

Abstract
Background
Prenatal tobacco smoke exposure is associated with alterations in motivated behavior in offspring, such as increased consumption of highly palatable foods and abused drugs. Animal models show that gestational nicotine (GN) exposure mediates changes in responding for sucrose and drug reward.
Methods
A novel, intermittent low-dose intravenous (IV) exposure model was used to administer nicotine (0.05 mg/kg/injection) or saline 3×/day to rats on gestational days 8-21. Two experiments investigated the effect of IV GN on 1) the habituation of spontaneous locomotor activity and on 2) sucrose reinforced responding in offspring. For the operant experiments, animals acquired fixed-ratio (FR-3) responding for sucrose, 26% (w/v), and were tested on varying concentrations (0, 3, 10, 30, 56%; Latin-square) according to a FR-3, and then a progressive-ratio (PR) schedule. Male and female adult offspring were used.
Results
IV GN did not alter birth or growth weight, or the number of pups born. No between-group differences in habituation to spontaneous locomotor activity were observed. FR testing produced an inverted U-shaped response curve, and rats showed peak responding for 10% sucrose reinforcement. Neither gestation nor sex affected responding, suggesting equivalent sensitivity to varying sucrose concentrations. PR testing revealed that GN rats showed greater motivation for sucrose reinforcement relative to controls.
Conclusions
A low-dose, IV GN exposure model resulted in increased motivation to respond for sucrose reinforcement in adult offspring. This suggests that using a low number of cigarettes throughout pregnancy will result in increased motivation for highly palatable foods in adult, and perhaps, adolescent offspring.
Keywords: gestational nicotine, locomotor activity, sucrose-maintained responding, sex, rats
1. Introduction
In the United States, approximately 13% of women continue to use tobacco cigarettes throughout pregnancy (Martin et al., 2008; Tong et al., 2009). Prenatal tobacco smoke exposure results in numerous health risks for the infant such as intrauterine growth retardation, reduced birth weight, and sudden infant death syndrome (see Pauly and Slotkin, 2008; Salihu and Wilson, 2007). Moreover, it is associated with neurodevelopmental deficits in offspring like attention deficit hyperactivity disorder (Button et al., 2007; Button et al., 2005; Linnet et al., 2003; Thapar et al., 2003) and alterations in auditory and cognitive processing (Fried et al., 1998, 2003; Jacobsen et al., 2007; Jacobsen et al., 2006; McCartney et al., 1994; Picone et al., 1982; Saxton, 1978; Sexton et al., 1990). Of particular interest are the tobacco smoke-induced neurodevelopmental alterations in motivated behavior manifested during adolescent and adult development. Individuals exposed to prenatal tobacco smoke are more likely to exhibit increased abuse of recreational drugs and increased consumption of highly palatable foods relative to non-exposed individuals (Al Mamun et al., 2006a; Al Mamun et al., 2006b; Buka et al., 2003; Kandel et al., 1994; Toschke et al., 2003; Weissman et al., 1999).
Nicotine is the primary psychoactive compound in tobacco that maintains smoking behavior (Benowitz et al., 2009; Corrigall and Coen, 1989; Corrigall et al., 1992). Several preclinical animal models are being used to investigate the neurobehavioral effects of gestational nicotine (GN) exposure (see Dwyer et al., 2009). The most common method is to administer continuous nicotine via osmotic mini-pump to maintain steady-state nicotine levels throughout gestation (Franke et al., 2008; Slotkin et al., 2006; Wei et al., 2011). Pregnant dams are also administered nicotine through drinking water (Pauly et al., 2004; Paz et al., 2007; Zhu et al., 1996), and by IV injection (Harrod et al., 2011; Lacy et al., 2011; LeSage et al., 2006; Pentel et al., 2006). These models provide converging evidence that GN exposure alone produces neurochemical and behavioral changes in offspring (Dwyer et al., 2008).
Preclinical animal studies confirm that offspring exposed to continuous GN exhibit neurodevelopmental alterations in the mesocorticolimbic dopamine (DA) system (Kane et al., 2004; Muneoka et al., 1997; Navarro et al., 1989; Park et al., 2006; Ribary and Lichtensteiger, 1989; Richardson and Tizabi, 1994; Wei et al., 2011), which organizes goal-directed behavior (Edwards and Koob, 2010; Everitt et al., 2008; Kalivas, 2009; Robinson and Berridge, 2003; Wise and Bozarth, 1987). Moreover, GN-exposed offspring exhibit hyper- and hypoactive spontaneous locomotor activity (LeSage et al., 2006; Pauly et al., 2004; Paz et al., 2007; Peters and Tang, 1982; Peters et al., 1979; Richardson and Tizabi, 1994; Romero and Chen, 2004; Tizabi et al., 2000; Vaglenova et al., 2004), and alterations in cocaine and nicotine self-administration (Franke et al., 2008; Levin et al., 2006).
Interest is growing in the association between gestational tobacco smoke exposure and the increased consumption of palatable foods by offspring (Al Mamun et al., 2006a; Franke et al., 2008). For example, individuals exposed to gestational tobacco smoke show greater preference for soft drinks relative to those not prenatally exposed. Further, prenatal tobacco smoke-exposed individuals are reported to have a greater body mass index and increased odds of being obese during postnatal development (Al Mamun et al., 2006a; Toschke et al., 2003). Experiments using the continuous GN exposure model reveal that rat offspring show decreased motivation for sucrose reinforcement, relative to saline treated controls. This research thus indicates that GN exposure alone influences motivation for sucrose-based reward in offspring (Franke et al., 2008).
The present experiments tested the effect of a novel low-dose, intermittent prenatal IV nicotine exposure model on 1) spontaneous locomotor activity and 2) sucrose-maintained operant responding in adult rat offspring. The use of the IV route of administration to deliver prenatal nicotine is of interest because this route closely mimics the absorption and distribution of nicotine through smoke inhalation (Benowitz et al., 2009; Booze et al., 1999; Mactutus, 1989; Russell and Feyerabend, 1978). Previous research using a high-dose IV exposure method to deliver multiple nicotine boli per day throughout gestation (e.g., every 14 min for 16 h/day) resulted in hypoactive spontaneous locomotor behavior in preweanling-aged offspring (LeSage et al., 2006). The effects of low-dose, IV nicotine exposure have not been investigated with spontaneous locomotor activity or sucrose-maintained reinforcement. It was hypothesized that low-dose IV prenatal nicotine exposure would alter spontaneous locomotor behavior and the motivation to respond for sucrose reward in rats.
2. Methods
2.1 Animals
A total of 40 female and 15 male adult, virgin Sprague-Dawley rats were acquired from Harlan Industries, Inc. (Indianapolis, IN). All rats were transported to the animal care facilities in the psychology department at the University of South Carolina and rodent food (ProLab Rat/Mouse/Hamster Chow 3000) and water were provided ad libitum throughout the experiments, except when otherwise specified. The animal colony was maintained at 21 ± 2° C, 50% ± 10% relative humidity and a 12L: 12D cycle with lights on at 0700 h. The protocol for this research methodology was approved by the Institutional Animal Care and Use Committee (IACUC) at the University of South Carolina.
2.2 Surgery
The internalized jugular catheters used in the present experiment are commercially available from Harlan Industries. The catheterization was performed at Harlan Industries (Indianapolis, IN), prior to arrival at the University of South Carolina, according the methods of Mactutus et al. (1994). Briefly, animals were anesthetized with a mixture of ketamine hydrochloride (100 mg/kg/ml) and xylazine (3.3 mg/kg/ml). Following anesthesia a sterile Intracath IV catheter (Becton, Dickinson and Co., Franklin Lakes, NJ) with a Luer-Lok injection cap (Medex, Inc., Carlsbad, CA) was implanted dorsally in a subcutaneous pouch. The distal end of the catheter was inserted into the left jugular vein, advanced toward the heart, and the catheter was bound with sterile suture. Animals were kept under periodic post-operative observation and returned to the colony upon recovery. On the day following surgery, catheters were flushed with 0.2 ml of heparinized saline.
2.3 Breeding
Following a seven-day habituation period, female rats were housed three per cage, and one male was placed with each triad from approximately 1700 to 0900. Lavage samples were taken daily and analyzed with a microscope to assess the result of the overnight breeding. When a sample was confirmed as sperm-positive the corresponding female was single-caged and that day was considered gestational day (GD) 0. The weights of the pregnant dams were recorded daily during pregnancy.
2.4 Prenatal Nicotine Treatment
Pregnant dams were randomly assigned to either the GN or GS groups. Nicotine (0.05 mg/kg/injection) or saline was administered 3×/day via internalized IV catheters from gestational days 8-21. The dose of IV nicotine was chosen based on previous pharmacokinetic and behavioral data. The elimination half-life for IV 0.05 mg/kg/injection nicotine is ~ 50 minutes (Booze et al., 1999), and thus, this model is unique in that the dam and fetuses experience the bolus delivery of nicotine to the brain followed by a rather precipitous clearance (LeSage et al., 2006; Mactutus, 1989; Russell and Feyerabend, 1978). The 3× daily injection of this dose, together with an inter-injection interval of 3 hours allows for substantial (2-3 half-lives), but not “complete” (6 half-lives) elimination of nicotine. Due to the increased volume of distribution associated with advancing pregnancy some accumulation of nicotine levels that also occurs with cigarette smoking is expected following repeated injections. Behaviorally, repeated IV injection of this dose produces behavioral sensitization (Booze et al., 1999; Harrod et al., 2004), and changes DA transporter and DA receptor expression in adult rats, demonstrating that this dose and route of administration alters motivational circuitry in rodents. Moreover, the 0.05 mg/kg/injection dose is within the dose range that supports nicotine self-administration in rats (Chaudhri et al., 2005).
Intravenous injections were delivered through the Luer-Lok injection cap of the subcutaneously implanted injection port (20 seconds in duration). Following the first and second injections catheters were flushed with 0.2 ml of 0.9% physiological saline because 0.2 ml represents the approximate volume of the catheter. Injection of 0.2 ml of heparinized saline was used to flush the catheter and to maintain catheter patency after the final daily nicotine or saline injection. All post-flush injections were 20 seconds in duration. All injections were performed during the light portion of the photoperiod, and injections were administered daily at approximately 1000, 1300, and 1600.
2.5 Drugs
Nicotine hydrogen tartrate (Sigma, St. Louis, MO) was weighed as base and was dissolved in physiological saline (0.9%; Hospira, Inc. Lake Forest, IL). The pH of the nicotine solution was neutralized to ~ 7.0 with NaOH. Control dams were administered saline at the volumetric equivalent of nicotine. Heparin (APP Pharmaceuticals, Schaumburg, IL; 1000 U) was added to saline and the 2.5% heparinized saline solution was used to flush the IV catheters once per day.
2.6 Litter composition, surrogate fostering, and postnatal testing
The day of birth was considered postnatal day (PND) 0. On PND 1 litters were culled to 10 with 5 males and 5 females whenever possible. All pups were surrogate-fostered to timed-pregnant, drug naïve dams to prevent poor maternal care (Vorhees, 1986). The developmental milestones of the righting reflex, negative geotaxis, and eye opening were assessed on PND 3-5, 8-10, and 13-17, respectively. For each experiment, only one male and one female randomly selected from each litter were assigned to each treatment group (Holson and Pearce, 1992). The righting reflex was assessed by placing animals on to their backs. The time from release to the completion of the righting response was recorded. This procedure occurred in blocks of 3 consecutive trials during PND 3-5, the maximum latency for each animal to right itself was 25 sec per trial. Negative geotaxis was measured by placing animals on a wire mesh grid, positioned in a 25° downward angle. Rats were placed with their heads facing towards the downward slope of the apparatus. The latency for animals to turn 180° to face up the slope was recorded for every trial. A maximum latency of 30 sec was allowed for these trials. Animals were tested in three trial blocks across three consecutive days (PND 8-10) for negative geotaxis. Each animal’s eyes (left and right) were checked for degree of openness. The degree of openness was rated on a scale of 0-3: 0 = completely closed; 1 = any part of the eye visible; 2 = partially open; 3 = fully open. All animals’ weights were recorded on PND 1, 7, 14, and 21. Rats were weaned and pair housed, same sex, on PND 21.
2.7 Experiment 1: Effect of prenatal IV nicotine exposure on spontaneous locomotor activity
2.7.1 Apparatus
Locomotor activity was assessed with 16 square (40 × 40 cm) chambers (Kinder Inc., Poway, CA). Movement was detected by infrared photocell interruptions; each chamber had 32 emitter/detector pairs capable of measuring horizontal and vertical activity. The chambers were converted to round compartments (~ 40 cm) by adding Plexiglas inserts. The addition of the Plexiglas required each emitter/detector pair to be specifically tuned by the manufacturer for the extra perspex width. The chambers were housed in an isolated room and controlled by MotorMonitor software by Hamilton-Kinder (Kinder Inc., Poway, CA) on a Pentium class computer running Windows® XP.
2.7.2 Procedure
Subjects were 32 adult, male and female offspring of IV GN or gestational saline (GS; representing 18 litters). Subjects were divided into four groups: the GS-exposed males (GS-M; n=8) and females (GS-F; n=7), and the GN-exposed males (GN-M; n=9) and females (GN-F; n=8). Locomotor activity was assessed across three consecutive days. The range of ages tested across habituation days 1, 2, and 3 correspond to PND 95-106, 96-107, 97-108, respectively. At the beginning of each test, rats were placed in the rear half of the chamber with their head facing the back wall of the apparatus. Males and females from each gestational treatment were represented during each locomotor session. Activity tests were 60 minutes in duration.
2.8 Experiment 2: Effect of prenatal IV nicotine exposure on operant responding for sucrose reinforcement
2.8.1 Apparatus
Operant chambers (ENV-008; Med-Associates, St. Albans, VT), housed within sound-attenuating enclosures, were controlled by Med-PC computer interface software. The front and back panels of the chamber were stainless steel and the sides and top were constructed of polycarbonate. The front panel of the chamber allowed access to a recessed food dipper (ENV-202M) through a 5 × 5 cm opening. Two retractable metal levers (ENV-112BM) on either side of the opening were located 7.3 cm above a metal grid floor. A dipper equipped with a 0.1 ml cup attached to the end of the arm was raised into the food receptacle, which allowed access to sucrose or H2O upon completion of the response requirement. An infrared sensor (ENV-254-CB) was used to detect head entries into the food receptacle.
2.8.2 Preliminary Training
Thirty male and female, adult offspring representing 15 litters were used as subjects. Rats were maintained at 85% body weight for three days prior to the beginning of dipper training. Dipper training and autoshaping were conducted according to the methods of Reichel et al. (2008). Sucrose, 26%; w/v, was used as the reinforcer.
2.8.3 Fixed-ratio training
Both levers were presented in the chamber, and rats learned to respond for continuous reinforcement during 30 min sessions. A response on the active lever resulted in 4 seconds of access to sucrose, whereas responding on the inactive lever was recorded, but not reinforced. After the first day on continuous reinforcement, food was returned to the animal’s home cage allowing ad libitum access for the remainder of the experiment. Animals were maintained on the FR-1 schedule for a minimum of 5 days, and once stable responding was exhibited animals were shifted to an FR-3 schedule of reinforcement. Stable responding was operationally defined as less than 20% variability across 2 consecutive sessions, greater than 2:1 ratio of active to inactive responses, and a minimum of 10 reinforcements per session.
2.8.4 Fixed-ratio responding for the 0, 3, 10, 30, 56 % sucrose concentrations
Following 2 consecutive days of stable responding on the FR-3 schedule animals were allowed to lever press for various concentrations of sucrose according to the same schedule. The sucrose concentrations were 0, 3, 10, 30, and 56%, and were presented to each rat according to a Latin-square design. The maintenance trials, which made 26% sucrose available, occurred every other day (i.e., between each test). After all testing concentrations were assessed, animals reestablished FR-3 responding on the 26% training concentration for 2 days before responding for sucrose according to a progressive-ratio (PR) schedule of reinforcement.
2.8.5 Progressive-ratio responding for the 0, 3, 10, 30, 56 % sucrose concentrations
PR testing occurred in a similar manner as the fixed-ratio tests. Testing concentrations of 0, 3, 10, 30, and 56 % were assessed using the PR schedule with maintenance sessions (i.e., FR-3; 26%) occurring between each PR test. The PR tests were a maximum of 2 hours in length. The sequence of progressive-ratio requirements was: 1, 2, 4, 6, 9, 12, 15, 20, 25, 32, 40, 50, 62, 77, 95, 118, 145, 179, 603, 737, 901, 1102, 1347, 1646, 2012, according to the methods of Richardson and Roberts (1996).
2.9 Data analysis
2.9.1 Litter parameters
The between-subjects factors for the litter parameter analyses were Sex and gestational nicotine or saline treatment (Gestation). The within-subjects factors were PND and GD. A one-way analysis of variance (ANOVA) was conducted for the total number of pups born to GN and GS dams. A Sex × Gestation factorial ANOVA was used to analyze the ratio of males to females born to GN and GS dams. A Sex × Gestation × PND mixed-factorial ANOVA was conducted for the pup weight gain, righting reflex, negative geotaxis and eye opening data. A Gestation × GD mixed-factorial ANOVA determined if there were differences between GS and GN dams on the measure of maternal weight gain.
2.9.2 Locomotor activity
The data were analyzed using a 2 × 2 × 3 × 12 mixed-factorial ANOVA. The between-subjects measures were Sex and Gestation and the within-subjects variables were Day and Time. The dependent measures were total horizontal activity and rearing, which were all measured at 12, 5-minute intervals per session for 3 consecutive days.
2.9.3 Sucrose maintained responding on FR and PR schedules of reinforcement
The dependent measure was active lever responding. The data were analyzed using a 2 × 2 × 5 mixed-factorial ANOVA. For FR and PR data the between-subjects measures were Sex and Gestation and the within-subjects measure was Sucrose Concentration (0, 3, 10, 30, and 56%). Breakpoint data from the PR experiment were analyzed, as well. Fisher’s exact tests were used to assess the rate that GN and GS rats acquired operant responding on the FR-1 and FR-3 schedules (Siegel and Castellan, 1988). For all analyses an alpha level of 0.05 was used to determine statistical significance. Also, Greenhouse-Geisser corrections were used to correct for violations of sphericity (Maulchy’s W test) and this is notated with the subscript “G-G”.
3. Results
3.1 Litter Parameters
No significant differences between GN- and GS-exposed rats were observed for the total number of pups born to dams, the number of male vs. female pups, the righting reflex, negative geotaxis, or eye opening. There was no effect of gestational treatment on maternal weight gain across GD 1, 7, 14, 21 (Figure 1A), or in pup weight gain across PND 1, 7, 14, 21 (Figure 1B). Analysis of pups’ absolute weights revealed a significant effect of Sex [F(1, 159) = 6.98, p<0.01], which represents the standard sex difference in rodent weight. No other main effects or interactions were observed. Together, these analyses indicate that gestational IV nicotine treatment did not differentially impact pup weights.
Figure 1.
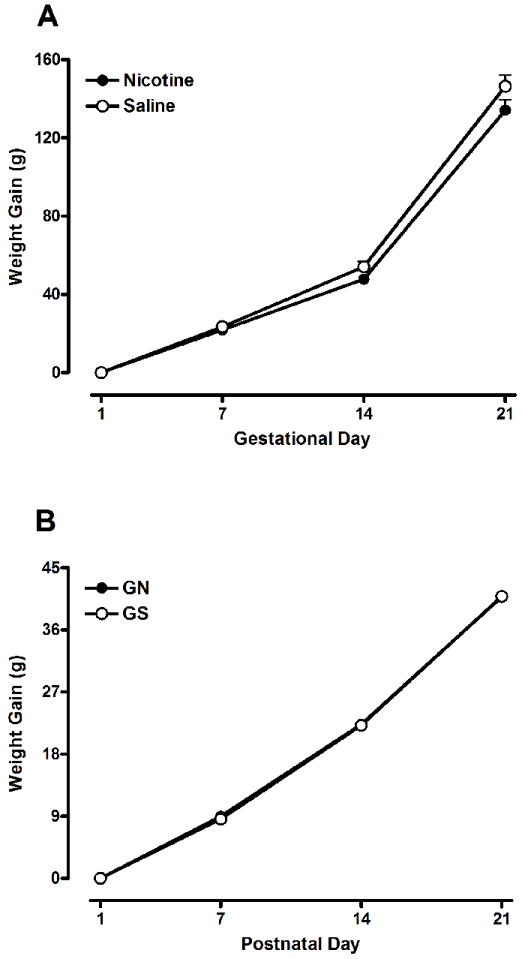
(A) Mean (±SEM) weight gain exhibited by the dams during gestational days 1-21. Dams were treated with IV nicotine (0.05 mg/kg/injection) or saline on gestational days 8-21. There were no differences between the weights of the nicotine and saline females. (B) Mean (±SEM) weight gain of the pups across postnatal days 1-21. The offspring were prenatally exposed to IV GS or GN. There were no significant differences between the GN and GS groups.
3.2 Experiment 1: Locomotor Activity
3.2.1 Total Horizontal Activity
Figure 2 shows the spontaneous locomotor data presented across the three habituation days as a function of Time and Gestation. A mixed-design ANOVA did reveal main effects of Day [F(2, 56) = 15.041, p<0.001] and Time [F(11, 308) = 422.825, p<0.001], and a significant Day × Time interaction [F(22, 616) = 1.985, p<0.05]. Rats exhibited the most activity during the first 5-min interval following placement into the activity chamber and this did not change across testing days. Rats did exhibit habituation but this was indicated by a progressive decrease in horizontal activity on minutes 10-25 across testing days, with relatively lower levels of activity during the remainder of the hour (see Figure 2). The Day × Time interaction confirms this downward and leftward shift in mean horizontal activity as a function of day. Gestation and Sex were not significant factors in this analysis.
Figure 2.
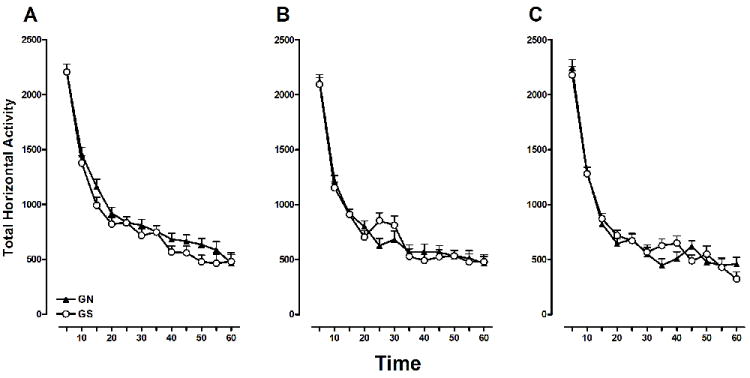
Mean (±SEM) total horizontal activity counts in GN and GS rats, over the 60-minute activity session, for habituation days 1, 2, and 3 (A, B, and C, respectively). These testing days correspond to PND 95-106, 96-107, 97-108, respectively. A significant Day × Time interaction (p< .05) indicates that habituation to spontaneous locomotor activity was revealed as a leftward and downward shift in activity across the three testing days.
3.2.2 Rearing
Analysis of the rearing data revealed main effects of Day [FG-G (2, 56) = 13.881, p<0.001] and Time [FG-G (11, 308) = 270.889, p<0.001]. There was also a significant Day × Time interaction [F G-G (22, 616) = 2.793, p<0.01]. Like the horizontal activity data, the Day × Time interaction indicates a downward and leftward shift in mean rearing as a function of day. Neither sex nor gestational treatment affected the rate or magnitude of the habituated response (data not shown).
3.3 Experiment 2: Sucrose Maintained Operant Responding
3.3.1 Acquisition Criterion
Fisher’s exact tests were conducted on the training trials-to-criterion sessions for the FR-1 and FR-3 schedules. Sixty percent of GN and GS rats achieved criterion on the FR-1 by the 5th day of responding on that schedule. The percentage of GN and GS animals meeting the criterion progressively increased across days and 100% of the GN and GS rats met the criterion for the FR-1 on day 11. There were no significant differences between the GN and GS groups’ rate to acquire criterion on the continuous reinforcement schedule (data not shown). Approximately 75 percent of the GN and GS rats met the criterion for the FR-3 after two days of moving from the FR-1 schedule; and 100 percent of animals reached criterion after three days on the FR-3 schedule. There were no significant differences between the GN and GS groups’ rate to acquire the criterion on the FR-3 schedule (data not shown).
3.3.2 FR-Responding
The FR data are presented in Figure 3. A mixed-design ANOVA revealed a significant main effect of Concentration [F G-G (4, 92) = 38.792, p<0.001] and no other significant effects were indicated. The significant effect of Concentration is derived from a quadratic function that represents an inverted U shaped curve. Rats exhibited peak responding at the 10% sucrose concentration and responding decreased as the sucrose concentration increased to 30 and 56%. These results indicate that the GN and GS animals showed equivalent sensitivity to various sucrose concentrations.
Figure 3.
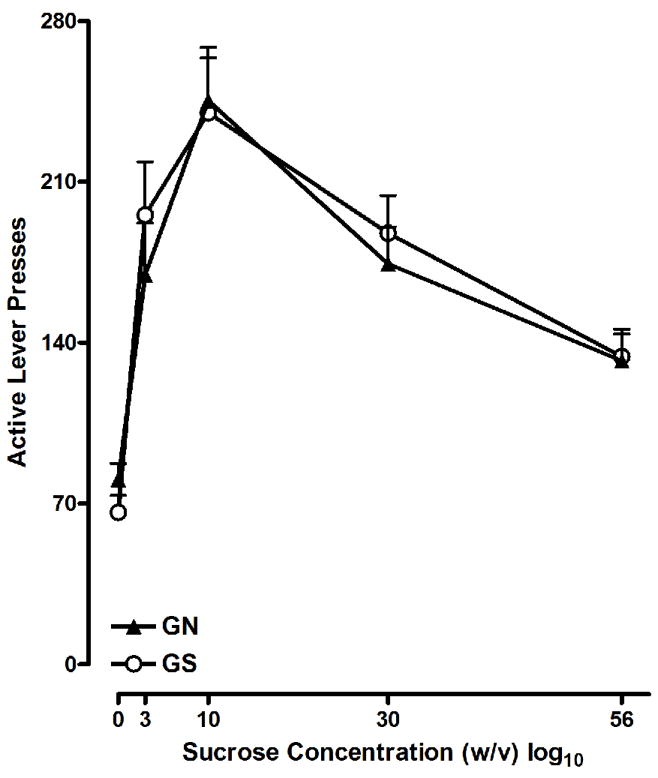
Mean (±SEM) active level presses for 0, 3, 10, 30, and 56% (w/v) sucrose on an FR-3 schedule. GN and GS rats showed similar concentration response curves. Peak responding occurred on trials that made the 10% sucrose concentration available.
3.3.3 PR-Responding
The PR data are presented in Figure 4. Analysis of the PR-maintained responding identified significant main effects of Concentration [F(4, 104) = 23.733, p<0.001] and Gestation [F(1, 26) = 5.500, p<0.05]. Animals showed increased active lever responding as a function of increasing sucrose concentration, but the Gestation × Concentration interaction did not reach significance. Sex was not a significant factor in this analysis. Analysis of inactive lever pressing revealed a main effect of Gestation [F(1,26) = 4.38, p<0.05], which indicates that the GN group showed more inactive responding (mean ± SEM: 21.4 ± 2.7) than the GS group (13.0 ± 2.9). Analysis of the breakpoint data also revealed significant main effects of Concentration [F(4, 104) = 21.609, p<0.001] and Gestation [F(1, 26) = 4.663, p<0.05; data not shown]. These results show that GN rats exhibited increased motivation for sucrose reward relative to GS animals (Figure 4B).
Figure 4.
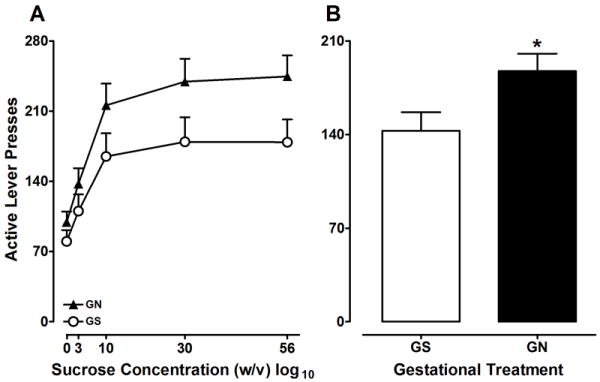
(A) Mean (±SEM) active level presses for 0, 3, 10, 30, and 56% (w/v) sucrose from the PR schedule by GN and GS rats. The Concentration × Gestation interaction did not reach significance. (B) Mean (±SEM) active level presses for sucrose (collapsed across concentration) in GN and GS rats. The asterisk represents the main effect of Gestation (p<0.05).
4. Discussion
The present experiments assessed the behavioral effects of a low dose, intermittent prenatal IV nicotine exposure model on adult offspring’s habituation to novelty exploration and on sucrose maintained operant responding, according to FR and PR schedules of reinforcement. Data from experiment 1 suggest that there was no overall effect of GN exposure on habituation to locomotor activity, elicited by context novelty, across three consecutive days of testing. In experiment 2, there were no differences between the prenatal exposure groups when varying concentrations of sucrose were tested using the FR-3 schedule; however the GN exposed rats exhibited increased responding for sucrose when the same concentrations were tested using the PR schedule. Thus, the FR-3 findings indicate that there were no differences in GN or GS rats’ sensitivity to detect a range of sucrose concentrations. When tested using the PR schedule, the GN rats exhibited more responding than the GS animals, indicating greater motivation for sucrose.
Previous research indicates that GN exposure resulted in either hyperactivity (Pauly et al., 2004; Paz et al., 2007; Peters et al., 1979; Tizabi et al., 1997; Tizabi et al., 2000), hypoactivity (LeSage et al., 2006; Peters and Tang, 1982; Romero and Chen, 2004), or no change (Franke et al., 2007; Gaworski et al., 2004; Paulson et al., 1993) in spontaneous locomotor behavior (see LeSage et al., 2006). The findings from the low-dose IV exposure model used in the present experiment are in accord with previous studies reporting no effect of prenatal nicotine on locomotor behavior in adult offspring. These results are also consistent with previous locomotor activity data from our laboratory, which show that there were also no differences attributable to gestational treatment in the habituation of spontaneous locomotor activity over three days of testing when offspring were adolescents, i.e., PND 28-35 (Harrod et al., 2011). This lack of GN effect on habituation may be related to the daily amount of nicotine delivered. For example, LeSage et al. (2006) administered multiple high dose, IV nicotine boli during gestation and showed that preweanling, male offspring exhibited hypoactivity relative to the GS controls. In their study, the dam was exposed to 0.03 mg/kg/injection of nicotine every 14 minutes for 16 hours/day on GD 4-21. This procedure achieves nicotine levels comparable to that delivered to animals via osmotic mini-pump (e.g., 2 mg/kg/day; see also Dwyer et al., 2009). The dose of IV nicotine used in the present experiment was 0.05 mg/kg/injection (0.15 mg/kg/day), which is considerably lower than that achieved in previous studies (LeSage et al. 2006; Tizabi et al., 1997).
This daily concentration of IV GN did produce changes in motivated behavior. The conclusion that GN animals exhibited increased motivation to respond for sucrose reward is consistent with the findings of a prospective study, which shows that individuals exposed to in utero tobacco smoke consume greater amounts of highly palatable foods, such as soft drinks and red meat, compared to non-exposed offspring (Al Mamun et al., 2006a). This study also reported that maternal smoking during pregnancy was significantly correlated with a higher body mass index, and increased odds of being overweight and/or obese at 14 years of age. The increased motivation for sucrose reward exhibited by the GN animals in the present experiment suggests that nicotine may be the important constituent in tobacco smoke that influences the increased consumption of sucrose-laden foods during adolescence in human offspring. Although the GN animals exhibited greater motivation for sucrose, there were no weight differences between the GN and GS rats prior to dipper training, which was initiated in adulthood.
The overall exposure levels to GN may also differentially impact offsprings’ response to sucrose reinforcement in preclinical studies. For example, Franke et al. (2008) reported that adolescent male rat offspring exposed to continuous prenatal nicotine (~2 mg/kg/day) showed decreased motivation for sucrose pellets acquired on a continuous reinforcement schedule. When the response requirement was raised to an FR-2 and then to an FR-5, the difference in the motivation to respond for sucrose by the GN- relative to GS-exposed rats was resolved. The results from the present experiment show that adult GN and GS rats exhibited equivalent sensitivity to a range of sucrose concentrations when a FR-3 schedule of reinforcement was used: the shape of the concentration response curve was identical in GN and GS rats. During PR tests, the GN rats, which were prenatally treated with 0.15 mg/kg/day showed more motivation for sucrose reinforcement compared to the GS group, and this was more evident at the higher concentrations of sucrose. These findings suggest that the amount of GN administration may differentially impact operant behavior in offspring. Notably, another methodological difference, such as age at testing may have contributed to the different findings. The general effects of GN exposure on the motivational thresholds for sucrose reinforcement may be manifested differently in GN adolescents and GN adults, given that dopaminergic tone increases drastically throughout the mesocorticolimbic pathway during adolescence (Chambers et al., 2003; Spear, 2000; Teicher et al., 1995), and further, that GN has been shown to alter development of the mesocorticolimbic system during adolescence (Harrod et al., 2011; Kane et al., 2004; Muneoka et al., 1997; Navarro et al., 1989; Park et al., 2006; Ribary and Lichtensteiger, 1989; Richardson and Tizabi, 1994). Thus it is possible that age of testing contributes to the effect of gestational nicotine exposure on responding for sucrose reward. In general, these findings demonstrate that two different methods of prenatal nicotine administration alter the motivation for sucrose reward in adolescent and adult offspring.
The administration of 3×/day low-dose IV nicotine during gestation resulted in long-term neurobehavioral changes in offspring. The present results add to previous findings from our laboratory, which report that IV nicotine exposure altered sensorimotor gating of the acoustic startle response, and resulted in increased levels of brain-derived neurotrophic factor in the nucleus accumbens, striatum, and frontal cortex in GN animals compared to GS controls (Harrod et al., 2011; Lacy et al., 2011). These findings suggest that pregnant women who smoke a “low” number of daily tobacco cigarettes may produce enduring changes within the mesocorticolimbic circuitry, and alter motivated behavior, particularly for highly palatable foods in later stages of development, such as adolescence and adulthood. Future experiments will need to determine the neurophysiological factors that are affected by prenatal, low-dose IV nicotine exposure, to gain a better understanding of how in utero exposure to nicotine alters the motivation for palatable foods, thereby increasing the risk for adolescent and adult obesity.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Al Mamun A, Lawlor DA, Alati R, O’Callaghan MJ, Williams GM, Najman JM. Does maternal smoking during pregnancy have a direct effect on future offspring obesity? Evidence from a prospective birth cohort study. Am J Epidemiol. 2006a;164:317–325. doi: 10.1093/aje/kwj209. [DOI] [PubMed] [Google Scholar]
- Al Mamun A, O’Callaghan FV, Alati R, O’Callaghan M, Najman JM, Williams GM, Bor W. Does maternal smoking during pregnancy predict the smoking patterns of young adult offspring? A birth cohort study. Tob Control. 2006b;15:452–457. doi: 10.1136/tc.2006.016790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benowitz NL, Hukkanen J, Jacob P., 3rd Nicotine chemistry, metabolism, kinetics and biomarkers. Handb Exp Pharmacol. 2009:29–60. doi: 10.1007/978-3-540-69248-5_2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Booze RM, Welch MA, Wood ML, Billings KA, Apple SR, Mactutus CF. Behavioral sensitization following repeated intravenous nicotine administration: gender differences and gonadal hormones. Pharmacol Biochem Behav. 1999;64:827–839. doi: 10.1016/s0091-3057(99)00169-0. [DOI] [PubMed] [Google Scholar]
- Buka SL, Shenassa ED, Niaura R. Elevated risk of tobacco dependence among offspring of mothers who smoked during pregnancy: a 30-year prospective study. Am J Psychiatry. 2003;160:1978–1984. doi: 10.1176/appi.ajp.160.11.1978. [DOI] [PubMed] [Google Scholar]
- Button TM, Maughan B, McGuffin P. The relationship of maternal smoking to psychological problems in the offspring. Early Hum Dev. 2007;83:727–732. doi: 10.1016/j.earlhumdev.2007.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Button TM, Thapar A, McGuffin P. Relationship between antisocial behaviour, attention-deficit hyperactivity disorder and maternal prenatal smoking. Br J Psychiatry. 2005;187:155–160. doi: 10.1192/bjp.187.2.155. [DOI] [PubMed] [Google Scholar]
- Chambers RA, Taylor JR, Potenza MN. Developmental neurocircuitry of motivation in adolescence: a critical period of addiction vulnerability. Am J Psychiatry. 2003;160:1041–1052. doi: 10.1176/appi.ajp.160.6.1041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaudhri N, Caggiula AR, Donny EC, Booth S, Gharib MA, Craven LA, Allen SS, Sved AF, Perkins KA. Sex differences in the contribution of nicotine and nonpharmacological stimuli to nicotine self-administration in rats. Psychopharmacology (Berl) 2005;180:258–266. doi: 10.1007/s00213-005-2152-3. [DOI] [PubMed] [Google Scholar]
- Corrigall WA, Coen KM. Nicotine maintains robust self-administration in rats on a limited-access schedule. Psychopharmacology (Berl) 1989;99:473–478. doi: 10.1007/BF00589894. [DOI] [PubMed] [Google Scholar]
- Corrigall WA, Franklin KB, Coen KM, Clarke PB. The mesolimbic dopaminergic system is implicated in the reinforcing effects of nicotine. Psychopharmacology (Berl) 1992;107:285–289. doi: 10.1007/BF02245149. [DOI] [PubMed] [Google Scholar]
- Dwyer JB, Broide RS, Leslie FM. Nicotine and brain development. Birth Defects Res C Embryo Today Rev. 2008;84:30–44. doi: 10.1002/bdrc.20118. [DOI] [PubMed] [Google Scholar]
- Dwyer JB, McQuown SC, Leslie FM. The dynamic effects of nicotine on the developing brain. Pharmacol Ther. 2009;122:125–139. doi: 10.1016/j.pharmthera.2009.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edwards S, Koob GF. Neurobiology of dysregulated motivational systems in drug addiction. Future Neurol. 2010;5:393–401. doi: 10.2217/fnl.10.14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Everitt BJ, Belin D, Economidou D, Pelloux Y, Dalley JW, Robbins TW. Review. Neural mechanisms underlying the vulnerability to develop compulsive drug-seeking habits and addiction. Philos Trans R Soc Lond B Biol Sci. 2008;363:3125–3135. doi: 10.1098/rstb.2008.0089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franke RM, Belluzzi JD, Leslie FM. Gestational exposure to nicotine and monoamine oxidase inhibitors influences cocaine-induced locomotion in adolescent rats. Psychopharmacology (Berl) 2007;195:117–124. doi: 10.1007/s00213-007-0876-y. [DOI] [PubMed] [Google Scholar]
- Franke RM, Park M, Belluzzi JD, Leslie FM. Prenatal nicotine exposure changes natural and drug-induced reinforcement in adolescent male rats. Eur J Neurosci. 2008;27:2952–2961. doi: 10.1111/j.1460-9568.2008.06253.x. [DOI] [PubMed] [Google Scholar]
- Fried PA, Watkinson B, Gray R. Differential effects on cognitive functioning in 9- to 12-year olds prenatally exposed to cigarettes and marihuana. Neurotoxicol Teratol. 1998;20:293–306. doi: 10.1016/s0892-0362(97)00091-3. [DOI] [PubMed] [Google Scholar]
- Fried PA, Watkinson B, Gray R. Differential effects on cognitive functioning in 13- to 16-year-olds prenatally exposed to cigarettes and marihuana. Neurotoxicol Teratol. 2003;25:427–436. doi: 10.1016/s0892-0362(03)00029-1. [DOI] [PubMed] [Google Scholar]
- Gaworski CL, Carmines EL, Faqi AS, Rajendran N. In utero and lactation exposure of rats to 1R4F reference cigarette mainstream smoke: effect on prenatal and postnatal development. Toxicol Sci. 2004;79:157–169. doi: 10.1093/toxsci/kfh083. [DOI] [PubMed] [Google Scholar]
- Harrod SB, Lacy RT, Zhu J, Hughes BA, Perna MK, Brown RW. Gestational IV nicotine produces elevated brain-derived neurotrophic factor in the mesocorticolimbic dopamine system of adolescent rat offspring. Synapse. 2011;65:1382–1392. doi: 10.1002/syn.20975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrod SB, Mactutus CF, Bennett K, Hasselrot U, Wu G, Welch M, Booze RM. Sex differences and repeated intravenous nicotine: behavioral sensitization and dopamine receptors. Pharmacol Biochem Behav. 2004;78:581–592. doi: 10.1016/j.pbb.2004.04.026. [DOI] [PubMed] [Google Scholar]
- Holson RR, Pearce B. Principles and pitfalls in the analysis of prenatal treatment effects in multiparous species. Neurotoxicol Teratol. 1992;14:221–228. doi: 10.1016/0892-0362(92)90020-b. [DOI] [PubMed] [Google Scholar]
- Jacobsen LK, Slotkin TA, Mencl WE, Frost SJ, Pugh KR. Gender-specific effects of prenatal and adolescent exposure to tobacco smoke on auditory and visual attention. Neuropsychopharmacology. 2007;32:2453–2464. doi: 10.1038/sj.npp.1301398. [DOI] [PubMed] [Google Scholar]
- Jacobsen LK, Slotkin TA, Westerveld M, Mencl WE, Pugh KR. Visuospatial memory deficits emerging during nicotine withdrawal in adolescents with prenatal exposure to active maternal smoking. Neuropsychopharmacology. 2006;31:1550–1561. doi: 10.1038/sj.npp.1300981. [DOI] [PubMed] [Google Scholar]
- Kalivas PW. The glutamate homeostasis hypothesis of addiction. Nat Rev Neurosci. 2009;10:561–572. doi: 10.1038/nrn2515. [DOI] [PubMed] [Google Scholar]
- Kandel DB, Wu P, Davies M. Maternal smoking during pregnancy and smoking by adolescent daughters. Am J Public Health. 1994;84:1407–1413. doi: 10.2105/ajph.84.9.1407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kane VB, Fu Y, Matta SG, Sharp BM. Gestational nicotine exposure attenuates nicotine-stimulated dopamine release in the nucleus accumbens shell of adolescent Lewis rats. J Pharmacol Exp Ther. 2004;308:521–528. doi: 10.1124/jpet.103.059899. [DOI] [PubMed] [Google Scholar]
- Lacy RT, Mactutus CF, Harrod SB. Prenatal IV nicotine exposure produces a sex difference in sensorimotor gating of the auditory startle reflex in adult rats. Int J Dev Neurosci. 2011;29:153–161. doi: 10.1016/j.ijdevneu.2010.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LeSage MG, Gustaf E, Dufek MB, Pentel PR. Effects of maternal intravenous nicotine administration on locomotor behavior in pre-weanling rats. Pharmacol Biochem Behav. 2006;85:575–583. doi: 10.1016/j.pbb.2006.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levin ED, Lawrence S, Petro A, Horton K, Seidler FJ, Slotkin TA. Increased nicotine self-administration following prenatal exposure in female rats. Pharmacol Biochem Behav. 2006;85:669–674. doi: 10.1016/j.pbb.2006.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Linnet KM, Dalsgaard S, Obel C, Wisborg K, Henriksen TB, Rodriguez A, Kotimaa A, Moilanen I, Thomsen PH, Olsen J, Jarvelin MR. Maternal lifestyle factors in pregnancy risk of attention deficit hyperactivity disorder and associated behaviors: review of the current evidence. Am J Psychiatry. 2003;160:1028–1040. doi: 10.1176/appi.ajp.160.6.1028. [DOI] [PubMed] [Google Scholar]
- Mactutus CF. Developmental neurotoxicity of nicotine, carbon monoxide, and other tobacco smoke constituents. Ann N Y Acad Sci. 1989;562:105–122. doi: 10.1111/j.1749-6632.1989.tb21010.x. [DOI] [PubMed] [Google Scholar]
- Mactutus CF, Herman AS, Booze RM. Chronic intravenous model for studies of drug (ab)use in the pregnant and/or group-housed rat: an initial study with cocaine. Neurotoxicol Teratol. 1994;16:183–191. doi: 10.1016/0892-0362(94)90116-3. [DOI] [PubMed] [Google Scholar]
- Martin JA, Kung HC, Mathews TJ, Hoyert DL, Strobino DM, Guyer B, Sutton SR. Annual summary of vital statistics: 2006. Pediatrics. 2008;121:788–801. doi: 10.1542/peds.2007-3753. [DOI] [PubMed] [Google Scholar]
- McCartney JS, Fried PA, Watkinson B. Central auditory processing in school-age children prenatally exposed to cigarette smoke. Neurotoxicol Teratol. 1994;16:269–276. doi: 10.1016/0892-0362(94)90048-5. [DOI] [PubMed] [Google Scholar]
- Muneoka K, Ogawa T, Kamei K, Muraoka S, Tomiyoshi R, Mimura Y, Kato H, Suzuki MR, Takigawa M. Prenatal nicotine exposure affects the development of the central serotonergic system as well as the dopaminergic system in rat offspring: involvement of route of drug administrations. Brain Res Dev Brain Res. 1997;102:117–126. doi: 10.1016/s0165-3806(97)00092-8. [DOI] [PubMed] [Google Scholar]
- Navarro HA, Seidler FJ, Eylers JP, Baker FE, Dobbins SS, Lappi SE, Slotkin TA. Effects of prenatal nicotine exposure on development of central and peripheral cholinergic neurotransmitter systems. Evidence for cholinergic trophic influences in developing brain. J Pharmacol Exp Ther. 1989;251:894–900. [PubMed] [Google Scholar]
- Park MK, Loughlin SE, Leslie FM. Gestational nicotine-induced changes in adolescent neuronal activity. Brain Res. 2006;1094:119–126. doi: 10.1016/j.brainres.2006.04.001. [DOI] [PubMed] [Google Scholar]
- Paulson RB, Shanfeld J, Vorhees CV, Sweazy A, Gagni S, Smith AR, Paulson JO. Behavioral effects of prenatally administered smokeless tobacco on rat offspring. Neurotoxicol Teratol. 1993;15:183–192. doi: 10.1016/0892-0362(93)90014-f. [DOI] [PubMed] [Google Scholar]
- Pauly JR, Slotkin TA. Maternal tobacco smoking, nicotine replacement and neurobehavioural development. Acta Paediatr. 2008;97:1331–1337. doi: 10.1111/j.1651-2227.2008.00852.x. [DOI] [PubMed] [Google Scholar]
- Pauly JR, Sparks JA, Hauser KF, Pauly TH. In utero nicotine exposure causes persistent, gender-dependant changes in locomotor activity and sensitivity to nicotine in C57Bl/6 mice. Int J Dev Neurosci. 2004;22:329–337. doi: 10.1016/j.ijdevneu.2004.05.009. [DOI] [PubMed] [Google Scholar]
- Paz R, Barsness B, Martenson T, Tanner D, Allan AM. Behavioral teratogenicity induced by nonforced maternal nicotine consumption. Neuropsychopharmacology. 2007;32:693–699. doi: 10.1038/sj.npp.1301066. [DOI] [PubMed] [Google Scholar]
- Pentel PR, Dufek MB, Roiko SA, Lesage MG, Keyler DE. Differential effects of passive immunization with nicotine-specific antibodies on the acute and chronic distribution of nicotine to brain in rats. J Pharmacol Exp Ther. 2006;317:660–666. doi: 10.1124/jpet.105.097873. [DOI] [PubMed] [Google Scholar]
- Peters DA, Tang S. Sex-dependent biological changes following prenatal nicotine exposure in the rat. Pharmacol Biochem Behav. 1982;17:1077–1082. doi: 10.1016/0091-3057(82)90497-x. [DOI] [PubMed] [Google Scholar]
- Peters DA, Taub H, Tang S. Postnatal effects of maternal nicotine exposure. Neurobehav Toxicol. 1979;1:221–225. [PubMed] [Google Scholar]
- Picone TA, Allen LH, Olsen PN, Ferris ME. Pregnancy outcome in North American women. II. Effects of diet, cigarette smoking, stress, and weight gain on placentas, and on neonatal physical and behavioral characteristics. Am J Clin Nutr. 1982;36:1214–1224. doi: 10.1093/ajcn/36.6.1214. [DOI] [PubMed] [Google Scholar]
- Reichel CM, Linkugel JD, Bevins RA. Bupropion differentially impacts acquisition of methamphetamine self-administration and sucrose-maintained behavior. Pharmacol Biochem Behav. 2008;89:463–472. doi: 10.1016/j.pbb.2008.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ribary U, Lichtensteiger W. Effects of acute and chronic prenatal nicotine treatment on central catecholamine systems of male and female rat fetuses and offspring. J Pharmacol Exp Ther. 1989;248:786–792. [PubMed] [Google Scholar]
- Richardson NR, Roberts DC. Progressive ratio schedules in drug self-administration studies in rats: a method to evaluate reinforcing efficacy. J Neurosci Methods. 1996;66:1–11. doi: 10.1016/0165-0270(95)00153-0. [DOI] [PubMed] [Google Scholar]
- Richardson SA, Tizabi Y. Hyperactivity in the offspring of nicotine-treated rats: role of the mesolimbic and nigrostriatal dopaminergic pathways. Pharmacol Biochem Behav. 1994;47:331–337. doi: 10.1016/0091-3057(94)90018-3. [DOI] [PubMed] [Google Scholar]
- Robinson TE, Berridge KC. Addiction. Annu Rev Psychol. 2003;54:25–53. doi: 10.1146/annurev.psych.54.101601.145237. [DOI] [PubMed] [Google Scholar]
- Romero RD, Chen WJ. Gender-related response in open-field activity following developmental nicotine exposure in rats. Pharmacol Biochem Behav. 2004;78:675–681. doi: 10.1016/j.pbb.2004.04.033. [DOI] [PubMed] [Google Scholar]
- Russell MA, Feyerabend C. Cigarette smoking: a dependence on high-nicotine boli. Drug Metab Rev. 1978;8:29–57. doi: 10.3109/03602537808993776. [DOI] [PubMed] [Google Scholar]
- Salihu HM, Wilson RE. Epidemiology of prenatal smoking and perinatal outcomes. Early Hum Dev. 2007;83:713–720. doi: 10.1016/j.earlhumdev.2007.08.002. [DOI] [PubMed] [Google Scholar]
- Saxton DW. The behaviour of infants whose mothers smoke in pregnancy. Early Hum Dev. 1978;2:363–369. doi: 10.1016/0378-3782(78)90063-4. [DOI] [PubMed] [Google Scholar]
- Sexton M, Fox NL, Hebel JR. Prenatal exposure to tobacco: II. effects on cognitive functioning at age three. Int J Epidemiol. 1990;19:72–77. doi: 10.1093/ije/19.1.72. [DOI] [PubMed] [Google Scholar]
- Siegel S, Castellan NJ. Nonparametric Statistics for the Behavioral Sciences. McGraw-Hill; New York: 1988. [Google Scholar]
- Slotkin TA, Tate CA, Cousins MM, Seidler FJ. Prenatal nicotine exposure alters the responses to subsequent nicotine administration and withdrawal in adolescence: Serotonin receptors and cell signaling. Neuropsychopharmacology. 2006;31:2462–2475. doi: 10.1038/sj.npp.1300988. [DOI] [PubMed] [Google Scholar]
- Spear LP. The adolescent brain and age-related behavioral manifestations. Neurosci Biobehav Rev. 2000;24:417–463. doi: 10.1016/s0149-7634(00)00014-2. [DOI] [PubMed] [Google Scholar]
- Teicher MH, Andersen SL, Hostetter JC., Jr Evidence for dopamine receptor pruning between adolescence and adulthood in striatum but not nucleus accumbens. Brain Res Dev Brain Res. 1995;89:167–172. doi: 10.1016/0165-3806(95)00109-q. [DOI] [PubMed] [Google Scholar]
- Thapar A, Fowler T, Rice F, Scourfield J, van den Bree M, Thomas H, Harold G, Hay D. Maternal smoking during pregnancy and attention deficit hyperactivity disorder symptoms in offspring. Am J Psychiatry. 2003;160:1985–1989. doi: 10.1176/appi.ajp.160.11.1985. [DOI] [PubMed] [Google Scholar]
- Tizabi Y, Popke EJ, Rahman MA, Nespor SM, Grunberg NE. Hyperactivity induced by prenatal nicotine exposure is associated with an increase in cortical nicotinic receptors. Pharmacol Biochem Behav. 1997;58:141–146. doi: 10.1016/s0091-3057(96)00461-3. [DOI] [PubMed] [Google Scholar]
- Tizabi Y, Russell LT, Nespor SM, Perry DC, Grunberg NE. Prenatal nicotine exposure: effects on locomotor activity and central [125I]alpha-BT binding in rats. Pharmacol Biochem Behav. 2000;66:495–500. doi: 10.1016/s0091-3057(00)00171-4. [DOI] [PubMed] [Google Scholar]
- Tong VT, Jones JR, Dietz PM, D’Angelo D, Bombard JM. Trends in smoking before, during, and after pregnancy - Pregnancy Risk Assessment Monitoring System (PRAMS), United States, 31 sites, 2000-2005. MMWR Surveill Summ. 2009;58:1–29. [PubMed] [Google Scholar]
- Toschke AM, Ehlin AG, von Kries R, Ekbom A, Montgomery SM. Maternal smoking during pregnancy and appetite control in offspring. J Perinat Med. 2003;31:251–256. doi: 10.1515/JPM.2003.034. [DOI] [PubMed] [Google Scholar]
- Vaglenova J, Birru S, Pandiella NM, Breese CR. An assessment of the long-term developmental and behavioral teratogenicity of prenatal nicotine exposure. Behav Brain Res. 2004;150:159–170. doi: 10.1016/j.bbr.2003.07.005. [DOI] [PubMed] [Google Scholar]
- Vorhees CV. Principles of behavioral teratology. In: Riley EP, Vorhees CV, editors. Handbook of Behavioral Teratology. Plenum Press; New York, NY US: 1986. pp. 23–48. [Google Scholar]
- Wei J, Wang J, Dwyer JB, Mangold J, Cao J, Leslie FM, Li MD. Gestational nicotine treatment modulates cell death/survival-related pathways in the brains of adolescent female rats. Int J Neuropsychopharmacol. 2011;14:91–106. doi: 10.1017/S1461145710000416. [DOI] [PubMed] [Google Scholar]
- Weissman MM, Warner V, Wickramaratne PJ, Kandel DB. Maternal smoking during pregnancy and psychopathology in offspring followed to adulthood. J Am Acad Child Adolesc Psychiatry. 1999;38:892–899. doi: 10.1097/00004583-199907000-00020. [DOI] [PubMed] [Google Scholar]
- Wise RA, Bozarth MA. A psychomotor stimulant theory of addiction. Psychol Rev. 1987;94:469–492. [PubMed] [Google Scholar]
- Zhu J, Takita M, Konishi Y, Sudo M, Muramatsu I. Chronic nicotine treatment delays the developmental increase in brain muscarinic receptors in rat neonate. Brain Res. 1996;732:257–260. doi: 10.1016/0006-8993(96)00704-4. [DOI] [PubMed] [Google Scholar]