

Abstract
In addition to cellular immune responses, humoral immune responses, mediated by natural antibodies, autoantibodies, and alloantibodies, have increasingly been recognized as causes of organ transplant rejection. In our previous studies, we have demonstrated the induction of antinuclear antibodies against histone H1 and high-mobility group box 1 (HMGB1), in both experimental and clinical liver transplant tolerance. The active induction of antinuclear antibodies is usually an undesirable phenomenon, but it is often observed after liver transplantation. However, the release of nuclear antigens and its suppression by neutralizing antibodies are proposed to be important in the initiation and regulation of immune responses. In this review article, we summarize the current understanding of nuclear antigens and corresponding antinuclear regulatory antibodies (Abregs) on infection, injury, inflammation, transplant rejection, and tolerance induction and discuss the significance of nuclear antigens as diagnostic and therapeutic targets.
1. Introduction
Transplantation of cells, tissues, or organs is now widely used to cure patients with life-threatening diseases or traumatic injuries. Except for the use of self-derived grafts or grafts from an identical twin, allograft rejection can be observed acutely and/or chronically [1, 2]. In the current practice of transplantation, the administration of immunosuppressants, such as tacrolimus (FK506) and cyclosporin A, is indispensable for the prevention of allograft rejection [3]. However, the use of these immunosuppressants has limitations, including the necessity of long-term medication and serious side effects, such as nephrotoxicity [4], cardiovascular toxicity [5], and cancer [6]. Therefore, the development of safer and more effective immunosuppressants as well as useful diagnostic tools for the prediction of rejection is an important subject for further improvement of the quality of life of patients and their families after transplantation.
Since the early days of experimental and clinical liver transplantation, it has been known that this organ does not always obey the normal rules of transplant rejection (Medawar's rule of transplantation); for example, all grafts are rejected between unrelated individuals, and the survival rate following liver transplantation is higher than that following the transplantation of other organs [7, 8]. In Dark Agouti (DA) donor livers transplanted into Piebald Virol Glaxo (PVG) recipients, allograft rejection is spontaneously overcome after orthotopic liver transplantation (OLT), resulting in a state of long-lasting and donor-specific tolerance without pharmacological immunosuppression, although PVG recipients acutely reject skin, heart, and renal grafts from DA rats [9]. Interestingly, PVG recipients bearing DA livers could accept skin, heart, and kidney transplants from the DA donor rats but rejected them from third-party strains of rats [10, 11]. The molecular and cellular basis of liver transplant tolerogenicity has not been fully elucidated, but the unique repertoires of nonparenchymal cells including liver antigen-presenting cells (e.g., dendritic cells (DCs), Kupffer cells, and liver sinusoidal endothelial cells) and unconventional lymphoid cells (e.g., NK cells, B-1 cells, and γδ T cells), which are rarely present in the blood, may explain the immune privilege of the liver [12]. Our recent study also suggested that mast cells in the donor grafts may play important roles in the induction/maintenance of immune tolerance and liver regeneration, resulting in the replacement of hepatic cells from donor to recipient [13]. In addition, several humoral factors in the serum of a rat tolerogenic OLT model have been identified as immunosuppressive factors, including donor-soluble MHC class I molecules [14], antidonor MHC class II antibodies [15], liver suppressor factor-1 (LSF-1; 40 kDa) [16, 17], LSF-2 (87 kDa), and LSF-3 (10 kDa) [18]. However, most of these humoral factors are found only in the experimental OLT model, and it is hard to translate the findings of this animal study to clinical practice.
In the past decade, we further evaluated humoral factors, specifically IgG antibodies, which are immediately elevated and maintained at a higher level even after the recipients accept the donor liver allografts and demonstrated strong immunosuppressive activity in vitro [19, 20]. The screening of autoantigens recognized by immunosuppressive IgG antibodies in the post-OLT sera revealed the spontaneous induction of antinuclear antibodies against histone H1 and high-mobility group box 1 (HMGB1), both in the DA-PVG natural tolerance model and in a patient with operational tolerance [19–22].
In this review article, we summarize the current understanding of nuclear antigens and corresponding antinuclear regulatory antibodies (Abregs) on infection, injury, inflammation, transplant rejection, and tolerance induction and discuss the significance of nuclear antigens as diagnostic and therapeutic targets.
2. Induction of Humoral Immune Responses after Transplantation: Link to Rejection or Tolerance?
In the past, organ transplant rejection and tolerance were believed to be mediated almost exclusively by cellular immune responses. Although improvements in T-cell-directed immunosuppression have decreased the incidence of acute cellular rejection, humoral immune responses, mediated by natural antibodies, autoantibodies, and alloantibodies, have increasingly been recognized as causes of organ transplant rejection [23, 24]. The overall incidence of antibody-mediated rejection (AMR) is estimated to be 20%–30% for renal transplant recipients [25]. However, AMR is mainly discussed in ABO blood type-incompatible liver transplantation [26]. Natural antibodies against A/B carbohydrate determinants are likely to develop as a result of exposure to environmental bacteria that express similar determinants. The response of the B-cell compartment to environmental antigens/microbial products and autoantigens has been thought to be derived preferentially from the activation of CD5+ B-1 cells. Therefore, CD5+ B-1 cells have been speculated to be the major B-cell subset responding to A/B determinants in both mice and humans [27–29]. B-1 cells are present in low numbers in the lymph nodes and spleen and are instead found predominantly in the peritoneal and pleural cavity [30, 31]. Recent reports suggest that splenic CD1dhiCD5+ B cells are potent regulatory cells that produce IL-10 in models of contact hypersensitivity and experimental allergic encephalomyelitis [32, 33]. Furthermore, Moritoki et al. reported that B cells or B-cell subsets may affect the induction and function of regulatory T cells (Tregs) as suppressors of the T-cell component [34]. Chhabra et al. recently reported the prevention of autoimmune diabetes and the prolongation of islet allograft survival by the administration of naturally occurring IgM autoantibodies [35]. These findings strongly suggest that the induction of natural antibodies or autoantibodies may play an important role in immune regulation and tolerance induction after transplantation.
In our previous studies, we have demonstrated the induction of antinuclear antibodies against histone H1 and HMGB1 both in a rat tolerogenic OLT model and in a patient with operational tolerance [19–22]. In the field of liver transplantation, the induction of autoantibodies (e.g., antinuclear antibody, smooth muscle antibody, and liver-kidney microsomal antibody) has often been observed, particularly in pediatric recipients [36], while the incidence of de novo autoimmune hepatitis in children with elevated serum autoantibodies and liver function tests, hypergammaglobulinemia, and liver pathology showing necroinflammatory disease and fibrosis has been found to be just 1%–7% [37–39]. We also confirmed the significance of antinuclear antibody for protection and recovery from the concanavalin A-(Con A-) induced liver injury mimic of autoimmune hepatitis [40]. Therefore, the induction of autoantibodies in most recipients after liver transplantation may not be associated with any clinical manifestations of autoimmune disorders. A recent study also demonstrated that the long-term administration of tacrolimus to liver transplant recipients induces the production of antinuclear antibodies, whereas the autoimmune disease susceptibility of recipients treated with tacrolimus has not been elucidated [41]. The active induction of antinuclear antibodies is usually an undesirable phenomenon, but why is it often observed after liver transplantation? Is it linked to the immune privilege of the liver? The answers to these questions are still uncertain, but we speculate that the existence of antinuclear antibodies against histone H1 and HMGB1, which possess strong immunosuppressive activity in the systemic circulation, may regulate uncontrollable immune responses such as acute/chronic rejection after transplantation. In other words, the induction of antinuclear antibodies may be a “lethal weapon” to escape the breakdown of our immune system at least in transplant immunology. Our hypothesis is supported by Barnay-Verdier et al., who recently demonstrated that autoantibodies against HMGB1 are produced during sepsis and are associated with a favorable outcome in patients with septic shock [42].
3. Nuclear Antigens and Immunogenicity
Why are antinuclear antibodies against histone H1 and HMGB1 elevated in the specific condition of liver transplantation, and do they act as Abregs? An initial mechanism for the induction of antinuclear antibodies is the release of nuclear antigens, and the primary source of nuclear antigens would be damaged hepatic cells due to peritransplant ischemia/reperfusion injury and posttransplant rejection. Specifically, hepatic cell death by necrosis, apoptosis, and autophagy during cold ischemia and warm reperfusion during the course of liver transplantation triggers liver graft dysfunction [43–45]. Indeed, the release of nuclear antigens and its suppression by neutralizing antibodies are proposed to be important in the initiation and regulation of immune responses. HMGB1 is a ubiquitous and abundant chromatin component, and it is currently well known as one of the damage-associated molecular pattern molecules (DAMPs) interacting with the receptor for advanced glycation end product (RAGE), toll-like receptor (TLR)2, TLR4, and TLR9 [46]. Wang et al. first reported the proinflammatory role of HMGB1 in endotoxin lethality in mice and in septic patients [47]. Since then, the proinflammatory roles of HMGB1 in the pathogenesis of many diseases have been reported, including acute lung inflammation [48], atherosclerosis and restenosis after vascular damage [49], severe acute pancreatitis [50], rheumatoid arthritis [51], pulmonary fibrosis [52], stroke [53], Kawasaki disease [54], cold ischemia/reperfusion-induced inflammation [55], liver fibrosis [56], systemic inflammatory response syndrome [57, 58], febrile seizures [59], hyperlipidemia [60], preeclampsia [61], and acute-on-chronic liver failure [62].
However, the roles of histones in immune responses are poorly understood in comparison with HMGB1. Histone H1 has been reported to possess various important functions including a role in transmitting apoptotic signals from the nucleus to the mitochondria, which release apoptogenic factors into the cytoplasm, following DNA double-strand breaks [63] and in normal DC differentiation, based on evidence demonstrating that the production and differentiation of DCs in histone H10-deficient mice are significantly reduced [64]. Our previous study has demonstrated that the translocation of histone H1 from the nucleus to the cytoplasm and the release of their own histone H1 are necessary for DC maturation and the T-cell activation [65]. This function is also similar to the role of HMGB1 in DC maturation [66]. In addition, recent work has clearly demonstrated the induction of inflammatory responses by extracellular histones from dying cells via TLR2 and TLR4 in acute kidney injury [67].
Taken together, these results strongly suggest the significance of nuclear antigens such as histones and HMGB1 that are released from damaged cells or actively secreted from activated immune cells such as DCs and macrophages in the initiation of immune responses during rejection as well as infection, injury, and inflammation (Figure 1). We speculate that the sensitivity to nuclear antigens (i.e., easy production of antinuclear Abregs) may be one of the key factors determining the acceptance or rejection of donor liver allografts [68]. To be exact, antinuclear antibodies include both auto- and alloantibodies due to the different sources of antigens (liver allografts: alloantigens, immune cells: autoantigens) in the case of liver transplantation. In this review article, however, we have defined the induction of antinuclear antibodies as an autoimmune response due to the homological similarity of nuclear antigens even in different species.
Figure 1.
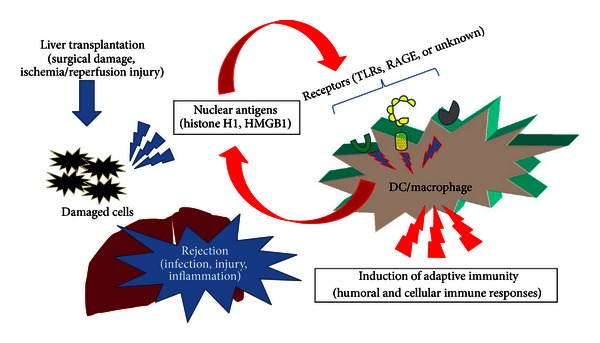
Histone H1 and HMGB1 as “nuclear weapons” in innate and adaptive immune responses. Nuclear antigens released from damaged cells or actively secreted from activated immune cells such as DCs and macrophages are important initiators for the activation of humoral and cellular immune responses during peritransplant ischemia/reperfusion injury and posttransplant rejection as well as infection, injury, and inflammation.
4. Nuclear Antigens as a Prognostic Marker for Rejection
The release of nuclear antigens into the blood stream has been associated with the progression of several diseases. Hatada et al. reported the elevation of plasma HMGB1 levels in patients with infectious diseases, malignancies, and traumas and suggested that HMGB1 is a potentially suitable prognostic marker of organ failure or disseminated intravascular coagulation [69]. The serum level of HMGB1 in patients with nonsmall cell lung cancer (NSCLC) was significantly higher compared to patients with chronic obstructive pulmonary disease, suggesting that HMGB1 may be a useful marker for evaluating NSCLC progression [70]. A positive association between the circulating HMGB1 level and cardiovascular mortality or traumatic brain injury has also been reported [71, 72]. As shown in Figure 2, the elevation of circulating histone H1 and HMGB1 was confirmed during the rejection phase (day 7) after OLT in a rat acute rejection combination (DA-LEW). However, mild or no elevation of circulating histone H1 and HMGB1 was confirmed in a rat tolerogenic combination (DA-PVG), suggesting the diagnostic potential for the prediction of acute rejection after transplantation. In our previous studies, we have confirmed the induction of humoral immune responses against histone H1 and HMGB1 only in the DA-PVG combination [21, 22], suggesting the blockade of the exposure of nuclear antigens by the induction of corresponding antinuclear Abregs. The induction of antinuclear Abregs could also suppress alloantibody production during the rejection phase (day 7) after OLT (Figure 3). Therefore, the balance between autoimmunity and alloimmunity is important for the prolongation of allograft survival (Figure 4).
Figure 2.
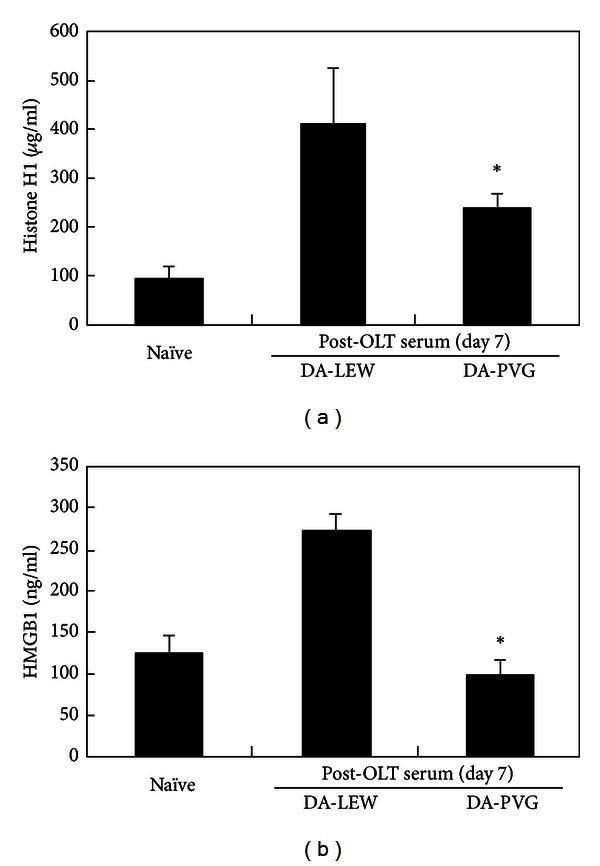
The elevation of circulating histone H1 and HMGB1 during the rejection phase after OLT. The levels of histone H1 and HMGB1 in naive or post-OLT sera were determined by ELISA. For the quantitative determination of histone H1, 0.1 μg of antihistone H1 polyclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) in 100 mM NaHCO3 (pH 9.3) was coated onto a 96-well microtiter plate (Nalge Nunc International, Roskilde, Denmark) by overnight incubation at 4°C. The plate was then blocked with SuperBlock T20 (PBS) Blocking Buffer (Thermo Fisher Scientific Inc., Rockford, IL, USA), and serum samples (each n = 3) (50 μL, ×25 dilution with 10 mM Tris-HCl (pH 8.0), 0.9% (w/v) NaCl, 0.5% (w/v) Tween 20) were added to the wells. Calf thymus histone H1 (Upstate, Charlottesville, VA, USA) was used as a standard. The mixture was incubated at room temperature for 1 hr. Antihistone H1 monoclonal antibody (×500 dilution; Abcam, Cambridge, MA, USA) was then added, and the mixture was incubated at room temperature for 1 hr. Peroxidase-conjugated anti-mouse IgG (×2,000 dilution; Santa Cruz Biotechnology) was then added, and the mixture was incubated at room temperature for 1 hr, followed by the addition of 1-Step Ultra TMB substrate solution (Thermo Fisher Scientific Inc.). For the quantitative determination of HMGB1, a rat HMGB1 ELISA kit (MyBioSource Inc., San Diego, CA, USA) was used according to the manufacturer's protocol. The absorbance (450 nm) was then measured using a Victor X4 Multilabel Plate Reader (PerkinElmer, Shelton, CT, USA). The results are expressed as the mean of three individuals ± SD. *Significantly different compared with the DA-LEW combination (P < 0.01, Student's t-test).
Figure 3.
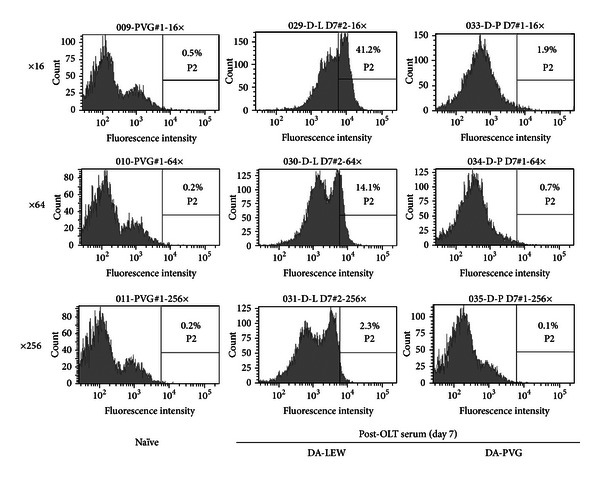
Alloantibody response during the rejection phase after OLT. The alloantibody response was measured by flow cytometry on a single cell suspension of DA rat splenocytes. Briefly, 50 μL aliquots containing 5 × 105 splenocytes was incubated with 50 μL of diluted naive or post-OLT sera (1 : 16, 1 : 64, 1 : 256) for 45 min at 4°C. The washed cells were reacted with 50 μL of a mixture of FITC-conjugated goat antibody specific for the Fc portion of rat IgG (×100 dilution) (Jackson ImmunoResearch Laboratories, West Grove, PA, USA) in PBS containing 1% BSA and 0.02% NaN3. After staining, the cells were washed, fixed, and analyzed using an LSRII flow cytometer (BD Biosciences, San Jose, CA, USA). Histograms (representative of three individuals) show the percentage of DA splenocytes recognized by alloantibody (IgG) in the post-OLT sera at day 7 after OLT.
Figure 4.
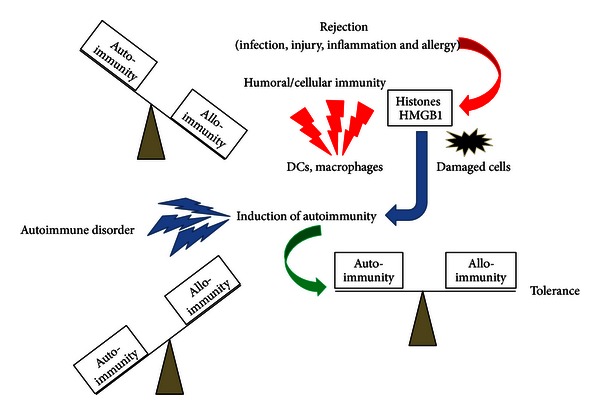
The balance of autoimmunity and alloimmunity (against alloantigens, pathogens, or allergens) is important for immune regulation. During the rejection phase (or when suffering from infection, injury, or allergy), alloimmunity is predominant, and nuclear antigens such as histones and HMGB1 are released from damaged cells, tissues, or organs or are actively secreted from activating immune cells such as DCs and macrophages. The induction of autoimmunity against nuclear antigens (i.e., induction of antinuclear Abregs) may regulate the balance and induce immunological tolerance. Excessive activation of autoimmunity may cause autoimmune disorders.
5. Nuclear Antigens as a Therapeutic Target
To prevent the release of nuclear antigens such as histone H1 and HMGB1, resulting in the activation of innate and adaptive immune responses, several strategies have been proposed. The therapeutic potential of anti-HMGB1 antibody, soluble RAGE, and anti-RAGE neutralizing antibody has been demonstrated in experimental sepsis [73, 74], liver ischemia/reperfusion injury [75], Con-A-induced hepatic injury [76], traumatic brain injury [77], and organ transplantation [78, 79]. The therapeutic potential of antihistone H1 polyclonal antibody for overcoming rejection and liver inflammation was also confirmed by our group [19, 40]. We also confirmed the great potential of histone H1 vaccination in transplant recipients for tolerance induction [80, 81]. To further explore the roles of histone H1 and its future clinical application, we have generated antihistone H1 monoclonal antibodies (clone: 16G9, IgM), which possess immunosuppressive activity in vitro [82]. In addition, we have identified the functional epitope (SSVLYGGPPSAA) responsible for the immunosuppressive activity of 16G9 and have confirmed the diagnostic and therapeutic potential of histone H1 peptide [83]. In addition to neutralizing antibody therapy, an HMGB1 absorption column (polymyxin B-immobilized fibers) has been developed and clinically applied for the removal of circulating HMGB1 in patients with septic shock, acute respiratory distress syndrome, and idiopathic pulmonary fibrosis with acute exacerbation [84–90]. The therapeutic potential of HMGB1 antagonists such as HMGB1 A box peptide has also been reported [91, 92].
6. Summary and Future Directions
In this review, we have discussed the diagnostic and therapeutic potential of nuclear antigens (histone H1 and HMGB1) and the corresponding antinuclear Abregs on infection, injury, inflammation, and transplant rejection. One of the immunosuppressive mechanisms of antinuclear Abregs is the direct binding of circulating nuclear antigens, which triggers the immune response (Figure 1). In addition, our previous study strongly suggested the binding of antihistone H1 Abregs to histone H1-like molecules, which may be transiently expressed on the cell membrane of splenocytes [19]. We have also demonstrated that antihistone H1 Abregs may selectively suppress the MAPK, NF-κB, and calcineurin-NFAT signaling pathways during T-cell activation [40], coordinate the Th1/Th2 balance [81], and induce CD4+CD25+ T cells [65]. Recent evidence suggests that antihistone H1 Abregs negatively regulate the harmful T-cell response, in part through collaboration with Tregs [93]. Although further investigation is needed, the direct effects of nuclear antigens and corresponding antinuclear Abregs on immune cells may play important roles in inflammation, rejection, and tolerance induction. Interestingly, the induction of antinuclear Abregs (i.e., the autoimmune response against nuclear antigens) may suppress alloantibody production during rejection after OLT (Figure 3). A crucial issue is why cell death-associated moieties and corresponding autoantibodies, which elicit clinical autoimmunity in patients with autoimmune diseases, could be indispensable for immune regulation in other settings. In our previous study, nuclear histone H1 and Freund's complete adjuvant were injected into naive rats and resulted in different autoantibody responses against histone H1 in tolerogenic PVG OLT recipients and rejecting LEW OLT recipients [68]. The transient induction of autoantibodies in normal mice challenged with dying cells and adjuvants (Freund's incomplete adjuvant or DCs) was also reported without clinical or histological features of autoimmunity, while clinical autoimmunity develops in autoimmune-prone mice [94, 95]. Therefore, we speculate that the response to dying cells in OLT recipients may be one of the key factors determining the clinical outcome. How to modulate the balance between autoimmunity and alloimmunity is an important issue for the extrinsic regulation of unwanted immune responses and the induction of immune tolerance (Figure 4). Our present data also reveal the diagnostic significance of nuclear antigens for the prediction of acute rejection after liver transplantation (Figure 2). The development of fast, accurate, and precise diagnostic tools by measuring the blood level of nuclear histone H1 and HMGB1 would allow clinicians to evaluate immune status and modulate the dose of immunosuppressants for rejection control. The development of absorption columns for circulating nuclear antigens (histone H1 and HMGB1) as well as neutralizing humanized monoclonal antibodies may help to establish novel immunotherapies for infection, injury, inflammation, and transplant rejection.
Conflict ofInterests
The authors have declared that no competing interests exist.
Acknowledgments
This work was supported in part by Grants from the National Science Council (NSC98-2320-B-182-029-MY3 and NSC101-2320-B-182-037-MY3 to T. Nakano; NSC98-2314-B-182A-058-MY3 and NSC101-2314-B-182A-031-MY3 to C.-L. Chen.) and the Chang Gung Memorial Hospital (CMRPD891671 and CMRPD8A0701 to T. Nakano.; CMRPG891061/2 to C.-L. Chen.) of Taiwan.
Abbreviations
- HMGB1:
High-mobility group box 1
- DA:
Dark Agouti
- PVG:
Piebald Virol Glaxo
- OLT:
Orthotopic liver transplantation
- DCs:
Dendritic cells
- LSF:
Liver suppressor factor
- AMR:
Antibody-mediated rejection
- Tregs:
Regulatory T cells
- Con A:
Concanavalin A
- DAMPs:
Damage-associated molecular pattern molecules
- RAGE:
Receptor for advanced glycation endproduct
- TLR:
Toll-like receptor
- NSCLC:
Nonsmall cell lung cancer.
References
- 1.Haeney M. The immunological background to transplantation. Journal of Antimicrobial Chemotherapy. 1995;36(5):1–9. doi: 10.1093/jac/36.suppl_b.1. [DOI] [PubMed] [Google Scholar]
- 2.Starzl TE. History of clinical transplantation. World Journal of Surgery. 2000;24(7):759–782. doi: 10.1007/s002680010124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Calne RY, Rolles K, White DJ, et al. Cyclosporin A initially as the only immunosuppressant in 34 recipients of cadaveric organs: 32 kidneys, 2 pancreases, and 2 livers. The Lancet. 1979;2(8151):1033–1036. doi: 10.1016/s0140-6736(79)92440-1. [DOI] [PubMed] [Google Scholar]
- 4.Platz KP, Mueller AR, Blumhardt G, et al. Nephrotoxicity following orthotopic liver transplantation: a comparison between cyclosporine and FK506. Transplantation. 1994;58(2):170–178. [PubMed] [Google Scholar]
- 5.Miller LW. Cardiovascular toxicities of immunosuppressive agents. The American Journal of Transplantation. 2002;2(9):807–818. doi: 10.1034/j.1600-6143.2002.20902.x. [DOI] [PubMed] [Google Scholar]
- 6.Hojo M, Morimoto T, Maluccio M, et al. Cyclosporine induces cancer progression by a cell-autonomous mechanism. Nature. 1999;397(6719):530–534. doi: 10.1038/17401. [DOI] [PubMed] [Google Scholar]
- 7.Calne RY, Sells RA, Pena JR, et al. Induction of immunological tolerance by porcine liver allografts. Nature. 1969;223(5205):472–476. doi: 10.1038/223472a0. [DOI] [PubMed] [Google Scholar]
- 8.Crispe IN. Hepatic T cells and liver tolerance. Nature Reviews Immunology. 2003;3(1):51–62. doi: 10.1038/nri981. [DOI] [PubMed] [Google Scholar]
- 9.Kamada N, Davies HS, Roser B. Reversal of transplantation immunity by liver grafting. Nature. 1981;292(5826):840–842. doi: 10.1038/292840a0. [DOI] [PubMed] [Google Scholar]
- 10.Kamada N, Wight DGD. Antigen-specific immunosuppression induced by liver transplantation in the rat. Transplantation. 1984;38(3):217–221. doi: 10.1097/00007890-198409000-00004. [DOI] [PubMed] [Google Scholar]
- 11.Kamada N. Transfer of specific immunosuppression of graft rejection using lymph from tolerant liver-grafted rats. Immunology. 1985;55(2):241–247. [PMC free article] [PubMed] [Google Scholar]
- 12.Doherty D, O'Farrelly C. Lymphocyte repertoires in healthy liver. In: Gershwin ME, Vierling JM, Manns MP, editors. Liver Immunology. Philadelphia, Pa, USA: Hanley & Belfus; 2003. pp. 31–46. [Google Scholar]
- 13.Nakano T, Lai CY, Goto S, et al. Immunological and regenerative aspects of hepatic mast cells in liver allograft rejection and tolerance. PLoS ONE. 2012;7(5) doi: 10.1371/journal.pone.0037202.e37202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sumimoto R, Kamada N. Specific suppression of allograft rejection by soluble class I antigen and complexes with monoclonal antibody. Transplantation. 1990;50(4):678–682. doi: 10.1097/00007890-199010000-00029. [DOI] [PubMed] [Google Scholar]
- 15.Tsurufuji M, Ishiguro K, Shinomiya T, Uchida T, Kamada N. Immunosuppressive activity of serum from liver-grafted rats: in vitro specific inhibition of mixed lymphocyte reactivity by antibodies against class II RT1 alloantigens. Immunology. 1987;61(4):421–428. [PMC free article] [PubMed] [Google Scholar]
- 16.Lord R, Kamada N, Kobayashi E, Goto S, Sunagawa M. Isolation of a 40 kDa immunoinhibitory protein induced by rat liver transplantation. Transplant Immunology. 1995;3(2):174–179. doi: 10.1016/0966-3274(95)80045-x. [DOI] [PubMed] [Google Scholar]
- 17.Edwards-Smith C, Goto S, Lord R, Shimizu Y, Vari F, Kamada N. Allograft acceptance and rejection, mediated by a liver suppressor factor, LSF-1, purified from serum of liver transplanted rats. Transplant Immunology. 1996;4(4):287–292. doi: 10.1016/s0966-3274(96)80049-9. [DOI] [PubMed] [Google Scholar]
- 18.Goto S, Lord R, Kobayashi E, Vari F, Edwards-Smith C, Kamada N. Novel immunosuppressive proteins purified from the serum of liver-retransplanted rats. Transplantation. 1996;61(8):1147–1151. doi: 10.1097/00007890-199604270-00004. [DOI] [PubMed] [Google Scholar]
- 19.Nakano T, Kawamoto S, Lai CY, et al. Liver transplatation-induced antihistone H1 autoantibodies suppress mixed lymphocyte reaction. Transplantation. 2004;77(10):1595–1603. doi: 10.1097/01.tp.0000123079.10650.71. [DOI] [PubMed] [Google Scholar]
- 20.Nakano T, Kawamoto S, Lai CY, et al. Characterization of immunosuppressive factors expressed in serum by rat tolerogenic liver transplantation. Transplantation Proceedings. 2005;37(1):80–81. doi: 10.1016/j.transproceed.2004.12.290. [DOI] [PubMed] [Google Scholar]
- 21.Nakano T, Goto S, Lai CY, et al. Experimental and clinical significance of antinuclear antibodies in liver transplantation. Transplantation. 2007;83(8):1122–1125. doi: 10.1097/01.tp.0000258646.54562.c7. [DOI] [PubMed] [Google Scholar]
- 22.Nakano T, Lai CY, Goto S, et al. Role of antinuclear antibodies in experimental and clinical liver transplantation. Transplantation Proceedings. 2006;38(10):3605–3606. doi: 10.1016/j.transproceed.2006.10.076. [DOI] [PubMed] [Google Scholar]
- 23.Win TS, Pettigrew GJ. Humoral autoimmunity and transplant vasculopathy: when allo is not enough. Transplantation. 2010;90(2):113–120. doi: 10.1097/TP.0b013e3181e25a59. [DOI] [PubMed] [Google Scholar]
- 24.Kirk AD, Turgeon NA, Iwakoshi NN. B cells and transplantation tolerance. Nature Reviews Nephrology. 2010;6(10):584–593. doi: 10.1038/nrneph.2010.111. [DOI] [PubMed] [Google Scholar]
- 25.Sureshkumar KK, Hussain SM, Carpenter BJ, Sandroni SE, Marcus RJ. Antibody-mediated rejection following renal transplantation. Expert Opinion on Pharmacotherapy. 2007;8(7):913–921. doi: 10.1517/14656566.8.7.913. [DOI] [PubMed] [Google Scholar]
- 26.Egawa H, Ohdan H, Haga H, et al. Current status of liver transplantation across ABO blood-type barrier. Journal of Hepato-Biliary-Pancreatic Surgery. 2008;15(2):131–138. doi: 10.1007/s00534-007-1298-2. [DOI] [PubMed] [Google Scholar]
- 27.Kantor AB, Herzenberg LA. Origin of murine B cell lineages. Annual Review of Immunology. 1993;11:501–538. doi: 10.1146/annurev.iy.11.040193.002441. [DOI] [PubMed] [Google Scholar]
- 28.Neron S, Lemieux R. Type 2 T-cell-independent murine immune response to the human ABO blood group antigens. Vox Sanguinis. 1994;67(1):68–74. doi: 10.1111/j.1423-0410.1994.tb05041.x. [DOI] [PubMed] [Google Scholar]
- 29.Néron S, Lemieux R. CD5+ B cell-dependent regulation of the murine T-cell independent immune response against the human blood group A antigen. Immunological Investigations. 1997;26(5-7):631–647. doi: 10.3109/08820139709088546. [DOI] [PubMed] [Google Scholar]
- 30.Montecino-Rodriguez E, Dorshkind K. New perspectives in B-1 B cell development and function. Trends in Immunology. 2006;27(9):428–433. doi: 10.1016/j.it.2006.07.005. [DOI] [PubMed] [Google Scholar]
- 31.Tung JW, Herzenberg LA. Unraveling B-1 progenitors. Current Opinion in Immunology. 2007;19(2):150–155. doi: 10.1016/j.coi.2007.02.012. [DOI] [PubMed] [Google Scholar]
- 32.Matsushita T, Yanaba K, Bouaziz JD, Fujimoto M, Tedder TF. Regulatory B cells inhibit EAE initiation in mice while other B cells promote disease progression. Journal of Clinical Investigation. 2008;118(10):3420–3430. doi: 10.1172/JCI36030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yanaba K, Bouaziz JD, Haas KM, Poe JC, Fujimoto M, Tedder TF. A regulatory B cell subset with a unique CD1dhiCD5+ phenotype controls T cell-dependent inflammatory responses. Immunity. 2008;28(5):639–650. doi: 10.1016/j.immuni.2008.03.017. [DOI] [PubMed] [Google Scholar]
- 34.Moritoki Y, Zhang W, Tsuneyama K, et al. B cells suppress the inflammatory response in a mouse model of primary biliary cirrhosis. Gastroenterology. 2009;136(3):1037–1047. doi: 10.1053/j.gastro.2008.11.035. [DOI] [PubMed] [Google Scholar]
- 35.Chhabra P, Schlegel K, Okusa MD, Lobo PI, Brayman KL. Naturally occurring immunoglobulin M (nIgM) autoantibodies prevent autoimmune diabetes and mitigate inflammation after transplantation. Annals of Surgery. 2012;256(4):634–641. doi: 10.1097/SLA.0b013e31826b4ba9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Richter A, Grabhorn E, Helmke K, Manns MP, Ganschow R, Burdelski M. Clinical relevance of autoantibodies after pediatric liver transplantation. Clinical Transplantation. 2007;21(3):427–432. doi: 10.1111/j.1399-0012.2007.00667.x. [DOI] [PubMed] [Google Scholar]
- 37.Kerkar N, Hadzic N, Davies ET, et al. De-novo autoimmune hepatitis after liver transplantation. The Lancet. 1998;351(9100):409–413. doi: 10.1016/S0140-6736(97)06478-7. [DOI] [PubMed] [Google Scholar]
- 38.Mieli-Vergani G, Vergani D. De novo autoimmune hepatitis after liver transplantation. Journal of Hepatology. 2004;40(1):3–7. doi: 10.1016/j.jhep.2003.10.022. [DOI] [PubMed] [Google Scholar]
- 39.Czaja AJ. Diagnosis, pathogenesis, and treatment of autoimmune hepatitis after liver transplantation. Digestive Diseases and Sciences. 2012;57(9):2248–2266. doi: 10.1007/s10620-012-2179-3. [DOI] [PubMed] [Google Scholar]
- 40.Nakano T, Goto S, Lai CY, et al. Immunological aspects and therapeutic significance of an autoantibody against histone H1 in a rat model of concanavalin A-induced hepatitis. Immunology. 2010;129(4):547–555. doi: 10.1111/j.1365-2567.2009.03149.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wu Y, Cai B, Tang J, Bai Y, Wang L. Tacrolimus may induce the production of nucleolar anti-nuclear antibody in liver transplant patients. Journal of Gastrointestinal and Liver Diseases. 2011;20(3):267–270. [PubMed] [Google Scholar]
- 42.Barnay-Verdier S, Fattoum L, Borde C, Kaveri S, Gibot S, Maréchal V. Emergence of autoantibodies to HMGB1 is associated with survival in patients with septic shock. Intensive Care Medicine. 2011;37(6):957–962. doi: 10.1007/s00134-011-2192-6. [DOI] [PubMed] [Google Scholar]
- 43.Gujral JS, Bucci TJ, Farhood A, Jaeschke H. Mechanism of cell death during warm hepatic ischemia-reperfusion in rats: apoptosis or necrosis? Hepatology. 2001;33(2):397–405. doi: 10.1053/jhep.2001.22002. [DOI] [PubMed] [Google Scholar]
- 44.Cursio R. Caspase inhibition in liver transplantation: from basic research to clinical studies. HPB. 2010;12(1):1–3. doi: 10.1111/j.1477-2574.2009.00123.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gotoh K, Lu Z, Morita M, et al. Participation of autophagy in the initiation of graft dysfunction after rat liver transplantation. Autophagy. 2009;5(3):351–360. doi: 10.4161/auto.5.3.7650. [DOI] [PubMed] [Google Scholar]
- 46.Lotze MT, Tracey KJ. High-mobility group box 1 protein (HMGB1): nuclear weapon in the immune arsenal. Nature Reviews Immunology. 2005;5(4):331–342. doi: 10.1038/nri1594. [DOI] [PubMed] [Google Scholar]
- 47.Wang H, Bloom O, Zhang M, et al. HMG-1 as a late mediator of endotoxin lethality in mice. Science. 1999;285(5425):248–251. doi: 10.1126/science.285.5425.248. [DOI] [PubMed] [Google Scholar]
- 48.Abraham E, Arcaroli J, Carmody A, Wang H, Tracey KJ. Cutting edge: HMG-1 as a mediator of acute lung inflammation. Journal of Immunology. 2000;165(6):2950–2954. doi: 10.4049/jimmunol.165.6.2950. [DOI] [PubMed] [Google Scholar]
- 49.Degryse B, Bonaldi T, Scaffidi P, et al. The high mobility group (HMG) boxes of the nuclear protein HMG1 induce chemotaxis and cytoskeleton reorganization in rat smooth muscle cells. The Journal of Cell Biology. 2001;152(6):1197–1206. doi: 10.1083/jcb.152.6.1197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Yasuda T, Ueda T, Takeyama Y, et al. Significant increase of serum high-mobility group box chromosomal protein 1 levels in patients with severe acute pancreatitis. Pancreas. 2006;33(4):359–363. doi: 10.1097/01.mpa.0000236741.15477.8b. [DOI] [PubMed] [Google Scholar]
- 51.Goldstein RS, Bruchfeld A, Yang L, et al. Cholinergic anti-inflammatory pathway activity and high mobility group box-1 (HMGB1) serum levels in patients with rheumatoid arthritis. Molecular Medicine. 2007;13(3-4):210–215. doi: 10.2119/2006-00108.Goldstein. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hamada N, Maeyama T, Kawaguchi T, et al. The role of high mobility group box1 in pulmonary fibrosis. The American Journal of Respiratory Cell and Molecular Biology. 2008;39(4):440–447. doi: 10.1165/rcmb.2007-0330OC. [DOI] [PubMed] [Google Scholar]
- 53.Muhammad S, Barakat W, Stoyanov S, et al. The HMGB1 receptor RAGE mediates ischemic brain damage. The Journal of Neuroscience. 2008;28(46):12023–12031. doi: 10.1523/JNEUROSCI.2435-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hoshina T, Kusuhara K, Ikeda K, Mizuno Y, Saito M, Hara T. High mobility group box 1 (HMGB1) and macrophage migration inhibitory factor (MIF) in Kawasaki disease. Scandinavian Journal of Rheumatology. 2008;37(6):445–449. doi: 10.1080/03009740802144143. [DOI] [PubMed] [Google Scholar]
- 55.Kaczorowski DJ, Nakao A, Vallabhaneni R, et al. Mechanisms of toll-like receptor 4 (TLR4)-mediated inflammation after cold ischemia/reperfusion in the heart. Transplantation. 2009;87(10):1455–1463. doi: 10.1097/TP.0b013e3181a36e5e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Albayrak A, Uyanik MH, Cerrah S, et al. Is HMGB1 a new indirect marker for revealing fibrosis in chronic hepatitis and a new therapeutic target in treatment? Viral Immunology. 2010;23(6):633–638. doi: 10.1089/vim.2010.0080. [DOI] [PubMed] [Google Scholar]
- 57.Kohno T, Anzai T, Shimizu H, et al. Impact of serum high-mobility group box 1 protein elevation on oxygenation impairment after thoracic aortic aneurysm repair. Heart and Vessels. 2011;26(3):306–312. doi: 10.1007/s00380-010-0056-6. [DOI] [PubMed] [Google Scholar]
- 58.Takahata R, Ono S, Tsujimoto H, et al. Postoperative serum concentrations of high mobility group box chromosomal protein-1 correlates to the duration of SIRS and pulmonary dysfunction following gastrointestinal surgery. The Journal of Surgical Research. 2011;170(1):135–140. doi: 10.1016/j.jss.2011.04.040. [DOI] [PubMed] [Google Scholar]
- 59.Choi J, Min HJ, Shin JS. Increased levels of HMGB1 and pro-inflammatory cytokines in children with febrile seizures. Journal of Neuroinflammation. 2011;8(article 135) doi: 10.1186/1742-2094-8-135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Haraba R, Suica VI, Uyy E, Ivan L, Antohe F. Hyperlipidemia stimulates the extracellular release of the nuclear high mobility group box 1 protein. Cell and Tissue Research. 2011;346(3):361–368. doi: 10.1007/s00441-011-1277-4. [DOI] [PubMed] [Google Scholar]
- 61.Naruse K, Sado T, Noguchi T, et al. Peripheral RAGE (receptor for advanced glycation endproducts)-ligands in normal pregnancy and preeclampsia: novel markers of inflammatory response. Journal of Reproductive Immunology. 2012;93(2):69–74. doi: 10.1016/j.jri.2011.12.003. [DOI] [PubMed] [Google Scholar]
- 62.Zhou RR, Liu HB, Peng JP, et al. High mobility group box chromosomal protein 1 in acute-on-chronic liver failure patients and mice with ConA-induced acute liver injury. Experimental and Molecular Pathology. 2012;93(2):213–219. doi: 10.1016/j.yexmp.2012.05.006. [DOI] [PubMed] [Google Scholar]
- 63.Konishi A, Shimizu S, Hirota J, et al. Involvement of histone H1.2 in apoptosis induced by DNA double-strand breaks. Cell. 2003;114(6):673–688. doi: 10.1016/s0092-8674(03)00719-0. [DOI] [PubMed] [Google Scholar]
- 64.Gabrilovich DI, Cheng P, Fan Y, et al. H1° histone and differentiation of dendritic cells. A molecular target for tumor-derived factors. Journal of Leukocyte Biology. 2002;72(2):285–296. [PubMed] [Google Scholar]
- 65.Hsu LW, Chen CL, Nakano T, et al. The role of a nuclear protein, histone H1, on signalling pathways for the maturation of dendritic cells. Clinical and Experimental Immunology. 2008;152(3):576–584. doi: 10.1111/j.1365-2249.2008.03652.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Rovere-Querini P, Capobianco A, Scaffidi P, et al. HMGB1 is an endogenous immune adjuvant released by necrotic cells. EMBO Reports. 2004;5(8):825–830. doi: 10.1038/sj.embor.7400205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Allam R, Scherbaum CR, Darisipudi MN, et al. Histones from dying renal cells aggravate kidney injury via TLR2 and TLR4. Journal of the American Society of Nephrology. 2012;23(8):1375–1388. doi: 10.1681/ASN.2011111077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Nakano T, Goto S, Lai CY, et al. Involvement of autoimmunity against nuclear histone H1 in liver transplantation tolerance. Transplant Immunology. 2008;19(2):87–92. doi: 10.1016/j.trim.2008.04.001. [DOI] [PubMed] [Google Scholar]
- 69.Hatada T, Wada H, Nobori T, et al. Plasma concentrations and importance of high mobility group box protein in the prognosis of organ failure in patients with disseminated intravascular coagulation. Thrombosis and Haemostasis. 2005;94(5):975–979. doi: 10.1160/TH05-05-0316. [DOI] [PubMed] [Google Scholar]
- 70.Shang GH, Jia CQ, Tian H, et al. Serum high mobility group box protein 1 as a clinical marker for non-small cell lung cancer. Respiratory Medicine. 2009;103(12):1949–1953. doi: 10.1016/j.rmed.2009.05.019. [DOI] [PubMed] [Google Scholar]
- 71.Hashimoto T, Ishii J, Kitagawa F, et al. Circulating high-mobility group box 1 and cardiovascular mortality in unstable angina and non-ST-segment elevation myocardial infarction. Atherosclerosis. 2012;221(2):490–495. doi: 10.1016/j.atherosclerosis.2012.01.040. [DOI] [PubMed] [Google Scholar]
- 72.Wang KY, Yu GF, Zhang ZY, Huang Q, Dong XQ. Plasma high-mobility group box 1 levels and prediction of outcome in patients with traumatic brain injury. Clinica Chimica Acta. 2012;413(21-22):1737–1741. doi: 10.1016/j.cca.2012.07.002. [DOI] [PubMed] [Google Scholar]
- 73.Andersson U, Erlandsson-Harris H, Yang H, Tracey KJ. HMGB1 as a DNA-binding cytokine. Journal of Leukocyte Biology. 2002;72(6):1084–1091. [PubMed] [Google Scholar]
- 74.Wang H, Ward MF, Sama AE. Novel HMGB1-inhibiting therapeutic agents for experimental sepsis. Shock. 2009;32(4):348–357. doi: 10.1097/SHK.0b013e3181a551bd. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Tsung A, Sahai R, Tanaka H, et al. The nuclear factor HMGB1 mediates hepatic injury after murine liver ischemia-reperfusion. Journal of Experimental Medicine. 2005;201(7):1135–1143. doi: 10.1084/jem.20042614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Gong Q, Zhang H, Li JH, et al. High-mobility group box 1 exacerbates concanavalin A-induced hepatic injury in mice. Journal of Molecular Medicine. 2010;88(12):1289–1298. doi: 10.1007/s00109-010-0681-7. [DOI] [PubMed] [Google Scholar]
- 77.Okuma Y, Liu K, Wake H, et al. Anti-high mobility group box-1 antibody therapy for traumatic brain injury. Annals of Neurology. 2012;72(3):373–384. doi: 10.1002/ana.23602. [DOI] [PubMed] [Google Scholar]
- 78.Moser B, Szabolcs MJ, Ankersmit HJ, et al. Blockade of RAGE suppresses alloimmune reactions in vitro and delays allograft rejection in murine heart transplantation. The American Journal of Transplantation. 2007;7(2):293–302. doi: 10.1111/j.1600-6143.2006.01617.x. [DOI] [PubMed] [Google Scholar]
- 79.Duan L, Wang CY, Chen J, et al. High-mobility group box 1 promotes early acute allograft rejection by enhancing IL-6-dependent Th17 alloreactive response. Laboratory Investigation. 2011;91(1):43–53. doi: 10.1038/labinvest.2010.141. [DOI] [PubMed] [Google Scholar]
- 80.Nakano T, Ono K, Goto S, et al. Histone H1 vaccine therapy for overcoming acute rejection in experimental organ transplantation. Transplantation Proceedings. 2006;38(10):3247–3248. doi: 10.1016/j.transproceed.2006.10.078. [DOI] [PubMed] [Google Scholar]
- 81.Nakano T, Goto S, Lai CY, et al. Impact of vaccine therapy using nuclear histone H1 on allograft survival in experimental organ transplantation. Transplant Immunology. 2007;17(3):147–152. doi: 10.1016/j.trim.2007.01.003. [DOI] [PubMed] [Google Scholar]
- 82.Shimada Y, Goto T, Kawamoto S, et al. Development of a two-step chromatography procedure that allows the purification of a high-purity anti-histone H1 monoclonal immunoglobulin M antibody with immunosuppressant activity. Biomedical Chromatography. 2008;22(1):13–19. doi: 10.1002/bmc.887. [DOI] [PubMed] [Google Scholar]
- 83.Chiang KC, Shimada Y, Nakano T, et al. A novel peptide mimotope identified as a potential immunosuppressive vaccine for organ transplantation. Journal of Immunology. 2009;182(7):4282–4288. doi: 10.4049/jimmunol.0800641. [DOI] [PubMed] [Google Scholar]
- 84.Sakamoto Y, Mashiko K, Matsumoto H, et al. Effect of direct hemoperfusion with a polymyxin B immobilized fiber column on high mobility group box-1 (HMGB-1) in severe septic shock: report of a case. ASAIO Journal. 2006;52(6):e37–e39. doi: 10.1097/01.mat.0000248996.22865.ce. [DOI] [PubMed] [Google Scholar]
- 85.Sakamoto Y, Mashiko K, Matsumoto H, Hara Y, Kutsukata N, Yamamoto Y. Relationship between effect of polymyxin B-immobilized fiber and high-mobility group box-1 protein in septic shock patients. ASAIO Journal. 2007;53(3):324–328. doi: 10.1097/MAT.0b013e3180340301. [DOI] [PubMed] [Google Scholar]
- 86.Sakamoto Y, Mashiko K, Obata T, et al. Clinical responses and improvement of some laboratory parameters following polymyxin B-immobilized fiber treatment in septic shock. ASAIO Journal. 2007;53(5):646–650. doi: 10.1097/MAT.0b013e3181492395. [DOI] [PubMed] [Google Scholar]
- 87.Enomoto N, Suda T, Uto T, et al. Possible therapeutic effect of direct haemoperfusion with a polymyxin B immobilized fibre column (PMX-DHP) on pulmonary oxygenation in acute exacerbations of interstitial pneumonia. Respirology. 2008;13(3):452–460. doi: 10.1111/j.1440-1843.2008.01290.x. [DOI] [PubMed] [Google Scholar]
- 88.Nakamura T, Fujiwara N, Sato E, et al. Effect of polymyxin B-immobilized fiber hemoperfusion on serum high mobility group box-1 protein levels and oxidative stress in patients with acute respiratory distress syndrome. ASAIO Journal. 2009;55(4):395–399. doi: 10.1097/MAT.0b013e3181a5290f. [DOI] [PubMed] [Google Scholar]
- 89.Yamamoto T, Ono T, Ito T, Yamanoi A, Maruyama I, Tanaka T. Hemoperfusion with a high-mobility group box 1 adsorption column can prevent the occurrence of hepatic ischemia-reperfusion injury in rats. Critical Care Medicine. 2010;38(3):879–885. doi: 10.1097/CCM.0b013e3181c58951. [DOI] [PubMed] [Google Scholar]
- 90.Abe S, Hayashi H, Seo Y, et al. Reduction in serum high mobility group box-1 level by polymyxin B-immobilized fiber column in patients with idiopathic pulmonary fibrosis with acute exacerbation. Blood Purification. 2011;32(4):310–316. doi: 10.1159/000330325. [DOI] [PubMed] [Google Scholar]
- 91.Zhang CL, Shu MG, Qi HW, Li LW. Inhibition of tumor angiogenesis by HMGB1 A box peptide. Medical Hypotheses. 2008;70(2):343–345. doi: 10.1016/j.mehy.2007.05.024. [DOI] [PubMed] [Google Scholar]
- 92.Li Y, Gong W, Zhang L, et al. Expression and purification of the fusion protein HMGB1Abox-TMD1, a novel HMGB1 antagonist. Biochemistry. 2010;75(4):466–471. doi: 10.1134/s0006297910040103. [DOI] [PubMed] [Google Scholar]
- 93.Takaoka Y, Kawamoto S, Katayama A, et al. Unexpected T cell regulatory activity of anti-histone H1 autoantibody: its mode of action in regulatory T cell-dependent and -independent manners. Biochemical and Biophysical Research Communications. 2013;431(2):246–252. doi: 10.1016/j.bbrc.2012.12.125. [DOI] [PubMed] [Google Scholar]
- 94.Bondanza A, Zimmermann VS, Dell’Antonio G, et al. Cutting edge: dissociation between autoimmune response and clinical disease after vaccination with dendritic cells. Journal of Immunology. 2003;170(1):24–27. doi: 10.4049/jimmunol.170.1.24. [DOI] [PubMed] [Google Scholar]
- 95.Bondanza A, Zimmermann VS, Dell’Antonio G, et al. Requirement of dying cells and environmental adjuvants for the induction of autoimmunity. Arthritis and Rheumatism. 2004;50(5):1549–1560. doi: 10.1002/art.20187. [DOI] [PubMed] [Google Scholar]