

Abstract
Enhanced perception of detail has long been regarded a hallmark of autism spectrum conditions (ASC), but its origins are unknown. Normal sensitivity on all fundamental perceptual measures—visual acuity, contrast discrimination, and flicker detection—is strongly established in the literature. If individuals with ASC do not have superior low-level vision, how is perception of detail enhanced? We argue that this apparent paradox can be resolved by considering visual attention, which is known to enhance basic visual sensitivity, resulting in greater acuity and lower contrast thresholds. Here, we demonstrate that the focus of attention and concomitant enhancement of perception are sharper in human individuals with ASC than in matched controls. Using a simple visual acuity task embedded in a standard cueing paradigm, we mapped the spatial and temporal gradients of attentional enhancement by varying the distance and onset time of visual targets relative to an exogenous cue, which obligatorily captures attention. Individuals with ASC demonstrated a greater fall-off in performance with distance from the cue than controls, indicating a sharper spatial gradient of attention. Further, this sharpness was highly correlated with the severity of autistic symptoms in ASC, as well as autistic traits across both ASC and control groups. These findings establish the presence of a form of “tunnel vision” in ASC, with far-reaching implications for our understanding of the social and neurobiological aspects of autism.
Introduction
Atypical perception is a defining characteristic of autism spectrum conditions (ASC), recently proposed as a diagnostic criterion in the DSM-V. In particular, autistic visual experience is marked by superior perception of local details and inefficient integration of information across space (Shah and Frith, 1983). However, autistic individuals do not evidence superior local visual sensitivity on measures such as visual acuity (Kéïta et al., 2010; Tavassoli et al., 2011), contrast discrimination (De Jonge et al., 2007; Koh et al., 2010), and flicker detection (Bertone et al., 2005; Pellicano et al., 2005), leaving unresolved the question of how quick and accurate perception of detail arises. In this study, we investigated whether differences in attention could account for the enhanced perception of local details in ASC.
Attention is the mechanism by which we orient our perception of the visual world, enabling us to efficiently parse and process complex scenes. For example, the sudden onset of a stimulus not only draws focus to a point in space reflected in improved detection of stimuli, but also increases in acuity and contrast sensitivity in nearby locations (Cameron et al., 2002; Golla et al., 2004). Importantly, this enhancement extends across the visual field along a two-dimensional gradient (Mangun and Hillyard, 1988), facilitating the processing of some stimuli over others, in effect organizing the contents of visual perception (Kravitz and Behrmann, 2011).
The aim of this study was to establish whether the enhanced perception of details in ASC reflects sharper spatial and temporal gradients of attention than in matched controls. Specifically, we varied the distance and time of onset of a target relative to an exogenous cue. We found that individuals with ASC have a sharper spatial gradient of attention with a strong relationship between the sharpness of the spatial gradient and higher-order autistic symptomatology, suggesting that the perturbation in attention is a useful index of, or contributor to, higher-level perceptual deficits in ASC. Because the spatial gradient of attention is thought to closely reflect neural circuitry (Brefczynski-Lewis et al., 2009), these results may also provide insight into atypical neural organization in the autistic brain. Overall, these findings demonstrate a fundamental alteration in the distribution of attention in individuals with ASC and provide a new quantitative and low-level index of high-level autistic symptomatology.
Materials and Methods
Participants.
Twenty adult controls (5 male, 15 female) and 22 adults with high-functioning autism (13 male, 7 female) were recruited from Cambridge's Autism Research Centre volunteer database, autism clinics, and support centers throughout the United Kingdom. All patients met international criteria for ASC according to the DSM-IV, as judged by clinicians specialized in the diagnosis of ASC. The two groups were matched for age (p > 0.23) and nonverbal IQ (p > 0.59) (Table 1). Participants had normal or corrected-to-normal vision and individuals with other psychiatric conditions, such as attention deficit-hyperactivity disorder, were not recruited. Written consent was obtained from all participants in accordance with a protocol approved by Cambridge's Psychology Research Ethics Committee.
Table 1.
Psychometric data
| N | Minimum | Maximum | Mean | SD | |
|---|---|---|---|---|---|
| ASC | |||||
| Age | 20 | 19 | 46 | 32.60 | 8.255 |
| IQ | 20 | 86.00 | 140.00 | 114.55 | 14.288 |
| Gender | 13M:7F | — | — | — | — |
| Control | |||||
| Age | 20 | 22 | 52 | 30.10 | 7.847 |
| IQ | 20 | 102.00 | 133.00 | 117.95 | 8.899 |
| Gender | 5M:15F | — | — | — | — |
Psychometric testing.
All participants completed the Wechsler Abbreviated Scale of Intelligence (WASI) (Wechsler, 1999). The ASC and control groups were matched in nonverbal IQ (Table 1). One ASC participant was excluded due to an inability to complete the IQ test. All participants also completed the Autism Spectrum Quotient (AQ), a self-report questionnaire which quantifies autistic traits across both ASC and control populations (Baron-Cohen et al., 2001). Additionally, an hour-long diagnostic protocol was administered to all ASC participants (Autism Diagnostic Observation Schedule, ADOS) (Lord et al., 2000).
Stimulus presentation.
Stimuli were presented using Psychtoolbox (Brainard, 1997) on a MacBook Pro and presented on a 17-inch TFT-LCD display (1280 × 1024 resolution). All testing took place in a darkened room with viewing distance from screen center (57 cm) fixed using a chin rest. Auditory feedback was provided for incorrect responses.
Procedure.
On each trial, participants fixated on a cross at the center of the screen and were briefly presented with a peripheral circular target (radius, 0.5°; duration,67 ms) and asked to report whether a small gap (0.4°) was on the top or bottom (Fig. 1). The contrast of the target was set to each individual's 75% correct detection threshold, obtained by a standard staircase procedure after thorough training on the task (see Thresholding, below).
Figure 1.

Procedure used to map attentional gradients. A, Time course of one trial. Each trial began with a fixation cross followed by a brief cue that captured participants' attention to a discrete point on the horizontal meridian to the left or right of fixation. After a variable ISI (67, 135, or 210 ms), a target appeared briefly, on the same side of the screen as the cue. Simultaneously, a distractor circle was presented 180° from the target. Participants indicated whether the opening was on the top or bottom of the target. B, Stimulus locations and sizes. The numbers next to the possible target locations give their distances in degrees from the cue location.
To measure the spatial and temporal gradients of attentional enhancement, this visual acuity task was embedded in an exogenous attention paradigm (Fig. 1a). Specifically, before presentation of the target, a brief cue (radius: 0.125°, duration: 67 ms) was presented, capturing participants' attention to a point on the screen's horizontal meridian. This point was at the minimum mean Euclidean distance from all possible target locations (see below). Participants were instructed to use the cue covertly and maintain fixation. Following this cue there was a variable delay (interstimulus interval (ISI); short, 67; medium, 135; or long, 210 ms) before the target was presented in one of three possible isoeccentric locations above or below the horizontal meridian in the same hemifield as the cue (distance from the cue: near, 2.46; mid, 4.51; or far, 6.56°) (Fig. 1b). A distractor stimulus, a circle identical to the target but without a gap, was presented 180° from the target in the opposite quadrant of the visual field to strengthen the effect of the cue. Participants' response accuracy and reaction time were recorded on each trial.
Before the experiment, participants were familiarized with the task through both verbal description and a slow-motion demonstration in which the target remained on the screen until response (10 trials). Each participant's individual contrast threshold was then established (see Thresholding, below) before a complete run of the task was practiced at full speed (96 trials). Performance was evaluated to determine the stability of the detection threshold. If between 65 and 85% accuracy at the farthest cue position, the experiment continued. Otherwise, participants repeated the thresholding run and began the experiment with the newly obtained threshold. One participant (ASC) whose accuracy during the experiment was lower than 65% correct at the threshold location was excluded from further analyses.
Following the practice run, participants completed 12 blocks of 96 trials each, resulting in 128 samples per distance (collapsed across the four quadrants of the screen) at each ISI.
Thresholding.
During thresholding, target-contrast was adaptively determined by response on each trial to obtain a 75% correct contrast threshold measurement at the longest ISI and farthest spatial location from the cue tested in the experiment. Using a standard staircase procedure consisting of five large reversals (step-size, 7.5% of the previous contrast) and 10 smaller reversals (step-size, 2.5% of the previous contrast), each correct response lowered the target contrast by one step, and each incorrect response raised the target contrast by three. The mean of the last seven reversals was taken as the participant's 75% contrast threshold.
Gray-level contrast thresholds were defined as follows: (target luminance − background luminance)/(target luminance + background luminance). Two participants had contrast thresholds more than two SDs outside the mean (one control, one ASC). Analyses were performed both with and without these participants; they did not qualitatively influence the results and were included in the analyses reported here.
Thresholding allowed us to objectively match participants on baseline difficulty in the task, so that any effects observed in our experiment could be attributed to attentional enhancement rather than visual sensitivity. Confirmation this aim was successful was manifest in the absence of any difference in accuracy between ASC and control participants, both overall (F(2,37) = 1.255, p < 0.270) and at only the ISI and distance at which threshold was performed (p < 0.126).
Performance analysis.
To investigate the spatial and temporal distribution of attention, we calculated a combined measure of individual accuracy and response times that is commonly used to account for individual speed-accuracy tradeoffs in attention studies, inverse efficiency scores (Townsend and Ashby, 1978). Speed-accuracy trade-offs are routinely observed in spatial cuing paradigms (Chica et al., 2011), where the instruction to respond both quickly and with a high degree of accuracy leads to individual variation in which measure best reflects task difficulty. Specifically, inverse efficiency scores were computed as (−1 × median reaction time/accuracy), with higher scores reflecting more efficient processing. This performance measure was used in all subsequent ANOVAs describing the spatial gradient of attention. Qualitatively similar results were obtained when using reaction time alone as a measure of performance. In all statistical analyses, Greenhouse–Geisser corrections were used for nonspherical data.
Eye tracking and gaze analysis.
The LCD display had a built-in binocular eye-tracker (Tobii Technology; sampling rate, 60 Hz; spatial precision, 0.5°), which was used to monitor the fixation stability of each participant. Participants successfully completed a 9-point calibration routine with an error threshold of 0.92° before the first experimental run. Fixation data from the right eye were analyzed using custom MATLAB (MathWorks) scripts. Data from time points during which the eye-tracker did not receive input from the eye (e.g., blinks) were removed.
Results
Comparable contrast sensitivity
The thresholding procedure confirmed the previously reported similarity in contrast thresholds in ASC and controls (14 and 10%, respectively; p > 0.20). Thresholds correlated with neither intelligence (WASI, ρ(40) = −0.21; p > 0.19), nor autistic symptomatology (AQ, ρ(40) = 0.22, p < 0.17; ADOS, ρ(40) = 0.18, p < 0.31).
Sharper gradient of attention in ASC
As expected, in both ASC and controls, performance decreased with distance from the cue, reflecting the spatial gradient of attention (Fig. 2a). To test for differences in the temporal and spatial gradients of attention, we conducted a three-way ANOVA with distance (near, mid, far) and ISI (short, medium, long) as repeated measures factors and diagnosis as a between-subjects factor.
Figure 2.
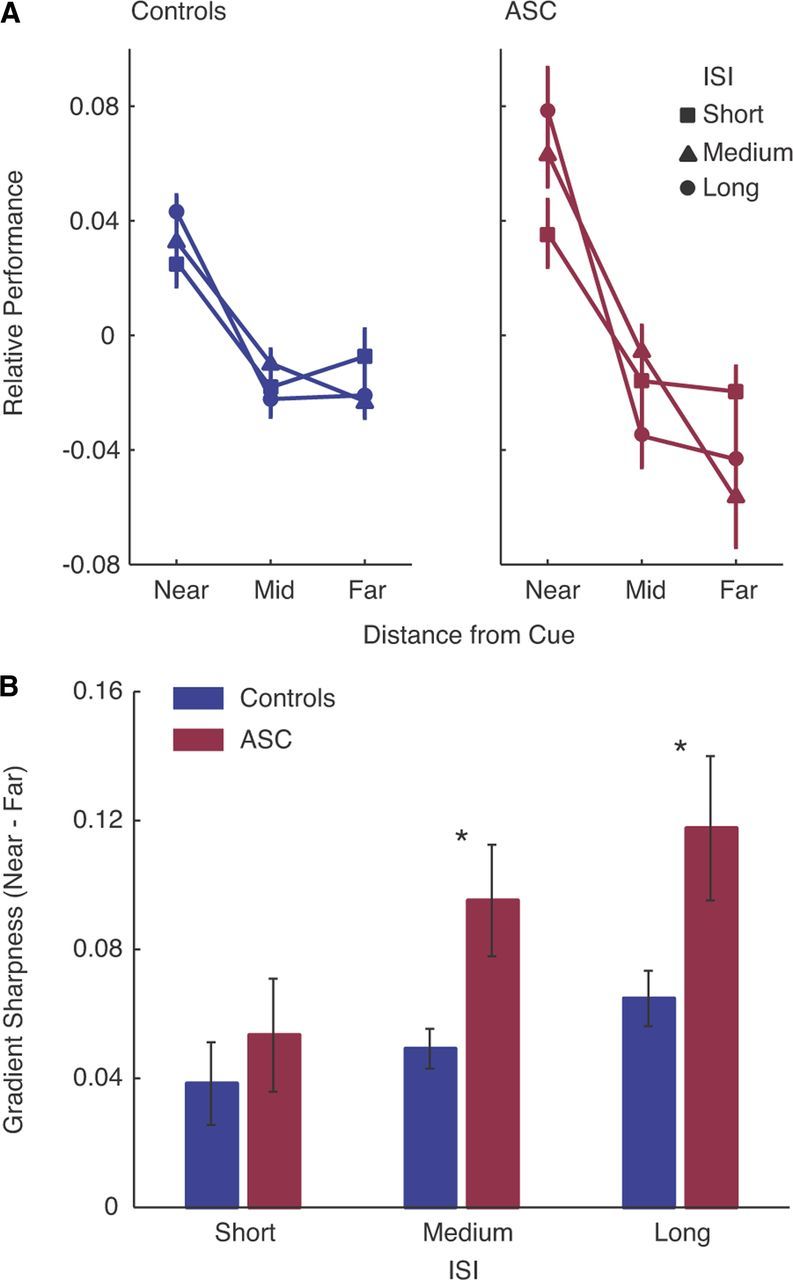
Sharper gradient of spatial attention in autism. A, Performance was measured as negative inverse efficiency scores [−1 × (reaction time/accuracy)]. To highlight the spatial gradient of attention for the purpose of the plot, performance scores were normalized by subtracting the mean performance for each combination of diagnosis and ISI. All statistics were calculated without this normalization being applied. In both groups, performance was better the closer the target was to the cue. This fall-off was much stronger in participants with ASC than controls, demonstrating a much sharper gradient of attentional enhancement. B, Sharpness of the gradient across ISIs. The slope of the attentional function increased with ISI in both groups. This effect was more pronounced in ASC, although the interaction between ISI and diagnosis did not reach significance. Error bar, SEM. *p < 0.05.
In terms of the spatial distribution of attention, the ANOVA revealed a highly significant main effect of distance (F(1,76) = 62.065, p < 0.001), indicating the expected fall-off in performance with increasing distance from the cue. Critically, this effect was exaggerated in individuals with ASC, resulting in a highly significant interaction between diagnosis and distance (F(2,76) = 5.051, p < 0.009). This finding indicates a much sharper spatial gradient of attention in ASC compared with control participants (Fig. 2a).
In terms of the temporal distribution of attention, the ANOVA revealed a highly significant main effect of ISI (F(1,76) = 5.564, p < 0.006), resulting from the expected better performance with longer ISIs. We also observed a significant interaction between ISI and distance (F(4,152) = 4.36, p < 0.009), reflecting an increase in the sharpness of the gradient with increasing ISI in both ASC and controls. The evolution of this sharpness with ISI suggests that the gradient reflects the distribution of spatial attention, rather than a general performance field difference (Abrams et al., 2012). Finally, there were no significant interactions involving ISI and diagnosis (all p values > 0.57), although a significant main effect of diagnosis was also observed (F(1,38) = 5.848, p < 0.02), indicating slightly poorer performance in the ASC group overall.
To quantify the form of the interaction between distance and diagnosis and to estimate the overall sharpness of the attention gradient in each participant, we computed gradient difference scores for each ISI separately (near–far distance) (Fig. 2b). These scores were used for all further psychometric correlation analyses (see below). A two-way ANOVA of gradient difference scores, with ISI as a repeated measures factor and Diagnosis as a between-subjects factor, revealed a main effect of diagnosis (F(1,38) = 9.207, p < 0.007), confirming the sharper spatial gradient of attention in ASC, and of ISI (F(2,76) = 4.060, p < 0.031), resulting from the increase in sharpness in both groups at longer ISIs. This exaggerated sharpness with increasing ISI was particularly evident in ASC. Post hoc one-tailed t tests revealed significantly larger gradient difference scores in the ASC group at the medium (t = 2.299, p < 0.013) and long (t = 1.932, p < 0.031) ISIs, with no difference at the short ISI (t = −0.833, p < 0.410), though this analysis of gradient difference scores did not produce a significant interaction between diagnosis and ISI (p > 0.59 in the main analysis). Although this interaction did not reach significance, it is possible that the spatial gradient in the two groups is similar at short ISIs and particularly sharper in ASC at longer ISIs. This effect would suggest a temporal component to the evolution of the difference between the groups, but requires further study with a broader range of ISIs. Gradient difference scores remained greater in the ASC group when age, IQ, or gender was treated as a covariate of no interest (all p < 0.027).
This increased sharpness of the spatial gradient in the ASC group was also evident in reaction time, illustrated by a main effect of diagnosis in a two-way ANOVA of mean reaction time difference scores (F(1,38) = 4.824, p < 0.034; controls, 25 ms; ASC, 44 ms). There was also a numerical difference in accuracy difference scores, suggesting a slightly sharper drop in accuracy with distance, but this did not reach significance (p > 0.1; controls, 3.1%; ASC, 4.2%). Overall, mean reaction times were slightly slower in ASC [t = 2.11, p < 0.041; controls, 550 ms (±70); ASC, 660 ms (±20)], and accuracy was comparable between the two groups [t = 1.11, p > 0.1; controls, 81% (±0.1); ASC, 84% (±0.07)].
Sharpness of the attentional gradient predicts autistic symptomatology
Not only did individuals with ASC display a sharper spatial gradient of visual attention as a group, the overall sharpness of this function was highly associated with measures of autistic symptoms and traits across individuals in both the ASC and control groups. First, there was a striking correlation between the sharpness of the attentional gradient and the ADOS scores of individuals with ASC (Fig. 3a) (ρ(20) = 0.527, p < 0.017). Second, sharpness was also strongly correlated with autistic traits across both the ASC and control groups, as measured by the AQ (Fig. 3b) (ρ(40) = 0.501, p < 0.001). These results show that this relatively low-level attentional marker of ASC is associated with symptomatology defined at much more complex levels of behavior.
Figure 3.

Sharpness of the attentional gradient correlates with autistic symptoms and traits in both ASC and controls. A, ASC participants with higher ADOS scores (an interview-based measure of autistic traits) had sharper attentional gradients averaged across ISIs (p < 0.01). B, Across both groups (ASC, red; controls, blue), participants with higher AQ scores (a social survey-based measure of autistics traits) evidence sharper attentional gradients (p < 0.001).
Results cannot be explained by differences in gaze-position
There were no quantitative differences in gaze-positions between the two groups [degrees from fixation: controls (x, 0.17 ± 1.17°; y, 0.23 ± 0.72°); ASC (x, 0.61 ± 2.01°; y, 0.07 ± 0.77°)]. Repeated-measures ANOVA performed separately on participants' mean x and y gaze-positions during target presentation for each trial-type, with ISI, distance, and target hemifield as within-group factors and diagnosis as a between-group factor, revealed only a main effect of target hemifield for the x-coordinates (p < 0001), as participants' gaze-positions were slightly biased toward the side of the screen on which the cue/target appeared. This effect was slight: mean gaze-position fell within 1° of the fixation point for both groups, regardless whether the cue/target was in the left or right hemifield. No other main effects or interactions involving ISI, distance, target hemifield, or diagnosis (all p > 0.359) were observed. Further, gradient difference scores remained larger in the ASC group when any trials during which gaze-position was >1.5° from fixation were excluded (F(1,38) = 5.220, p < 0.028).
Discussion
We have demonstrated that the spatial gradient over which attention enhances visual processing is much sharper in autistic than in matched control participants. Further, the degree of sharpness is associated both with the severity of autistic symptoms in people with ASC and with autistic traits across both ASC individuals and controls.
A sharper distribution of spatial attention is likely to dramatically affect the way individuals with autism engage with the visual world. Here we tested the dimension of space, but attention is known to operate over other complex features (e.g., color, shape), which may also have associated gradients that may be sharper in ASC (Kravitz and Behrmann, 2011). If the basic mechanisms of attention are atypical, these effects may extend to other sensory modalities as well.
A sharper gradient of attention in ASC may contribute to a robust finding in the literature: individuals with autism routinely display faster detection of targets in conjunctive visual search tasks (Plaisted et al., 1998; Joseph et al., 2009). Sharper spatial gradients around target locations would predict less interference from distractors and quicker response times in conjunctive visual search, and efficient perception of details in visual scenes.
Greater efficiency in detail perception has also been noted in other local perceptual tasks (Dakin and Frith, 2005). In particular, reduced effects of crowding (Baldassi et al., 2009; Kéïta et al., 2010) and enhanced facilitation from spatially proximal compared with more distant collinear Gabors (Kéïta et al., 2010) have been reported in ASC. A sharper gradient of spatial attention is consistent with both of these findings, as it might reflect a sharper decrease in lateral interactions with distance from the locus of attention. One might also expect a sharper gradient of attention to reduce the interference/enhancement from incongruent/congruent Eriksen flankers in ASC, but the literature is mixed, with some reports of equivalent interference in ASC and controls (Sanderson and Allen, 2012) and others reporting increased interference (Adams and Jarrold, 2012). The only study to manipulate target-flanker distance showed an increased interference effect across all distances in ASC (Adams and Jarrold, 2012) contrary to the intuitive prediction. Future research is needed to understand the relationship between this paradigm and our own, and the relative contributions of the facilitative/inhibitory interactions which might contribute to these effects and shape of the spatial gradient of attention (Müller et al., 2005).
Possible neural correlates of the increased sharpness of the spatial attentional gradient could be a narrower allocation of top-down signals to primary visual areas of the brain, altered dynamics within and among the early visual neurons that receive such top-down modulation, or a combination of both. For example, neurons in the banks of the monkey lateral intraparietal sulcus (LIP) have been implicated in the maintenance of a “priority map” of space, in which cells monitoring a cued area of the visual field show a sustained, heightened, retinotopically specific response in anticipation of a target (Gee et al., 2008). This signal is thought to facilitate detection and discrimination through top-down modulation of primary visual responses (Baluch and Itti, 2011). Differences in the distribution of such a parietal response could underlie the increased sharpness of the attentional gradient in ASC.
On the other hand, the receptive field sizes of early visual neurons are thought to spatially restrict the distribution of attentional modulation from extrastriate areas (Reynolds and Desimone, 1999), and could therefore account for this finding at a more basic level of visual processing. Such an account would lead to the prediction that smaller receptive field sizes may underlie the narrower attentional gradient we observe in ASC, a hypothesis which is arguably consistent with neurobiological findings of narrower minicolumns in the autistic brain (Casanova et al., 2002). Finally, our finding may result from an imbalance between the spatial extent of both low-level (receptive field) and higher-order (attentional signal) components of the autistic visual system (Reynolds and Heeger, 2009).
People with autism are often described as having tunnel vision, attracted to details of a visual scene while neglecting surrounding stimuli, as if attention were sharply pinpointed to the peaks of their visual world. The physiological roots of this behavior have never been known. We demonstrate a sharper gradient of spatial attention in the autistic brain, which may contribute to sharp perceptual enhancement of visual details. This finding not only provides insight into mechanisms underlying how individuals with ASC perceive the world, but also a starting point in a well characterized domain for modeling atypical neural circuitry in the autistic brain.
Footnotes
This work was supported by the Intramural Research Program of the National Institute of Mental Health, and was conducted in association with the NIHR CLAHRC for Cambridgeshire and Peterborough NHS Foundation Trust. We gratefully acknowledge the Gates-Cambridge Trust and the NIH-Cambridge Fellowship (to C.E.R.). S.B-C. was supported by the MRC and the Wellcome Trust during the period of this work. We thank Bonnie Auyeung, Marlene Behrmann, Alex Martin, John Mollon, Kate Plaisted-Grant, and Teresa Tavassoli for their comments, Carrie Allison for help with recruitment, and Steffen Lösch, Andra Adams, Richard Bethlehem, and Orian Welling for their help with stimulus design.
The authors declare no competing financial interests.
References
- Abrams J, Nizam A, Carrasco M. Isoeccentric locations are not equivalent: the extent of the vertical meridian asymmetry. Vision Res. 2012;52:70–78. doi: 10.1016/j.visres.2011.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adams NC, Jarrold C. Inhibition in autism: children with autism have difficulty inhibiting irrelevant distractors but not prepotent responses. J Autism Dev Disord. 2012;42:1052–1063. doi: 10.1007/s10803-011-1345-3. [DOI] [PubMed] [Google Scholar]
- Baldassi S, Pei F, Megna N, Recupero G, Viespoli M, Igliozzi R, Tancredi R, Muratori F, Cioni G. Search superiority in autism within, but not outside the crowding regime. Vision Res. 2009;49:2151–2156. doi: 10.1016/j.visres.2009.06.007. [DOI] [PubMed] [Google Scholar]
- Baluch F, Itti L. Mechanisms of top-down attention. Trends Neurosci. 2011;34:210–224. doi: 10.1016/j.tins.2011.02.003. [DOI] [PubMed] [Google Scholar]
- Baron-Cohen S, Wheelwright S, Skinner R, Martin J, Clubley E. The autism-spectrum quotient (AQ): evidence from Asperger syndrome/high-functioning autism, males and females, scientists and mathematicians. J Autism Dev Disord. 2001;31:5–17. doi: 10.1023/A:1005653411471. [DOI] [PubMed] [Google Scholar]
- Bertone A, Mottron L, Jelenic P, Faubert J. Enhanced and diminished visuo-spatial information processing in autism depends on stimulus complexity. Brain. 2005;128:2430–2441. doi: 10.1093/brain/awh561. [DOI] [PubMed] [Google Scholar]
- Brainard DH. The psychophysics toolbox. Spat Vis. 1997;10:433–436. doi: 10.1163/156856897X00357. [DOI] [PubMed] [Google Scholar]
- Brefczynski-Lewis JA, Datta R, Lewis JW, DeYoe EA. The topography of visuospatial attention as revealed by a novel visual field mapping technique. J Cogn Neurosci. 2009;21:1447–1460. doi: 10.1162/jocn.2009.21005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cameron EL, Tai JC, Carrasco M. Covert attention affects the psychometric function of contrast sensitivity. Vision Res. 2002;42:949–967. doi: 10.1016/S0042-6989(02)00039-1. [DOI] [PubMed] [Google Scholar]
- Casanova MF, Buxhoeveden DP, Switala AE, Roy E. Minicolumnar pathology in autism. Neurology. 2002;58:428–432. doi: 10.1212/WNL.58.3.428. [DOI] [PubMed] [Google Scholar]
- Chica AB, Bartolomeo P, Valero-Cabré A. Dorsal and ventral parietal contributions to spatial orienting in the human brain. J Neurosci. 2011;31:8143–8149. doi: 10.1523/JNEUROSCI.5463-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dakin S, Frith U. Vagaries of visual perception in autism. Neuron. 2005;48:497–507. doi: 10.1016/j.neuron.2005.10.018. [DOI] [PubMed] [Google Scholar]
- de Jonge MV, Kemner C, de Haan EH, Coppens JE, van den Berg TJ, van Engeland H. Visual information processing in high-functioning individuals with autism spectrum disorders and their parents. Neuropsychology. 2007;21:65–73. doi: 10.1037/0894-4105.21.1.65. [DOI] [PubMed] [Google Scholar]
- Gee AL, Ipata AE, Gottlieb J, Bisley JW, Goldberg ME. Neural enhancement and pre-emptive perception: the genesis of attention and the attentional maintenance of the cortical salience map. Perception. 2008;37:389–400. doi: 10.1068/p5874. [DOI] [PubMed] [Google Scholar]
- Golla H, Ignashchenkova A, Haarmeier T, Thier P. Improvement of visual acuity by spatial cueing: a comparative study in human and non-human primates. Vision Res. 2004;44:1589–1600. doi: 10.1016/j.visres.2004.01.009. [DOI] [PubMed] [Google Scholar]
- Joseph RM, Keehn B, Connolly C, Wolfe JM, Horowitz TS. Why is visual search superior in autism spectrum disorder? Dev Sci. 2009;12:1083–1096. doi: 10.1111/j.1467-7687.2009.00855.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kéïta L, Mottron L, Bertone A. Far visual acuity is unremarkable in autism: do we need to focus on crowding? Autism Res. 2010;3:333–341. doi: 10.1002/aur.164. [DOI] [PubMed] [Google Scholar]
- Koh HC, Milne E, Dobkins K. Spatial contrast sensitivity in adolescents with autism spectrum disorders. J Autism Dev Disord. 2010;40:978–987. doi: 10.1007/s10803-010-0953-7. [DOI] [PubMed] [Google Scholar]
- Kravitz DJ, Behrmann M. Space-, object-, and feature-based attention interact to organize visual scenes. Atten Percept Psychophys. 2011;73:2434–2447. doi: 10.3758/s13414-011-0201-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lord C, Risi S, Lambrecht L, Cook EH, Jr, Leventhal BL, DiLavore PC, Pickles A, Rutter M. The autism diagnostic observation schedule-generic: a standard measure of social and communication deficits associated with the spectrum of autism. J Autism Dev Disord. 2000;30:205–223. doi: 10.1023/A:1005592401947. [DOI] [PubMed] [Google Scholar]
- Mangun GR, Hillyard SA. Spatial gradients of visual attention: behavioral and electrophysiological evidence. Electroencephalogr Clin Neurophysiol. 1988;70:417–428. doi: 10.1016/0013-4694(88)90019-3. [DOI] [PubMed] [Google Scholar]
- Müller NG, Mollenhauer M, Rösler A, Kleinschmidt A. The attentional field has a Mexican hat distribution. Vision Res. 2005;45:1129–1137. doi: 10.1016/j.visres.2004.11.003. [DOI] [PubMed] [Google Scholar]
- Pellicano E, Gibson L, Maybery M, Durkin K, Badcock DR. Abnormal global processing along the dorsal visual pathway in autism: a possible mechanism for weak visuospatial coherence? Neuropsychologia. 2005;43:1044–1053. doi: 10.1016/j.neuropsychologia.2004.10.003. [DOI] [PubMed] [Google Scholar]
- Plaisted K, O'Riordan M, Baron-Cohen S. Enhanced visual search for a conjunctive target in autism: a research note. J Child Psychol Psychiatry. 1998;39:777–783. doi: 10.1111/1469-7610.00376. [DOI] [PubMed] [Google Scholar]
- Reynolds JH, Desimone R. The role of neural mechanisms of attention in solving the binding problem. Neuron. 1999;24:19–29. 111–125. doi: 10.1016/S0896-6273(00)80819-3. [DOI] [PubMed] [Google Scholar]
- Reynolds JH, Heeger DJ. The normalization model of attention. Neuron. 2009;61:168–185. doi: 10.1016/j.neuron.2009.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanderson C, Allen ML. The specificity of inhibitory impairments in autism and their relation to ADHD-type symptoms. J Autism Dev Disord. 2012 doi: 10.1007/s10803-012-1650-5. [DOI] [PubMed] [Google Scholar]
- Shah A, Frith U. An islet of ability in autistic children: a research note. J Child Psychol Psychiatry. 1983;24:613–620. doi: 10.1111/j.1469-7610.1983.tb00137.x. [DOI] [PubMed] [Google Scholar]
- Tavassoli T, Latham K, Bach M, Dakin SC, Baron-Cohen S. Psychophysical measures of visual acuity in autism spectrum conditions. Vision Res. 2011;51:1778–1780. doi: 10.1016/j.visres.2011.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Townsend JT, Ashby FG. Methods of modeling capacity in simple processing systems. In: Castellan NJ, Restle F, editors. Cognitive theory. Vol 3. Hillsdale, NJ: Erlbaum; 1978. pp. 199–239. [Google Scholar]
- Wechsler D. Wechsler abbreviated scale of intelligence (WASI) San Antonio, TX: The Psychological Corporation; 1999. [Google Scholar]