

Abstract
There is strong preclinical evidence that cancer, including breast cancer, undergoes immune surveillance. This continual monitoring, by both the innate and the adaptive immune systems, recognizes changes in protein expression, mutation, folding, glycosylation, and degradation. Local immune responses to tumor antigens are amplified in draining lymph nodes, and then enter the systemic circulation. The antibody response to tumor antigens, such as p53 protein, are robust, stable, and easily detected in serum, may exist in greater concentrations than their cognate antigens, and are potential highly specific biomarkers for cancer. However, antibodies have limited sensitivities as single analytes, and differences in protein purification and assay characteristics have limited their clinical application. For example, p53 autoantibodies in the sera are highly specific for cancer patients, but are only detected in the sera of 10-20% of patients with breast cancer. Detection of p53 autoantibodies is dependent on tumor burden, p53 mutation, rapidly decreases with effective therapy, but is relatively independent of breast cancer subtype. Although antibodies to hundreds of other tumor antigens have been identified in the sera of breast cancer patients, very little is known about the specificity and clinical impact of the antibody immune repertoire to breast cancer. Recent advances in proteomic technologies have the potential for rapid identification of immune response signatures for breast cancer diagnosis and monitoring. We have adapted programmable protein microarrays for the specific detection of autoantibodies in breast cancer. Here, we present the first demonstration of the application of programmable protein microarray ELISAs for the rapid identification of breast cancer autoantibodies.
Keywords: Breast Cancer, Autoantibodies, Tumor Antigen, Biomarker, Proteomics
Introduction
The early diagnosis of breast cancer is associated with a significant improvement in morbidity and overall survival1. While mammography is a highly successful screening modality, there remains a need for serologic biomarkers for the early diagnosis, monitoring, and classification of breast cancer. Despite this, very few serologic biomarkers have been developed for the diagnosis and management of breast cancer. CA27.29, CA15-3, and CEA are currently used in clinical practice, but by the 2007 biomarker guidelines for the American Society of Clinical Oncology, their use is limited to management of late stage disease2. Despite advances in genomics and proteomics, few, if any, biomarkers for early detection are in late stage clinical development for breast cancer. This may reflect the molecular heterogeneity of breast cancer3, a physical isolation of early breast cancer from the systemic circulation, or technologic limits of detection of early molecular changes.
The molecular changes in breast carcinogenesis are reflected in antigen upregulation, mutation, altered glycosylation, and altered degradation. These changes in protein structure can be specifically detected by the immune system, which carries altered antigens to draining lymph nodes for amplification of the immune response, to then enter the systemic circulation. This in vivo amplification system is highly specific and adaptive for changes in protein structure. Many tumor-specific autoantibodies have been identified in the sera of patients with breast cancer4, 5. Of these, several have been analyzed in detail with hundreds of serum samples, including p53 antigen 6, her2/neu 7, MUC18,c-myc 9, ECPKA 10, and NY-ESO-111, 12. While the specificity of these antigens for cancer is high (>90%), the sensitivities of individual antigens are low (10-20%). Multiplexed analysis using several antigens can increase sensitivity of detection up to 70% 5, 9. The majority of the published assays, while promising, depend on individual recombinant protein production and small, single-institution serum sets. The structure, quantity, and purity of recombinant protein used in ELISAs can dramatically affect the detection of autoantibodies, demonstrating that contaminating or misfolded proteins can lead to false-positive antibody detection13. Many of these assays must be cautiously interpreted, and they remain to be validated in standard biomarker multi-institutional phase 3 blinded retrsopective longitudinal repository studies or phase 4 prospective screening studies14.
Of the known tumor antigens that induce detectable antibodies in early cancer, mutated p53 protein was the first identified 15 and the most studied 6. The development of autoantibodies to p53 protein is thought to be due to accumulation of mutant p53 antigen in the cell nucleus. P53-specific autoantibodies correlate with cancer (p<10−4), and are associated with p53 point mutations, but epitope mapping using overlapping peptides has suggested that immunogenic regions are in the N- and C-termini of the molecule and not the mutation hotspots in the core region16-18. P53 autoantibodies have been detected in high-risk individuals months prior to the diagnosis of lung cancer and angiosarcoma19, and prior to relapse of early-stage breast cancer20.
A number of techniques have been developed to detect novel autoantibodies in patient sera. Traditionally, this has been done with serologic expression cloning (SEREX) of phage expression libraries derived from tumor cells. Over 2000 autoantigens recognized by patient sera have been identified with SEREX11, 21, 22, though relative few have gone on to validation studies. A modification of SEREX screening is combinatoryal phage display, which expresses tumor antigens as fusions with phage proteins. Phage display relies on biopanning of phage libraries using patient serum to identify candidate antigens, then spotting proteins on phage-protein microarrays. This approach has been used to identify tumor antigens in cancer, many of which are out-of-frame translation products or expressed from the 3′ UTR23-26. Autoantigens can also be detected by fractionation of tumor cell lysates and then immunoblotting with patient or control sera. This has been used to demonstrate antibodies in the sera of lung and prostate cancer27-30.
The advent of proteomic technologies, in particular protein microarrays, allows for rapid, flexible, high-throughput screening of thousands of tumor antigens for the detection of autoantibodies in the sera of cancer patients. This approach has been successfully used to screen autoimmune patient sera31 and to identify antigens expressed in ovarian cancer32. Because the identity of each feature on the array is known, this is a powerful technique for the detection of novel antibodies, but production of protein microarrays is challenging. Protein arrays produced by printing isolated proteins, which are now commercially available, require production and purification of recombinant antigen, raising issues of batch-to-batch variation, purity, and limited shelf-life. Glycan arrays are also being developed to detect alterations in glycosylation, which can also be highly immunogenic33.
While established and highly quantitative analytical methods are available for validation and large-scale serum screening, there is a clear need for discovery tools for the rapid detection of antibodies to thousands of tumor antigens in patient sera. Here, we present evidence of the adaptation of programmable protein microarrays34 for the detection of autoantibodies in breast cancer sera. Just-in-time protein expression avoids the potentially denaturing steps of protein isolation, storage, printing, and storage that occur with printed protein microarrays. To demonstrate that programmable protein microarrays can be used for serologic screening, we provide a direct comparison with a recombinat protein p53 autoantibody ELISA. We show that of 1,705 non redundant expressed antigens, dominant antibodies in melanoma and cancers of the breast and ovaries are detected. The application of these assays for biomarker detection will be discussed.
Materials and Methods
Patient Sera
Sera used in these analyses were obtained from the Lurie Breast Cancer Tissue and Blood Repository and the Specialized Research Program in Breast Cancer at the Dana-Farber Cancer Institute. Sera derived from breast cancer patients were obtained at the time of presentation with invasive breast cancer. Sera from patients with early stage ovarian cancer and matched healthy women were obtained from the Brigham and Women's Hospital. Control sera were from healthy women undergoing blood donation. Written consent was obtained from all subjects under institutional review board approval.
Generation of recombinant Survivin-GST and GST protein
The pGEX-4T-1 plasmid (Amersham, Piscataway, NJ) encoding full-length glutathione S-transferase (GST) protein and GST fused to the full-length survivin gene (Survivin-GST) were expressed in BL21 DE3 cells (Stratagene, La Jolla, CA) with 0.1M IPTG induction for 4 hours and purified according to manufacturer's recommendations. Bacterial cells were sonicated in cold PBS with protease inhibitors and lysozyme, then 1% Triton X-100 was added. Cleared supernatants were applied to glutathione CL-4B Sepharaose columns (Pharmacia, Piscataway, NJ) and bound protein was eluted in 50 mM Tris pH8.0 with 10 mM reduced glutathione. Protein purity was confirmed with SDS-PAGE and concentration determined at OD 280nM.Recombinant EBNA-1 protein was obtained by ABI (Columbia, MD)
P53 antibody ELISA
The p53 protein ELISA assay was performed per manufacturer's recommendations (p53 ELISAPLUS Autoantibody Kit, Calbiochem, Gibbstown, NJ). 100 μl each of calibrators, controls, and serum samples were added in duplicate. The calibrators were used neat, 1:1.5, 1:2, 1:3, 1:4, and 1:6 to obtain a calibration curve. Serum samples were diluted 1:100 and bound for 1 hour at room temperature (rt). The samples were removed and the wells washed 5 times with wash buffer. The wells were incubated with 100 μl of detector antibody conjugate for 1 hour at room temperature and washed 5 times with wash buffer. To develop the wells, 100 μl of substrate solution was added and incubated in the dark at room temperature. After 30 minutes, 50 μl of stop solution was added. The absorbance was determined at 450 nm using a Wallac plate reader. A calibration curve was created using the absorbances of the diluted calibrators. Dilutions of 1:1.5, 1:2, 1:3, 1:4, and 1:6 correspond to titers of 1, 0.67, 0.5, 0.33, 0.25, and 0.16 Units [U]. A p53 autoimmune index was calculated for each sample based on the formula (Absorbancesample-Absorbancecut-off)/(Absorbanceundiluted calibrator-Absorbancecut-off). The cut-off value is defined as 0.15 U. Any sample below this value is designated p53 negative. Samples within 20% of the cut-off were designated critical, and samples above 20% of the cut-off were designated p53 positive.
Survivin and EBNA-1 antibody ELISA
Recombinant GST, survivin-GST, or EBNA-1 protein was applied at 5 μg/ml to Nunc C96 Maxisorp plates (Fisher, Pittsburgh, PA) in carbonate buffer (pH 9.6) overnight at 4°C. Plates were washed in PBS-0.05% Tween (PBST) and blocked with PBS-Tween with 2% milk, overnight at 4°C. Serum was added in duplicate at 1:500 dilution in blocking buffer, overnight at 4°C. After washing, 1:1000 goat anti-human IgG-HRP secondary antibody was added (Zymax, Invitrogen, Carlsbad, CA) for one hour at rt. After washing, TMB-Plus (Dako, Carpinteria, CA) was added and the reaction stopped with 1N H2SO4. Absorbance was read at 450 nM, and GST signal was subtracted from Survivin-GST signal.
Generation of NAPPA protein microarrays
The arrays were prepared by printing purified plasmid DNA. Expression plasmid DNA in DH5alpha cells was grown overnight at 37°C in 1.5 mL liquid culture (Terrific Broth) and purified using Nucleobond anion exchange resin. The DNA (1 μg/μL) was printed along with the capture antibody (anti-GST antibody, 50 ng/μL, Amersham), protein crosslinker (BS3, 2mM, Pierce) and bovine serum albumin (3 mg/mL). Printing was done using an Affymetrix 427 or Genetix arrayer with 300um solid pins. The slides were expressed using previously published protocols34. For testing serum, the arrays were incubated with 300 uL of 1:300 diluted sera for 1hr at room temperature. Serum antibody binding was detected by probing the array with a HRP conjugated anti human IgG (1:500, Jackson lab) for 1hr at room temperature. The slides were developed for fluorescence detection by treating the slides with Tyramide signal amplification (Perkin Elmer) as per manufacturer's instructions. Data analysis was performed using Microvigene software (Vigenetech 2.9.9.2).
Statistical Analysis
Protein ELISA analyses were performed in duplicate or triplicate and positive autoantibodies were confirmed on repeat experiments. Individual antigens on microarrays were spotted as duplicates or quadruplicates. We defined an autoantibody signal as positive if it had at least a 3fold change compared to control sera (Figure 2), to the median of the sera tested (Figure 4), or to the median of all antigens on the array (Figure 5). To compare levels of antibody signal between two groups (as in Figures 1A and 1C) we used the Wilcoxon Rank Sum non-parametric test. To measure agreement in NAPPA and standard ELISA's detection of positive signals in sera, we used KAPPA analysis. When examining levels of p53-specific antibody in two patients who had positive signals prior to neoadjuvant chemotherapy (Figure 1B), we fitted a linear regression to log-transformed levels of antibody signal. To determine if the levels declined over time after chemotherapy was administered we tested if the regression slope was different from 0.
Figure 2.

Development of protein microarrays for detection of autoantibodies. A. Detection of p53 antigen with standard ELISA, using reference p53-antibody positive sera and negative sera. B and C. 4 antigens (S100A7, p21, ML-IAP, and p53) are expressed in microarray format, and p53 antigen is specifically detected with the reference sera. All four proteins are strongly expressed, as measured by detection of GST tag using anti-GST antibody. D. Sensitivity of detection. P53 positive serum was serially diluted and tested by standard ELISA and NAPPA. Standard deviation for ELISA was based on two samples and for NAPPA was based on four features on the array.
Figure 4.
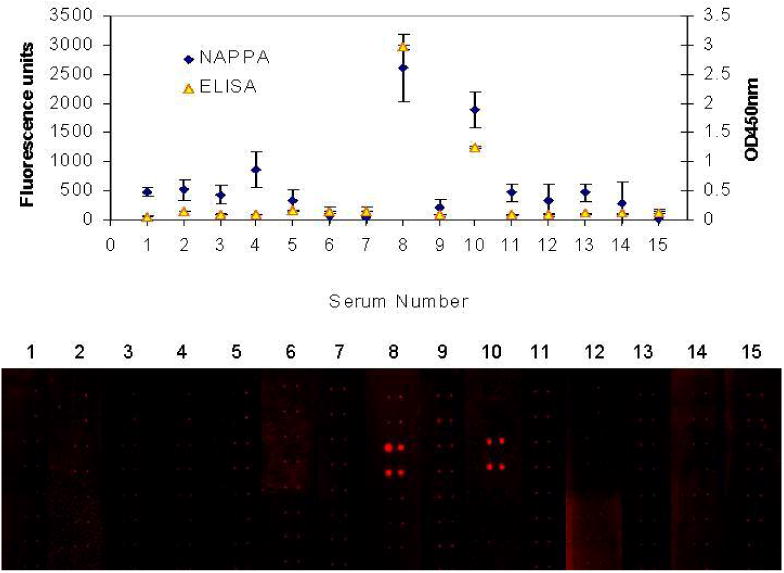
Comparable specificities of NAPPA microarray and protein ELISA for the detection of p53 antibodies in metastatic breast cancer patient sera. 15 sera from patients with metastatic breast cancer were diluted 1:100 and tested for p53-specific antibodies by protein ELISA and by NAPPA microarray. Two sera were positive by protein ELISA and were also positive by NAPPA microarray. The images of the quadruplicate-spotted antigens detected are shown in red on the bottom.
Figure 5.
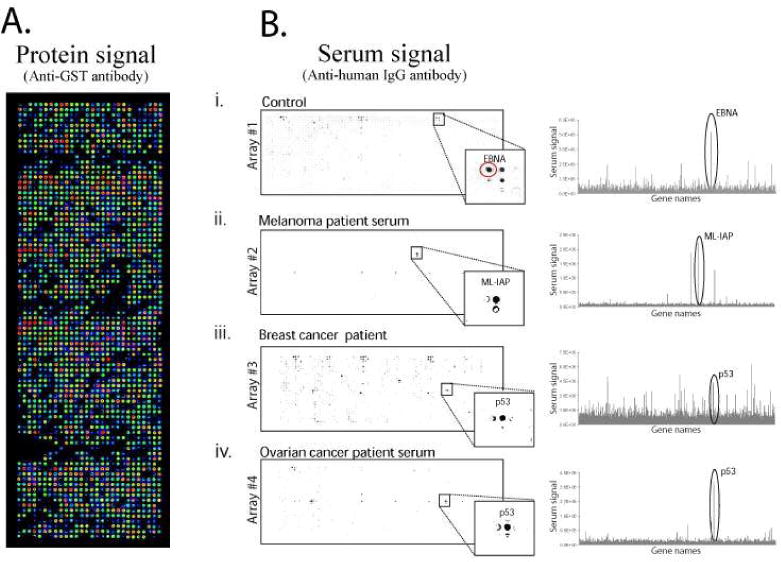
Development of custom tumor antigen high-density programmable protein microarrays for the detection of autoantibodies in patient sera. A) Over 1,700 candidate tumor antigens were expressed and captured in a microarray format, and protein expression was detected using anti-GST antibody. B. The arrays were probed with sera from a healthy individual, and patients with melanoma, breast and ovarian cancer. All arrays were incubated with 1:300 diluted serum and developed using a HRP conjugated anti-human IgG and TSA. (i) Shows EBNA response from healthy serum (red circle, CV=40% across 4 slides) (ii) Response to ML-IAP from melanoma serum (CV not tested) (iii) Response to p53 from breast cancer sera (CV=11% across 4 slides) (iv) Response to p53 from ovarian cancer sera (CV=49% across 2 slides).
Figure 1.
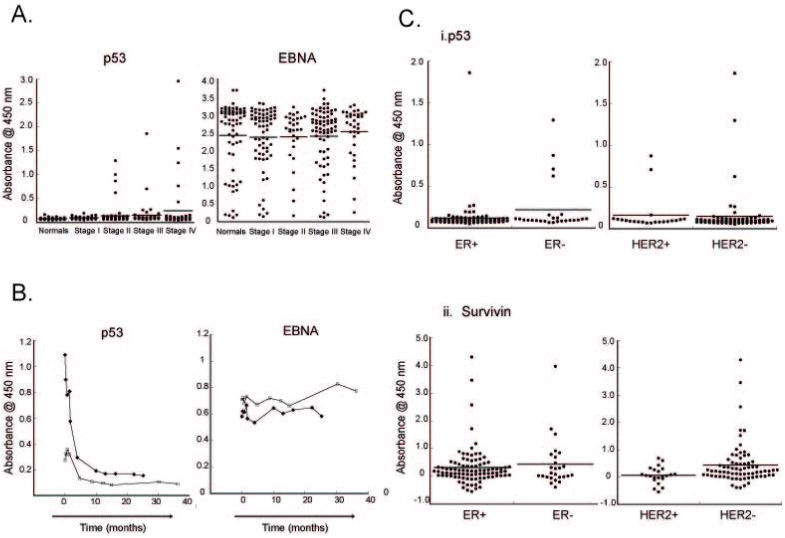
Detection of p53-specific antibodies in early-stage breast cancer. A. Sera derived from breast cancer patients and healthy female donors were tested for p53-specific antibodies (left) and EBNA-1 specific antibodies (right) by protein ELISA. The sera used consisted of normals (n=46); Stage I (n=29); Stage II (n=70); Stage III (n=47) and Stage IV breast cancer (n=40). The median level is shown with a bar. P53 antibody detection correlates with presence of breast cancer (p<0.0001). In contrast, EBNA-1 antibodies are detected at comparable intensity in normals (n=64) and patients (n=132), independent of stage. B. Rapid and durable loss of p53 antibodies with neoadjuvant chemotherapy. Two patients with early-stage Her2+ breast cancer who had detectable antibodies to p53, underwent neoadjuvant chemotherapy treatment with trastuzumab and chemotherapy over a 14-week period. P53 antibodies were detected in serial serum samples by ELISA, and show a rapid loss with the onset of treatment. C. Autoantibodies in breast cancer can be subtype-specific. i) Detection of p53-specific antibodies in breast cancer subtypes. Sera from Figure 1A were divided by tumor subtype. Subgroup analysis showed borderline significant preferential detection of anti-p53 antibodies in patients with ER- (n=26) compared with ER+(n=93) cancer, but no preference between HER2+ (n=22) vs. HER2- (n=73) breast cancer. ii) Selective detection of survivin-specific antibodies in HER2- breast cancer. Sera from the breast cancer patients in (A) were tested for anti-survivin specific antibodies using recombinant GST-tagged survivin. Subgroup analysis showed selective detection of anti-survivin antibodies in patients with HER2- breast cancer, independent of estrogen receptor (ER) status.
Results
Autoantibody Detection in Breast Cancer: Clinical Parameters
The clinical application of tumor antigen-specific antibodies as biomarkers require that they be specifically detected in cancer patients, be detectable in early stages, and/or be associated with tumor burden or disease outcome. The most well-studied autoantigen in breast cancer is p53. As with other p53-associated cancers, p53-specific antibodies are detected in the sera of 10-15% of breast cancer patients, and are strongly associated with missense mutations in the core domain of the molecule35-37. There have been multiple, but sometimes conflicting, studies correlating p53 antibodies to clinical stage, subtype, or outcome. These conflicting results may reflect underlying tumor heterogeneity, sample collection, or variation across different patient populations, but most likely is directly related to differences in antigen preparation and structure used in the assays38. For example, screening for antibodies with protein was more specific for cancer patients than screening with overlapping peptides 39. This is not surprising, since many B cell epitopes are conformational40, 41. P53 antibodies have been shown to correlate with poor prognosis in breast cancer 42, 43, but improved prognosis in ovarian cancer44, although the specificity of the p53 point mutations in the two cancers are quite similar.
The greatest potential impact of a biomarker in breast cancer would be to detect early-stage disease. Autoantibodies have been detected as early as DCIS45, but, again, the association of p53-specific antibodies and tumor stage has shown mixed results. A higher frequency of autoantibodies was observed in late stage breast cancer43, 46. However, studies in the U.K. and in Sweden found that p53 antibodies did not correlate with disease status47-49. In lung cancer, p53 autoantibodies decreased in most patients during therapy50, 51, arguing that the presence of these antibodies is associated with tumor burden, and they could be used as biomarkers to monitor response to therapy.
To demonstrate the detection of p53-specific antibodies in breast cancer, we used a set of well-annotated sera from 46 healthy female donors and 186 breast cancer patients, obtained at the time of diagnosis. The detection assay used was a commercially-available ELISA assay that uses recombinant, baculoviral-generated p53 antigen coated onto 96-well plates. The left panel of Figure 1 shows the levels of p53-specific antibodies in normal individuals and in cancer patients stratified by stage. p53-specific antibody levels were significantly lower in normal controls than in cancer patients using Wilcoxon Rank Sum test (p-value<0.0001). In comparison, levels of antibody to the EBV viral antigen EBNA-1 did not differ significantly between normal controls and cancer patients (p-value=0.1764, right panel of Figure 1A). Responses to p53 were detected as early as stage II.
To confirm that p53 antibodies are associated with tumor burden, figure 1B demonstrates p53 antibodies that were detected in two patients at time of diagnosis. Linear regression was fitted to log-transformed levels of p53-specific antibodies versus times. After neoadjuvant chemotherapy, the antibody concentration in the sera markedly declined over time (p-value=0.0002). The loss of these antibodies persisted long after chemotherapy completion. In contrast, we do not find any change in antibody concentration to the viral antigen, EBNA-1 (Figure 1B, right). This suggests that the rapid decline in anti-p53 antibody titer is antigen-specific, but we cannot rule out a more global effect of chemotherapy on antibody-specific immunity.
The three major subtypes of breast cancer, hormone receptor positive, her2/neu positive, and ‘triple-negative’ breast cancer, have markedly different RNA microarray expression profiles protein expression, and biologic behavior52. As shown in Figure 1C, the ER-negative breast cancer subtype had higher levels of p53-specific antibodies than the ER-positive subtype (p-value=0.0373). In comparison, there was no significant difference between the levels of p53-specific antibodies between the Her2-positive and Her2-negative subtypes of breast cancer (p-value=0.6698).
Antibodies other than p53 antigen have been detected in early-stage breast cancer patients, with varying frequencies. Survivin-specific antibodies have been detected in 11 of 46 patients breast cancer in Japan53 but only 5 of 64 patients in the U.S. 9. We generated recombinant survivin protein as a GST-fusion in bacteria. Using the same sera as in Figure 1C(i), we examined survivin-specific antibody levels (Figure 1C(ii)). We found no significant difference in survivin-specific antibody levels between ER-positive and ER-negative subtypes of breast cancer (p-value=0.9720). However, survivin-specific antibodies had higher levels of expression in the Her2-negative subgroup of breast cancer than in the Her2-positive subgroup (p-value=0.0076).
Development of NAPPA Tumor Antigen Protein Microarrays
Standard ELISAs offer a simple, rapid method to identify immune responses to a single antigen. However, this approach has low to moderate throughput and requires the expression and purification of every antigen to be tested. Therefore, ELISA has limited utility for proteome-scale applications, where the intent is to assess the global immune responses to thousands of proteins. High density antigen microarrays offer a significant advantage in throughput but can be difficult to produce given the expense and difficulty in producing large numbers of purified antigens.
We have developed programmable protein microarrays that comprise anchored cDNAs that are transcribed and translated in situ to produce corresponding epitope-tagged proteins34. Upon protein synthesis, the proteins are rapidly captured at the site of synthesis through a high affinity capture molecule, usually an antibody, that recognizes the epitope tag. This allows for a large number of antigens to be freshly synthesized and captured for profiling the immune response against entire proteomes.
Adaptation of NAPPA protein microarrays for Serologic Screening
To validate the assay, we obtained commercially available p53 antibody positive and negative sera (Calbiochem). The responses of the sera were confirmed by ELISA analysis using recombinant p53 antigen adhered to 96-well plates (Figure 2A). p53 autoantibodies were also detected by NAPPA protein microarray, and three control antigens (S100A7, p21, and ML-IAP) were not detected (Figure 2B,C). To confirm expression of all four antigens, the arrays were spotted in quadruplicate and probed with anti-GST antibody (Figure 2B, left), p53 negative sera (middle) and p53 positive sera (right). The p53 positive and negative sera were provided as internal standard with the commercially-available ELISA assay. Postive signal on NAPPA was defined as 10% above signal from non antigen containing negative control. The sensitivity of p53 detection by NAPPA and standard ELISA were tested by diluting the p53 positive sera, figure 2C. Comparable detection levels were observed with diluting the serum six fold.
Accessibility of p53 antigenic epitopes on NAPPA protein microarrays
Nearly all protein microarrays and many ELISAs are produced with proteins that bear epitope tags on either or both of their termini. In the context of detecting immune responses to these proteins, one potential concern is that these polypeptide tags might block access to antigenic epitopes via steric hindrance. As accessibility of antigenic epitopes to applied sera is essential for the detection of autoantibodies, we wished to confirm that multiple regions of known epitopes of the p53 autoantigen were accessible to antibodies directed at them. In figure 3, 7 different monoclonal antibodies directed at epitopes spread throughout the p53 protein (D01, D02, D07,D013,D014,240, and 243) were used to detect determinants on the p53 molecule expressed by NAPPA. The antigenic epitopes of these monoclonal antibodies have been well-characterized54 and are depicted in Figure 3. As shown with the anti-GST control which detects the c-terminal GST fusion protein, all p53 antigenic determinants are specifically detected by NAPPA fluorescence, 96-well ELISA, and by western blotting of expressed recombinant protein. The negative control (no antigen) and the control antigen p21-GST are not detected. This demonstrates that multiple regions of the molecule are accessible to antibody detection on the array. Moreover, the trends of detection are similar for these antibodies for both NAPPA and ELISA. Data on the quantitation of NAPPA and ELISA signals are provided in Supplemental Table 1, but the sensitivity of detection of epitopes must be interpreted with caution, as each monoclonal antibody has unique binding characteristics. Furthermore, one advantage of NAPPA is that, if needed, tumor antigens on the array can also be readily expressed with N-terminal tags or with non-GST tagged antigens to allow for flexibility of epitope expression (data not shown).
Figure 3.
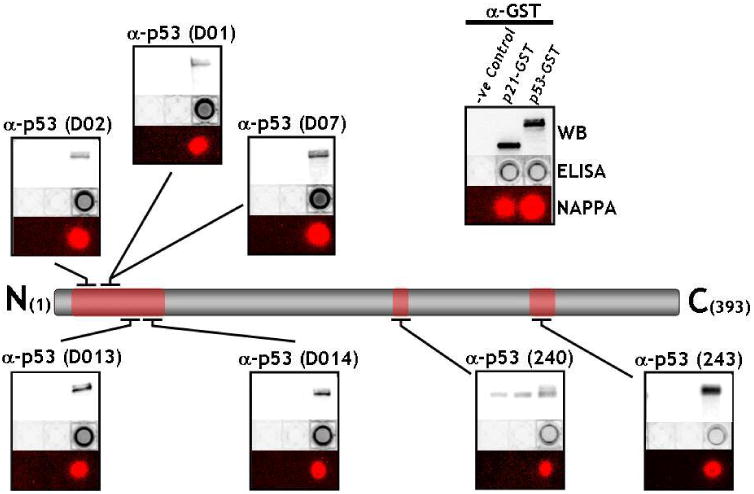
Detection of multiple distinct epitopes of p53 antigen expressed by NAPPA. 7 different monoclonal antibodies to p53 antigen (D01, D02, D07,D013,D014,240, and 243) were used to detect determinants on the p53 molecule expressed by NAPPA. As shown with the anti-GST control which detects the c-terminal GST fusion protein, all p53 antigenic determinants are specifically detected by NAPPA fluorescence, 96-well ELISA, and by western blotting of expressed recombinant protein. The negative control (no antigen) and the control antigen p21-GST are not detected.
NAPPA protein microarrays specifically detect p53 autoantibodies in breast cancer patient sera
To confirm the selectivity of autoantibody detection by tumor antigen expressed on NAPPA protein microarrays, sera from 15 patients with metastatic breast cancer were screened for p53 autoantibodies using a commercially-available protein ELISA kit (Calbiochem). Based on published reports and our own data (Figure 1A), we assumed that majority of sera did not have detectable antibodies to p53 antigen. We defined sera to have a detectable level of antibodies to p53 antigen on NAPPA and standard ELISA as those that have fold change of 3 or more when compared to the median. Fold changes were computed as ratios of the serum signal to the median signal of all 15 sera assayed. We found a significant concordance between NAPPA and standard ELISA (Kappa=1.0, p-value=0.0095) with the same 2 of the 15 patients having detectable autoantibodies to p53 antigen in both NAPPA and ELISA (Figure 4)
Application of Proteomic Technologies for Antibody Detection
Although specific for cancer patient sera, p53-specific autoantibodies occur with limited frequency in patients and alone have limited utility as cancer biomarkers. For new autoantigen discovery, we selected over 1,700 antigen clones from our PlasmID repository to build a custom high density antigen array, which was printed onto a derivatized glass surface 55. The gene collection represented 1,116 cancer related genes of which 539 were linked to breast cancer, as determined by the literature mining tool, MedGene 56. The protein types included signalling molecules like kinases (84), transcription factors (206), and transmembrane proteins (535). The success of protein expression and capture on the array was measured by probing the array with an anti GST antibody that recognizes a C terminal GST epitope on every antigen, figure 5A. We determined that 90% of the proteins had readily detected protein signal over background (10% above the average of non antigen containing spots).
To detect an immune response, we probed the high density cancer antigen arrays with serum from a healthy control and sera from patients with melanoma, ovarian and breast cancer. As a positive control, we chose to express the Epstein Barr nuclear antigen (EBNA), which derives from the highly prevalent Epstein Barr Virus (EBV). More than 90% of the population responds to this antigen, which acts as a good internal control indicating that the antigen detection assay is working)57.
In figure 5B, we detected responses to the EBNA antigen from the control, breast, and ovarian cancer sera. The serum from the melanoma patient did not have detectable antibodies to EBNA, although a strong response to a well-documented melanoma antigen called ML-IAP (Melanoma inhibitor of apoptosis protein) was detected, figure 5B (ii)58. For breast and ovarian sera, we detected responses to the tumour antigen p53 as well as other potential cancer antigens, figure 5B (iii,iv). Responses to p53 and ML-IAP were confirmed by standard ELISA (data not shown).
Discussion
Host factors that drive immune recognition of cancer
The evidence for active immunosurveillance of breast cancer is mounting. Polymorphisms in MHC molecules have long been associated with the development of clinical autoimmune disorders, and epidemiologic studies have shown mild assocation with protective HLA-DQ and HLA-DR class II alleles in breast cancer59. Lymphocytic infiltrates, as in ovarian cancer, colon cancer, and melanoma, have been shown to be associated with improved overall survival in breast cancer60, 61. However, less than half of patients with mutated p53 antigen develop a detectable antibody response. In longitudinal studies, patients who fail to make antibody responses at presentation, do not have detectable antibodies at disease relapse6, arguing that host immune memory and genetic variability drive the development of autoantibodies to tumor antigens.
Antibody binding to cognate antigen results in uptake of antigen/antibody complexes by Fc gamma receptors (FcGR) on antigen presenting cells. This enhances MHC class II antigen presentation and amplification of memory B cells bearing the specific immunoglobulin receptor. Polymorphisms in subsets of FcGR (IIa and IIIa) alter the affinity of FcGR binding to immunoglobulin and are associated with the development of clinical autoimmune disorders62. These polymorphisms are associated with altered response to antibody-mediated therapy in lymphoma63. In particular, we have found that structural polymorphisms in complement factor H are associated with clinical response to trastuzumab 64. Furthermore, dying and apoptotic tumor cells elicit danger signals via toll-like receptors (TLR) for dendritic cell activation and antigen presentation65. Not surprisingly, polymorphisms in TLR4, which impact innate immunity, affect response to chemotherapy in breast cancer66. Thus, host factors that impact antibody binding (FCG receptors), and activation of innate and adaptive immunity (complement and toll-like receptors) could profoundly influence the ability of patients to develop autoantibodies to tumor-derived antigens.
Protein structure, immunogenicity, and lessons from p53
The structural features that enhance immunogenicity of proteins are not well known, but charged surfaces, coiled-coil motifs, and nucleic acid binding are associated with autoantibody development in autoimmunity67. Alterations in post-translational modifications, such as phosphorylation and aberrant glycosylation, can also be strongly immunogenic68. Cancer-associated changes in apoptosis and cleavage by granzymes also render antigens more immunogenic69.
Given these factors, it is perhaps surprising that immunity to p53 antigen is detected across multiple cancer types and across multiple ethnic backgrounds in Europe, the U.S. and Asia. In our multiplexed protein microarray analysis of over 1,700 unique antigens, p53 is an immunodominant antigen which induces striking immunoreactivity compared with other tumor antigens. We have detected these antibodies in multiple stages and subtypes of breast and ovarian cancer. The structural cause of this immunodominance is not known, especially since epitope mapping studies have suggested that antibody binding to p53 is localized outside of the mutational hotspots. It has been hypothesized that one reason for p53 immunogenicity is due to protein binding to hsp70, causing immune activation70. Understanding the structural factors that drive antibody-mediated immunity will be critical for biomarker detection as well as immunotherapies.
Proteomic approaches to discovery of autoantibody signatures
The advent of proteomics allows for a global assessment of the breadth of the immune response to cancer. For biomarker development in particular, multiplexing autoantibody assays is critical to enhance sensitivities, while maintaining specificities5, 9, 23. Since the protein structure, content, and post-translational modifications are different for different proteomic techniques, it is not surprising that different antigens are identified. SEREX relies on phage expression and immunoblotting, and results in the detection of denatured epitopes that can be expressed in phage. Antibody detection by phage display has led to out-of-frame and 3′UTR epitopes, perhaps reflecting the cDNA libraries used, or defective ribosomal products present in cancer cells71. Fractionation of tumor cell lysates can detect cancer-specific post-translational modifications.
The programmable protein microarrays presented here may have distinct advantages for autoantibody detection in cancer. First, antigen selection is only limited by the availability of full-length sequence-verified open reading frames. Second, the cDNA's that are spotted on the arrays can be readily manipulated for epitope identification and mutational analysis. For serological assays, anitgens produced in situ by NAPPA behaved similarly to standard ELISA methods. Sensitivity of detection was comparable between NAPPA and commercial ELISA tests in titration experiments with the same antibodies. Accessbility to the epitopes of the antigen was unaffected by the differences in the methods of display. Moreover, good concordance of signal was observed between the two methods. The difference is that NAPPA can easily be multiplexed without sacrificing the performance features of a standard ELISA. Large number of antigens can be readily expressed with high levels of success for large scale studies.
The application of autoantibody biomarkers in breast cancer
There are a number of clinical settings where the development of biomarkers for breast cancer would have a significant clinical impact. Biomarkers for early detection of primary breast cancer would be particularly useful for patients at increased risk of cancer development or as a complement to mammography or magnetic resonance imaging (MRI) to identify occult cancers. Monitoring for early relapse will have a greater impact if therapies that target minimal residual disease are proven to be beneficial. The antibodies that are associated with improved outcome (tumor rejection) may also have therapeutic potential.
Supplementary Material
Acknowledgments
We thank Yanhui Hu, Andreas Rolf, Janice Williamson, and Mauricio Fernandez from the Harvard Institute of Proteomics, Harvard Medical School for their support. We thank Dr. David Lane for providing the p53 monoclonal antibodies. This study was supported by a research grant from the Early Detection Research Network 5U01CA117374-02 and the Friends for an Earlier Breast Cancer Test.
References
- 1.Effects of chemotherapy and hormonal therapy for early breast cancer on recurrence and 15-year survival: an overview of the randomised trials. Lancet. 2005;365(9472):1687–717. doi: 10.1016/S0140-6736(05)66544-0. [DOI] [PubMed] [Google Scholar]
- 2.Harris L, Fritsche H, Mennel R, Norton L, Ravdin P, Taube S, Somerfield MR, Hayes DF, Bast RC., Jr American Society of Clinical Oncology 2007 update of recommendations for the use of tumor markers in breast cancer. J Clin Oncol. 2007;25(33):5287–312. doi: 10.1200/JCO.2007.14.2364. [DOI] [PubMed] [Google Scholar]
- 3.Shipitsin M, Campbell LL, Argani P, Weremowicz S, Bloushtain-Qimron N, Yao J, Nikolskaya T, Serebryiskaya T, Beroukhim R, Hu M, Halushka MK, Sukumar S, Parker LM, Anderson KS, Harris LN, Garber JE, Richardson AL, Schnitt SJ, Nikolsky Y, Gelman RS, Polyak K. Molecular definition of breast tumor heterogeneity. Cancer Cell. 2007;11(3):259–73. doi: 10.1016/j.ccr.2007.01.013. [DOI] [PubMed] [Google Scholar]
- 4.Coronella-Wood JA, Hersh EM. Naturally occurring B-cell responses to breast cancer. Cancer Immunol Immunother. 2003;52(12):715–38. doi: 10.1007/s00262-003-0409-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chapman C, Murray A, Chakrabarti J, Thorpe A, Woolston C, Sahin U, Barnes A, Robertson J. Autoantibodies in breast cancer: their use as an aid to early diagnosis. Ann Oncol. 2007;18(5):868–73. doi: 10.1093/annonc/mdm007. [DOI] [PubMed] [Google Scholar]
- 6.Soussi T. p53 Antibodies in the sera of patients with various types of cancer: a review. Cancer Res. 2000;60(7):1777–88. [PubMed] [Google Scholar]
- 7.Disis ML, Pupa SM, Gralow JR, Dittadi R, Menard S, Cheever MA. High-titer HER-2/neu protein-specific antibody can be detected in patients with early-stage breast cancer. J Clin Oncol. 1997;15(11):3363–7. doi: 10.1200/JCO.1997.15.11.3363. [DOI] [PubMed] [Google Scholar]
- 8.von Mensdorff-Pouilly S, Petrakou E, Kenemans P, van Uffelen K, Verstraeten AA, Snijdewint FG, van Kamp GJ, Schol DJ, Reis CA, Price MR, Livingston PO, Hilgers J. Reactivity of natural and induced human antibodies to MUC1 mucin with MUC1 peptides and n-acetylgalactosamine (GalNAc) peptides. Int J Cancer. 2000;86(5):702–12. doi: 10.1002/(sici)1097-0215(20000601)86:5<702::aid-ijc16>3.0.co;2-1. [DOI] [PubMed] [Google Scholar]
- 9.Koziol JA, Zhang JY, Casiano CA, Peng XX, Shi FD, Feng AC, Chan EK, Tan EM. Recursive partitioning as an approach to selection of immune markers for tumor diagnosis. Clin Cancer Res. 2003;9(14):5120–6. [PubMed] [Google Scholar]
- 10.Nesterova MV, Johnson N, Cheadle C, Bates SE, Mani S, Stratakis CA, Khan IU, Gupta RK, Cho-Chung YS. Autoantibody cancer biomarker: extracellular protein kinase A. Cancer Res. 2006;66(18):8971–4. doi: 10.1158/0008-5472.CAN-06-1049. [DOI] [PubMed] [Google Scholar]
- 11.Scanlan MJ, Gout I, Gordon CM, Williamson B, Stockert E, Gure AO, Jager D, Chen YT, Mackay A, O'Hare MJ, Old LJ. Humoral immunity to human breast cancer: antigen definition and quantitative analysis of mRNA expression. Cancer Immun. 2001;1:4. [PubMed] [Google Scholar]
- 12.Chen YT, Scanlan MJ, Sahin U, Tureci O, Gure AO, Tsang S, Williamson B, Stockert E, Pfreundschuh M, Old LJ. A testicular antigen aberrantly expressed in human cancers detected by autologous antibody screening. Proc Natl Acad Sci U S A. 1997;94(5):1914–8. doi: 10.1073/pnas.94.5.1914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Schmetzer O, Moldenhauer G, Riesenberg R, Pires JR, Schlag P, Pezzutto A. Quality of recombinant protein determines the amount of autoreactivity detected against the tumor-associated epithelial cell adhesion molecule antigen: low frequency of antibodies against the natural protein. J Immunol. 2005;174(2):942–52. doi: 10.4049/jimmunol.174.2.942. [DOI] [PubMed] [Google Scholar]
- 14.Pepe MS, Etzioni R, Feng Z, Potter JD, Thompson ML, Thornquist M, Winget M, Yasui Y. Phases of biomarker development for early detection of cancer. J Natl Cancer Inst. 2001;93(14):1054–61. doi: 10.1093/jnci/93.14.1054. [DOI] [PubMed] [Google Scholar]
- 15.Crawford LV, Pim DC, Bulbrook RD. Detection of antibodies against the cellular protein p53 in sera from patients with breast cancer. Int J Cancer. 1982;30(4):403–8. doi: 10.1002/ijc.2910300404. [DOI] [PubMed] [Google Scholar]
- 16.Winter SF, Minna JD, Johnson BE, Takahashi T, Gazdar AF, Carbone DP. Development of antibodies against p53 in lung cancer patients appears to be dependent on the type of p53 mutation. Cancer Res. 1992;52(15):4168–74. [PubMed] [Google Scholar]
- 17.Dowell SP, Wilson PO, Derias NW, Lane DP, Hall PA. Clinical utility of the immunocytochemical detection of p53 protein in cytological specimens. Cancer Res. 1994;54(11):2914–8. [PubMed] [Google Scholar]
- 18.Casey G, Lopez ME, Ramos JC, Plummer SJ, Arboleda MJ, Shaughnessy M, Karlan B, Slamon DJ. DNA sequence analysis of exons 2 through 11 and immunohistochemical staining are required to detect all known p53 alterations in human malignancies. Oncogene. 1996;13(9):1971–81. [PubMed] [Google Scholar]
- 19.Trivers GE, De Benedetti VM, Cawley HL, Caron G, Harrington AM, Bennett WP, Jett JR, Colby TV, Tazelaar H, Pairolero P, Miller RD, Harris CC. Anti-p53 antibodies in sera from patients with chronic obstructive pulmonary disease can predate a diagnosis of cancer. Clin Cancer Res. 1996;2(10):1767–75. [PubMed] [Google Scholar]
- 20.Regele S, Vogl FD, Kohler T, Kreienberg R, Runnebaum IB. p53 autoantibodies can be indicative of the development of breast cancer relapse. Anticancer Res. 2003;23(1B):761–4. [PubMed] [Google Scholar]
- 21.Jager D, Unkelbach M, Frei C, Bert F, Scanlan MJ, Jager E, Old LJ, Chen YT, Knuth A. Identification of tumor-restricted antigens NY-BR-1, SCP-1, and a new cancer/testis-like antigen NW-BR-3 by serological screening of a testicular library with breast cancer serum. Cancer Immun. 2002;2:5. [PubMed] [Google Scholar]
- 22.Jager D, Taverna C, Zippelius A, Knuth A. Identification of tumor antigens as potential target antigens for immunotherapy by serological expression cloning. Cancer Immunol Immunother. 2004;53(3):144–7. doi: 10.1007/s00262-003-0470-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wang X, Yu J, Sreekumar A, Varambally S, Shen R, Giacherio D, Mehra R, Montie JE, Pienta KJ, Sanda MG, Kantoff PW, Rubin MA, Wei JT, Ghosh D, Chinnaiyan AM. Autoantibody signatures in prostate cancer. N Engl J Med. 2005;353(12):1224–35. doi: 10.1056/NEJMoa051931. [DOI] [PubMed] [Google Scholar]
- 24.Chatterjee M, Mohapatra S, Ionan A, Bawa G, Ali-Fehmi R, Wang X, Nowak J, Ye B, Nahhas FA, Lu K, Witkin SS, Fishman D, Munkarah A, Morris R, Levin NK, Shirley NN, Tromp G, Abrams J, Draghici S, Tainsky MA. Diagnostic markers of ovarian cancer by high-throughput antigen cloning and detection on arrays. Cancer Res. 2006;66(2):1181–90. doi: 10.1158/0008-5472.CAN-04-2962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sioud M, Hansen MH. Profiling the immune response in patients with breast cancer by phage-displayed cDNA libraries. Eur J Immunol. 2001;31(3):716–25. doi: 10.1002/1521-4141(200103)31:3<716::aid-immu716>3.0.co;2-9. [DOI] [PubMed] [Google Scholar]
- 26.Fernandez-Madrid F, Tang N, Alansari H, Granda JL, Tait L, Amirikia KC, Moroianu M, Wang X, Karvonen RL. Autoantibodies to Annexin XI-A and Other Autoantigens in the Diagnosis of Breast Cancer. Cancer Res. 2004;64(15):5089–96. doi: 10.1158/0008-5472.CAN-03-0932. [DOI] [PubMed] [Google Scholar]
- 27.Qiu J, Madoz-Gurpide J, Misek DE, Kuick R, Brenner DE, Michailidis G, Haab BB, Omenn GS, Hanash S. Development of natural protein microarrays for diagnosing cancer based on an antibody response to tumor antigens. J Proteome Res. 2004;3(2):261–7. doi: 10.1021/pr049971u. [DOI] [PubMed] [Google Scholar]
- 28.Zhang JY, Casiano CA, Peng XX, Koziol JA, Chan EK, Tan EM. Enhancement of antibody detection in cancer using panel of recombinant tumor-associated antigens. Cancer Epidemiol Biomarkers Prev. 2003;12(2):136–43. [PubMed] [Google Scholar]
- 29.Bouwman K, Qiu J, Zhou H, Schotanus M, Mangold LA, Vogt R, Erlandson E, Trenkle J, Partin AW, Misek D, Omenn GS, Haab BB, Hanash S. Microarrays of tumor cell derived proteins uncover a distinct pattern of prostate cancer serum immunoreactivity. Proteomics. 2003;3(11):2200–7. doi: 10.1002/pmic.200300611. [DOI] [PubMed] [Google Scholar]
- 30.Brichory FM, Misek DE, Yim AM, Krause MC, Giordano TJ, Beer DG, Hanash SM. An immune response manifested by the common occurrence of annexins I and II autoantibodies and high circulating levels of IL-6 in lung cancer. Proc Natl Acad Sci U S A. 2001;98(17):9824–9. doi: 10.1073/pnas.171320598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Robinson WH, Steinman L, Utz PJ. Protein arrays for autoantibody profiling and fine-specificity mapping. Proteomics. 2003;3(11):2077–84. doi: 10.1002/pmic.200300583. [DOI] [PubMed] [Google Scholar]
- 32.Hudson ME, Pozdnyakova I, Haines K, Mor G, Snyder M. Identification of differentially expressed proteins in ovarian cancer using high-density protein microarrays. Proc Natl Acad Sci U S A. 2007;104(44):17494–9. doi: 10.1073/pnas.0708572104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chen S, LaRoche T, Hamelinck D, Bergsma D, Brenner D, Simeone D, Brand RE, Haab BB. Multiplexed analysis of glycan variation on native proteins captured by antibody microarrays. Nat Methods. 2007;4(5):437–44. doi: 10.1038/nmeth1035. [DOI] [PubMed] [Google Scholar]
- 34.Ramachandran N, Hainsworth E, Bhullar B, Eisenstein S, Rosen B, Lau AY, Walter JC, LaBaer J. Self-assembling protein microarrays. Science. 2004;305(5680):86–90. doi: 10.1126/science.1097639. [DOI] [PubMed] [Google Scholar]
- 35.Hollstein M, Sidransky D, Vogelstein B, Harris CC. p53 mutations in human cancers. Science. 1991;253(5015):49–53. doi: 10.1126/science.1905840. [DOI] [PubMed] [Google Scholar]
- 36.Schlichtholz B, Legros Y, Gillet D, Gaillard C, Marty M, Lane D, Calvo F, Soussi T. The immune response to p53 in breast cancer patients is directed against immunodominant epitopes unrelated to the mutational hot spot. Cancer Res. 1992;52(22):6380–4. [PubMed] [Google Scholar]
- 37.Saleh J, Brunner C, Golzer R, Nastainczyk W, Montenarh M. p53 autoantibodies from patients with head and neck cancer recognise common epitopes on the polypeptide chain of p53. Cancer Lett. 2006;233(1):48–56. doi: 10.1016/j.canlet.2005.02.040. [DOI] [PubMed] [Google Scholar]
- 38.Volkmann M, Sinn HP, Gaugel D, Frey M, Hajjar Y, Ludwig J, Hansel S, Bastert G, Wallwiener D, Fiehn W, Zentgraf H, Huober J. Anti-p53 in breast cancer: concordance of different assay procedures and association with p53 antigen expression. Oncology. 2002;63(3):297–305. doi: 10.1159/000065472. [DOI] [PubMed] [Google Scholar]
- 39.Vennegoor CJ, Nijman HW, Drijfhout JW, Vernie L, Verstraeten RA, von Mensdorff-Pouilly S, Hilgers J, Verheijen RH, Kast WM, Melief CJ, Kenemans P. Autoantibodies to p53 in ovarian cancer patients and healthy women: a comparison between whole p53 protein and 18-mer peptides for screening purposes. Cancer Lett. 1997;116(1):93–101. doi: 10.1016/s0304-3835(97)00168-7. [DOI] [PubMed] [Google Scholar]
- 40.Laver WG, Air GM, Webster RG, Smith-Gill SJ. Epitopes on protein antigens: misconceptions and realities. Cell. 1990;61(4):553–6. doi: 10.1016/0092-8674(90)90464-p. [DOI] [PubMed] [Google Scholar]
- 41.Tan EM, Shi FD. Relative paradigms between autoantibodies in lupus and autoantibodies in cancer. Clin Exp Immunol. 2003;134(2):169–77. doi: 10.1046/j.1365-2249.2003.02259.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lubin R, Schlichtholz B, Bengoufa D, Zalcman G, Tredaniel J, Hirsch A, de Fromentel CC, Preudhomme C, Fenaux P, Fournier G, Mangin P, Laurent-Puig P, Pelletier G, Schlumberger M, Desgrandchamps F, Le Duc A, Peyrat JP, Janin N, Bressac B, Soussi T, et al. Analysis of p53 antibodies in patients with various cancers define B-cell epitopes of human p53: distribution on primary structure and exposure on protein surface. Cancer Res. 1993;53(24):5872–6. [PubMed] [Google Scholar]
- 43.Sangrajrang S, Arpornwirat W, Cheirsilpa A, Thisuphakorn P, Kalalak A, Sornprom A, Soussi T. Serum p53 antibodies in correlation to other biological parameters of breast cancer. Cancer Detect Prev. 2003;27(3):182–6. doi: 10.1016/s0361-090x(03)00066-7. [DOI] [PubMed] [Google Scholar]
- 44.Goodell V, Salazar LG, Urban N, Drescher CW, Gray H, Swensen RE, McIntosh MW, Disis ML. Antibody immunity to the p53 oncogenic protein is a prognostic indicator in ovarian cancer. J Clin Oncol. 2006;24(5):762–8. doi: 10.1200/JCO.2005.03.2813. [DOI] [PubMed] [Google Scholar]
- 45.Regele S, Kohlberger P, Vogl FD, Bohm W, Kreienberg R, Runnebaum IB. Serum p53 autoantibodies in patients with minimal lesions of ductal carcinoma in situ of the breast. Br J Cancer. 1999;81(4):702–4. doi: 10.1038/sj.bjc.6690751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gao RJ, Bao HZ, Yang Q, Cong Q, Song JN, Wang L. The presence of serum anti-p53 antibodies from patients with invasive ductal carcinoma of breast: correlation to other clinical and biological parameters. Breast Cancer Res Treat. 2005;93(2):111–5. doi: 10.1007/s10549-005-4321-9. [DOI] [PubMed] [Google Scholar]
- 47.Metcalfe S, Wheeler TK, Picken S, Negus S, Jo Milner A. P53 autoantibodies in 1006 patients followed up for breast cancer. Breast Cancer Res. 2000;2(6):438–43. doi: 10.1186/bcr91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lenner P, Wiklund F, Emdin SO, Arnerlov C, Eklund C, Hallmans G, Zentgraf H, Dillner J. Serum antibodies against p53 in relation to cancer risk and prognosis in breast cancer: a population-based epidemiological study. Br J Cancer. 1999;79(5-6):927–32. doi: 10.1038/sj.bjc.6690148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Willsher PC, Pinder SE, Robertson L, Nicholson RI, Ellis IO, Bell JA, Blamey RW, Green JA, Robertson JF. The significance of p53 autoantibodies in the serum of patients with breast cancer. Anticancer Res. 1996;16(2):927–30. [PubMed] [Google Scholar]
- 50.Zalcman G, Schlichtholz B, Tredaniel J, Urban T, Lubin R, Dubois I, Milleron B, Hirsch A, Soussi T. Monitoring of p53 autoantibodies in lung cancer during therapy: relationship to response to treatment. Clin Cancer Res. 1998;4(6):1359–66. [PubMed] [Google Scholar]
- 51.Mitsudomi T, Suzuki S, Yatabe Y, Nishio M, Kuwabara M, Gotoh K, Hatooka S, Shinoda M, Suyama M, Ogawa M, Takahashi T, Ariyoshi Y. Clinical implications of p53 autoantibodies in the sera of patients with non-small-cell lung cancer. J Natl Cancer Inst. 1998;90(20):1563–8. doi: 10.1093/jnci/90.20.1563. [DOI] [PubMed] [Google Scholar]
- 52.Perou CM, Sorlie T, Eisen MB, van de Rijn M, Jeffrey SS, Rees CA, Pollack JR, Ross DT, Johnsen H, Akslen LA, Fluge O, Pergamenschikov A, Williams C, Zhu SX, Lonning PE, Borresen-Dale AL, Brown PO, Botstein D. Molecular portraits of human breast tumours. Nature. 2000;406(6797):747–52. doi: 10.1038/35021093. [DOI] [PubMed] [Google Scholar]
- 53.Yagihashi A, Ohmura T, Asanuma K, Kobayashi D, Tsuji N, Torigoe T, Sato N, Hirata K, Watanabe N. Detection of autoantibodies to survivin and livin in sera from patients with breast cancer. Clin Chim Acta. 2005;362(1-2):125–30. doi: 10.1016/j.cccn.2005.06.009. [DOI] [PubMed] [Google Scholar]
- 54.Stephen CW, Helminen P, Lane DP. Characterisation of epitopes on human p53 using phage-displayed peptide libraries: insights into antibody-peptide interactions. J Mol Biol. 1995;248(1):58–78. doi: 10.1006/jmbi.1995.0202. [DOI] [PubMed] [Google Scholar]
- 55.Database P. http://plasmid.med.harvard.edu/PLASMID/
- 56.Hu Y, Hines LM, Weng H, Zuo D, Rivera M, Richardson A, LaBaer J. Analysis of genomic and proteomic data using advanced literature mining. J Proteome Res. 2003;2(4):405–12. doi: 10.1021/pr0340227. [DOI] [PubMed] [Google Scholar]
- 57.Ascherio A, Munger KL, Lennette ET, Spiegelman D, Hernan MA, Olek MJ, Hankinson SE, Hunter DJ. Epstein-Barr virus antibodies and risk of multiple sclerosis: a prospective study. Jama. 2001;286(24):3083–8. doi: 10.1001/jama.286.24.3083. [DOI] [PubMed] [Google Scholar]
- 58.Schmollinger JC, Vonderheide RH, Hoar KM, Maecker B, Schultze JL, Hodi FS, Soiffer RJ, Jung K, Kuroda MJ, Letvin NL, Greenfield EA, Mihm M, Kutok JL, Dranoff G. Melanoma inhibitor of apoptosis protein (ML-IAP) is a target for immune-mediated tumor destruction. Proc Natl Acad Sci U S A. 2003;100(6):3398–403. doi: 10.1073/pnas.0530311100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Chaudhuri S, Cariappa A, Tang M, Bell D, Haber DA, Isselbacher KJ, Finkelstein D, Forcione D, Pillai S. Genetic susceptibility to breast cancer: HLA DQB*03032 and HLA DRB1*11 may represent protective alleles. Proc Natl Acad Sci U S A. 2000;97(21):11451–4. doi: 10.1073/pnas.97.21.11451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Aaltomaa S, Lipponen P, Eskelinen M, Kosma VM, Marin S, Alhava E, Syrjanen K. Lymphocyte infiltrates as a prognostic variable in female breast cancer. Eur J Cancer. 1992;28A(4-5):859–64. doi: 10.1016/0959-8049(92)90134-n. [DOI] [PubMed] [Google Scholar]
- 61.Menard S, Tomasic G, Casalini P, Balsari A, Pilotti S, Cascinelli N, Salvadori B, Colnaghi MI, Rilke F. Lymphoid infiltration as a prognostic variable for early-onset breast carcinomas. Clin Cancer Res. 1997;3(5):817–9. [PubMed] [Google Scholar]
- 62.Koener HR, Kleije M, Algra J, Roos D, von dem Borne AE, de Haas M. Fc gammaRIIIa-158V/F polymorphism influences the binding of IgG by natural killer cell Fc gammaRIIIa, independently of the Fc gammaRIIIa-48L/R/H phenotype. Blood. 1997;90(3):1109–14. [PubMed] [Google Scholar]
- 63.Weng WK, Levy R. Two immunoglobulin G fragment C receptor polymorphisms independently predict response to rituximab in patients with follicular lymphoma. J Clin Oncol. 2003;21(21):3940–7. doi: 10.1200/JCO.2003.05.013. [DOI] [PubMed] [Google Scholar]
- 64.Anderson K, Aronzon D, Keshaviah A, Keung EZ, Kamma M, Winer EP, Burstein HJ, Harris LN. Role of host immune response genes in the clinical response to trastuzumab-based therapies. 2006;2006 [Google Scholar]
- 65.Matzinger P. The danger model: a renewed sense of self. Science. 2002;296(5566):301–5. doi: 10.1126/science.1071059. [DOI] [PubMed] [Google Scholar]
- 66.Apetoh L, Ghiringhelli F, Tesniere A, Obeid M, Ortiz C, Criollo A, Mignot G, Maiuri MC, Ullrich E, Saulnier P, Yang H, Amigorena S, Ryffel B, Barrat FJ, Saftig P, Levi F, Lidereau R, Nogues C, Mira JP, Chompret A, Joulin V, Clavel-Chapelon F, Bourhis J, Andre F, Delaloge S, Tursz T, Kroemer G, Zitvogel L. Toll-like receptor 4-dependent contribution of the immune system to anticancer chemotherapy and radiotherapy. Nat Med. 2007;13(9):1050–9. doi: 10.1038/nm1622. [DOI] [PubMed] [Google Scholar]
- 67.Plotz PH. The autoantibody repertoire: searching for order. Nat Rev Immunolv. 2003;3(1):73–8. doi: 10.1038/nri976. [DOI] [PubMed] [Google Scholar]
- 68.Tarp MA, Sorensen AL, Mandel U, Paulsen H, Burchell J, Taylor-Papadimitriou J, Clausen H. Identification of a novel cancer-specific immunodominant glycopeptide epitope in the MUC1 tandem repeat. Glycobiology. 2007;17(2):197–209. doi: 10.1093/glycob/cwl061. [DOI] [PubMed] [Google Scholar]
- 69.Casciola-Rosen L, Andrade F, Ulanet D, Wong WB, Rosen A. Cleavage by granzyme B is strongly predictive of autoantigen status: implications for initiation of autoimmunity. J Exp Med. 1999;190(6):815–26. doi: 10.1084/jem.190.6.815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kaur J, Srivastava A, Ralhan R. Serum p53 antibodies in patients with oral lesions: correlation with p53/HSP70 complexes. Int J Cancer. 1997;74(6):609–13. doi: 10.1002/(sici)1097-0215(19971219)74:6<609::aid-ijc9>3.0.co;2-y. [DOI] [PubMed] [Google Scholar]
- 71.Meister S, Schubert U, Neubert K, Herrmann K, Burger R, Gramatzki M, Hahn S, Schreiber S, Wilhelm S, Herrmann M, Jack HM, Voll RE. Extensive immunoglobulin production sensitizes myeloma cells for proteasome inhibition. Cancer Res. 2007;67(4):1783–92. doi: 10.1158/0008-5472.CAN-06-2258. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.