

Abstract
The innate immune system senses pathogens by pattern recognition receptors in different cell compartments. In the endosome, bacteria are generally recognized by TLRs; facultative intracellular bacteria such as Listeria, however, can escape the endosome. Once in the cytosol, they become accessible to cytosolic pattern recognition receptors, which recognize components of the bacterial cell wall, metabolites or bacterial nucleic acids and initiate an immune response in the host cell. Current knowledge has been focused on the type I IFN response to Listeria DNA or Listeria-derived second messenger c-di-AMP via the signaling adaptor STING. Our study focused on the recognition of Listeria RNA in the cytosol. With the aid of a novel labeling technique, we have been able to visualize immediate cytosolic delivery of Listeria RNA upon infection. Infection with Listeria as well as transfection of bacterial RNA induced a type-I-IFN response in human monocytes, epithelial cells or hepatocytes. However, in contrast to monocytes, the type-I-IFN response of epithelial cells and hepatocytes was not triggered by bacterial DNA, indicating a STING-independent Listeria recognition pathway. RIG-I and MAVS knock-down resulted in abolishment of the IFN response in epithelial cells, but the IFN response in monocytic cells remained unaffected. By contrast, knockdown of STING in monocytic cells reduced cytosolic Listeria-mediated type-I-IFN induction. Our results show that detection of Listeria RNA by RIG-I represents a non-redundant cytosolic immunorecognition pathway in non-immune cells lacking a functional STING dependent signaling pathway.
Introduction
Multicellular organisms evolved efficient host-defense mechanisms to sense invading pathogens such as bacteria and viruses in order to block their replication and spread. Components of the bacterial cell wall and conserved bacterial proteins are recognized by germ line encoded Toll-like receptors (TLRs) on the surface of cells and by receptors of the nucleotide-binding domain and leucine-rich repeat containing gene family (NLRA,B,C,P,X) in the cytosol [1]. Recognition of nucleic acids plays a major role in immune responses during viral infection. The pathogenicity of foreign genetic material is either recognized through its location, specific structural features or modification of nucleic acids. Nucleic acids are recognized by TLRs in the endosome. TLR3 detects double stranded RNA. TLR7 and TLR8 detect double and single stranded RNA respectively, with a preference for certain sequence motifs (reviewed in [2]). TLR9 is activated by DNA containing unmethylated CpG motifs [3], [4], [5]. In the cytosol, the helicase MDA5 [6] recognizes long double stranded RNA with additional structural requirements as yet not well defined [7], [8] and the helicase RIG-I [6] binds to and is activated by 5′triphosphorylated double stranded RNA [9], [10], [11], [12], which is present either in viral genomic RNA or is formed during viral replication. Two independent studies described a TLR9-independent pathway of cytosolic recognition of double stranded DNA leading to type I IFN induction [13], [14]. So far, numerous candidate cytosolic DNA receptors including DAI, DDX41, IFI16, LRRFIP1, Ku70, DHX36, DHX9 and DNA-PK have been suggested [15]. All studies agree on the fact that the mitochondrial adaptor protein STING (also known as MITA) downstream of the putative DNA receptor is essential for sensing cytosolic DNA [16], [17]. A recent study by Sun et al. identified Cyclic GMP-AMP Synthase (cGAS) as the so far most convincing DNA recognizing candidate receptor [18]. Upon recognition of DNA, cGAS synthesize the second messenger Cyclic GMP-AMP (cGAMP) which activates STING [18], [19].
Several groups [20], [21], [22] identified AIM2 to be a cytosolic dsDNA sensing receptor, which activates caspase 1, leading to the release of IL-1. While the role of AIM2 in the activation of caspase-1 is non-redundant, AIM2 is not involved in the activation of transcription factors. Recent work demonstrated that AT-rich DNA can be sensed by an indirect recognition mechanism. It was demonstrated that the endogenous RNA polymerase III (pol III) uses AT-rich DNA (poly(dA-dT)) as a template, leading to generation of 5′triphosphorylated poly AU-RNA, which forms an dsRNA and therefore represents a strong RIG-I ligand [11], [23], [24]. Due to the ubiquitous expression of RIG-I, poly(dA-dT)-mediated type I IFN induction appears to occur in all cell types analyzed so far. However, this pol III/RIG-I dependent pathway for type I IFN induction is not activated upon transfection of mixed DNA sequences lacking AT rich regions such as plasmid DNA or PCR products. In the human system only certain immune cells with an intact STING-dependent DNA sensing pathway, e.g. monocytic cells, are able to induce type I IFN by PCR products or plasmid DNA.
IFN induction by RNA from RNA viruses has been explored and understood far better than type I IFN induction by DNA. Binding of RIG-I and MDA5 to the mitochondrial adaptor molecule MAVS (also known as IPS1, Cardif or VISA [25], [26], [27], [28]) leads to the binding and the activation of IRF3, which induces type I IFN expression. MAVS is essential for RIG-I and MDA5 signaling, but not for RIG-I independent ( = STING dependent) pathways of dsDNA mediated type I IFN induction [29].
Bacteria trigger IFN-β responses through stimulation of TLR4 on the cell surface or TLR9 in endosomes. Investigations in the labs of Decker and Portnoy suggested that TLR-independent pathways exist, which lead to the induction of type I IFN in mouse macrophages infected with Listeria monocytogenes [30], [31]. Interestingly, type I IFN induction depended on cytosolic localization of the bacteria [30], [31] but was shown to be NOD2-independent [32]. Later on, the requirement of MAVS and, consequently, MDA5 or RIG-I in type I IFN induction was excluded in murine bone marrow-derived or peritoneal macrophages and MEFs [29], [33]. Stetson and Medzhitov [13] observed that DNA represents the type I IFN inducing agent in the lysate of Listeria monocytogenes when transfected into murine monocytes. From their experiments, they concluded that intracellular bacteria (L. monocytogenes and Legionella pneumophila) with cytosolic access or cytosolic contact, induce a type I IFN response upon recognition of bacterial DNA in the cytosol. Moreover, cyclic diadenosine monophosphate (c-di-AMP), a metabolite of L. monocytogenes similar to the endogenous second messenger cGAMP was found to induce type I IFN induction directly via STING [34], [35]. The fact that the DNA-sensing AIM2 inflammasome [20], [21], [22] is involved in Listeria-induced caspase 1 activation [36], [37], [38], clearly supports the contribution of released bacterial DNA to the immune response.
Here we hypothesized that, like DNA, bacterial RNA can enter the cytosol, where it is recognized by cytosolic RNA receptors, for example RIG-I-like helicases. Indeed, in contrast to eukaryotic mRNA, bacterial mRNA is not capped but contains 15% 5′triphosphorylated RNA [39], which would render bacterial RNA an ideal PAMP due to its ability to activate RIG-I. In this study, we used the bacterium L. monocytogenes as a model organism for intracellular bacteria-host interaction. L. monocytogenes is an opportunistic bacterium responsible for human food-borne infections leading to meningitis and miscarriages. After crossing the intestinal barrier it enters lymph nodes, spleen and liver. In immunocompromised individuals, bacterial multiplication can occur in hepatocytes with further release of the bacteria into the blood and spread to the brain and the placenta (reviewed in [40]). Crossing the host barriers involves bacterial invasion and survival within a large variety of nonphagocytic cells [41]. Internalins (InlA and InlB) allow invasion via E-cadherin on the surface of epithelial cells and via hepatocyte growth factor receptor (MET), which is expressed on a wide range of cells [41]. With the current study, we provide direct evidence that RNA isolated from bacteria induces type I IFN in a 5′ phosphorylation-dependent manner. Using an advanced RNA labeling technique, we were able to show that RNA from L. monocytogenes translocates to the cytosol of the host cell during infection. This RNA sensing pathway induced both type I IFN and CXCL10 during infection with L. monocytogenes in non-monocytic cells, including hepatocytes and colon epithelial cells. Using RNAi we revealed that RIG-I is crucial for L. monocytogenes-induced type I IFN and CXCL10 induction in cell types, such as non-immune cells, without a functional STING-dependent immune response, as indicated by the absence of a direct sensing mechanism for cytosolic DNA.
Results
Bacterial RNA is Recognized by Human Monocytes by a TLR-independent but RNA 5′-phosphate Dependent Pathway
Initially, we evaluated whether bacterial DNA and RNA isolated from extracellular and intracellular bacteria can trigger a TLR-independent type I IFN response. For this purpose, PBMCs were preincubated with chloroquine to block endosomal TLRs (TLR7, TLR8 and TLR9) and were then transfected with bacterial DNA (bacDNA) or bacterial RNA (bacRNA) extracted from the indicated bacteria (Fig. 1A). Chloroquine suppresses endosomal TLR-activity [46] as monitored by CpG ODN transfection (Fig. S1). DNase I-treated bacRNA still induced type I IFN to the same degree as non-treated bacRNA, indicating that DNA contamination did not account for the stimulatory activity (Fig. 1A). As shown previously, triphosphates at the 5′ end are critical for RIG-I-mediated recognition of RNA [10]. To address the involvement of RIG-I in bacRNA recognition, RNA was treated with alkaline phosphatase to remove triphosphates from the 5′ ends (Fig. 1B). Indeed, dephosphorylation of bacRNA diminished IFN-α-inducing activity, suggesting a triphosphate-dependent activation pathway (Fig. 1B). This activation pattern was similar for all analyzed extracellular or facultative intracellular bacteria including E. coli (extracellular), L. monocytogenes (facultative intracellular), Staphylococcus aureus (facultative intracellular, [47]) and Acinetobacter baumannii (facultative intracellular [48]) (Fig. 1B). Altogether, these data indicate that the RNA of all tested bacteria activate a triphosphate dependent, TLR independent, type I IFN-inducing pathway when transfected into cells.
Figure 1. Bacterial RNA is recognized by human monocytes in a TLR-independent and 5′phosphorylation-dependent pathway.
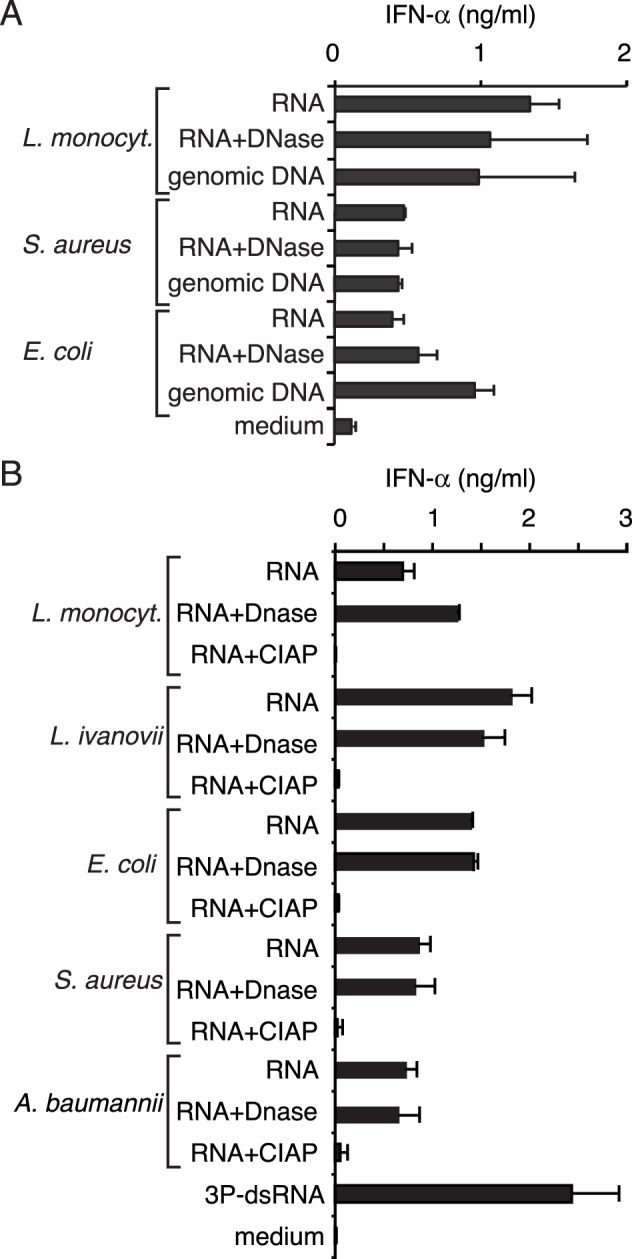
Human PBMC were preincubated with chloroquine and transfected with indicated nucleic acids. IFN-α production was analyzed 24 hours after stimulation. Error bars represent s.d. A: The IFN-α-inducing activity of bacterial RNA (untreated or DNase treated) and DNA of L. monocytogenes, Staphylococcus aureus and Escherichia coli was analyzed. B: The IFN-α-inducing activity of RNAs from L. monocytogenes, L. ivanovii, E. coli, S. aureus and Acinetobacter baumannii, treated with DNase or calf intestine alkaline phosphatase (CIAP), were compared.
RNA of Listeria monocytogenes has Access to the Cytosol of the Host Cell during Infection
To assess the relevance of our findings in the host-pathogen interaction during infection we investigated whether bacRNA is able to reach RIG-I localized in the cytosol of the host cell. The bacterium L. monocytogenes is a well-characterized model organism for studying intracellular bacteria-host interaction. In particular, it is able to escape the endosome and migrate into the cytosol of macrophages, human (not murine) epithelial cells [49], [50] or hepatocytes [51], a process mediated by the bacterial pore-forming toxin “listeriolysin” (LLO, encoded by the hly gene). In the following experiments, we selected cell lines that are derived from tissues or cell types involved in L. monocytogenes pathogenesis in vivo. THP-1 is an acute monocytic leukemia cell line similar to human monocyte-derived macrophages. We visualized transfer of bacRNA into the cytosol of cells using a recently developed sensitive, non-radioactive but non-toxic method to label RNA in living cells [52] (Fig. 2): 5-ethynyluridine (EU) was shown to be incorporated into RNA transcripts generated by RNA polymerases I, II and III in mammalian cells but not into DNA [52]. Listeria were grown in medium containing EU and FITC. The host cells were then infected with labeled bacteria. One or four hours post infection, host cells containing Listeria in the cytosol were fixed, permeabilized and incubated with fluorescence dye coupled to a reactive azide group (Alexa594-azide). In this setting, the azide selectively couples to the ethynyl group of EU incorporated into the bacRNA. Whole Listeria are labeled green with FITC; RNA is visible as red fluorescence (Alexa594), nuclei are stained by DAPI (blue). As evident from fluorescence images, the cytosol of THP-1 cells was clearly labeled for EU-containing RNA when infected with wild type (wt) L. monocytogenes (Fig. 2A left and middle panel) but not when infected with a mutant lacking LLO (hly-) (Fig. 2A right panel) and therefore impaired in its escape from the endosome.
Figure 2. RNA of L. monocytogenes has access to the cytosol of the host cell during infection.

THP-1, A549 and HepG2 were infected with FITC-tagged and EU-labeled wt and hly- L. monocytogenes for the indicated duration. Cells were then fixed, stained with Alexa594-azide and counterstained with DAPI. Left column, wt L. monocytogenes infection 1 hr. Middle column, wt L. monocytogenes infection 4 hrs. Right column, hly- L. monocytogenes infection 4 hrs. A: THP-1 cells. B: A549 cells. C: HepG2 cells. As determined by counting of single bacteria in cells (50 cells per slide were counted) the average bacterial load was 9(wt) and 4(hly-) bacteria per cell for THP-1 cells, 6(wt) and 4(hly-) bacteria per cell for A549 cells and 6(wt) and 5(hly-) bacteria per cell for HepG2 cells, one representative experiment out of two is shown. Whole L. monocytogenes are labeled green with FITC and RNA is visible as red fluorescence (Alexa594), nuclei are stained by DAPI (blue).
Bacterial RNA accumulated in the cytosol of host cells upon prolonged infection time with wt L. monocytogenes (Fig. 2A, left and middle panel). In concordance with findings from THP-1 cells, similar results were obtained with epithelial (A549, Fig. 2B ) and hepatocarcinoma cell lines (HepG2, Fig. 2C). During Listeria infection only bacRNA delivered to the cytosol of the host cell could be detected. As direct labeling of RNA from gram negative E. coli, which lack a gram positive cell wall, was possible (Fig. S2A), the gram positive cell wall appeared to be the reason for the absence of RNA staining in the bacteria.
To confirm equal incorporation of EU in the RNA of L. monocytogenes strains, bacterial RNA of wt and hly- L. monocytogenes incubated in EU-containing medium was extracted and labeled with Alexa594-azide in vitro. The data confirm that EU was equally incorporated in wt and hly- L. monocytogenes RNA (Fig. S2B). The absence of labeled RNA in the host cell nucleoli, the site of ribosomal RNA production, excluded transfer of EU nucleotides from L. monocytogenes with subsequent incorporation into host RNA. In fact, direct labeling of cells in conditioned culture medium led to strong staining of the nucleoli (Fig. S2C). Of note, EU labeling was performed in starvation medium that facilitates the incorporation of EU. Indeed, EU is not quantitatively incorporated in host RNA when cultured in standard culture medium for this short time span (Fig. S2D). Since we cultivated cell lines in standard culture medium during infection, we additionally excluded the possibility that host cell RNA is stained by non-incorporated EU released from L. monocytogenes.
From the data we conclude that during infection, significant amounts of bacRNA are transferred into the cytosol of cells where it can be detected by cytosolic immune receptors.
THP-1 cells infected with L. monocytogenes lacking the secA2 secretion system, which was recently identified to be responsible for secretion of bacterial RNA of L. monocytogenes, were not labeled for EU-containing RNA (Fig. S3), suggesting that translocation of bacterial RNA is rather mediated by active secretion than by lysis of bacteria [53].
Type I IFN induction by L. monocytogenes in epithelial cells and hepatocytes is triggered by recognition of bacterial RNA.
Work by Ishii et al. [54] suggested that cytosolic recognition of DNA represents an ubiquitous pathway expressed in a wide range of cell lines and cell types. More recent studies exhibited that in human cell lines this refers only to the recognition of AT-rich [23], [55] sequences. Long AT-rich DNA sequences (poly(dA-dT)) were found to be the template of RNA polymerase III (pol III) leading to synthesis of triphosphorylated RNA, the ligand of RIG-I [23], [24]. However, non-AT rich dsDNA molecules in the size range of poly(dA-dT) still induced a type I IFN response in human monocyte-derived dendritic cells (MoDC) in which RIG-I is silenced [23]. This points to two distinct DNA recognition pathways in immune cells: pol III/RIG-I dependent recognition of AT-rich DNA and STING dependent (polIII/RIG-I independent) type I IFN induction by long random DNA. This redundancy in DNA sensing mechanisms complicates the delineation of pattern recognition receptors involved in L. monocytogenes sensing. Recent data suggested that the STING dependent pathway functions mainly in immune cells, whereas tumor cell lines such as HEK293 cells only responded to poly(dA-dT) in a pol III/RIG-I dependent fashion [23]. Indeed, we also observed that while THP-1 cells responded to all kinds of transfected DNA including poly(dA-dT), plasmid DNA (pDNA), genomic bacterial DNA and an 84mer double stranded DNA oligonucleotide (dsODN), the lung epithelial cell line A549 only responded to poly(dA-dT) and double stranded 5′ triphosphate RNA (3P-dsRNA) (Fig. 3A). Analogous results were obtained when we used bacRNA or bacDNA derived from Listeria (Fig. 3B). Both transfected bacRNA and bacDNA induced equal amounts of type I IFN in monocytic THP-1 cells (Fig. 3B). By contrast, A549, HepG2 (hepatocarcinoma) and Colo205 (colon carinoma) cells were able to sense transfected bacRNA but did not induce a type I IFN response upon transfection of bacDNA (Fig. 3B). We thereby conclude that type I IFN induction by L. monocytogenes bacDNA is restricted to immune cells with intact STING- dependent recognition pathways, which are not functional in human non-immune cells. By contrast, non-immune cells are exclusively triggered by bacRNA to induce type I IFN.
Figure 3. Recognition of bacterial RNA or DNA varies for different cell types.

A: THP-1 and A549 cells were transfected with double stranded triphosphorylated RNA (3P-dsRNA), poly(dA-dT), bacterial DNA (bacDNA), plasmid DNA (pDNA) or double stranded 84 mer DNA oligonucleotides (dsODN); B: THP-1, A549, HepG2 and Colo205 cells were transfected with L. monocytogenes RNA, L. monocytogenes DNA or 3P-dsRNA. Type I IFN (THP-1, A549, Colo205) or CXCL10 (HepG2) production was analyzed 24 hours after stimulation. The relative induction of the indicated cytokine is depicted as percentage of induction by transfected L. monocytogenes (L.M.) RNA. C, D, E and F: THP-1, A549, HepG2 and Colo205 cells were infected with wt and hly- L. monocytogenes at the indicated MOI. Type I IFN (THP-1, A549, Colo205) or CXCL10 production (HepG2) was analyzed 24 hours after stimulation. Error bars represent s.d.
Subsequently we tested whether L. monocytogenes infection of cell lines, unresponsive to bacDNA, can still induce a type I IFN response. To this end, we infected the indicated cell lines with wild type (wt) or LLO deficient (hly-) L. monocytogenes at the MOI shown and assessed the type I IFN secretion (Fig. 3C,D, E, F). THP-1 cells, which can respond to bacDNA, raised a robust type I IFN response upon infection with wt L. monocytogenes (Fig. 3C). Strikingly, cell lines derived from non-immune cells lacking a type I IFN response to bacDNA (A549, HepG2, Colo205) also induced substantial amounts of type I IFN or the type I IFN regulated chemokine CXCL10 (HepG2) when infected with wt L. monocytogenes (Fig. 3D, E, F). In all analyzed cell lines L. monocytogenes induced type I IFN/CXCL10 in a LLO dependent manner, strongly suggesting cytosolic recognition of bacRNA (Fig. 3C, D, E, F). Together these results indicate that bacRNA translocated to the cytosol is the causative agent for L. monocytogenes-mediated type I IFN induction in non-immune cells.
Type I IFN Induction by L. monocytogenes Infection Requires RIG-I in Epithelial but not in Monocytic Cells
Since epithelial cells were able to respond to transfected bacRNA but not transfected bacDNA, we examined if cytosolic RNA receptors are essential for recognition of bacterial RNA during infection. RIG-I, recognizing 5′triphosphorylated RNAs, is a likely candidate cytosolic RNA sensor for L. monocytogenes. To determine the RNA receptor responsible for bacRNA-mediated type I IFN induction we used bone marrow-derived dendritic cells (BM-DC) of RIG-I or MDA5-deficient mice. Indeed, as shown using wild type, RIG-I or MDA5 deficient bone marrow-derived dendritic cells (BM-DC), the Listeria RNA-triggered IFN-α induction was exclusively RIG-I dependent (Fig. 4A). Treatment of A549 cells with siRNAs against RIG-I and MAVS abolished the response to the 3P-dsRNA RIG-I ligand (Fig. 4B) 12fold and fourfold, documenting an efficient knock-down of the RIG-I signaling pathway. Accordingly, as anticipated, siRNA mediated knock-down MAVS extensively inhibited type I IFN production of A549 cells during infection with L. monocytogenes (Fig. 4B; Fig. S4A). By contrast, in THP-1 cells, which can recognize bacDNA and bacRNA, type I IFN induction did not appear to be influenced by siRNA-mediated knockdown of MAVS during infection with L. monocytogenes (Fig. 4C), while the type I IFN response to the RIG-I ligand 3P-dsRNA was efficiently downregulated. The response to DNA was still intact after knock-down of MAVS, excluding the involvement of a polIII/RIG-I stimulatory effect of this DNA type in these cells. Currently, STING is the only known adaptor protein upstream of IRF3 in the DNA recognition signaling pathway and has been shown to be essential for the type I IFN response induced upon L. monocytogenes infection [16], [17], [42]. Knockdown of STING strongly inhibited the type I IFN induction response to plasmid DNA(pDNA) in THP-1 cells (Fig. 4C). In contrast to MAVS, knockdown of STING significantly reduced type I IFN induction during L. monocytogenes infection of THP-1 cells. We conclude that induction of type I IFN during Listeria infection of STING pathway deficient cells is dependent on RIG-I, while the RIG-I pathway is redundant in immune cells such as monocytes, as they possess a STING-dependent pathway and are therefore able to sense c-di-AMP and bacDNA in the cytosol in a pol III/RIG-I independent manner.
Figure 4. Knockdown of RIG-I abrogates L. monocytogenes-induced type I IFN induction in epithelial but not in monocytic cells.

A: Murine BM-DCs were transfected with indicated stimuli. One out of two experiments is shown. Murine IFN-α secretion was analyzed 24 hours after stimulation. Error bars represent SEM. B: A549 cells were transfected with siRNA against RIG-I, MAVS or Luciferase (control). Cells were then infected with L. monocytogenes or transfected with L. monocytogenes RNA (L.m. RNA), L. monocytogenes DNA (L.m. RNA) or 3P-dsRNA 48 hours after knock-down. Type I IFN production was analyzed 24 hours after stimulation. B) THP-1 cells were electroporated with control siRNA or siRNAs against MAVS or STING. 72 hours after electroporation, THP-1 cells were infected with hly- or wt L. monocytogenes at indicated MOI or transfected with plasmid DNA (pDNA) or RIG-I ligand (3P-dsRNA). Type I IFN production was analyzed 24 hours after stimulation. Relative type I IFN induction was normalized to cells transfected with siRNA against Luciferase and stimulated with pDNA. Error bars represent s.d.
Discussion
RIG-I is considered to be essential for the immune recognition of most negative strand and positive strand RNA viruses (reviewed in [56]). Little is known, however about the involvement of RIG-I and of bacterial RNA in the innate immune recognition of bacteria. Here we found that both the RNA extracted from extracellular and facultative intracellular bacteria induced type I IFN in a 5′-phosphorylation-dependent manner. We analyzed the effect of L. monocytogenes infection on immune cells and non-immune cells, both involved in the pathogenesis of L. monocytogenes. Infection with wt L. monocytogenes led to the induction of type I IFN in all cell lines tested, including monocytic cells and cell lines derived from lung (A549), intestine (Colo205) or liver (HepG2). Using a recently developed sensitive RNA fluorescence labeling technique [52] we demonstrate that during natural infection, the RNA of L. monocytogenes indeed gains access to the cytosol of the host cells. Both cytosolic translocation of bacRNA and type I IFN induction were dependent on the presence of the pore forming protein LLO. Although transfer of shRNA from LLO-overexpressing E. coli has been reported [57], this is the first time that translocation of endogenous bacterial RNA into the cytosol of the host cell could be visualized.
Previous studies on human cells revealed that LLO is not needed for egress from the vacuole. Paschen et al. [58] observed that in human dendritic cells lack of listeriolysin retarded but did not prevent egress of L. monocytogenes from the vacuole. It is tempting to speculate that LLO-deficient Listeria are at least impaired in entering the cytosol, leading to less or retarded secretion of RNA. The fact that secA2-deficient L. monocytogenes fails to translocate bacRNA into the cytosol confirm recent findings that RNA is released by secretion and not by lysis of bacteria [53]. Nevertheless, as suggested previously, cytosolic bacterial DNA or c-di-AMP released at the same time as bacRNA could represent an equal or even dominant source of type I IFN induction [13], [59]. However, we found that while transfected bacRNA and bacDNA induced equal amounts of type I IFN in monocytic cells (primary monocytes and THP-1 cells), non-immune cell types were able to sense transfected bacRNA but did not raise a type I IFN response upon transfection of bacDNA, despite a robust response to poly(dA-dT), indicating the absence of functional STING-dependent recognition pathways. The latter finding also excludes an involvement of the pol III/RIG-I dependent pathway of bacDNA detection as suggested by Chu et al. [24] but corroborates data of Monroe et al. [59] for L. pneumophila bacterial DNA, which was shown not to trigger the pol III/RIG-I dependent pathway.
Despite translocation of bacRNA into the cytosol, RNAi of RIG-I or MAVS in monocytic cells (THP-1) did not impair type I IFN induction during infection with L. monocytogenes. This suggests a redundant role of RIG-I regarding type I IFN induction in monocytes during L. monocytogenes infection, as shown for L. pneumophila [59]. The minor contribution of the RIG-I/MAVS pathway in monocytic cells may be due to a dominance in concentration or effect of other stimuli released by L. monocytogenes, including bacDNA or the recently discovered mediator cyclic diadenosine monophosphate (c-di-AMP) [34].
By contrast, induction of type I IFN during L. monocytogenes infection of epithelial cells was nearly completely abolished by RNAi-mediated knockdown of RIG-I or MAVS, implicating that neither DNA nor c-di-AMP plays a role for L. monocytogenes mediated type I IFN induction in these cells. Together, we conclude that RIG-I plays a major role in the L. monocytogenes-induced secretion of type-I-IFN in cell types lacking functional STING-dependent pathways.
Cytosolic recognition of bacterial RNA is controversial. Mancuso and colleagues [60] showed that phagosomal bacteria, but not cytosolic bacteria, potently induce IFN in conventional dendritic cells by a mechanism that required the RNA-sensing endosomal TLR 7. Although purified transfected L. monocytogenes RNA was observed to induce type I IFN [61], previous studies suggested that IFN induction by L. monocytogenes occurs in a MAVS-independent, and thereby cytosolic RIG-I independent way [29], [33]. However, in line with our current findings some bacterial RNA species represent a perfect RIG-I target structure: In contrast to eukaryotes, bacteria do not harbor 5′end mRNA cap structures but 5′mono or 5′triphosphate, a major requirement for RIG-I recognition [10]. In E. coli, one third of mRNA remains 5′triphosphorylated [39]. Recent studies reported that the 5′phosphorylation status of bacterial mRNA is regulated by the pyrophosphatase RppH [62] regulating mRNA decay [63]. The 5′triphosphate moiety was described to protect mRNA from decay by the bacterial RNase E [63]. Interestingly, the pyrophosphatase RppH was shown to strongly prefer dephosphorylation of single-stranded triphosphorylated 5′nucleotides over base paired ends [62]. This finding was correlated with the fact that bacterial RNAs can be stabilized by a 5′-terminal stem-loop [64], [65]. Therefore the occurrence of 5′base paired triphosphorylated RNA appears to be characteristic for bacteria and represents a pathogen-associated molecular pattern (PAMP). 5′end base paired triphosphorylated RNA was shown to be the ligand for RIG-I [11].
Indeed, studies on recognition of the intracellular bacterium Legionella pneumophila exhibited a MAVS-dependent pathway leading to IFN-β stimulation of lung epithelial cells (A549) [66]. More recent studies revealed that crude RNA isolated from Legionella pneumophila is recognized in a RIG-I dependent manner when transfected [59]. However, in that study infection with bacteria was examined in murine macrophages, which harbor several type I IFN inducing receptor systems, including direct STING-dependent DNA recognition. This complicates the evaluation of the contribution of RIG-I to the type I IFN response. In macrophages, the RIG-I pathway appeared to play a rather redundant role for immune recognition of intracellular bacteria [59]. Nonetheless, this finding might be cell type specific. So far, the impact of RIG-I during L. monocytogenes infection of other cell types than monocytes has not been addressed. In vivo, L. monocytogenes enters the body by crossing the intestinal barrier into the blood (reviewed in [40]). This involves bacterial invasion and survival within a large variety of non-phagocytic cells [41]. In immune-compromised individuals, multiplication can occur in hepatocytes.
Together, our current data show that RNA of the facultative intracellular bacterium L. monocytogenes has quantitative access to the cytosol of the host cell, and that detection of bacterial RNA by the cytosolic immune receptor RIG-I plays a non-redundant role in non-immune cells with impaired DNA/c-di-AMP (STING dependent) recognition pathway such as hepatocytes, a cell type involved in the propagation of bacteria in chronic infection, and colon epithelial cells, a cell type that is critically involved in the enteral route of infection. Interestingly, Li et al. found that RNA of commensal bacteria is recognized in a MAVS-dependent manner and that MAVS in cells of non-hematopoietic origin plays a dominant role in preventing DSS-induced colitis [61].
Considering the fact that mice with a defect in the type I IFN pathway exhibit a strong resistance to Listeria-induced pathogenesis, it remains to be determined if the observed RIG-I dependent recognition of bacterial RNA contributes more to the pathogenicity or to the clearance of Listeria [67], [68].
Materials and Methods
Ethics Statement
The PBMC studies were approved by the local ethics committee (Ethikkommission der Medizinischen Fakultät Bonn) according to the ICH-GCP guidelines. Written informed consent was provided by voluntary blood donors.
The accommodation and care of animals used for experimental and other scientific purposes occurred in the mouse core facility (HET) of the University Hospital Bonn according to the guidelines of the Federation of European Laboratory Animal Science Associations (FELASA) and GLP guidelines of the OECD.
Bacteria
The L. monocytogenes wild-type strain EGD and isogenic deletion mutant strains Δhly, ΔsecA were cultured in brain-heart infusion (BHI) until they reached log phase and then used for experiments.
Stimulatory Nucleic Acids
Bacterial RNA was isolated using the Qiagen RNeasy mini kit according to manufacturer’s instructions (Qiagen). Genomic DNA was isolated by lysing the cells with lysozyme and then precipitating the DNA using phenol-chloroform extraction. 3P-dsRNA was generated by in vitro transcription as described [11].
Cell Culture and Stimulation
Human PBMC were isolated from whole human blood of healthy, voluntary donors by Ficoll-Hypaque density gradient centrifugation (Biochrom). PBMCs were cultured in RPMI medium supplemented with 10% FCS and 1% penicillin and streptomycin. Cell lines were cultured in RPMI (THP-1, HepG2) or DMEM (Colo205) medium supplemented with 10% FCS and 1% Penicillin and Streptomycin. For stimulation cells were seeded into 96-well plates. Nucleic acid stimuli were used at a final concentration of 0,8 µg/mL. RNA and DNA stimuli were complexed with Lipofectamine 2000 (Invitrogen). Cell lines were incubated for 2 hours (THP-1) or 4 hours (A549, HepG2 and Colo205) with L. monocytogenes and then inactivated using Gentamycin (THP-1) or penicillin (for A549, HepG2 and Colo205). For infection of A549, HepG2 and Colo205 bacteria were pretreated with 10% human AB serum for 30 min at 37°C and washed with PBS.
Secretion of cytokines was measured using ELISA kits supplied by ebioscience (human IFN-α) and BD Pharmingen (human CXCL10). Murine IFN-α was determined by sandwich ELISA using standard protein and antibodies from R&D Systems and BIORAD. Human type I IFN activity was quantified by incubation of the type I IFN-sensing reporter cell line HEK-Blue™ (Invivogene) with supernatants of stimulated cells. After 24 h incubation, supernatants were assessed for SEAP activity using substrate pNPP (Sigma) according to the manufacturer’s protocol. RIG-I and MDA5-deficient mice were generated as described [7], [8]. Murine BM-DC were generated by culturing murine bone marrow cells for 7 days with GM-CSF.
RNAi in Cell Lines
A549 cells in 96 well plates were incubated with 0.1 µg siRNA against RIG-I, MAVS and Luciferase complexed to 1.5 µL Hiperfect (Qiagen; Fig. 4B) or 0.25 µL Lipofectamin2000 (Fig. S4A). Infection or transfection of stimuli into the cells was performed 36 (Fig. S4A) or 48 (Fig. 4B) hours after siRNA transfection. THP-1 cells were electroporated with siRNA 72 hours prior to stimulation. SiRNAs with 3′dTdT were purchased from Biomers (Ulm, Germany) and targeted the following sequences: STING #1: GCAUCAAGGAUCGGGUUU [42]; STING #2: GGUCAUAUUACAUCGGAU; MAVS #1: UUAAAGGAGUUUAUCGAUGUA; MAVS #2: CCCAGAGGAGAAUGAGUAUAA; RIG-I #1: AAGGCUGGUUCCGUGGCUUUU [23]; RIG-I #2: AAGGGAACGAUUCCAUCACU Controls were: Luciferase : CAUAAGGCAAUGAAGAGAUAG; nonsense#1: GAAGUCCUUAACGCGGCAA (Fig. 4C, Fig. S4A).
Labeling of RNA
L. monocytogenes were grown in an overnight culture and then 2 mL of HT-Medium [43] inoculated with 500 µL L. monocytogenes suspension. 1 mM EU (Invitrogen) was added and L. monocytogenes grown for 4 hours. Bacteria were labeled with 2 mg/mL FITC (Sigma-Aldrich) and washed with 5 µM EDTA/PBS, then resuspended in Opti-MEM prior to infection of host cells [44]. L. monocytogenes were used for infection at the appropriate MOIs, then washed off after 1 or 4 hours. Cells were washed with PBS and fixed with 4% PFA in PBS, then treated with Alexa 594 azide and the labeling kit Click-iT (Invitrogen) according to manufacturer’s instructions. Briefly, cells were permeabilized with 0.5% Triton X-100 in PBS, then blocked with 1% BSA and washed with PBS. The Click-iT reaction cocktail was assembled and mixed with Alexa 594 azide at 5 µM final concentration. Cells were incubated with the Alexa 594 azide-containing Click-iT reaction cocktail for 30 minutes at room temperature, then washed and treated for fluorescence microscopy.
For labeling of extracted bacterial RNA, overnight wt and hly L. monocytogenes cultures were diluted 1∶5 in Fraser Half Medium (FHM). Ethynyl Uridine (EU, Invitrogen) was added to a final concentration of 10 µM and bacteria grown for 4 hours at 37°C. Bacteria RNA was isolated using a modified protocol applied by Toledo-Arana [45]: Bacteria were pelleted and resuspended in solution A (10% w/v glucose, Tris 12.5 mM pH 7.6, 10 mM EDTA). 60 µL 0.5M EDTA and 60 µL of a 10 mg/mL Lysozyme solution were added and bacteria lysed for 2 hours at 37°C. RNA was isolated by standard phenol/chloroform extraction using TRIzol reagent (Invitrogen). RNA was incubated with the Click-iT reaction cocktail containing Alexa Fluor 488 azide. The quantification of EU presence in Listeria RNA was performed by measuring the fluorescence of Alexa 488 Fluor-stained RNA from hly- and wt Listeria in a Perkin-Elmer EnVision reader. As a control, equal amounts of RNA from L. monocytogenes grown in medium without EU was incubated in the Click-iT reaction cocktail. The background fluorescence of non-EU containing RNA after Click-iT reaction was subtracted from EU containing RNA and normalized on fluorescence of RNA from wt Listeria.
Fluorescence Microscopy
Freshly trypsinized and washed A549 or HepG2 were seeded at the appropriate concentrations onto uncoated coverslips, then left to adhere for 3 hours at 37°C. FITC-tagged and EU-labeled L. monocytogenes were then added to the cells and left for infection for the indicated duration. After fixation and treatment with the Click-iT reaction cocktail, cells were washed with PBS and counterstained with DAPI (Thermo scientific). THP-1 cells were infected and stained in solution and resuspended in mounting medium. Cells were then visualized using a fluorescence microscope (Zeiss).
Supporting Information
Effect of chloroquin incubation. Human PBMC were preincubated with chloroquine or left untreated and transfected with indicated nucleic acids. IFN-α production was analyzed 24 hours after stimulation.
(EPS)
Specificity of Click RNA labeling. A: E. coli bacteria were incubated with EU during log phase growth and EU labeled RNA stained with Alexa Fluor 594 azide. B: Whole RNA was extracted from wild type (wt) and LLO deficient (hly-) L. monocytogenes bacteria grown in EU-containing medium and coupled to Alexa Fluor 488 azide. Fluorescence of EU/Alexa Fluor 488 azide-labeled RNA was measured in solution. Background fluorescence of non-EU containing RNA after the Click-iT reaction was subtracted from EU containing RNA and normalized on fluorescence of RNA from wt L. monocytogenes. Mean values+s.d. (% of wt RNA) of three independent experiments are shown. THP-1 cells were grown for 12 hrs in starvation medium (RPMI without FCS) (C) or in regular medium (D) and then incubated for 4 hrs with equal concentrations of EU. Cells were fixed and EU-labeled RNA counterstained with Alexa 594.
(EPS)
Release of bacterial RNA into the cytosol depends on the SecA2 secretion system. THP-1 were infected with FITC-tagged and EU-labeled wild type (L.m.wt; left column) and SecA2 deficient (L.m. SecA-; left column) L. monocytogenes for two hours. Cells were then fixed, stained with Alexa594-azide and counterstained with DAPI. Whole L. monocytogenes are labeled green with FITC and RNA is visible as red fluorescence (Alexa594), nuclei are stained by DAPI (blue). Three examples per condition are shown. As determined by counting of single stained bacteria in cells (100 cells per slide were counted) the average bacterial load was 0.5(wt) and 0.5(SecA-) bacteria per cell. One representative experiment out of three is shown.
(EPS)
A: A549 cells were transfected with siRNA against RIG-I, MAVS or a control sequence. Cells were then infected with L. monocytogenes 40 hours after knock-down. Type I IFN production was analyzed 24 hours after stimulation. B: mRNA expression levels of RIG-I (left panel) or MAVS (right panel) normalized on GAPDH expression, 40 h after transfection of siRNAs into A549 cells. C: mRNA expression levels of STING (left panel) or MAVS (right panel) normalized of GAPDH expression, 72 h after electroporation. Error bars represent s.d.
(EPS)
Acknowledgments
We thank Jasper van den Boorn and Percy Knolle for critical reading of the manuscript and T. Chakraborty for providing SecA2 deficient L. monocytogenes.
Funding Statement
This study was supported by Grants BMBF Biofuture 0311896, BMBF-grant 01KI0771 (www.bmbf.de/) to GH and from the Deutsche Forschungsgemeinschaft (www.dfg.de; SFB670 to MS, GH and VH, SFB704 to GH, SFB832 and KFO177 to GH). GH, WB and VH are members of the DFG Excellence Cluster ImmunoSensation. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Chen G, Shaw MH, Kim YG, Nunez G (2009) NOD-like receptors: role in innate immunity and inflammatory disease. Annu Rev Pathol 4: 365–398. [DOI] [PubMed] [Google Scholar]
- 2. Schlee M, Hornung V, Hartmann G (2006) siRNA and isRNA: two edges of one sword. Mol Ther 14: 463–470. [DOI] [PubMed] [Google Scholar]
- 3. Krieg AM, Yi AK, Matson S, Waldschmidt TJ, Bishop GA, et al. (1995) CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 374: 546–549. [DOI] [PubMed] [Google Scholar]
- 4. Hemmi H, Takeuchi O, Kawai T, Kaisho T, Sato S, et al. (2000) A Toll-like receptor recognizes bacterial DNA. Nature 408: 740–745. [DOI] [PubMed] [Google Scholar]
- 5. Coch C, Busch N, Wimmenauer V, Hartmann E, Janke M, et al. (2009) Higher activation of TLR9 in plasmacytoid dendritic cells by microbial DNA compared with self-DNA based on CpG-specific recognition of phosphodiester DNA. J Leukoc Biol 86: 663–670. [DOI] [PubMed] [Google Scholar]
- 6. Yoneyama M, Kikuchi M, Natsukawa T, Shinobu N, Imaizumi T, et al. (2004) The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat Immunol 5: 730–737. [DOI] [PubMed] [Google Scholar]
- 7. Kato H, Takeuchi O, Sato S, Yoneyama M, Yamamoto M, et al. (2006) Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 441: 101–105. [DOI] [PubMed] [Google Scholar]
- 8. Gitlin L, Barchet W, Gilfillan S, Cella M, Beutler B, et al. (2006) Essential role of mda-5 in type I IFN responses to polyriboinosinic:polyribocytidylic acid and encephalomyocarditis picornavirus. Proc Natl Acad Sci U S A 103: 8459–8464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Pichlmair A, Schulz O, Tan CP, Naslund TI, Liljestrom P, et al. (2006) RIG-I-mediated antiviral responses to single-stranded RNA bearing 5′-phosphates. Science 314: 997–1001. [DOI] [PubMed] [Google Scholar]
- 10. Hornung V, Ellegast J, Kim S, Brzozka K, Jung A, et al. (2006) 5′-Triphosphate RNA is the ligand for RIG-I. Science 314: 994–997. [DOI] [PubMed] [Google Scholar]
- 11. Schlee M, Roth A, Hornung V, Hagmann CA, Wimmenauer V, et al. (2009) Recognition of 5′ triphosphate by RIG-I helicase requires short blunt double-stranded RNA as contained in panhandle of negative-strand virus. Immunity 31: 25–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Wang Y, Ludwig J, Schuberth C, Goldeck M, Schlee M, et al. (2010) Structural and functional insights into 5′-ppp RNA pattern recognition by the innate immune receptor RIG-I. Nat Struct Mol Biol 17: 781–787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Stetson DB, Medzhitov R (2006) Recognition of cytosolic DNA activates an IRF3-dependent innate immune response. Immunity 24: 93–103. [DOI] [PubMed] [Google Scholar]
- 14. Ishii KJ, Coban C, Kato H, Takahashi K, Torii Y, et al. (2006) A Toll-like receptor-independent antiviral response induced by double-stranded B-form DNA. Nat Immunol 7: 40–48. [DOI] [PubMed] [Google Scholar]
- 15. Cavlar T, Ablasser A, Hornung V (2012) Induction of type I IFNs by intracellular DNA-sensing pathways. Immunol Cell Biol 90: 474–482. [DOI] [PubMed] [Google Scholar]
- 16. Ishikawa H, Ma Z, Barber GN (2009) STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 461: 788–792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Zhong B, Yang Y, Li S, Wang YY, Li Y, et al. (2008) The adaptor protein MITA links virus-sensing receptors to IRF3 transcription factor activation. Immunity 29: 538–550. [DOI] [PubMed] [Google Scholar]
- 18. Sun L, Wu J, Du F, Chen X, Chen ZJ (2012) Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 339: 786–791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Wu J, Sun L, Chen X, Du F, Shi H, et al. (2012) Cyclic GMP-AMP is an endogenous second messenger in innate immune signaling by cytosolic DNA. Science 339: 826–830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hornung V, Ablasser A, Charrel-Dennis M, Bauernfeind F, Horvath G, et al.. (2009) AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating inflammasome with ASC. Nature. [DOI] [PMC free article] [PubMed]
- 21.Fernandes-Alnemri T, Yu JW, Datta P, Wu J, Alnemri ES (2009) AIM2 activates the inflammasome and cell death in response to cytoplasmic DNA. Nature. [DOI] [PMC free article] [PubMed]
- 22.Burckstummer T, Baumann C, Bluml S, Dixit E, Durnberger G, et al.. (2009) An orthogonal proteomic-genomic screen identifies AIM2 as a cytoplasmic DNA sensor for the inflammasome. Nat Immunol. [DOI] [PubMed]
- 23.Ablasser A, Bauernfeind F, Hartmann G, Latz E, Fitzgerald KA, et al.. (2009) RIG-I-dependent sensing of poly(dA:dT) through the induction of an RNA polymerase III-transcribed RNA intermediate. Nat Immunol. [DOI] [PMC free article] [PubMed]
- 24. Chiu YH, Macmillan JB, Chen ZJ (2009) RNA polymerase III detects cytosolic DNA and induces type I interferons through the RIG-I pathway. Cell 138: 576–591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Kawai T, Takahashi K, Sato S, Coban C, Kumar H, et al. (2005) IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction. Nat Immunol 6: 981–988. [DOI] [PubMed] [Google Scholar]
- 26. Meylan E, Curran J, Hofmann K, Moradpour D, Binder M, et al. (2005) Cardif is an adaptor protein in the RIG-I antiviral pathway and is targeted by hepatitis C virus. Nature 437: 1167–1172. [DOI] [PubMed] [Google Scholar]
- 27. Seth RB, Sun L, Ea CK, Chen ZJ (2005) Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-kappaB and IRF 3. Cell 122: 669–682. [DOI] [PubMed] [Google Scholar]
- 28. Xu LG, Wang YY, Han KJ, Li LY, Zhai Z, et al. (2005) VISA is an adapter protein required for virus-triggered IFN-beta signaling. Mol Cell 19: 727–740. [DOI] [PubMed] [Google Scholar]
- 29. Sun Q, Sun L, Liu HH, Chen X, Seth RB, et al. (2006) The specific and essential role of MAVS in antiviral innate immune responses. Immunity 24: 633–642. [DOI] [PubMed] [Google Scholar]
- 30. O’Riordan M, Yi CH, Gonzales R, Lee KD, Portnoy DA (2002) Innate recognition of bacteria by a macrophage cytosolic surveillance pathway. Proc Natl Acad Sci U S A 99: 13861–13866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Stockinger S, Materna T, Stoiber D, Bayr L, Steinborn R, et al. (2002) Production of type I IFN sensitizes macrophages to cell death induced by Listeria monocytogenes. J Immunol 169: 6522–6529. [DOI] [PubMed] [Google Scholar]
- 32. Stockinger S, Reutterer B, Schaljo B, Schellack C, Brunner S, et al. (2004) IFN regulatory factor 3-dependent induction of type I IFNs by intracellular bacteria is mediated by a TLR- and Nod2-independent mechanism. J Immunol 173: 7416–7425. [DOI] [PubMed] [Google Scholar]
- 33. Soulat D, Bauch A, Stockinger S, Superti-Furga G, Decker T (2006) Cytoplasmic Listeria monocytogenes stimulates IFN-beta synthesis without requiring the adapter protein MAVS. FEBS Lett 580: 2341–2346. [DOI] [PubMed] [Google Scholar]
- 34. Woodward JJ, Iavarone AT, Portnoy DA (2010) c-di-AMP secreted by intracellular Listeria monocytogenes activates a host type I interferon response. Science 328: 1703–1705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Sauer JD, Sotelo-Troha K, von Moltke J, Monroe KM, Rae CS, et al. (2011) The N-ethyl-N-nitrosourea-induced Goldenticket mouse mutant reveals an essential function of Sting in the in vivo interferon response to Listeria monocytogenes and cyclic dinucleotides. Infect Immun 79: 688–694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Rathinam VA, Jiang Z, Waggoner SN, Sharma S, Cole LE, et al. (2010) The AIM2 inflammasome is essential for host defense against cytosolic bacteria and DNA viruses. Nat Immunol 11: 395–402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Wu J, Fernandes-Alnemri T, Alnemri ES (2010) Involvement of the AIM2, NLRC4, and NLRP3 inflammasomes in caspase-1 activation by Listeria monocytogenes. J Clin Immunol 30: 693–702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Warren SE, Armstrong A, Hamilton MK, Mao DP, Leaf IA, et al. (2010) Cutting edge: Cytosolic bacterial DNA activates the inflammasome via Aim2. J Immunol 185: 818–821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Bieger CD, Nierlich DP (1989) Distribution of 5′-triphosphate termini on the mRNA of Escherichia coli. J Bacteriol 171: 141–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Pizarro-Cerda J, Kuhbacher A, Cossart P (2012) Entry of Listeria monocytogenes in mammalian epithelial cells: an updated view. Cold Spring Harb Perspect Med 2. [DOI] [PMC free article] [PubMed]
- 41. Seveau S, Pizarro-Cerda J, Cossart P (2007) Molecular mechanisms exploited by Listeria monocytogenes during host cell invasion. Microbes Infect 9: 1167–1175. [DOI] [PubMed] [Google Scholar]
- 42. Ishikawa H, Barber GN (2008) STING is an endoplasmic reticulum adaptor that facilitates innate immune signalling. Nature 455: 674–678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Tsai HN, Hodgson DA (2003) Development of a synthetic minimal medium for Listeria monocytogenes. Appl Environ Microbiol 69: 6943–6945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Popov A, Abdullah Z, Wickenhauser C, Saric T, Driesen J, et al. (2006) Indoleamine 2,3-dioxygenase-expressing dendritic cells form suppurative granulomas following Listeria monocytogenes infection. J Clin Invest 116: 3160–3170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Toledo-Arana A, Dussurget O, Nikitas G, Sesto N, Guet-Revillet H, et al. (2009) The Listeria transcriptional landscape from saprophytism to virulence. Nature 459: 950–956. [DOI] [PubMed] [Google Scholar]
- 46. Rutz M, Metzger J, Gellert T, Luppa P, Lipford GB, et al. (2004) Toll-like receptor 9 binds single-stranded CpG-DNA in a sequence- and pH-dependent manner. Eur J Immunol 34: 2541–2550. [DOI] [PubMed] [Google Scholar]
- 47. Clement S, Vaudaux P, Francois P, Schrenzel J, Huggler E, et al. (2005) Evidence of an intracellular reservoir in the nasal mucosa of patients with recurrent Staphylococcus aureus rhinosinusitis. J Infect Dis 192: 1023–1028. [DOI] [PubMed] [Google Scholar]
- 48. Choi CH, Lee JS, Lee YC, Park TI, Lee JC (2008) Acinetobacter baumannii invades epithelial cells and outer membrane protein A mediates interactions with epithelial cells. BMC Microbiol 8: 216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Mengaud J, Ohayon H, Gounon P, Mege RM, Cossart P (1996) E-cadherin is the receptor for internalin, a surface protein required for entry of L. monocytogenes into epithelial cells. Cell 84: 923–932. [DOI] [PubMed] [Google Scholar]
- 50. Lecuit M, Dramsi S, Gottardi C, Fedor-Chaiken M, Gumbiner B, et al. (1999) A single amino acid in E-cadherin responsible for host specificity towards the human pathogen Listeria monocytogenes. EMBO J 18: 3956–3963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Gregory SH, Barczynski LK, Wing EJ (1992) Effector function of hepatocytes and Kupffer cells in the resolution of systemic bacterial infections. J Leukoc Biol 51: 421–424. [DOI] [PubMed] [Google Scholar]
- 52. Jao CY, Salic A (2008) Exploring RNA transcription and turnover in vivo by using click chemistry. Proc Natl Acad Sci U S A 105: 15779–15784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Abdullah Z, Schlee M, Roth S, Mraheil MA, Barchet W, et al. (2012) RIG-I detects infection with live Listeria by sensing secreted bacterial nucleic acids. EMBO J 31: 4153–4164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Ishii KJ, Akira S (2006) Innate immune recognition of, and regulation by, DNA. Trends Immunol 27: 525–532. [DOI] [PubMed] [Google Scholar]
- 55. Cheng G, Zhong J, Chung J, Chisari FV (2007) Double-stranded DNA and double-stranded RNA induce a common antiviral signaling pathway in human cells. Proc Natl Acad Sci U S A 104: 9035–9040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Schlee M, Hartmann G (2010) The Chase for the RIG-I Ligand-Recent Advances. Mol Ther 18: 1254–1262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Xiang S, Fruehauf J, Li CJ (2006) Short hairpin RNA-expressing bacteria elicit RNA interference in mammals. Nat Biotechnol 24: 697–702. [DOI] [PubMed] [Google Scholar]
- 58. Paschen A, Dittmar KE, Grenningloh R, Rohde M, Schadendorf D, et al. (2000) Human dendritic cells infected by Listeria monocytogenes: induction of maturation, requirements for phagolysosomal escape and antigen presentation capacity. Eur J Immunol 30: 3447–3456. [DOI] [PubMed] [Google Scholar]
- 59. Monroe KM, McWhirter SM, Vance RE (2009) Identification of host cytosolic sensors and bacterial factors regulating the type I interferon response to Legionella pneumophila. PLoS Pathog 5: e1000665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Mancuso G, Gambuzza M, Midiri A, Biondo C, Papasergi S, et al. (2009) Bacterial recognition by TLR7 in the lysosomes of conventional dendritic cells. Nat Immunol 10: 587–594. [DOI] [PubMed] [Google Scholar]
- 61. Li XD, Chiu YH, Ismail AS, Behrendt CL, Wight-Carter M, et al. (2011) Mitochondrial antiviral signaling protein (MAVS) monitors commensal bacteria and induces an immune response that prevents experimental colitis. Proc Natl Acad Sci U S A 108: 17390–17395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Deana A, Celesnik H, Belasco JG (2008) The bacterial enzyme RppH triggers messenger RNA degradation by 5′ pyrophosphate removal. Nature 451: 355–358. [DOI] [PubMed] [Google Scholar]
- 63. Celesnik H, Deana A, Belasco JG (2007) Initiation of RNA decay in Escherichia coli by 5′ pyrophosphate removal. Mol Cell 27: 79–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Emory SA, Bouvet P, Belasco JG (1992) A 5′-terminal stem-loop structure can stabilize mRNA in Escherichia coli. Genes Dev 6: 135–148. [DOI] [PubMed] [Google Scholar]
- 65. Mackie GA (2000) Stabilization of circular rpsT mRNA demonstrates the 5′-end dependence of RNase E action in vivo. J Biol Chem 275: 25069–25072. [DOI] [PubMed] [Google Scholar]
- 66. Opitz B, Vinzing M, van Laak V, Schmeck B, Heine G, et al. (2006) Legionella pneumophila induces IFNbeta in lung epithelial cells via IPS-1 and IRF3, which also control bacterial replication. J Biol Chem 281: 36173–36179. [DOI] [PubMed] [Google Scholar]
- 67. O’Connell RM, Saha SK, Vaidya SA, Bruhn KW, Miranda GA, et al. (2004) Type I interferon production enhances susceptibility to Listeria monocytogenes infection. J Exp Med 200: 437–445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Auerbuch V, Brockstedt DG, Meyer-Morse N, O’Riordan M, Portnoy DA (2004) Mice lacking the type I interferon receptor are resistant to Listeria monocytogenes. J Exp Med 200: 527–533. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Effect of chloroquin incubation. Human PBMC were preincubated with chloroquine or left untreated and transfected with indicated nucleic acids. IFN-α production was analyzed 24 hours after stimulation.
(EPS)
Specificity of Click RNA labeling. A: E. coli bacteria were incubated with EU during log phase growth and EU labeled RNA stained with Alexa Fluor 594 azide. B: Whole RNA was extracted from wild type (wt) and LLO deficient (hly-) L. monocytogenes bacteria grown in EU-containing medium and coupled to Alexa Fluor 488 azide. Fluorescence of EU/Alexa Fluor 488 azide-labeled RNA was measured in solution. Background fluorescence of non-EU containing RNA after the Click-iT reaction was subtracted from EU containing RNA and normalized on fluorescence of RNA from wt L. monocytogenes. Mean values+s.d. (% of wt RNA) of three independent experiments are shown. THP-1 cells were grown for 12 hrs in starvation medium (RPMI without FCS) (C) or in regular medium (D) and then incubated for 4 hrs with equal concentrations of EU. Cells were fixed and EU-labeled RNA counterstained with Alexa 594.
(EPS)
Release of bacterial RNA into the cytosol depends on the SecA2 secretion system. THP-1 were infected with FITC-tagged and EU-labeled wild type (L.m.wt; left column) and SecA2 deficient (L.m. SecA-; left column) L. monocytogenes for two hours. Cells were then fixed, stained with Alexa594-azide and counterstained with DAPI. Whole L. monocytogenes are labeled green with FITC and RNA is visible as red fluorescence (Alexa594), nuclei are stained by DAPI (blue). Three examples per condition are shown. As determined by counting of single stained bacteria in cells (100 cells per slide were counted) the average bacterial load was 0.5(wt) and 0.5(SecA-) bacteria per cell. One representative experiment out of three is shown.
(EPS)
A: A549 cells were transfected with siRNA against RIG-I, MAVS or a control sequence. Cells were then infected with L. monocytogenes 40 hours after knock-down. Type I IFN production was analyzed 24 hours after stimulation. B: mRNA expression levels of RIG-I (left panel) or MAVS (right panel) normalized on GAPDH expression, 40 h after transfection of siRNAs into A549 cells. C: mRNA expression levels of STING (left panel) or MAVS (right panel) normalized of GAPDH expression, 72 h after electroporation. Error bars represent s.d.
(EPS)