

Abstract
The serotonin transporter, encoded by the SLC6A4 gene, influences the synaptic actions of serotonin and is responsive to stress hormones. We hypothesized that 5-HTTLPR, a functional SLC6A4 promoter polymorphism, and two tightly-linked, putatively functional 3' UTR SNPs (rs3813034, rs1042173) might have independent effects on suicidal behavior in the context of childhood trauma (CT).
DNA and Childhood Trauma Questionnaire scores were available for a total of 474 African Americans, including 112 suicide attempters and 362 non-suicide attempters. Genotyping was performed for the triallelic 5-HTTLPR polymorphism, 14 SLC6A4 haplotype-tagging SNPs, and 186 ancestry informative markers.
There were independent G × E interactive effects of 5-HTTLPR (p=0.017) and the rs3813034-rs1042173 diplotype (p=0.011) on suicidal behavior. In individuals exposed to high CT the risk of suicide attempt was 0.52 in carriers of the low activity 5-HTTLPR variant and 0.32 in medium/high activity variant carriers. Likewise, CT exposed carriers of the major rs3813034-rs1042173 ATAT diplotype had an increased risk of suicidal behavior relative to the ATCG/CGCG diplotype carriers (0.40 vs 0.31). Neither the 5' nor the 3' functional variants had an effect in individuals without CT: suicide attempt risk = 0.12 – 0.22. In individuals exposed to high CT the prevalence of suicide attempt was 0.56 in carriers of both 5' and 3' risk variants, 0.39 in carriers of one risk variant and 0.25 in individuals without either risk variant.
Our findings suggest that the 5' and 3' SLC6A4 functional variants have independent effects on the risk for suicidal behavior in CT exposed individuals.
Keywords: SLC6A4, 5-HTTLPR, substance dependence, rs3813034, rs1042173, suicide attempt
INTRODUCTION
Variation in central nervous system serotonin function has been associated with negative mood states including anxiety, depression and suicidal behavior. Decreased 5-HIAA, the serotonin metabolite, has been observed in violent suicide attempters, completed suicide and violent offenders (Asberg et al., 1976). The serotonin transporter (5-HTT) is largely responsible for terminating the synaptic actions of serotonin.
A functional promoter polymorphism, 5-HTTLPR, in the 5-HTT gene (SLC6A4) has been associated with changes in neuronal circuitry implicated in negative affect (Hariri et al., 2005; Heinz et al., 2005; Pezawas et al., 2005). 5-HTTLPR is a common polymorphism including a variable number of tandem repeats (44-basepair insertion/deletion) in the promoter region that alters transcription such that the less common, short `S' allele is associated with an approximately 50% reduction in 5-HTT availability and concomitant increase in synaptic serotonin, reduced neuronal uptake of serotonin and decreased 5-HIAA (Heils et al., 1996). Triallelic genotyping (incorporating the functional SNP rs25531) of 5-HTTLPR identifies the low transcriptional activity S allele but additionally separates the high activity LA allele from a third allele, LG which displays low transcriptional activity similar to the S allele (Hu et al., 2006). This is particularly important when studying individuals of African ancestry where the frequency of LG is comparatively high.
5-HTT is sensitive to stress since SLC6A4 has a glucocorticoid response element (GRE) in the promoter region (Glatz et al., 2003). It has been postulated that SLC6A4 variation may to some extent, through influence on early life development of corticolimbic circuitry, modulate the capacity to cope with stress (Hariri and Holmes, 2006). Indeed, numerous studies in humans and non-human primates have demonstrated interactive effects of 5-HTTLPR and trauma on stress-related disorders (Caspi et al., 2010) including suicidal ideation and suicidal behavior (Caspi et al., 2003; Gibb et al., 2006; Roy et al., 2007; Roy et al., 2009).
Recent studies have suggested that there may also be a functional variant at the distal end of the gene. The 3' untranslated region (UTR) of SLC6A4 mRNA exists in two forms that differ by a 123 bp element (Battersby et al., 1999). The more distal of the polyadenylation signals includes a common SNP rs3813034 that has been shown to alter the balance of the two transcripts in the brain and lymphoblasts (Gyawali et al., 2010). This SNP has been associated with impaired retention of fear extinction memory and increased anxiety / depression (Gyawali et al., 2010; Hartley et al., 2012). Other researchers have shown that another SNP rs1042173, in allelic identity and strong linkage disequilibrium (LD) with rs3813034, is functional and is associated with alcohol craving and drinking intensity (Ait-Daoud et al., 2012; Seneviratne et al., 2009).
In an earlier study of a subset of the sample of African American substance dependent patients and controls that is the focus of the current study, we showed that the low activity 5-HTTLPR variant interacted with childhood trauma exposure to increase the risk of suicidal behavior (Roy et al., 2007). The aim of the present study was to determine whether there were independent effects of the 5' (5-HTTLPR) and 3' (rs3813034, rs1042173) functional variants on risk for suicidal behavior, including in the context of childhood trauma. In addition, in order to undertake a comprehensive analysis across SLC6A4 we performed secondary haplotype and SNP association and gene × environment interaction (G × E) analyses of suicidal behavior for 12 additional tag SNPs.
METHODS AND MATERIALS
Participants
A detailed description of the sample has previously been provided (Roy et al., 2012). A total of 706 African-American patients (90% men) were recruited from the Substance Abuse Treatment Program (SATP) at the Department of Veteran Affairs New Jersey Healthcare System (VANJHCS), East Orange Campus. All patients were abstinent and nearly all were inpatients however a few were recruited from the SATP outpatient clinic or from the methadone clinic. Their mean (SD) age was 45.2 (7.9) years. The patients included individuals with single and comorbid diagnoses of cocaine, alcohol and heroin dependence: 42% were dependent only on cocaine (16%) or alcohol (15%) or heroin (11%). Other patients had dual dependencies: cocaine + alcohol (23%), cocaine + heroin (14%) and alcohol + heroin (8%) and a further 13% were dependent on all three substances.
Patients were interviewed by a psychiatrist (A.R.) using the substance abuse section of the Structured Clinical Interview for DSM-IV (SCID) (Spitzer et al., 1995) to establish lifetime substance dependence diagnoses. The diagnosis of suicide attempt was implemented by A.R. after interviewing the patient with a standard series of clinical questions and reviewing all available collateral information. A suicide attempt was defined as a self-destructive act with some intent to end one's life that was not self-mutilatory in nature. Actions without suicide intent such as delicate self cutting and accidental drug overdose were excluded.
Controls (759 African American men (n = 307) and women (n = 452)) were recruited from the same geographical area as the patients. Controls were recruited from diabetic outpatients at an ophthalmology clinic (54%) at the University of Medicine and Dentistry, New Jersey Medical School (UMDNJ), Newark, NJ and from churches and a blood bank in Newark, NJ, (46%). Their mean (SD) age was 34.3 (10.1) years. The diabetics underwent two physician-conducted structured interviews over a 6 year period that included questions about suicidal behavior and substance abuse (Roy, 2000). The participants recruited from churches and a blood bank underwent a semi-structured psychiatric interview conducted by a trained research worker that included a standard series of questions about lifetime suicidal behavior and substance abuse/dependence.
After a full description of the study was provided, all participants gave written informed consent to the study that was approved by the Institutional Review Boards of the VANJHCS and UMDNJ.
Childhood Trauma Questionnaire (CTQ)
The CTQ (28 item version) (Bernstein et al., 1998) was completed by 538 patients (92% men) with substance dependence; 166 (90% men) had attempted suicide. The CTQ was completed by 342 non-substance dependent participants (42% men), 32 (28% men) of whom had attempted suicide.
The CTQ yields scores (5–25 each) for physical abuse, physical neglect, emotional abuse, emotional neglect, and sexual abuse, as well as a total score (25–125). Reliability and validity of the CTQ has been demonstrated, including in drug abusers and African American populations (reviewed in Roy et al., 2012). It is important to note that the CTQ was used as a continuous measure in all logistic regression analyses.
A dichotomous total CTQ score was derived solely to illustrate the direction of interaction effects on suicide attempt. A total CTQ score ≥ 1 SD above the mean CTQ score of non substance dependent / no suicide attempt participants (36.49 (11.67), i.e. ≥ 48) was designated `high trauma' (N = 290); lower total CTQ scores (< 48) were designated `low trauma' (N = 590). This approach has been used in our earlier studies (Roy et al., 2010; 2012).
Summary of Dataset
DNA was available for a total of 830 participants: 141 suicide attempters and 689 non-suicide attempters. For G × E analyses, both DNA and CTQ scores were available for a total of 474 individuals, including 112 suicide attempters and 362 non-suicide attempters. Missing DNA and CTQ data was random and showed no selection bias.
A flow diagram detailing the dataset, including the proportions of participants with and without substance dependence, can be found in supplemental information: see Figure S1.
Genotyping
Addictions Array Genotyping
Genomic DNA was isolated from blood using standard protocols. A genomic region including sequence 5 kb upstream and 1 kb downstream of SLC6A4 was retrieved from NCBI Human Build 35.1. Haplotype tagging SNPs, derived from the African HapMap sample, were identified using a previously described design pipeline (Hodgkinson et al., 2008). Fourteen SLC6A4 SNPs, listed in Table 1, were genotyped using the Illumina GoldenGate platform (Hodgkinson et al., 2008). SNP rs6352 (Lys605Asn) was monomorphic. The remaining 13 SNPs were in Hardy-Weinberg equilibrium.
TABLE 1.
Description of SLC6A4 SNPs
| SNPs | Location | Af Am Base Variation | MAF | |||
|---|---|---|---|---|---|---|
| Chr 17* | Gene | Major > Minor Allele | African Americans | Finnish Caucasians | Plains Indians | |
| rs3813034 | 25548930 | 3'UTR | A > C | 0.24 | 0.44 | 0.73 |
| rs1042173 | 25549137 | 3'UTR | T > G | 0.24 | 0.44 | 0.73 |
| rs2054848 | 25554071 | intron | A > G | 0.07 | 0 | 0 |
| rs6352 | 25554319 | Lys605Asn | A > C | 0 | 0.05 | 0 |
| rs3794808 | 25555919 | intron | G > A | 0.35 | 0.40 | 0.66 |
| rs140701 | 25562658 | intron | G > A | 0.31 | 0.39 | 0.66 |
| rs4583306 | 25562841 | intron | A > G | 0.20 | 0.39 | 0.66 |
| rs2020942 | 25571040 | intron | G > A | 0.28 | 0.45 | 0.23 |
| rs6355 | 25572936 | Gly56Ala | G > C | 0.01 | 0 | 0 |
| rs2066713 | 25575791 | 5'UTR | C > T | 0.30 | 0.45 | 0.23 |
| rs16965628 | 25579551 | 5'UTR | G > C | 0.30 | 0.03 | 0.03 |
| rs2020933 | 25585881 | 5'UTR | T > A | 0.32 | 0.03 | 0.03 |
| rs9903062 | 25586450 | 5'UTR | G > A | 0.04 | 0 | 0 |
| rs25531** | 25588472 | 5'UTR | A > G | 0.23 | 0.10 | 0.01 |
| rs2020930 | 25590167 | intergenic | G > A | 0.03 | 0.03 | 0 |
Build 36.3
All SNPs were genotyped on the addictions array with the exception of rs25531 that is genotyped by size discrimination as part of the triallelic 5-HTTLPR polymorphism Allele frequencies are similar to HapMap freqs (for PI, frequencies approximate to HapMap Asians for all SNPs except for rs2020942 and rs2066713).
MAF = minor allele frequency
Triallelic 5-HTTLPR Genotyping
Genotyping was performed in two stages using size discrimination for the S (103bp) and L (146bp) alleles and for the rs25531 (LA (146bp) and LG (61bp)) alleles. The 5-HTTLPR region was amplified in a 20μl reaction: 1× Optimized Buffer A, 1× PCR enhancer, 0.25μM of each primer [FAM-ATCGCTCCTGCATCCCCCATTAT (forward primer), GAGGTGCAGGGGGATGCTGGAA (reverse primer)], 0.125μM of dNTP, 10ng of DNA, 1.25u of Platinum Taq polymerase (all from Invitrogen Corp). The PCR conditions were: 95°C (5 min), 40 cycles of 94°C (30sec), 52 °C (30sec), 68°C (1 min), and a final elongation, 68°C (10 min). S and L genotypes were discriminated directly from the PCR reaction products, The rs25531 LA and LG genotypes was determined by digesting 10μl PCR mix with 50units of MspI (37°C, for 1 hour, 1× restriction buffer). Samples were mixed with deionized formamide and GeneScan™-500 ROX Size Standard (Applied Biosystems), and the genotypes were resolved on a 3730 DNA Analyzer (Applied Biosystems).
Genotyping accuracy was determined empirically by duplicate genotyping of 25% of the samples selected randomly. The error rate was <0.005, and the completion rate was >0.95.
Triallelic genotyping revealed the following allele frequencies: S = 0.25, LA = 0.51, LG = 0.24. Alleles were grouped as low activity (SS, SLG, LGLG) (0.23) medium activity (SLA, LALG) (0.51) and high activity (LALA) (0.26) variants.
For comparisons of LD across SLC6A4, triallelic 5-HTTLPR genotyping, using the identical method, and the identical addictions array genotyping, was available for samples of Finnish Caucasians and Plains American Indians (see supplementary materials).
Assessment of Population Stratification using Ancestry Informative Markers (AIMS)
The samples were genotyped for 186 ancestry markers (Hodgkinson et al., 2008). The same AIMs were also genotyped in 1051 individuals from the 51 worldwide populations represented in the HGDP-CEPH Human Genome Diversity Cell Line Panel (http://www.cephb.fr/HGDP-CEPH-Panel). Ancestry factor scores were generated for each study subject by running Structure 2.2 (http://pritch.bsd.uchicago.edu/software.html). AIMs genotype data for each subject were analyzed individually along with the data for the 51 CEPH populations to identify population substructure and compute individual ethnic factor scores. This ancestry assessment showed that the European factor score was on average 0.09 (median value = 0.04). Both a Mid East factor and an Asian factor had an average score of 0.06 (median 0.04).
Statistical Analyses
Logistic regression analyses with attempted suicide as the dependent variable were undertaken using JMP 7 software and yielded likelihood ratio (L-R) χ2 results. Backward stepwise regression was performed with predictor variables being eliminated from the model in an iterative process if the level of significance was p > 0.1. The continuous CTQ score and African ethnic factor score and the dichotomous variables: age, sex and substance dependence diagnosis were initially included as predictor variables. Age and sex had no significant effects. Substance dependence and the continuous CTQ score were included since they had a significant effect. 5-HTTLPR genotype (low, medium, high activity) and rs3813034-rs1042173 diplotypes: ATAT and CGCG/ATCG were included as predictor variables. Interaction terms were included in the final model when significant (p < 0.05).
Haplotype frequencies were estimated using a Bayesian approach implemented with PHASE (Stephens & Donnelly, 2003). Haploview version 2.04 Software (Whitehead Institute for Biomedical Research, USA) was used to produce LD matrices.
RESULTS
From Figure 1 it can be seen that in this African American sample there is a semblance of a haplotype block in the distal region of SLC6A4 extending from rs2066713 to rs3813034. The same haplotype block occurs in Caucasians and American Indians but in these ethnic groups there is strong LD across the block (Figures S2 and S3).
FIGURE 1.
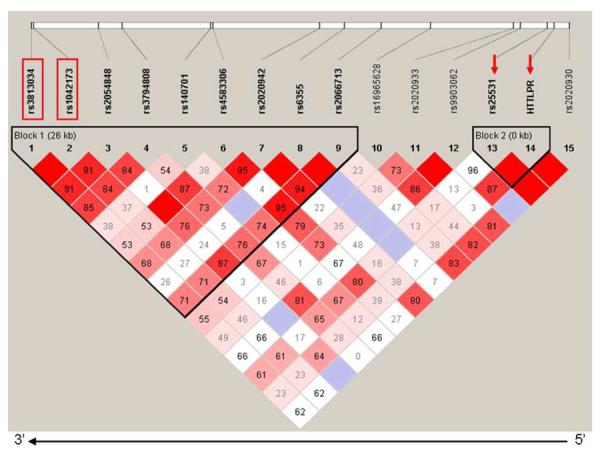
SLC6A4 haplotype block structure in African Americans
The numbers in the squares refer to pairwise linkage disequilibrium measured as D'. Haplotype blocks were defined using a setting of average pairwise D' within-block of ≥ 0.80. The direction of gene transcription is from right to left.
The position of the biallelic 5-HTTLPR polymorphism is indicated by an arrow at the proximal (5') end of the gene. The other arrow indicates rs25531 that is part of the triallelic polymorphism. The two putative functional SNPs at the distal (3') end of the gene are indicated by rectangles.
In the African Americans the two most distal SNPs (rs3813034 and rs1042173) are in strong LD and occur at identical frequencies within each ethnic group (Table 1). Two other SNPs rs4583306 and rs2020942 are in LD with each other. In particular, there is no evidence of LD between rs25531, located within 5-HTTLPR and the functional 3'UTR SNP rs3813034.
The 5-HTTLPR polymorphism was artificially introduced as a `SNP SL' in order to see whether there was LD between it and the distal SNPs rs1042173 and rs3813034. As validation, haplotype analyses between rs25531 A > G and 5-HTTLPR L > S showed the following haplotypes: AS 0.26; AL 0.52; GL 0.23. As expected, the GS haplotype did not occur. This validated the inclusion of `SNP' 5-HTTLPR. Figure 1 shows that there was no LD between the 5-HTTLPR construct and rs1042173 or rs3813034.
Haplotype analyses
Due to the poor LD structure across the 9 SNP distal haplotype block, PHASE estimation resulted in 23 haplotypes with ≥ 0.01 frequency that accounted for 0.83 of the total variance. There were only 4 haplotypes with ≥ 0.05 frequency that accounted for only 0.43 of the variance; the most abundant haplotype had a frequency of only 0.15. Therefore because of the poor LD structure it was not possible to do haplotype analyses within the total block.
Two-SNP haplotype analyses
Haplotypes were derived for the rs3813034 - rs1042173 haplotype block. There were two main haplotypes: AT (frequency = 0.76) and CG (frequency = 0.24). Diplotypes were derived: ATAT (0.59), ATCG (0.35) and CGCG (0.06). Because of low frequency, diplotypes CGCG and ATCG were combined and compared with ATAT.
Independent effects of 5-HTTLPR and rs3813034-rs1042173 diplotypes on suicidal behavior
Table 2 presents the results of the logistic regression model with total CTQ score, any substance dependence diagnosis, the African ethnic factor score, diplotypes and 5-HTTLPR genotype included as independent variables together with the interaction between any addiction and CTQ score and the interaction between genotype and CTQ score. The total model (N = 449) accounted for 15% of the variance in suicidal behavior: L-R χ2 = 74.6, 9 df, p < 0.0001. There was no main effect of genotype however there were independent G × E interactive effects of diplotype (p = 0.011) and 5-HTTLPR (p = 0.017). There was no diplotype × 5-HTTLPR interactive effect (p = 0.15) on suicidal behavior.
TABLE 2.
Independent effects of 5-HTTLPR and rs3813034-rsl042173 diplotypes on suicidal behavior
| Independent variables | L-R χ2 | DF | P value |
|---|---|---|---|
| Total CTQ score | 29.8 | 1 | < 0.0001 |
| Any substance dependence | 15.1 | 1 | 0.0001 |
| Any substance dependence x CTQ | 4.9 | 1 | 0.027 |
| African ancestry | 2.9 | 1 | 0.089 |
| Diplotypes: ATAT vs ATCG / CGCG | 0.5 | 1 | 0.480 |
| Diplotypes x CTQ | 6.5 | 1 | 0.011 |
| 5-HTTLPR high/med/low | 0.3 | 2 | 0.857 |
| 5-HTTLPR x CTQ | 8.2 | 2 | 0.017 |
N = 449, Total model: χ2 = 74.6, 9 df, p < 0.0001, accounting for 15% variance in suicidal behavior. Interaction terms are shown in italics.
'Any substance dependence' includes single or comorbid dependence on alcohol, cocaine or heroin.
When diplotype was excluded from the model there was still a G × E interactive effect for 5-HTTLPR (p = 0.023) and this model accounted for 13.6% of the variance in suicidal behavior. When 5-HTTLPR was excluded from the model there was a trend G × E interactive effect for diplotype (p = 0.054) and this model accounted for 12.7% of the variance in suicidal behavior. As a matter of interest, when 5-HTTLPR was excluded from the model and each of the SNPs replaced the diplotype, there was a significant G × E effect for rs1042173 TT (major homozygote) vs GT/GG, p = 0.040 and a trend effect for rs3813034 AA (major homozygote) vs AC/CC, p = 0.069.
The interactive effects are illustrated in Figure 2. As described previously, a dichotomous total CTQ score was derived solely to illustrate the direction of interaction effects on suicide attempt. In individuals exposed to high childhood trauma, the risk of suicide attempt was 0.52 in carriers of the low activity 5-HTTLPR variant but `only' 0.32 in carriers of the medium and high activity variants. In contrast, 5-HTTLPR genotype had no effect on suicide risk in individuals who had experienced low or no childhood trauma (risk = 0.12 – 0.20). Likewise, in individuals exposed to high childhood trauma, carriers of the major ATAT diplotype had an increased risk of suicidal behavior relative to carriers of the ATCG/CGCG diplotypes (0.40 vs 0.31) whereas genotype had no effect on suicide risk in individuals not exposed to childhood trauma (0.15 – 0.22).
FIGURE 2.
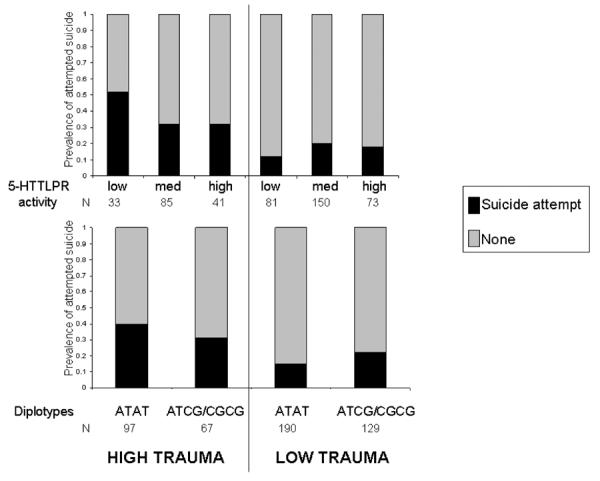
Independent effects of 5-HTTLPR and rs3813034-rs1042173 diplotypes on suicide risk after exposure to high childhood trauma
As can be seen in Figure 3 and Table 4, of the 154 individuals who had been exposed to childhood trauma, the prevalence of suicide attempt was 0.56 in carriers of both risk variants (5-HTTLPR low activity variant and the ATAT major diplotype), 0.39 in carriers of one risk variant and 0.25 in individuals without either risk variant. In the 296 individuals who had not experienced childhood trauma, the lowest prevalence of suicidal behavior was in carriers of the two `risk' variants (0.07) followed by carriers of one `risk' variant (0.17) and carriers of no risk variant (0.24). It should be noted that individuals without either risk variant were resilient to the deleterious effects of childhood trauma.
FIGURE 3.
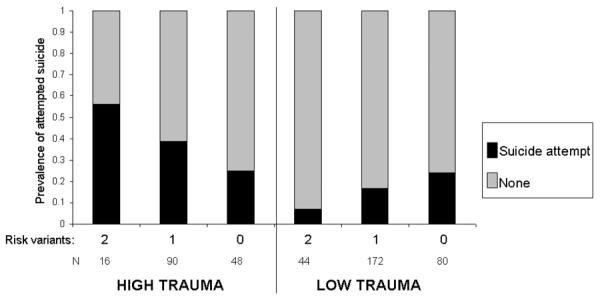
Cumulative effects of SLC6A4 risk variants on suicidal behavior in the context of childhood trauma
Risk variants: 5-HTTLPR low activity; rs3813034-rs1042173 major ATAT diplotype
TABLE 4.
Cumulative effects of SLC6A4 risk variants on suicidal behavior in the context of childhood trauma
| Risk Variants | HIGH TRAUMA | LOW TRAUMA | ||
|---|---|---|---|---|
| A: rs3813034-rsl042173 Diplotype ATAT | Suicide Attempt | Suicide Attempt | ||
| B: 5-HTTLPR low activity | Prevalence | Prevalence | ||
| N | N | |||
| No Risk Variant | 0.25 | 0.24 | ||
| ATCG/CGCG + high | 4/15 | 0.27 | 4/23 | 0.17 |
| ATCG/CGCG + med | 8/33 | 0.24 | 15/57 | 0.26 |
| total N: 48 | total: 80 | |||
| One Risk Variant | 0.39 | 0.17 | ||
| ATAT + high | 9/24 | 0.38 | 9/49 | 0.18 |
| ATAT + med | 18/50 | 0.36 | 14/86 | 0.16 |
| ATCG/CGCG + low | 8/16 | 0.50 | 7/37 | 0.19 |
| total N: 90 | total: 172 | |||
| Two Risk Variants | ||||
| ATAT + low | 9/16 | 0.56 | 3/44 | 0.07 |
rs3813034-rsl042173 diplotypes: ATAT, ATCG/CGCG: risk variant = ATAT 5-HTTLPR variants: low, medium and high activity: risk variant = low activity
Secondary individual SNP analyses
There were no significant results for any of the remaining SNPs.
DISCUSSION
We have found that in this sample of African Americans, suicidal behavior was, as expected, more common in individuals who had been exposed to childhood trauma (Figure 2). The participants in this study had for the most part been exposed to considerable childhood trauma; for example, using the dichotomous CTQ clinical cut-off scores that differentiate between the presence or absence of significant abuse/neglect (Walker et al., 1999) we found that only 27% of individuals did not meet criteria for significant abuse/neglect. In this group of high risk individuals, the prevalence of suicide attempts was highest in carriers of the 5-HTTLPR low activity variant (0.52) and carriers of the major rs3813034-rs1042173 diplotype ATAT (0.40); the highest rate was found in carriers of both variants (0.56) (Figure 3). In African Americans, 5-HTTLPR, and the two tightly linked SNPs rs3813034 and rs1042173, the functional 3' variants, are not in LD and therefore it appears that these variants at either end of the gene have independent and possibly additive effects on suicidal behavior in the context of childhood trauma. Indeed, it has been shown previously that rs1042173 and the triallelic 5-HTTLPR are not in LD in Caucasians and Hispanics ((Seneviratne et al., 2009) and we have also shown this in our samples of Finnish Caucasians and Plains American Indians (Figures S2, S3).
The 5-HTTLPR low activity variant has been associated with greater amygdala neuronal activity in response to fearful stimuli (Hariri et al., 2005) and impaired feedback circuitry for the extinction of negative affect (Pezawas et al., 2005). Moreover, during stress exposure the low activity variant has been associated with increased engagement of neural systems implicated in fear and arousal together with attention towards the emotional salience of threat (Drabant et al., 2012). Thus the low activity variant is associated with increased vulnerability to negative emotions on stress exposure. Since the promoter region of the 5-HTT gene includes a GRE, this gene is likely to be responsive to stress-induced changes in hypothalamic pituitary adrenal (HPA) axis activity that is associated with psychopathology including suicidal behavior. Indeed, Caspi et al., 2003, was one of the first to demonstrate that an interactive effect of the low activity 5-HTTLPR variant and stressful life events predicted depression and suicidal ideation/behavior in early adulthood. Other studies have found similar results but with childhood trauma as the stressor; for example Gibb et al (2006). As in our earlier study in this dataset of substance dependent African Americans (Roy et al., 2007) we found that CTQ-assessed childhood trauma interacted with the low activity 5-HTTLPR variant to increase the risk of a suicide attempt. It should be noted that the Roy et al., 2007 study was conducted in a subset of 257 male substance dependent patients (81 suicide attempters) using the 34 item CTQ. The current study includes all 474 participants (112 suicide attempters) in the dataset who have been administered the CTQ i.e. this is a larger dataset that also includes women and controls. Moreover, the current study is an update on the earlier study since AIMS have been included to account for population stratification and the 34 item CTQ has been converted to the 28 item CTQ because this is the most recent and commonly used form of the questionnaire.
As previously mentioned, our current study has also shown that functional variants in the 3' end of SLC6A4 influence suicidal behavior in the context of childhood trauma. The 3' untranslated region (UTR) of SLC6A4 mRNA exists in two forms that differ by a 123 bp element (Battersby et al., 1999). The distal polyadenylation signal includes rs3813034 that has been shown to alter the balance of the two mRNA transcripts in the brain and lymphoblasts (Gyawali et al., 2010). One group has found that the major A allele is associated with a higher distal polyadenylation fraction in a dose-dependent manner i.e. more efficient polyadenylation than the minor C allele and putatively increased 5-HTT expression (Gyawali et al., 2010). This group found that the rs3813034 minor C allele was associated with panic disorder, anxiety / depressive symptoms and impaired retention of fear extinction (Gyawali et al., 2010; Hartley et al., 2012). Moreover, since chronic administration of fluoxetine increased the 5-HTT distal polyadenylation fraction in mouse brain, it is possible that the therapeutic effects of SSRIs may in part be due to changes in 5-HTT polyadenylation (Hartley et al., 2012). In contrast, another research group has produced opposite results. Seneviratne et al., 2009 have shown that in HeLa cell cultures, rs1042173 minor G allele carriers have higher 5-HTT mRNA and protein levels. This group has shown that alcoholics who are carriers of the rs1042173 major TT homozygote are more intense drinkers amongst heavy drinking alcoholics and have increased craving when exposed to alcohol cues (Ait-Daioud et al., 2012; Johnson et al., 2011; Seneviratne et al., 2009). Our study has, in the first instance, shown a relationship between a stress-related disorder, suicidal behavior, and the 5-HTTLPR variant that results in lower levels of 5-HTT. The fact that we found a similar suicidal behavior association with the distal major ATAT diplotype and that the effects might be additive with the 5-HTTLPR variant, suggests that our findings tie in with that of Seneviratne et al., 2009 rather than that of Gyawali et al., 2010.
Finally, from Table 3 it can be seen that although the frequencies of the 5' and 3' risk variants are similar in Caucasians, the frequencies vary in other ethnicities such that the 5-HTTLPR low activity variant is the predominant risk factor in African Americans and the ATAT diplotype is the predominant risk factor in American Indians. Thus variation in the combinations of 5' and 3' functional variants may have pharmacogenetic implications for treatment with SSRIs.
TABLE 3.
Frequencies of SLC6A4 risk variants in different populations
| rs3813034-rsl042173 AT AT diplotype | 5-HTTLPR low activity variant | |
|---|---|---|
| African Americans | 0.59 | 0.23 |
| Finnish Caucasians | 0.32 | 0.26 |
| Plains Indians | 0.08 | 0.43 |
There are a few limitations to this study. Although the sample size decreased to 474 (112 suicide attempters) when examining G × E interactions, as mentioned earlier it is important to note that the participants in this study had for the most part been exposed to considerable childhood trauma and this could increase the strength of G × E interactions. Our finding that there was no statistically significant interactive effect between the 5' and 3' variants on suicidal behavior (p = 0.15) might be due to small sample size. As can be seen from Figure 3, although 60 individuals were carriers of both 5' and 3' risk variants, only 16 of these had experienced high childhood trauma. The CTQ is a self-report questionnaire although it has been shown to have high reliability and validity (reviewed in Roy et al., 2012). The nature and severity of each suicide attempt was not systematically recorded. Since the complete SCID was not administered, data about possible comorbid disorders such as depression, anxiety disorders and PTSD was not available. Finally this study was performed in African Americans predominantly with substance dependence and it is not clear that the results can be generalized to other populations or other patient groups. The results of this study should be treated with caution until replicated.
In conclusion, our results indicate that, at least in African American men with substance dependence, unlinked functional variants at both the 5' and 3' ends of SLC6A4 appear to influence a stress-related disorder, suicidal behavior, in individuals who have increased vulnerability because of exposure to childhood trauma. On the other hand, the risk - resilience balance is altered in individuals without either the 5' or the 3' risk variant as they are resilient to the adverse effects of childhood trauma.
Supplementary Material
ACKNOWLEDGEMENTS
There are no acknowledgements for this paper.
FUNDING SUPPORT This research was supported by the Intramural Research Program of the National Institute on Alcohol Abuse and Alcoholism, NIH and in part by grant RO1 DA 10336-02 to AR from the National Institute of Drug Abuse, NIH.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
DISCLOSURE /CONFLICT OF INTERES None of the authors have any disclosures or conflicts of interest to declare.
CONTRIBUTORS Dr Enoch designed the study, carried out the data analyses and wrote the paper.
Dr Hodgkinson was involved in the genotyping, contributed to the genetic analyses and participated in writing the paper.
Dr Gorodetsky performed the 5-HTTLPR genotyping, helped with constructing the figures and participated in writing the paper.
Dr Goldman supervised the genetic analyses.
Dr Roy wrote the protocol, collected the patients and participated in the writing of the paper.
All authors contributed to and have approved the final manuscript.
REFERENCES
- Ait-Daoud N, Seneviratne C, Smith JB, Roache JD, Dawes MA, Liu L, Wang XQ, Johnson BA. Preliminary Evidence for cue-induced Alcohol Craving Modulated by Serotonin Transporter Gene Polymorphism rs1042173. Frontiers in Psychiatry. 2012;3:6. doi: 10.3389/fpsyt.2012.00006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asberg M, Träskman L, Thorén P. 5-HIAA in the cerebrospinal fluid. A biochemical suicide predictor? Archives of General Psychiatry. 1976;33:1193–1197. doi: 10.1001/archpsyc.1976.01770100055005. [DOI] [PubMed] [Google Scholar]
- Battersby S, Ogilvie AD, Blackwood DH, Shen S, Muqit MM, Muir WJ, Teague P, Goodwin GM, Harmar AJ. Presence of multiple functional polyadenylation signals and a single nucleotide polymorphism in the 3' untranslated region of the human serotonin transporter gene. Journal of Neurochemistry. 1999;72:1384–1388. doi: 10.1046/j.1471-4159.1999.721384.x. [DOI] [PubMed] [Google Scholar]
- Bernstein D, Fink L. Childhood Trauma Questionnaire: A retrospective self-report manual. The Psychological Corporation; San Antonio, TX: 1998. [Google Scholar]
- Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H, McClay J, Mill J, Martin J, Braithwaite A, Poulton R. Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science. 2003;301:386–389. doi: 10.1126/science.1083968. [DOI] [PubMed] [Google Scholar]
- Caspi A, Hariri AR, Holmes A, Uher R, Moffitt TE. Genetic sensitivity to the environment: the case of the serotonin transporter gene and its implications for studying complex diseases and traits. American journal of Psychiatry. 2010;167:509–527. doi: 10.1176/appi.ajp.2010.09101452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drabant EM, Ramel W, Edge MD, Hyde LW, Kuo JR, Goldin PR, Hariri AR, Gross JJ. Neural mechanisms underlying 5-HTTLPR-related sensitivity to acute stress. American Journal of Psychiatry. 2012;169:397–405. doi: 10.1176/appi.ajp.2011.10111699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibb BE, McGeary JE, Beevers CG, Miller IW. Serotonin transporter (5-HTTLPR) genotype, childhood abuse, and suicide attempts in adult psychiatric inpatients. Suicide and Life Threatening Behavior. 2006;36:687–693. doi: 10.1521/suli.2006.36.6.687. [DOI] [PubMed] [Google Scholar]
- Glatz K, Mössner R, Heils A, Lesch KP. Glucocorticoid-regulated human serotonin transporter (5-HTT) expression is modulated by the 5-HTT gene-promotor-linked polymorphic region. Journal of Neurochemistry. 2003;86:1072–1078. doi: 10.1046/j.1471-4159.2003.01944.x. [DOI] [PubMed] [Google Scholar]
- Gyawali S, Subaran R, Weissman MM, Hershkowitz D, McKenna MC, Talati A, Fyer AJ, Wickramaratne P, Adams PB, Hodge SE, Schmidt CJ, Bannon MJ, Glatt CE. Association of a polyadenylation polymorphism in the serotonin transporter and panic disorder. Biological Psychiatry. 2010;67:331–338. doi: 10.1016/j.biopsych.2009.10.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hariri AR, Drabant EM, Munoz KE, Kolachana BS, Mattay VS, Egan MF, Weinberger DR. A susceptibility gene for affective disorders and the response of the human amygdala. Archives of General Psychiatry. 2005;62:146–152. doi: 10.1001/archpsyc.62.2.146. [DOI] [PubMed] [Google Scholar]
- Hariri AR, Holmes A. Genetics of emotional regulation: the role of the serotonin transporter in neural function. Trends in Cognitive Sciences. 2006;10:182–191. doi: 10.1016/j.tics.2006.02.011. [DOI] [PubMed] [Google Scholar]
- Hartley CA, McKenna MC, Salman R, Holmes A, Casey BJ, Phelps EA, Glatt CE. Serotonin transporter polyadenylation polymorphism modulates the retention of fear extinction memory. Proceedings of the National Academy of the Sciences USA. 2012;109:5493–5498. doi: 10.1073/pnas.1202044109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heils A, Teufel A, Petri S, Stöber G, Riederer P, Bengel D, Lesch KP. Allelic variation of human serotonin transporter gene expression. Journal of Neurochemistry. 1996;66:2621–2624. doi: 10.1046/j.1471-4159.1996.66062621.x. [DOI] [PubMed] [Google Scholar]
- Heinz A, Braus DF, Smolka MN, Wrase J, Puls I, Hermann D, Klein S, Grusser SM, Flor H, Schumann G, Mann K, Buchel C. Amygdala-prefrontal coupling depends on a genetic variation of the serotonin transporter. Nature Neuroscience. 2005;8:20–21. doi: 10.1038/nn1366. [DOI] [PubMed] [Google Scholar]
- Hodgkinson CA, Yuan Q, Xu K, Shen PH, Heinz E, Lobos EA, Binder EB, Cubells J, Ehlers CL, Gelernter J, Mann J, Riley B, Roy A, Tabakoff B, Todd RD, Zhou Z, Goldman D. Addictions biology: haplotype-based analysis for 130 candidate genes on a single array. Alcohol Alcohol. 2008;43:505–515. doi: 10.1093/alcalc/agn032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu X-Z, Lipsky RH, Zhu G, Akhtar LA, Taubman J, Greenberg BD, Xu K, Arnold PD, Richter MA, Kennedy JL, Murphy DL, Goldman D. Serotonin transporter promoter gain-of-function genotypes are linked to obsessive-compulsive disorder. American Journal of Human Genetics. 2006;78:815–826. doi: 10.1086/503850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson BA, Ait-Daoud N, Seneviratne C, Roache JD, Javors MA, Wang XQ, Liu L, Penberthy JK, DiClemente CC, Li MD. Pharmacogenetic approach at the serotonin transporter gene as a method of reducing the severity of alcohol drinking. American Journal of Psychiatry. 2011;168:265–275. doi: 10.1176/appi.ajp.2010.10050755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pezawas L, Meyer-Lindenberg A, Drabant EM, Verchinski BA, Munoz KE, Kolachana BS, Egan MF, Mattay VS, Hariri AR, Weinberger DR. 5-HTTLPR polymorphism impacts human cingulate-amygdala interactions: a genetic susceptibility mechanism for depression. Nature Neuroscience. 2005;8:828–834. doi: 10.1038/nn1463. [DOI] [PubMed] [Google Scholar]
- Roy A, Hu XZ, Janal MN, Goldman D. Interaction between childhood trauma and serotonin transporter gene variation in suicide. Neuropsychopharmacology. 2007;32:2046–2052. doi: 10.1038/sj.npp.1301331. [DOI] [PubMed] [Google Scholar]
- Roy A, Sarchiopone M, Carli V. Gene-environment interaction and suicidal behavior. Journal of Psychiatric Practice. 2009;15:282–288. doi: 10.1097/01.pra.0000358314.88931.b5. [DOI] [PubMed] [Google Scholar]
- Roy A, Gorodetsky E, Yuan Q, Goldman D, Enoch MA. Interaction of FKBP5, a stress-related gene, with childhood trauma increases the risk for attempting suicide. Neuropsychopharmacology. 2010;35:1674–1683. doi: 10.1038/npp.2009.236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy A, Hodgkinson CA, Deluca V, Goldman D, Enoch MA. Two HPA axis genes, CRHBP and FKBP5, interact with childhood trauma to increase the risk for suicidal behavior. Journal of Psychiatric Research. 2012;46:72–79. doi: 10.1016/j.jpsychires.2011.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy M. Diabetic retinopathy in African Americans with type 1 diabetes: The New Jersey 725. 1. Methodology, population, frequency of retinopathy, and visual impairment. Archives of Ophthalmology. 2000;118:97–104. doi: 10.1001/archopht.118.1.97. [DOI] [PubMed] [Google Scholar]
- Seneviratne C, Huang W, Ait-Daoud N, Li MD, Johnson BA. Characterization of a functional polymorphism in the 3' UTR of SLC6A4 and its association with drinking intensity. Alcoholism: Clinical and Experimental Research. 2009;33:332–339. doi: 10.1111/j.1530-0277.2008.00837.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spitzer RL, Williams JBW, Gibbon M. Structured Clinical Interview for DSM-IV (SCID) New York State Psychiatric Institute, Biometrics Research; New York: 1995. [Google Scholar]
- Stephens M, Donnelly P. A comparison of Bayesian methods for haplotype reconstruction from population genotype data. American Journal of Human Genetics. 2003;73:1162–1169. doi: 10.1086/379378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker EA, Unutzer J, Rutter C, Gelfand A, Saunders K, VonKorff, Koss MP, Katon W. Cost of health care use by women HMO members with a history of childhood abuse and neglect. Archives of General Psychiatry. 1999;56:609–613. doi: 10.1001/archpsyc.56.7.609. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.