestibular afferent projections into the hindbrain and their central targets. Brain Research Bulletin. 2003;60:497–510. doi: 10.1016/s0361-9230(03)00054-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubel EW, Fritzsch B. Auditory system development: primary auditory neurons and their targets. Annual Review of Neuroscience. 2002;25:51–101. doi: 10.1146/annurev.neuro.25.112701.142849. [DOI] [PubMed] [Google Scholar]
- Satoh T, Fekete DM. Lineage analysis of inner ear cells using genomic tags for clonal identification. Methods Mol. Biol. 2009;493:47–63. doi: 10.1007/978-1-59745-523-7_4. [DOI] [PubMed] [Google Scholar]
- Bok J, Chang W, Wu DK. Patterning and morphogenesis of the vertebrate inner ear. The International Journal of Developmental Biology. 2007;51:521–533. doi: 10.1387/ijdb.072381jb. [DOI] [PubMed] [Google Scholar]
- Appler JM, Goodrich LV. Connecting the ear to the brain: Molecular mechanisms of auditory circuit assembly. Progress in Neurobiology. 2011;93:488–508. doi: 10.1016/j.pneurobio.2011.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bulankina AV, Moser T. Neural circuit development in the mammalian cochlea. Physiology (Bethesda) 2012;27:100–112. doi: 10.1152/physiol.00036.2011. [DOI] [PubMed] [Google Scholar]
- Fekete DM, Campero AM. Axon guidance in the inner ear. The International Journal of Developmental Biology. 2007;51:549–556. doi: 10.1387/ijdb.072341df. [DOI] [PubMed] [Google Scholar]
- Momose-Sato Y, Glover JC, Sato K. Development of functional synaptic connections in the auditory system visualized with optical recording: afferent-evoked activity is present from early stages. Journal of Neurophysiology. 2006;96:1949–1962. doi: 10.1152/jn.00319.2006. [DOI] [PubMed] [Google Scholar]
- Marrs GS, Spirou GA. Embryonic assembly of auditory circuits: spiral ganglion and brainstem. The Journal of Physiology. 2012;590:2391–2408. doi: 10.1113/jphysiol.2011.226886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milo M, et al. Genomic analysis of the function of the transcription factor gata3 during development of the mammalian inner ear. PloS One. 2009;4:e7144. doi: 10.1371/journal.pone.0007144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fritzsch B, Eberl DF, Beisel KW. The role of bHLH genes in ear development and evolution: revisiting a 10-year-old hypothesis. Cellular and Molecular Life Sciences : CMLS. 2010;67:3089–3099. doi: 10.1007/s00018-010-0403-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jahan I, Kersigo J, Pan N, Fritzsch B. Neurod1 regulates survival and formation of connections in mouse ear and brain. Cell and Tissue Research. 2010;341:95–110. doi: 10.1007/s00441-010-0984-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang EJ, et al. Brn3a is a transcriptional regulator of soma size, target field innervation and axon pathfinding of inner ear sensory neurons. Development. 2001;128:2421–2432. doi: 10.1242/dev.128.13.2421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones JM, Warchol ME. Expression of the Gata3 transcription factor in the acoustic ganglion of the developing avian inner ear. The Journal of Comparative Neurology. 2009;516:507–518. doi: 10.1002/cne.22128. [DOI] [PubMed] [Google Scholar]
- Lu CC, Appler JM, Houseman EA, Goodrich LV. Developmental profiling of spiral ganglion neurons reveals insights into auditory circuit assembly. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 2011;31:10903–10918. doi: 10.1523/JNEUROSCI.2358-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]