

Abstract
Evidence for flexible impulse control over food consumption is rare in non-human animals. So far, only primates and corvids have been shown to be able to fully inhibit the consumption of a desirable food item in anticipation for a gain in quality or quantity longer than a minute. We tested Goffin cockatoos (Cacatua goffini) in an exchange task. Subjects were able to bridge delays of up to 80 s for a preferred food quality and up to 20 s for a higher quantity, providing the first evidence for temporal discounting in birds that do not cache food.
Keywords: delay of gratification, food exchange, impulse control, avian cognition
1. Introduction
The ability to inhibit the impulse to accept an instant option in anticipation for a delayed, more valuable one has long been believed to be a uniquely human attribute [1]. Indeed, rodents and most birds tested wait only few seconds for a delayed gain, and many monkeys wait less than a minute [2–7]. Some primates and dogs (possibly as an effect of domestication) do, however, accept delays over a minute and even show outstanding plasticity in decision-making relative to the benefits involved [8–11].
In delayed gratification tasks (DGTs), the point at which a subject decides to give up can provide information about the subject's representation of the delay: the subject should either renounce waiting early (rather than at a random point of time) or wait out the entire delay if it has a presumption about delay duration [10]. In exchange DGTs, subjects are given an immediate reward as a trade currency and can choose to keep it intact throughout the delay or finish the trial by starting to eat [7,10,12,13]. The value of the delayed reward can surpass the immediate one either in quality or quantity [7]. In the accumulated DGT, by contrast, the rewards mount up in quantity in the course of the delay and subjects can discontinue the accumulation through interference [8,14].
The superior performance of primates in DGTs was recently challenged by Dufour et al. [12], who showed that corvids can wait up to 5 min in a qualitative exchange DGT. Common ravens (Corvus corax) and carrion crows (Corvus corone) waited longer if the disproportion in value was large, and when they renounced waiting they tended to do so instantly rather than in the middle of the delay [12]. During longer delays, subjects temporarily deposited food before returning it, suggesting that their inhibition control was facilitated through being food hoarders [12]. Interestingly, the same crows failed to wait for higher quantities in an exchange DGT, despite choosing higher quantities in a binary preference test [13].
Parrots and corvids frequently are comparable with primates in problem-solving and innovative skills, and it has been suggested that many of the cognitive mechanisms of parrots and corvids are the result of convergent evolution [15]. Three African grey (Psittacus erithacus) parrots tested in an accumulated DGT, however, failed to inhibit their responses for longer than three seconds [14]. Possibly, their performance was hampered by the nature of the task (accumulation versus exchange) or the fact that they were asked to maximize quantity (which corvids failed as well). We thus tested another psittacine, the Goffin cockatoo (Cacatua goffini), on a qualitative and quantitative exchange DGT. These cockatoos previously showed high levels of persistence and patience in sequential problem-solving tasks (Auersperg et al. [16]) and, as feeding generalists [17], are likely to possess high levels of flexibility. If Goffins can inhibit food consummation in an exchange task, they should wait for a better quality and/or quantity. If reward type affects their performance, they should wait longer for their most-preferred than for their second-preferred food. If the inhibitory skills of corvids are enhanced by their feeding ecology (food caching), the performance of Goffins should be inferior to those of corvids.
2. Material and methods
We tested 14 subjects. They received a quality preference test in which preferences for three desirable foods (pecan nut, fried meat and cashew nut) were identified and a quantity preference test to confirm their ability to discriminate between different quantities of the same food (cashew; at 1 : 2 and 1 : 6 ratios), either with intact pieces of different sizes or one piece versus several equally sized pieces (see the electronic supplementary material). Thereafter, they were trained to return an inedible object into the experimenter's hands to receive a reward from the other hand (see the electronic supplementary material). In subsequent tests, subjects could exchange an intact initial food item for an expected one of different quality or quantity (see the electronic supplementary material, movie S1) after different time delays, starting at 2 s, then 5, 20, 40, 80 and 160 s (until they quit exchanging). The procedure went as follows: first both open hands of the experimenter (one containing the initial and one the expected item) were shown to the subject. Once the subject left its starting position, the hand containing the expected item was moved out of reach (with the food still visible). The subject was allowed to pick up the initial item and the (now empty) hand was closed to a fist. If the subject dropped the initial item off the table or started to nibble it, the experimenter immediately closed the hand containing the expected item and the trial ended. If the reward was still intact after the delay, the experimenter reopened the fist and the subject had the opportunity to return the initial item. In this case, the other hand moved back into reach and the subject could take the expected item. The delayed quality exchange task (DQL) comprised seven sessions of nine trials: three trials each, in which subjects could exchange the initial item (pecan) for cashew (most preferred by the majority of birds) or for meat (second in the food hierarchy for the majority of birds) and three control trials in which the initial item was cashew and subjects could exchange for meat (control 1) or pecan (control 2; the frequency of each control was balanced across sessions). In the delayed quantity exchange task (DQN), subjects received six trials with two ratios (1 : 2 and 1 : 6) both in pieces and as a whole. This, as well as the hand containing the larger quantity, was balanced (within one delay stage, every possible side and food combination was tested a minimum of three times). Each delay contained 12 controls in which the bigger quantity was the initial item. For further details see the electronic supplementary material.
3. Results
(a). Delayed quality exchange
All subjects readily exchanged pecan for preferred items at 2 and 5 s delays. Half of the birds waited 40 s and three even up to 80 s (see the electronic supplementary material, table S5).
The percentage of exchanges gradually decreased as waiting time increased (figure 1). Exchanges in control conditions started at low rates and vanished at 10 s. The mean number of exchanges for the most-preferred food item (MPF) varied between delays (MPF = cashew for 12 birds, meat for two birds; Friedman's ANOVA; χ2 = 16.435; p = 0.012), with subjects exchanging more in 5 than 10 s (Wilcoxon; z = 2.591, p = 0.01), 20 than 40 s (z = 2.521, p = 0.012) and 40 than 80 s (z = 2.201, p = 0.028). A similar pattern was found for the second preferred food (SPF; meat for 6 birds, cashew for 2 birds; Friedman's ANOVA; χ2 = 15.865; p = 0.014): the eight birds which preferred meat over pecan (see the electronic supplementary material, table S2) exchanged significantly more often for their MPF than their SPF in 5 s delays (z = 2.383; p = 0.017).
Figure 1.
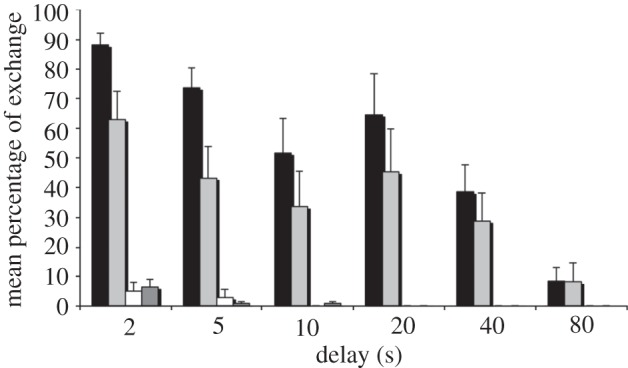
Mean percentage of successful exchanges in the DQL, for 2–80 s delays, for the two test and the two control conditions. Black bars represent test 1: pecan for most preferred; light grey bars represent test 2: pecan for second/least preferred; white bars represent control 1: cashew for meat and dark grey bars represents control 2: cashew for pecan.
Comparing our data with those of Dufour et al. [12], we found that cockatoos exchanged significantly more often than ravens in 2 and 5 s delays (Mann–Whitney U-test; z = 2.836; p = 0.005; z = 2.885; p = 0.004; paired tests for longer delays were not conducted owing to small sample sizes in corvids).
As observed in corvids, some Goffins renounced waiting earlier into a delay than predicted by chance (see the electronic supplementary material, table S6). The eight birds that preferred meat over pecan also gave up waiting significantly later in 5 s delays for their MPF, than for their SPF (Wilcoxon tests; z = 2.384, p = 0.017; note that paired testing of this for delays over 20 s was not possible due to decreased sample sizes). Furthermore, within a certain delay and condition birds either did not exchange at all or did so at high rates until ca 20 s delays before performance dropped (see the electronic supplementary material, tables S3 and S4; note: this is not reflected in figure 1, because subjects that had not yet been dropped from the test but did not exchange for a certain condition/delay are still accounted for).
(b). Delayed quantity exchange
Cockatoos also responded to the quantity of the offered reward. Eight of 14 cockatoos exchanged lower for higher quantities for up to 20 s delays. However, they did so at lower rates than in the DQL (figure 2). The exception was one female that still exchanged in 70 per cent of the cases in the 10 s delay in the 1 : 6 condition. Again, subjects mostly renounced waiting earlier into a delay than predicted by chance (see the electronic supplementary material, table S7).
Figure 2.
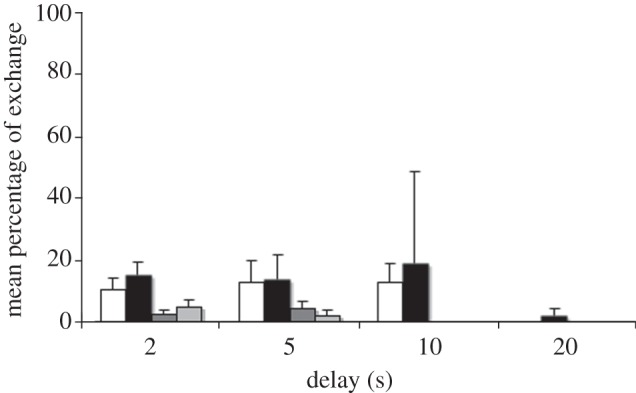
Mean percentage of successful exchanges in the DQN, for 2–20 s delays, for the two test (white, 1 : 2; black, 1 : 6) and the two control (dark grey, 2 : 1; light grey, 6 : 1) conditions.
Interestingly, we did not find any differences in the percentage of exchanges between the larger quantities presented in equally sized pieces or as a bigger piece, neither for the 1 : 2 nor for the 1 : 6 ratios (Wilcoxon test; all p > 0.05). Moreover, there were no significant differences in the percentage of exchange between bigger or smaller quantity (neither for the 1 : 2 nor for the 1 : 6 ratio), in the number of exchanges between the 1 : 6 and the 1 : 2 ratio, in the mean give up times between 1 : 6 and 1 : 2, or in the mean percentage of successful exchanges between the two ratios for any of the stages (Wilcoxon tests; all p > 0.05).
In cases where birds did not exchange, they usually ate the food (95.2%, DQL and DQN combined); rarely, the food was dropped off the experimental table. Unlike corvids, Goffins hardly ever deposited the food on the table and seemed to have difficulties leaving it there, immediately picking it up again. Longer delays (greater than 10 s) were often characterized by a number of frustration-induced, idiosyncratic behaviours (see the electronic supplementary material, table S8 and movie S2).
4. Discussion
Our results indicate that Goffin cockatoos clearly possess impulse control over extended delays in DGTs. They further seem to attribute value to tokens and to tolerate loss of an initial item in anticipation for a gained compensation. Like corvids and some primates, they acted much like economic agents, flexibly trading-off between immediate and future benefits relative not only to the length of delay, but also to the difference in trade value between the currency and the item on auction [6,10,12]. Goffins tended to trade the initial item more often for the most-preferred food than for a less desirable one and hardly ever exchanged in controls in which the initial value was higher than the expected. Similar to apes and corvids, Goffins tended to either renounce waiting early in a trial or waited the entire delay, indicating an ability to assess the duration of the delay [10,12]. Furthermore, they seemed to additionally judge if a gain was worth waiting for relative to its expected value, giving up later for a better gain.
Although Goffins initially exchanged at higher rates than corvids, their longest waiting times did not exceed 80 s, whereas some corvids waited up to 5 min [12]. Goffins exhibited a low inhibitory threshold for depositing the initial item onto the table before exchange opportunities, whereas crows and ravens temporarily discarded the initial food in all delays over 20 s [12]. Hence, food hoarding species may have an advantage over non-hoarders in exchange tasks in that they ‘can let go’ and do not have to hold the initial item in the beak, close to their taste organs, throughout the entire delay. However, the overall performance of Goffins indicates that food caching is not a precondition for birds to evolve mechanisms for coping with a delayed gratification.
Unlike crows [13], of which only three exchanged at very low rates and only at the 2 s delay in quantity DGTs, eight out of 14 Goffins still exchanged for up to 20 s and one at high rates until 10 s delays. However, Goffins’ exchange rates were much lower in DQN than DQL, fitting the assumption that birds tend to show more impulse control for maximizing quality than quantity. There are two potential explanations for this behaviour: either a failure to attribute to quantity the same value as to quality as being worth waiting for [13], or the rewards used in quantitative tasks in avian studies were too delectable to inhibit consummation. Future studies on quantitative DGTs should thus incorporate several reward types.
Taken together, our results provide the first evidence for high levels of impulse control in a parrot species in DGTs. Goffin cockatoos were not only able to wait for gains in quality but also, to some extent, quantity. These findings suggest that an extended temporal horizon for decisions concerning food may have evolved convergently within birds (corvids and parrots) and between birds and primates [12].
Acknowledgements
We thank Valerie Dufour, Claudia Wascher and Laurent Aman for their advice, and Philipp Stöger-Haselböck for interobserver reliability rating. This work was supported by the FWF (Austrian Science Fund) Project no. Y366-B03 to T.B. and Project no. J 3404-B19 to A.M.I.A.
References
- 1.Mischel W, Shoda Y, Rodriguez ML. 1989. Delay of gratification in children. Science 244, 933–938 10.1111/j.1461-0248.2009.01299.x (doi:10.1111/j.1461-0248.2009.01299.x) [DOI] [PubMed] [Google Scholar]
- 2.Richards JB, Mitchell SH, de Wit H, Seiden LS. 1997. Determination of discount functions in rats with an adjusting-amount procedure. J. Exp. Anal. Behav. 67, 353–366 10.1038/379718a0 (doi:10.1038/379718a0) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Abeyesinghe SM, Nicol CJ, Hartnell SJ, Wathes CM. 2005. Can domestic fowl, Gallus gallus domesticus, show self-control? Anim. Behav. 70, 1–11 10.1111/j.1461-0248.2009.01356.x (doi:10.1111/j.1461-0248.2009.01356.x) [DOI] [Google Scholar]
- 4.Ainslie G. 1974. Impulse control in pigeons. J. Exp. Anal. Behav. 21, 485–489 10.1111/j.1461-0248.2010.01548.x (doi:10.1111/j.1461-0248.2010.01548.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hidetoshi A, Ai K, Toshiya M. 2010. Social influence of competition on impulsive choices in domestic chicks. Biol. Lett. 6, 183–186 10.1126/science.1128326 (doi:10.1126/science.1128326) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Stevens JR, Hallinan EV, Hauser MD. 2005. The ecology and evolution of patience in two New World monkeys. Biol. Lett. 1, 223–226 10.1098/rsbl.2004.0285 (doi:10.1098/rsbl.2004.0285) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ramseyer A, Pele M, Dufour V, Chauvin C, Thierry B. 2006. Accepting loss: the temporal limits of reciprocity in brown capuchin monkeys. Proc. R. Soc. B 273, 179–184 10.1890/10-0999.1 (doi:10.1890/10-0999.1) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Beran MJ. 2002. Maintenance of self-imposed delay of gratification by four chimpanzees (Pan troglodytes) and an orangutan (Pongo pygmaeus). J. Gen. Psychol. 129, 49–66 10.1371/journal.pone.0044247 (doi:10.1371/journal.pone.0044247) [DOI] [PubMed] [Google Scholar]
- 9.Rosati AG, Stevens JR, Hare B, Hauser MD. 2007. The evolutionary origins of human patience: temporal preferences in chimpanzees, bonobos and human adults. Curr. Biol. 17, 1663–1668 10.1098/rspb.2011.0239 (doi:10.1098/rspb.2011.0239) [DOI] [PubMed] [Google Scholar]
- 10.Dufour V, Pele M, Sterck EHM, Thierry B. 2007. Chimpanzee (Pan troglodytes) anticipation of food return: coping with waiting time in an exchange task. J. Comp. Psych. 121, 145–155 10.1098/rspb.2012.0893 (doi:10.1098/rspb.2012.0893) [DOI] [PubMed] [Google Scholar]
- 11.Leonardi RJ, Vick S-J, Dufour V. 2012. Waiting for more: the performance of domestic dogs (Canis familiaris) on exchange tasks. Anim. Cogn. 15, 107–120 10.1007/s10071-011-0437-y (doi:10.1007/s10071-011-0437-y) [DOI] [PubMed] [Google Scholar]
- 12.Dufour V, Wascher CAF, Braun A, Miller R, Bugnyar T. 2011. Corvids can decide if a future exchange is worth waiting for. Biol. Lett. 8, 201–204 10.1371/journal.pone.0034403 (doi:10.1371/journal.pone.0034403) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wascher CAF, Dufour V, Bugnyar T. 2012. Carrion crows cannot overcome impulsive choice in a quantitative exchange task. Front. Comp. Psychol. 3, 118. 10.1038/35083573 (doi:10.1038/35083573) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Vick S-J, Bovet D, Anderson JR. 2009. How do African grey parrots (Psittacus erithacus) perform on a delay of gratification task? Anim. Cog. 13, 351–358 10.1086/587758 (doi:10.1086/587758) [DOI] [PubMed] [Google Scholar]
- 15.Emery NJ. 2006. Cognitive ornithology: the evolution of avian intelligence. Phil. Trans. R. Soc. B 361, 23–43 10.1007/s11829-009-9072-9 (doi:10.1007/s11829-009-9072-9) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Auersperg AMI, Kacelnik A, von Bayern AMP.Explorative learning and functional inferences on a five-step mechanical problem in Goffin's cockatoos (Cacatua goffini). Submitted. [DOI] [PMC free article] [PubMed]
- 17.Cahyadin Y, Jepson P, Manoppo BI. 1994. The status of Cacatua goffini and Eos reticulata on the Tanimbar islands. PHPA/Bird Life International Laporan No. 1