

Abstract
Extensive evidence indicates that heme oxygenase-1 (HO-1) exerts potent cytoprotective effects in response to stress. Previous studies have shown that gene therapy with HO-1 protects against myocardial ischemia/reperfusion injury for up to 8 weeks after gene transfer. However, the long-term effects of HO-1 gene therapy on myocardial ischemic injury and function are unknown. To address this issue, we created a recombinant adeno-associated viral vector carrying the HO-1 gene (rAAV/HO-1) that enables long-lasting transgene expression. Mice received injections in the anterior LV wall of rAAV/LacZ (LacZ group) or rAAV/HO-1 (HO-1 group); 1 year later, they were subjected to a 30-min coronary occlusion (O) and 4 h of reperfusion (R). Cardiac HO-1 gene expression was confirmed at 1 month and 1 year after gene transfer by immunoblotting and immunohistochemistry analyses. In the HO-1 group, infarct size (% of risk region) was dramatically reduced at 1 year after gene transfer (11.2 ± 2.1%, n = 12, vs. 44.7 ± 3.6%, n = 8, in the LacZ group; P < 0.05). The infarct-sparing effects of HO-1 gene therapy at 1 year were as powerful as those observed 24 h after ischemic PC (six 4-min O/4-min R cycles) (15.0 ± 1.7%, n = 10). There were no appreciable changes in LV fractional shortening, LV ejection fraction, or LV end-diastolic or end-systolic diameter at 1 year after HO-1 gene transfer as compared to the age-matched controls or with the LacZ group. Histology showed no inflammation in the myocardium 1 year after rAAV/HO-1-mediated gene transfer. These results demonstrate, for the first time, that rAAV-mediated HO-1 gene transfer confers long-term (1 year), possibly permanent, cardioprotection without adverse functional consequences, providing proof of principle for the concept of achieving prophylactic cardioprotection (i.e., “immunization against infarction”).
Keywords: Heme oxygenase-1, Gene therapy, Myocardial infarction, Cardiac function, Mouse
Introduction
Heme oxygenase-1 (HO-1) is induced in response to multiple cellular stresses [2, 13, 26]. HO-1 catalyzes the breakdown of potentially toxic heme to equimolar quantities of carbon monoxide, iron, and biliverdin, the latter being subsequently converted to bilirubin by biliverdin reductase [23]. These downstream products of heme degradation, which were once considered to be toxic metabolic waste molecules, have recently been shown to possess dose-dependent vasodilatory, antioxidant, antiapoptotic, and anti-inflammatory properties that are particularly desirable for tissue protection [2, 4, 8, 23].
Indeed, mounting experimental evidence suggests that induction of the HO-1 system is an important endogenous mechanism for cytoprotection and that the downstream products of heme degradation (carbon monoxide, iron, and biliverdin) may mediate these powerful beneficial effects. In fact, some investigators have reported that administration of exogenous carbon monoxide donors [13] or biliverdin [23] may offer a simple, inexpensive method to substitute for the cytoprotective effects of HO-1 in a variety of clinically applicable models [1, 3]. In addition, HO-1 is rapidly induced as a protective mechanism in response to various stimuli [3, 5, 26] among which is late preconditioning (PC) [32], an adaptation that enhances myocardial tolerance to ischemia 24–72 h after a sublethal ischemic stress [6]. Thus, HO-1 plays an important role in late PC [24, 32].
In view of these considerations, we have proposed that targeted induction of HO-1 may be an important therapeutic strategy for protection against ischemia/reperfusion injury [1]. Previous studies have shown that gene therapy with HO-1 reduces myocardial infarction 8 weeks after gene transfer [36]. However, to date, the long-term effects of HO-1 gene therapy on myocardial ischemic injury and function have not been explored. To address this issue, we have created a recombinant adeno-associated viral vector carrying the HO-1 gene (rAAV/HO-1). Because of chromosomal integration of the transgene and lack of inflammation, this vector enables long-lasting transgene expression (at least 1 year) in vivo [16, 19, 33].
The goal of this study was to use rAAV-mediated gene therapy to augment myocardial levels of HO-1 in mice and to determine the effects of this manipulation on myocardial infarct size and left ventricular (LV) function during a sustained period (1 year) of follow-up. A well-established murine model of myocardial infarction (MI) was used to compare the protection against myocardial ischemic injury afforded by rAAV/HO-1 gene therapy with that afforded by the late phase of ischemic PC. Our results indicate that the rAAV vector enables long-lasting HO-1 gene expression in the myocardium for up to 1 year and that this results in significant cardioprotection without adverse effects on cardiac function. The cardioprotection produced by rAAV-mediated HO-1 gene therapy was as powerful as that provided by late PC but dramatically longer (1 year vs. 3 days).
Methods
This study was performed in accordance with the Guide for the Care and Use of Laboratory Animals (Department of Health and Human Services, Publication No. [NIH] 86–23) and with the guidelines of the Animal Care and Use Committee of the University of Louisville, School of Medicine (Louisville, KY).
Construction of rAAV/HO-1 vector
The cDNA for the open reading frame sequence of mouse HO-1 [44] was cloned into the corresponding sites in the AAV2 shuttle plasmid. The recombinant AAV2 containing mouse HO-1 under the control of the cytomegalovirus promoter was generated by a triple-transfection technique using a helper virus-free system (Stratagene). rAAV/HO-1 was purified from 293 cell extracts by CsCl centrifugation and was routinely concentrated to approximately 2–4 × 1012 vector genomes (vg)/ml as determined by quantitative PCR [35, 43].
Gene transfer in vivo
Male ICR mice (8–12 weeks old; body weight 34.2 ± 2.4 g) were anesthetized with sodium pentobarbital (50 mg/kg i.p.). After opening the chest through a midline sternotomy, mice received two intramyocardial injections in the anterior left ventricular (LV) wall of rAAV/LacZ (1 × 1011 vg, LacZ group) or rAAV/HO-1 (1 × 1011 vg, HO-1 group). One year later, mice underwent cardiac tissue collection or the infarction protocol described below (Fig. 1). Each intramyocardial injection was 20 μl in volume and was performed with a 100-μl syringe using a 30-gauge needle; each mouse heart received two injections (totaling 40 μl in volume) in the soon-to-be-ischemic region of the LV [26, 28, 30].
Fig. 1.
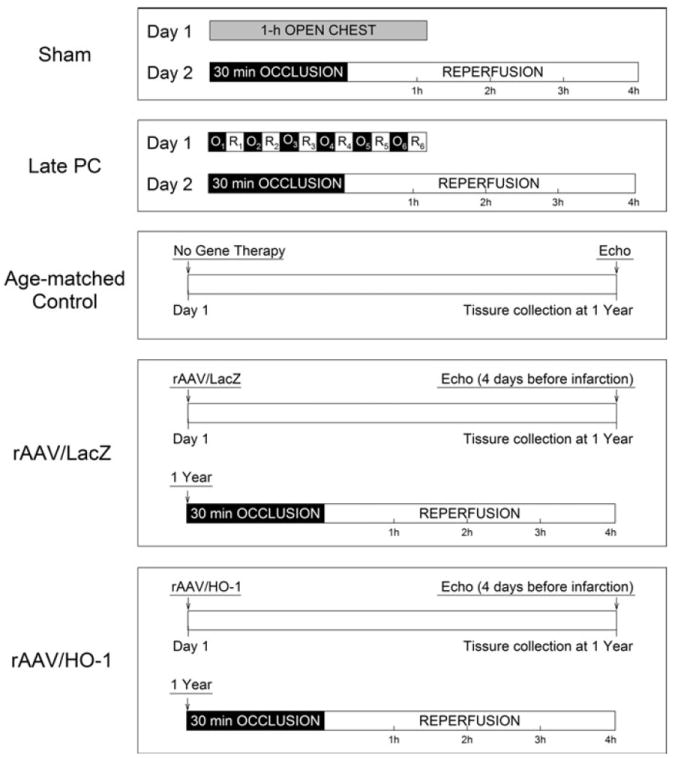
Experimental protocol. Five groups of mice were studied. The sham group underwent a thoracotomy and 1 h of open-chest state (without coronary occlusion) on day 1. The 1 h of open-chest state corresponded to the time interval necessary to perform 6 occlusion–reperfusion cycles in the late PC group; 24 h later (day 2), they underwent a 30-min coronary occlusion followed by 4 h of reperfusion. Mice in the late PC group were preconditioned with a sequence of 6 cycles of 4-min occlusion and 4-min reperfusion on day 1; on day 2, they were subjected to a 30-min coronary occlusion followed by 4 h of reperfusion. Mice in the age-matched control, LacZ, and HO-1 groups underwent the echocardiographic studies at 1 year (4 days prior to coronary occlusion) after gene transfer. The age-matched group did not receive any intervention during the period of the investigation. On day 1, mice in the LacZ and HO-1 groups were subjected to intramyocardial injections of rAAV/LacZ or rAAV/HO-1, respectively; at 1 year (4 days after echocardiographic measurements), both the LacZ and HO-1 groups underwent a 30-min coronary occlusion followed by 4 h of reperfusion for infarct size determination
Echocardiographic studies
Echocardiographic studies were performed using a HDI 5000 SonoCT ultrasound system (Philips Medical Systems, Bothell, WA) equipped with a 15- to 7-MHz linear transducer. Mice were anesthetized with isoflurane (3% induction and 1.5% maintenance). The chest was shaved, and mice were placed in a supine position. A rectal temperature probe was placed, and the body temperature was carefully maintained between 37.0 and 37.3°C with a heating pad throughout the study. The parasternal short-axis and modified parasternal long-axis views were used to obtain two-dimensional (2-D) and M-mode images. Digital images were analyzed off-line in a blinded fashion using ProSolv (version 2.5) image analysis software (Problem Solving Concepts, Indianapolis, IN), according to the American Society of Echocardiography standards [10, 25, 28].
Coronary occlusion/reperfusion protocol
The murine model of myocardial ischemia and reperfusion has been described in detail [12, 14]. Briefly, mice were anesthetized with sodium pentobarbital (50 mg/kg i.p.) and ventilated using carefully selected parameters. After administration of antibiotics, the chest was opened through a midline sternotomy, and a nontraumatic balloon occluder was implanted around the mid-left anterior descending coronary artery by using an 8-0 nylon suture. To prevent hypotension, blood from a donor mouse was given at serial times during surgery [14, 26]. Rectal temperature was carefully monitored and maintained between 36.7 and 37.3°C throughout the experiment. In all groups, MI was produced by a 30-min coronary occlusion followed by 4 h of reperfusion (Fig. 1). The LacZ and HO-1 groups received intramyocardial injections of rAAV/LacZ or rAAV/HO-1, respectively, as described above, 1 year before the 30-min occlusion. Successful performance of coronary occlusion and reperfusion was verified by visual inspection (i.e., by noting the development of a pale color in the distal myocardium after inflation of the balloon and the return of a bright red color due to hyperemia after deflation) and by observing S-T segment elevation and widening of the QRS on the ECG during ischemia and their resolution after reperfusion. After the coronary occlusion/reperfusion protocol was completed, the chest was closed in layers, and a small catheter was left in the thorax for 10–20 min to evacuate air and fluids. The mice were removed from the ventilator, kept warm with heat lamps, given fluids (1.0–1.5 ml of 5% dextrose in water i.p.), and allowed 100% oxygen via nasal cone [12, 14, 26, 28].
Postmortem tissue analysis
At the conclusion of the study, the heart was perfused with triphenyltetrazolium chloride (TTC) to delineate infarcted from viable myocardium [42]. The coronary artery was then tied at the site of the previous occlusion and the aortic root was perfused with phthalo blue dye to delineate the occluded–reperfused coronary vascular bed [12, 14]. The corresponding areas were measured by computerized video planimetry (Adobe Photoshop CS3) and from these measurements infarct size was calculated as a percentage of the region at risk [27, 28, 30].
Western immunoblotting analysis
Protein samples were isolated from heart tissues as previously described [26-28, 30]. The protein content in the membranous fractions was determined by the Bradford technique (Bio-Rad). The expression of HO-1 was assessed by standard SDS/PAGE Western immunoblotting techniques [26-28, 30]. Briefly, 80 μg of protein was separated on an SDS–polyacrylamide gel and transferred to a nitro-cellulose membrane. Gel transfer efficiency was recorded carefully by making photocopies of membranes dyed with reversible Ponceau staining; gel retention was determined by Coomassie blue staining [12, 26-28, 30]. Proteins were probed with the specific anti-HO-1 (Stressgen, San Diego, CA) and anti-GAPDH antibodies (Cell Signaling Technology, Inc. Danvers, MA). Immunoreactive bands were visualized with horseradish peroxidase–conjugated anti-rabbit IgG using an enhanced chemiluminescence detection kit (NEN), quantified by densitometry, and normalized to the Ponceau stain density. In all samples, the content of HO-1 protein was expressed as a percentage of the corresponding protein in the rAAV/LacZ group (control group) [12, 25, 26].
Bilirubin assay
Total bilirubin levels in myocardium were determined by using a bilirubin assay kit (Wako Chemicals USA, Inc., Richmond, VA) according to the manufacturer’s instruction [31]. The sample absorbance at 450 nm was normalized to a concomitantly performed standard curve constructed from commercially available bilirubin standard solution (Wako Chemicals USA, Inc., Richmond, VA) to determine the total bilirubin levels [26].
Immunohistochemistry analysis
Immunohistochemical analysis for HO-1 protein expression 1 year after gene transfer was performed by using the ABC kit (Vector Laboratories) according to the manufacturer’s instruction [26]. Frozen 10-μm sections were fixed in 10% formalin for 2 min at room temperature, washed in PBS, and blocked with Mouse-On-Mouse (MOM) IgG blocking reagent. Antigenic epitopes were equilibrated in MOM diluent and then incubated at 37°C with a specific anti-HO-1 antibody (Stressgen, San Diego, CA). Alternate sections were incubated in the absence of the primary antibody (negative control) as previously reported [26, 28].
Statistical analysis
Data are presented as means ± SEM. All data were analyzed with one-way ANOVA (ANOVA for normally distributed data, or Kruskal–Wallis one-way ANOVA on ranks for data that are not normally distributed), as appropriate, followed by unpaired Student’s t tests with the Bonferroni correction. The relationship between infarct size and risk region size was compared among groups with ANCOVA (SAS software system), with the size of the risk region as the covariate [11, 14]. A P value <0.05 was considered statistically significant. All statistical analyses were performed using the SigmaStat software system (3.5 V) [25, 26].
Results
Exclusions
A total of 97 mice were used for this study (72 for the infarct size and functional studies and 25 for biochemical analyses). Twelve mice died during or shortly after the surgical procedure: five after the first surgery (gene transfer) and seven after the second surgery (coronary occlusion). Five mice (5%) were excluded because of technical problems, including body temperature out of normal range (n = 1), balloon occluder malfunction (n = 1), and bleeding during the surgery (n = 3). Three mice (3%) died during the 1-year follow-up. Thus, a total of 77 mice were included in the final analyses. Nineteen mice were used for both this study and the study of rAAV/iNOS gene therapy reported in the companion manuscript [29], specifically, six mice in the sham group, six mice in the late PC group, and seven mice in the rAAV/LacZ group.
Fundamental physiological parameters
Heart rate and rectal temperature, fundamental physiological parameters that may impact infarct size [14, 26], were similar in the sham and late PC groups and in the LacZ and HO-1 groups (Table 1). Heart rate in the sham and late PC groups was higher than that in the gene therapy groups due to the effects of surgical trauma 24 h earlier [14]. However, heart rate was not statistically different between the sham and late PC groups or between the lacZ and HO-1 groups. Within the same group, heart rate did not differ significantly at any time-point before and during the 30-min occlusion or the ensuing reperfusion (Table 1). As shown in Table 1, rectal temperature remained within a narrow, physiologic range (36.8–37.2°C) in all groups. During the echocardiographic studies, heart rate (a parameter that may impact myocardial function) was similar in the age-matched control, LacZ, and HO-1 groups. The average heart rate in the three groups at 1 year after gene therapy ranged narrowly from 484 ± 17 to 461 ± 19 bpm (P > 0.05).
Table 1.
Heart rate and rectal temperature on the day of the 30-min coronary occlusion
| Groups | Preocclusion | Occlusion
|
Reperfusion
|
||
|---|---|---|---|---|---|
| 5 min | 30 min | 5 min | 15 min | ||
| Heart rate (beats/min) | |||||
| Sham (n = 12) | 609 ± 19 | 612 ± 17 | 620 ± 19 | 637 ± 27 | 635 ± 21 |
| Late PC (n = 10) | 621 ± 21 | 620 ± 27 | 615 ± 36 | 614 ± 34 | 639 ± 28 |
| rAAV/LacZ (n = 8) | 527 ± 14b | 538 ± 10 | 541 ± 12 | 542 ± 15 | 547 ± 19 |
| rAAV/HO-1 (n = 12) | 551 ± 17 | 576 ± 21 | 544 ± 25 | 547 ± 28 | 543 ± 25a,b |
| Body temperature (°C) | |||||
| Sham (n = 12) | 36.8 ± 0.1 | 37.0 ± 0.1 | 36.9 ± 0.0 | 36.9 ± 0.0 | 36.9 ± 0.1 |
| Late PC (n = 10) | 36.9 ± 0.1 | 37.0 ± 0.1 | 36.9 ± 0.1 | 36.9 ± 0.0 | 36.9 ± 0.1 |
| rAAV/LacZ (n = 8) | 37.1 ± 0.0 | 37.0 ± 0.1 | 37.1 ± 0.0 | 37.2 ± 0.0 | 37.2 ± 0.1 |
| rAAV/HO-1 (n = 12) | 37.1 ± 0.1 | 37.2 ± 0.0 | 37.0 ± 0.1 | 36.9 ± 0.1 | 36.9 ± 0.1 |
Data are means ± SEM. The heart rate and rectal temperature were recorded before the 30-min coronary occlusion (preocclusion), at 5 min and 30 min during the 30-min occlusion, and at 5 min and 15 min after reperfusion. Rectal temperature was continuously monitored and carefully controlled throughout the experiment, as detailed in the text
P < 0.05 versus sham group
P < 0.05 versus late PC group
HO-1 gene expression in vivo
The function of the rAAV/HO-1 vector was first assessed in the transduced myocardium 1 month after gene transfer, before the initiative of long-term (1 year) gene transfer experiments. One month or 1 year after intramyocardial injections of rAAV/LacZ or rAAV/HO-1, the myocardium surrounding the sites of injections (~40 mg) was harvested for measurement of HO-1 protein expression (each heart received two injections with a total volume of 40 μl). Western immunoblotting analyses demonstrated that cardiac HO-1 gene transfer mediated by the rAAV vector resulted in a pronounced increase in HO-1 protein expression at both 1 month (+5.9-fold) and 1 year (+7.7-fold) compared to mice transfected with the reporter gene LacZ, respectively (n = 4/group, P < 0.05; Fig. 2a). Consistent with the immunoblotting data, immunohistochemical analysis showed elevated expression of HO-1 in cardiac myocytes 1 year after transfection with rAAV/HO-1 (n = 3; Fig. 3). No HO-1 immunoreactivity was noted in the rAAV/LacZ group (n = 3) and in LV sections incubated with nonimmune serum (not shown). There was no evidence of inflammation in either the rAAV/LacZ or the rAAV/HO-1 group (Fig. 3), indicating that neither the administration of an irrelevant viral vector nor the intramyocardial injections are associated with the upregulation of HO-1 observed in the rAAV/HO-1 group at 1 year after gene transfer. Additionally, the content of bilirubin, a byproduct of HO-1, was significantly increased in the transduced myocardium 1 year after HO-1 gene transfer (1.79 ± 0.14 ng/μg protein [n = 4] vs. 0.22 ± 0.05 ng/μg protein [n = 4] in the LacZ group; P < 0.05; Fig. 2b), demonstrating that the transgenic HO-1 protein was functionally competent 1 year after rAAV/HO-1-mediated gene transfer, and that this rAAV vector enables long-term expression of the HO-1 transgene in myocardium in vivo.
Fig. 2.
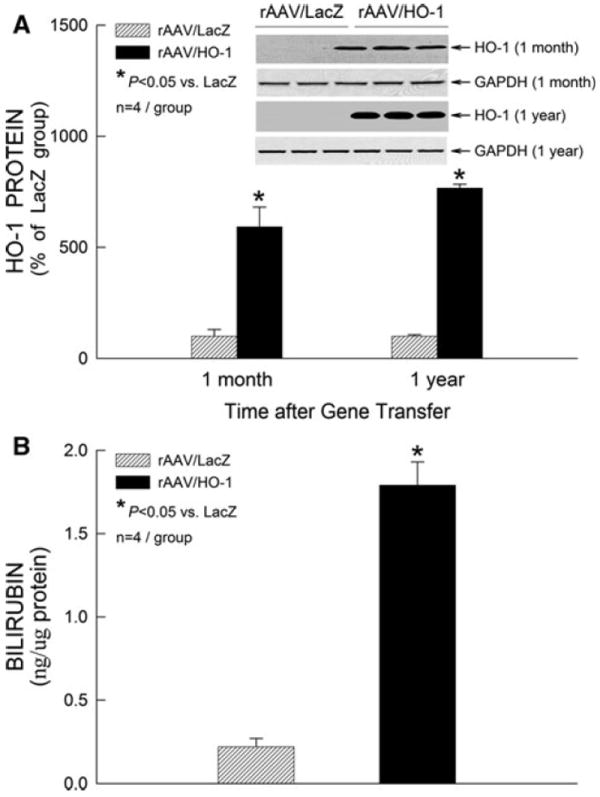
HO-1 gene expression in myocardium 1 month and 1 year after rAAV-mediated gene transfer. a Western immunoblots and densitometric analyses of HO-1 protein signals in membranous fractions of the transduced myocardium. b bilirubin content in cytosolic fractions of the transduced myocardium. Data are mean ± SEM
Fig. 3.
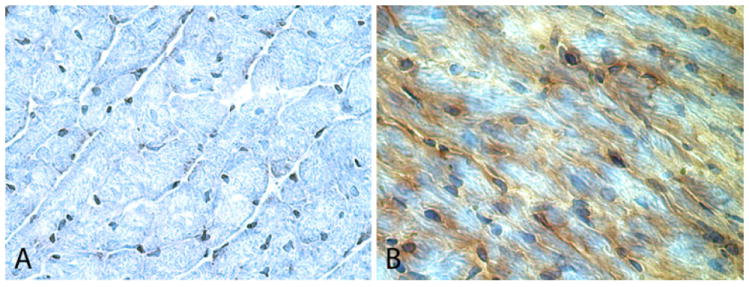
Expression of the HO-1 gene in myocardium 1 year after rAAV-mediated gene transfer. Hearts transfected with rAAV/LacZ (a ×400) or rAAV/HO-1 (b ×400), respectively, are shown. Robust expression of HO-1 (b brown stain) was observed in the transfected region of myocardium 1 year after gene transfer (n = 3)
Long-term effect of HO-1 gene transfer on infarct size
On the day of the 30-min occlusion, there were no appreciable differences among the four groups with respect to the region at risk (sham group, 39.7 ± 2.2% of LV; late PC group, 36.4 ± 3.2%; LacZ group, 40.4 ± 4.2%; and HO-1 group, 36.3 ± 2.1%; P > 0.05). When mice were preconditioned with six cycles of 4-min coronary occlusion/4-min reperfusion 24 h before the 30-min coronary occlusion, the mean infarct size was 69% smaller in the late PC group than in the sham group (15.0 ± 1.7% of the risk region, n = 10, vs. 49.6 ± 3.1%, n = 12, respectively; P < 0.05; Fig. 4), indicating a late PC effect against infarction. In the HO-1 gene therapy group, the average infarct size was 75% smaller than in the LacZ control group (11.2 ± 2.1% of the risk region, n = 12, vs. 44.7 ± 3.6%, n = 8, respectively; P < 0.05; Fig. 4), demonstrating that the expression of the HO-1 gene resulted in a marked cardioprotective effect 1 year after gene transfer.
Fig. 4.
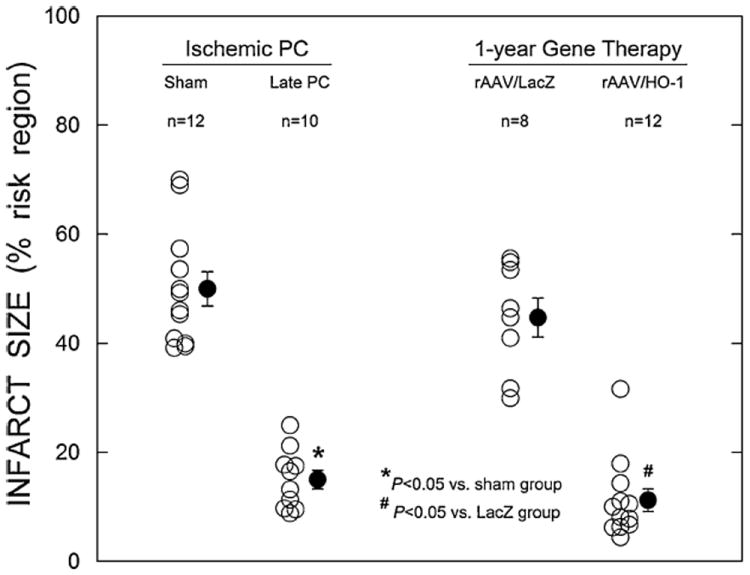
Myocardial infarct size expressed as a percentage of the region at risk. Open circles individual mice; solid circles means ± SE
Two representative examples of the infarcts observed in the LacZ (top) and HO-1 (bottom) groups are shown in Fig. 5. In the HO-1-treated mouse heart, patchy areas of infarction were noted instead of the confluent infarcts seen in the LacZ-treated mouse heart. This patchy pattern of cell death was characteristic of both the HO-1 group (Fig. 5) and the late PC group [14]. The similarity in infarct size between the late PC group (15.0 ± 1.7% of the risk region) and the HO-1 gene therapy group (11.2 ± 2.1% of the risk region) indicates that the protective effect of HO-1 gene therapy was comparable to that of late PC.
Fig. 5.
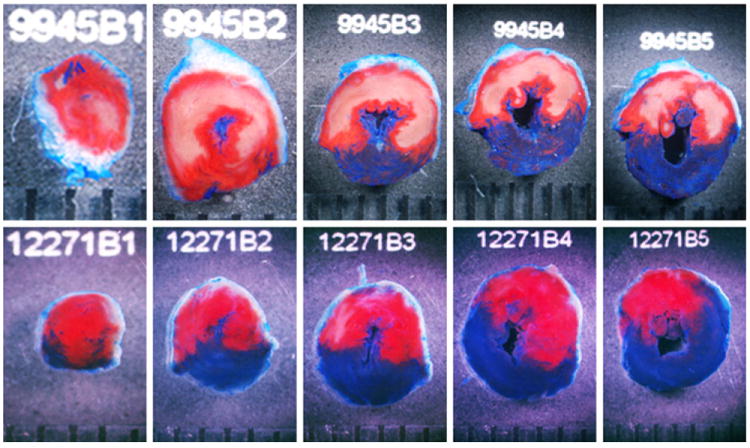
Representative LV slices from rAAV/LacZ and rAAV/HO-1 groups 1 year after gene therapy. The slices shown here were obtained from the apex to the base (B1–B5). Top panel rAAV/LacZ. Bottom panel rAAV/HO-1. The region at risk and the infarct were identified by postmortem perfusion with TTC and phthalo blue dye, as described in “Methods”. As a result of this procedure, the nonischemic portion of the LV was stained dark blue, and viable tissue within the region at risk was stained bright red, whereas infarcted tissue was light yellow/white. The LV endocardial surface was stained dark blue with phthalo blue to facilitate identification of the endocardial border of the slice. rAAV/LacZ-treated heart exhibited large, confluent areas of infarction. In contrast, the heart pretreated with rAAV/HO-1 exhibited small patchy areas of infarction, indicating a profound and long-term (1 year) cardioprotective effect of HO-1 gene therapy. The scale at bottom is in millimeters
In all four groups, the size of the infarct was positively and linearly related to the size of the region at risk (sham group, r = 0.85; late PC group, r = 0.61; LacZ group, r = 0.90; and HO-1 group, r = 0.59; Fig. 6). As expected [14], the regression line was shifted to the right in the late PC group compared with the control sham group (P < 0.05 by ANCOVA; Fig. 6, left). In the group pretreated with HO-1 gene therapy, the regression line was again significantly shifted to the right compared with the control LacZ-treated group (P < 0.05 by ANCOVA; Fig. 6, right) and was virtually indistinguishable from that of the late PC group, indicating both that, for any given size of the region at risk, infarct size was reduced by HO-1 gene therapy and that the magnitude of this effect was similar to that of the effect of ischemic PC.
Fig. 6.
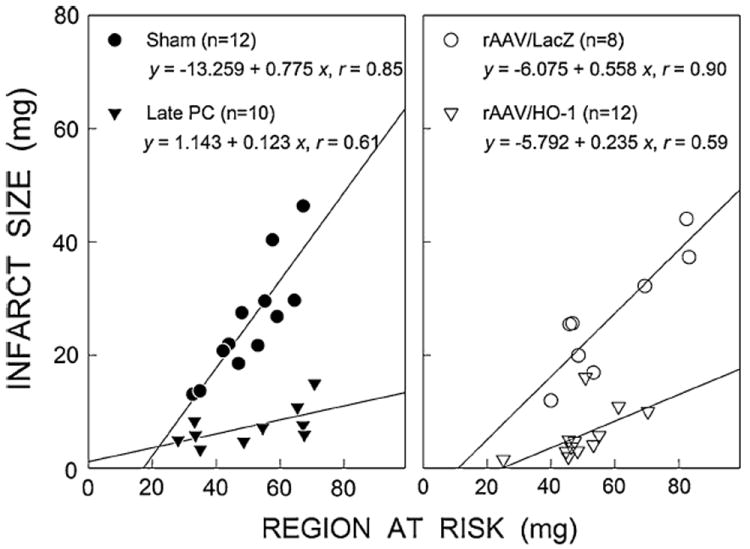
Relationship between size of region at risk and size of MI 1 year after rAAV/HO-1 gene therapy. Graphs show individual values and regression lines obtained by linear regression analysis for the various groups. Left sham and late PC groups. Right rAAV/LacZ and rAAV/HO-1 groups. In all groups, infarct size was positively and linearly related to risk region size. However, the slopes of the regression lines were smaller in the late PC and rAAV/HO-1 groups as compared to the sham and rAAV/LacZ control groups, respectively, demonstrating an attenuated infarct size response to myocardial injury in both of the late PC (24 h) and rAAV/HO-1 gene therapy (1 year)
Long-term effect of HO-1 gene transfer on cardiac function
Cardiac function was assessed by 2-D echocardiography at 1 year after gene transfer. To prevent any after-effects of the anesthesia used during the echocardiographic assessments, a 4-day interval was allowed between the echo-cardiographic assessment and the 30-min coronary occlusion followed by 4 h of reperfusion (Fig. 1). As shown in Fig. 7, there were no appreciable differences in LV fractional shortening, LV ejection fraction, or LV end-diastolic or end-systolic diameter at 1 year after HO-1 gene transfer as compared to age-matched controls or with the LacZ group. HO-1 gene transfer (n = 14) had no effect on LV dimensions or function up to 1 year as compared to the LacZ group (n = 11) (at 1 year: FS, 34 ± 1.5 vs. 32 ± 2.6%; EF, 63 ± 1.7 vs. 60 ± 2.9%; LVEDD, 4.5 ± 0.1 vs. 4.2 ± 0.2 mm; LVESD, 3.0 ± 0.1 vs. 2.9 ± 0.2 mm), demonstrating that rAAV-mediated HO-1 gene transfer in the heart does not induce adverse cardiac functional consequences for up to 1 year.
Fig. 7.
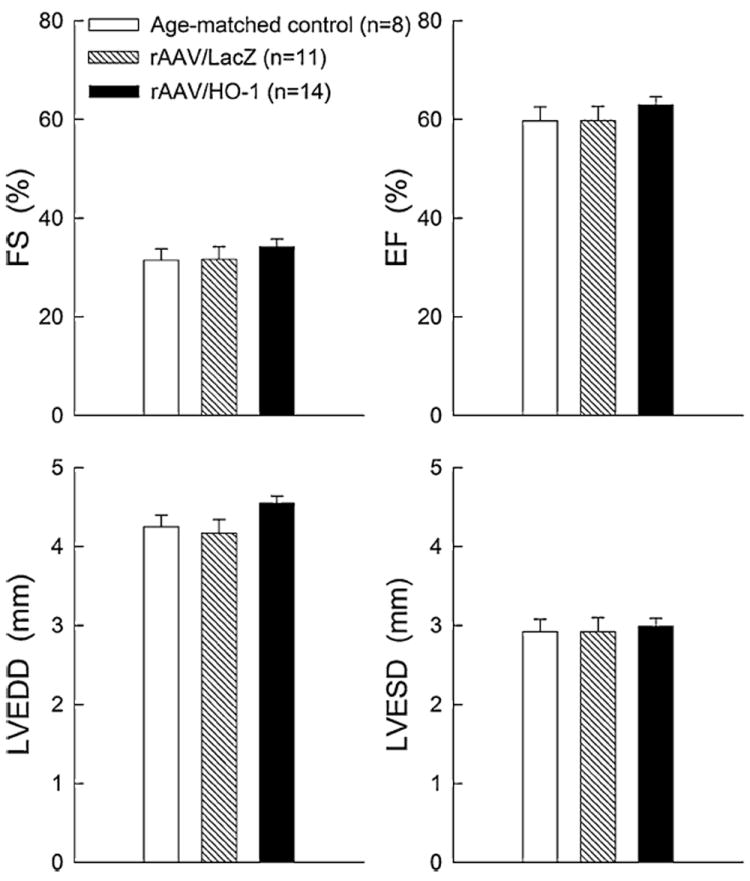
Quantitative analysis of echocardiographic LV functional parameters at 1 year after rAAV-mediated gene transfer. FS fractional shortening, EF ejection fraction, LVEDD left ventricular (LV) end-diastolic diameter, LVESD LV end-systolic diameter. Data are mean ± SEM
Discussion
This study demonstrates, for the first time, that transfer of the HO-1 gene to the myocardium via an rAAV vector enables long-lasting transgene expression (at least 1 year) and affords chronic, possibly permanent (1 year), protection against ischemia/reperfusion injury without adverse functional consequences. The finding that HO-1 gene transfer can reproduce the salubrious effects of the late phase of ischemic PC, but dramatically extend their duration (from 3 days to at least 1 year) supports a novel therapeutic strategy for achieving long-term prophylactic cardioprotection in patients with ischemic heart disease based on the transfer to the heart of HO-1 or other genes implicated in late preconditioning. This strategy can be thought of as an “immunization” of sorts against infarction.
Methodological considerations
The murine model of MI used in the present study has been well characterized [11, 12, 14, 26]. This murine model is physiologically relevant and provides reliable measurements. The experimental conditions in this murine model have been rigorously controlled to avoid a number of factors that could interfere with MI, including anesthesia, trauma, temperature fluctuations, and abnormal hemodynamics [14].
The choice of the HO-1 gene was predicated upon several considerations. Numerous genes have been reported to exert cardioprotective actions during myocardial ischemia/reperfusion [20, 46] and thus could, in principle, be candidates for gene transfer [7]. The rationale for studying HO-1 gene transfer is the discovery that HO-1 appears to be a central molecular mechanism whereby the heart protects itself from ischemia [13, 26] and that HO-1 exerts potent cytoprotective effects in the context of myocardial ischemia/reperfusion injury [34, 37, 44]. Myocardial ischemia leads to rapid induction of HO-1 [32-36] and pharmacologic induction of HO-1 by pretreatment with hemin ameliorates ischemia/reperfusion injury [17]. Either HO-1 gene therapy [36] or transgenic overexpression of HO-1 [44] provides powerful protection against ischemia/reperfusion injury; conversely, hearts isolated from heterozygous HO-1 deficient mice exhibit increased ischemia/reperfusion injury and oxidative stress [45]. At least in part, the salutary effects of HO-1 are due to the fact that this system serves as a critical defense against oxidative stress [8, 13, 21, 34]. However, despite the fact that the experimental evidence supporting a cytoprotective role of HO-1 activity is cogent, this knowledge has not yet been harnessed to develop long-term anti-ischemic strategies. Consequently, in the present study we created an rAAV vector carrying the therapeutic gene HO-1.
rAAV vectors are receiving increasing attention in the field of gene therapy because of their ability to program long-term, stable, and efficient gene expression in both dividing and nondividing cells in vivo with no inflammatory reaction, by integrating themselves into the host chromosome 19 in a site-specific manner [18, 40]. Although recombinant adenovirus 5 vectors (rAd5) provide a reliable method for high efficiency gene transfer into adult cardiomyocytes and for directly altering the cardiac phenotype [15, 22, 26, 27, 30], the long-term therapeutic use of rAd vectors is inherently limited by the transient expression of the transgene only for 1–2 weeks due to the host vigorous inflammatory response to viral proteins [38]. Furthermore, a number of human trials using rAAV vectors for cystic fibrosis and hemophilia B are ongoing, and so far the initial safety results from clinical trials of rAAV therapy are promising without reports of toxicity [9, 33], suggesting that rAAV vectors may have important applications for gene delivery in the clinical arena by affording long-lasting prophylactic protection in patients with coronary artery disease.
Effects of rAAV/HO-1-mediated gene transfer in vivo
Utilizing the rAAV system, we found that the rAAV/HO-1 vector enabled expression of HO-1 protein in the transduced myocardium not only at 1 month (+5.9-fold) but also at 1 year (+7.7-fold) after rAAV-mediated HO-1 gene transfer as compared to mice transfected with the rAAV/LacZ vector, respectively (n = 4/group, P < 0.05; Fig. 2a). Consistent with the Western immunoblotting results, immunohistochemistry analysis revealed increased levels of HO-1 protein expression in the myocardium (Fig. 3). Moreover, the content of bilirubin, a byproduct of HO-1, was significantly increased in the transduced myocardium 1 year after HO-1 gene transfer (Fig. 2b), indicating the ability of the rAAV/HO-1 vector to express functionally competent HO-1 protein for up to 1 year in vivo, which is critical to the long-term gene therapeutic approach against myocardial ischemia/reperfusion injury.
Long-term effects of HO-1 gene therapy on the heart
In the present study, we compared the long-term (1 year) effects of HO-1 gene therapy with the infarct-sparing effect of late PC, which is considered as the best example of a sustained cardioprotected phenotype for 3 days [6, 7, 39, 41]. As illustrated in Fig. 4, the reduction in infarct size (% of risk region) afforded by HO-1 gene therapy 1 year after gene transfer was comparable to that afforded by the late PC 24 h after the PC stimulus, demonstrating that rAAV-mediated HO-1 gene therapy produced robust and long-lasting cardioprotection.
Besides the control group subjected to rAAV/LacZ gene transfer, an age-matched control group was studied to monitor any significant changes in cardiac function associated with aging. As showed in Fig. 3, immunohistochemistry analysis demonstrated no significant changes in myocyte morphology 1 year after rAAV-mediated HO-1 gene transfer as compared to the control LacZ group. Furthermore, echocardiographic measurements of cardiac function at 1 year after gene transfer showed no evidence of a deleterious effect of HO-1 gene transfer on LV fractional shortening, LV ejection fraction, and LV end-diastolic or end-systolic diameter as compared to the LacZ group and to the age-matched controls (Fig. 7).
The use of rAAV-mediated HO-1 gene transfer in vivo to elevate endogenous HO-1 levels in the myocardium obviates the problems associated with using HO-1 inducers or HO-1 byproducts (such as hemin [17], carbon monoxide donors [13] or biliverdin [23]) to protect against MI, which need to be given parenterally and repeatedly and/or have short plasma half-lives. The long-term effects of HO-1 gene therapy in the heart, coupled with the fact that HO-1 protein expression in the transduced myocardium persisted for 1 year, suggest that rAAV-mediated HO-1 gene transfer in the heart may have potential clinical utility as a strategy for achieving “immunization” against MI in patients with coronary artery disease.
Conclusions
This study demonstrates, for the first time, that transfer of the HO-1 gene to the myocardium via rAAV vectors affords chronic, possibly permanent (1 year), protection against ischemia/reperfusion injury without adverse consequences on cardiac function. The results not only emphasize the central cardioprotective role of HO-1 in the heart, but also provide a framework for developing novel cardioprotective interventions aimed at rendering the myocardium permanently tolerant to ischemia/reperfusion injury. Unlike pharmacologically-induced protection by drugs given at the time of reperfusion or the 3-day protection afforded by late PC, rAAV/HO-1-mediated gene therapy enables chronic and uninterrupted protection, and thus alleviates ischemic damage whenever it occurs for at least 1 year. These findings provide a rationale for preclinical testing of HO-1 gene therapy as a prophylaxis against infarction.
Acknowledgments
This study was supported in part by NIH grants R01 HL55757, HL-70897, HL-76794, and P01HL78825.
Contributor Information
Qianhong Li, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA.
Yiru Guo, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA.
Qinghui Ou, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA.
Wen-Jian Wu, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA.
Ning Chen, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA.
Xiaoping Zhu, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA.
Wei Tan, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA.
Fangping Yuan, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA.
Buddhadeb Dawn, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA; Cardiovascular Research Institute, University of Kansas Medical Center, Kansas City, KS 66160, USA.
Li Luo, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA.
Gregory N. Hunt, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA
Roberto Bolli, Email: rbolli@louisville.edu, Institute of Molecular Cardiology, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA; Division of Cardiovascular Medicine, University of Louisville, 550 S. Jackson Street, Louisville, KY 40292, USA.
References
- 1.Abraham NG, Asija A, Drummond G, Peterson S. Heme oxygenase-1 gene therapy: recent advances and therapeutic applications. Curr Gene Ther. 2007;7:89–108. doi: 10.2174/156652307780363134. [DOI] [PubMed] [Google Scholar]
- 2.Abraham NG, Kappas A. Heme oxygenase and the cardiovascular-renal system. Free Radic Biol Med. 2005;39:1–25. doi: 10.1016/j.freeradbiomed.2005.03.010. [DOI] [PubMed] [Google Scholar]
- 3.Abraham NG, Kappas A. Pharmacological and clinical aspects of heme oxygenase. Pharmacol Rev. 2008;60:79–127. doi: 10.1124/pr.107.07104. [DOI] [PubMed] [Google Scholar]
- 4.Akagi R, Takahashi T, Sassa S. Cytoprotective effects of heme oxygenase in acute renal failure. Contrib Nephrol. 2005;148:70–85. doi: 10.1159/000086044. [DOI] [PubMed] [Google Scholar]
- 5.Alam J, Cook JL. How many transcription factors does it take to turn on the heme oxygenase-1 gene? Am J Respir Cell Mol Biol. 2007;36:166–174. doi: 10.1165/rcmb.2006-0340TR. [DOI] [PubMed] [Google Scholar]
- 6.Bolli R. The late phase of preconditioning. Circ Res. 2000;87:972–983. doi: 10.1161/01.res.87.11.972. [DOI] [PubMed] [Google Scholar]
- 7.Bolli R, Li QH, Tang XL, Guo Y, Xuan YT, Rokosh G, Dawn B. The late phase of preconditioning and its natural clinical application–gene therapy. Heart Fail Rev. 2007;12:189–199. doi: 10.1007/s10741-007-9031-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dawn B, Bolli R. HO-1 induction by HIF-1: a new mechanism for delayed cardioprotection? Am J Physiol Heart Circ Physiol. 2005;289:H522–H524. doi: 10.1152/ajpheart.00274.2005. [DOI] [PubMed] [Google Scholar]
- 9.Flotte TR, Schwiebert EM, Zeitlin PL, Carter BJ, Guggino WB. Correlation between DNA transfer and cystic fibrosis airway epithelial cell correction after recombinant adeno-associated virus serotype 2 gene therapy. Hum Gene Ther. 2005;16:921–928. doi: 10.1089/hum.2005.16.921. [DOI] [PubMed] [Google Scholar]
- 10.Gardin JM, Siri FM, Kitsis RN, Edwards JG, Leinwand LA. Echocardiographic assessment of left ventricular mass and systolic function in mice. Circ Res. 1995;76:907–914. doi: 10.1161/01.res.76.5.907. [DOI] [PubMed] [Google Scholar]
- 11.Guo Y, Bao W, Wu WJ, Shinmura K, Tang XL, Bolli R. Evidence for an essential role of cyclooxygenase-2 as a mediator of the late phase of ischemic preconditioning in mice. Basic Res Cardiol. 2000;95:479–484. doi: 10.1007/s003950070024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Guo Y, Jones WK, Xuan YT, Tang XL, Bao W, Wu WJ, Han H, Laubach VE, Ping P, Yang Z, Qiu Y, Bolli R. The late phase of ischemic preconditioning is abrogated by targeted disruption of the inducible NO synthase gene. Proc Natl Acad Sci USA. 1999;96:11507–11512. doi: 10.1073/pnas.96.20.11507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Guo Y, Stein AB, Wu WJ, Tan W, Zhu X, Li QH, Dawn B, Motterlini R, Bolli R. Administration of a CO-releasing molecule at the time of reperfusion reduces infarct size in vivo. Am J Physiol Heart Circ Physiol. 2004;286:H1649–H1653. doi: 10.1152/ajpheart.00971.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Guo Y, Wu WJ, Qiu Y, Tang XL, Yang Z, Bolli R. Demonstration of an early and a late phase of ischemic preconditioning in mice. Am J Physiol. 1998;275:H1375–H1387. doi: 10.1152/ajpheart.1998.275.4.H1375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hajjar RJ, Schmidt U, Matsui T, Guerrero JL, Lee KH, Gwathmey JK, Dec GW, Semigran MJ, Rosenzweig A. Modulation of ventricular function through gene transfer in vivo. Proc Natl Acad Sci USA. 1998;95:5251–5256. doi: 10.1073/pnas.95.9.5251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hajjar RJ, Zsebo K. AAV vectors and cardiovascular disease: targeting TNF receptor in the heart: clue to way forward with AAV? Gene Ther. 2007;14:1611–1612. doi: 10.1038/sj.gt.3303047. [DOI] [PubMed] [Google Scholar]
- 17.Hangaishi M, Ishizaka N, Aizawa T, Kurihara Y, Taguchi J, Nagai R, Kimura S, Ohno M. Induction of heme oxygenase-1 can act protectively against cardiac ischemia/reperfusion in vivo. Biochem Biophys Res Commun. 2000;279:582–588. doi: 10.1006/bbrc.2000.3973. [DOI] [PubMed] [Google Scholar]
- 18.Henckaerts E, Linden RM. Adeno-associated virus: a key to the human genome? Future Virol. 2010;5:555–574. doi: 10.2217/fvl.10.48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hester ME, Foust KD, Kaspar RW, Kaspar BK. AAV as a gene transfer vector for the treatment of neurological disorders: novel treatment thoughts for ALS. Curr Gene Ther. 2009;9:428–433. doi: 10.2174/156652309789753383. [DOI] [PubMed] [Google Scholar]
- 20.Heusch G, Boengler K, Schulz R. Cardioprotection: nitric oxide, protein kinases, and mitochondria. Circulation. 2008;118:1915–1919. doi: 10.1161/CIRCULATIONAHA.108.805242. [DOI] [PubMed] [Google Scholar]
- 21.Jiang Y, Chen L, Tang Y, Ma G, Shen C, Qi C, Zhu Q, Yao Y, Liu N. HO-1 gene overexpression enhances the beneficial effects of superparamagnetic iron oxide labeled bone marrow stromal cells transplantation in swine hearts underwent ischemia/reperfusion: an MRI study. Basic Res Cardiol. 2010;105:431–442. doi: 10.1007/s00395-009-0079-2. [DOI] [PubMed] [Google Scholar]
- 22.Jones JM, Wilson KH, Koch WJ, Milano CA. Adenoviral gene transfer to the heart during cardiopulmonary bypass: effect of myocardial protection technique on transgene expression. Eur J Cardiothorac Surg. 2002;21:847–852. doi: 10.1016/s1010-7940(02)00078-7. [DOI] [PubMed] [Google Scholar]
- 23.Kapitulnik J, Maines MD. Pleiotropic functions of biliverdin reductase: cellular signaling and generation of cytoprotective and cytotoxic bilirubin. Trends Pharmacol Sci. 2009;30:129–137. doi: 10.1016/j.tips.2008.12.003. [DOI] [PubMed] [Google Scholar]
- 24.Katori M, Buelow R, Ke B, Ma J, Coito AJ, Iyer S, Southard D, Busuttil RW, Kupiec-Weglinski JW. Heme oxygenase-1 overexpression protects rat hearts from cold ischemia/reperfusion injury via an antiapoptotic pathway. Transplantation. 2002;73:287–292. doi: 10.1097/00007890-200201270-00023. [DOI] [PubMed] [Google Scholar]
- 25.Li Q, Guo Y, Ou Q, Chen N, Wu WJ, Yuan F, O’Brien E, Wang T, Luo L, Hunt GN, Zhu X, Bolli R. Intracoronary administration of cardiac stem cells in mice: a new, improved technique for cell therapy in murine models. Basic Res Cardiol. 2011 doi: 10.1007/s00395-011-0180-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Li Q, Guo Y, Ou Q, Cui C, Wu WJ, Tan W, Zhu X, Lanceta LB, Sanganalmath SK, Dawn B, Shinmura K, Rokosh GD, Wang S, Bolli R. Gene transfer of inducible nitric oxide synthase affords cardioprotection by upregulating heme oxygenase-1 via a nuclear factor-{kappa}B-dependent pathway. Circulation. 2009;120:1222–1230. doi: 10.1161/CIRCULATIONAHA.108.778688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Li Q, Guo Y, Tan W, Ou Q, Wu WJ, Sturza D, Dawn B, Hunt G, Cui C, Bolli R. Cardioprotection afforded by inducible nitric oxide synthase gene therapy is mediated by cyclooxygenase-2 via a nuclear factor-kappaB dependent pathway. Circulation. 2007;116:1577–1584. doi: 10.1161/CIRCULATIONAHA.107.689810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Li Q, Guo Y, Tan W, Stein AB, Dawn B, Wu WJ, Zhu X, Lu X, Xu X, Siddiqui T, Tiwari S, Bolli R. Gene therapy with iNOS provides long-term protection against myocardial infarction without adverse functional consequences. Am J Physiol Heart Circ Physiol. 2006;290:H584–H589. doi: 10.1152/ajpheart.00855.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li Q, Guo Y, Wu W, Ou Q, Zhu X, Tan W, Yuan F, Chen N, Dawn B, Luo L, O’Brien E, Bolli R. Gene transfer as a strategy to achieve permanent cardioprotection I: rAAV-mediated gene therapy with inducible nitric oxide synthase limits infarct size 1 year later without adverse functional consequences. Basic Res Cardiol. 2011 doi: 10.1007/s00395-011-0207-7. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Li Q, Guo Y, Xuan YT, Lowenstein CJ, Stevenson SC, Prabhu SD, Wu WJ, Zhu Y, Bolli R. Gene therapy with inducible nitric oxide synthase protects against myocardial infarction via a cyclooxygenase-2-dependent mechanism. Circ Res. 2003;92:741–748. doi: 10.1161/01.RES.0000065441.72685.29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Liu Y, Li P, Lu J, Xiong W, Oger J, Tetzlaff W, Cynader M. Bilirubin possesses powerful immunomodulatory activity and suppresses experimental autoimmune encephalomyelitis. J Immunol. 2008;181:1887–1897. doi: 10.4049/jimmunol.181.3.1887. [DOI] [PubMed] [Google Scholar]
- 32.Lu R, Peng J, Xiao L, Deng HW, Li YJ. Heme oxygenase-1 pathway is involved in delayed protection induced by heat stress against cardiac ischemia-reperfusion injury. Int J Cardiol. 2002;82:133–140. doi: 10.1016/s0167-5273(01)00612-x. [DOI] [PubMed] [Google Scholar]
- 33.Manno CS, Chew AJ, Hutchison S, Larson PJ, Herzog RW, Arruda VR, Tai SJ, Ragni MV, Thompson A, Ozelo M, Couto LB, Leonard DG, Johnson FA, McClelland A, Scallan C, Skarsgard E, Flake AW, Kay MA, High KA, Glader B. AAV-mediated factor IX gene transfer to skeletal muscle in patients with severe hemophilia B. Blood. 2003;101:2963–2972. doi: 10.1182/blood-2002-10-3296. [DOI] [PubMed] [Google Scholar]
- 34.Marini M, Lapalombella R, Margonato V, Ronchi R, Samaja M, Scapin C, Gorza L, Maraldi T, Carinci P, Ventura C, Veicsteinas A. Mild exercise training, cardioprotection and stress genes profile. Eur J Appl Physiol. 2007;99:503–510. doi: 10.1007/s00421-006-0369-4. [DOI] [PubMed] [Google Scholar]
- 35.Matsushita T, Elliger S, Elliger C, Podsakoff G, Villarreal L, Kurtzman GJ, Iwaki Y, Colosi P. Adeno-associated virus vectors can be efficiently produced without helper virus. Gene Ther. 1998;5:938–945. doi: 10.1038/sj.gt.3300680. [DOI] [PubMed] [Google Scholar]
- 36.Melo LG, Agrawal R, Zhang L, Rezvani M, Mangi AA, Ehsan A, Griese DP, Dell’Acqua G, Mann MJ, Oyama J, Yet SF, Layne MD, Perrella MA, Dzau VJ. Gene therapy strategy for long-term myocardial protection using adeno-associated virus-mediated delivery of heme oxygenase gene. Circulation. 2002;105:602–607. doi: 10.1161/hc0502.103363. [DOI] [PubMed] [Google Scholar]
- 37.Morse D, Choi AM. Heme oxygenase-1: the “emerging molecule” has arrived. Am J Respir Cell Mol Biol. 2002;27:8–16. doi: 10.1165/ajrcmb.27.1.4862. [DOI] [PubMed] [Google Scholar]
- 38.Muruve DA. The innate immune response to adenovirus vectors. Hum Gene Ther. 2004;15:1157–1166. doi: 10.1089/hum.2004.15.1157. [DOI] [PubMed] [Google Scholar]
- 39.Post H, Heusch G. Ischemic preconditioning. Experimental facts and clinical perspective. Minerva Cardioangiol. 2002;50:569–605. [PubMed] [Google Scholar]
- 40.Samulski RJ, Zhu X, Xiao X, Brook JD, Housman DE, Epstein N, Hunter LA. Targeted integration of adeno-associated virus (AAV) into human chromosome 19. EMBO J. 1991;10:3941–3950. doi: 10.1002/j.1460-2075.1991.tb04964.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Thielmann M, Kottenberg E, Boengler K, Raffelsieper C, Neuhaeuser M, Peters J, Jakob H, Heusch G. Remote ischemic preconditioning reduces myocardial injury after coronary artery bypass surgery with crystalloid cardioplegic arrest. Basic Res Cardiol. 2010;105:657–664. doi: 10.1007/s00395-010-0104-5. [DOI] [PubMed] [Google Scholar]
- 42.Vivaldi MT, Kloner RA, Schoen FJ. Triphenyltetrazolium staining of irreversible ischemic injury following coronary artery occlusion in rats. Am J Pathol. 1985;121:522–530. [PMC free article] [PubMed] [Google Scholar]
- 43.Wright JF, Qu G, Tang C, Sommer JM. Recombinant adeno-associated virus: formulation challenges and strategies for a gene therapy vector. Curr Opin Drug Discov Devel. 2003;6:174–178. [PubMed] [Google Scholar]
- 44.Yet SF, Tian R, Layne MD, Wang ZY, Maemura K, Solovyeva M, Ith B, Melo LG, Zhang L, Ingwall JS, Dzau VJ, Lee ME, Perrella MA. Cardiac-specific expression of heme oxygenase-1 protects against ischemia and reperfusion injury in transgenic mice. Circ Res. 2001;89:168–173. doi: 10.1161/hh1401.093314. [DOI] [PubMed] [Google Scholar]
- 45.Yoshida T, Maulik N, Ho YS, Alam J, Das DK. H(mox-1) constitutes an adaptive response to effect antioxidant cardioprotection: a study with transgenic mice heterozygous for targeted disruption of the heme oxygenase-1 gene. Circulation. 2001;103:1695–1701. doi: 10.1161/01.cir.103.12.1695. [DOI] [PubMed] [Google Scholar]
- 46.Zhao HX, Wang XL, Wang YH, Wu Y, Li XY, Lv XP, Zhao ZQ, Zhao RR, Liu HR. Attenuation of myocardial injury by postconditioning: role of hypoxia inducible factor-1alpha. Basic Res Cardiol. 2010;105:109–118. doi: 10.1007/s00395-009-0044-0. [DOI] [PubMed] [Google Scholar]