

Abstract
Visual search plays an important role in guiding behavior. Children have more difficulty performing conjunction search tasks than adults. The present research evaluates whether developmental differences in children's ability to organize serial visual search (i.e., search organization skills) contribute to performance limitations in a typical conjunction search task. We evaluated 134 children between the ages of 2 and 17 on separate tasks measuring search for targets defined by a conjunction of features or by distinct features. Our results demonstrated that children organize their visual search better as they get older. As children's skills at organizing visual search improve they become more accurate at locating targets with conjunction of features amongst distractors, but not for targets with distinct features. Developmental limitations in children's abilities to organize their visual search of the environment are an important component of poor conjunction search in young children. In addition, our findings provide preliminary evidence that, like other visuospatial tasks, exposure to reading may influence children's spatial orientation to the visual environment when performing a visual search.
Keywords: visual search, search organization, executive function, normal development, search orientation, conjunction search
1. Introduction
Many of us can recall misplacing a well-used item in our childhood, perhaps a favored toy or a missing article of clothing. For example, consider a small child that has lost one of his shoes. His mother instructs him to go to his room and find the missing shoe. No matter how hard he looks, he cannot find his missing shoe. Concluding his unsuccessful search of the shoe's whereabouts, he tells his mother, “I can't find it, I looked everywhere.” Despite his assertion to the contrary, his mother insists that the shoe is located in his room. To his surprise, his mother quickly locates the missing shoe after briefly searching his room. From children finding their lost shoe in their messy room to adults locating their car in a crowded parking lot, visual search plays an important role in guiding behavior. As in our example, young children have more difficulty than their older counterparts with visual search (Donnelly et al., 2007; Trick & Enns, 1998). However, the underlying developmental processes responsible for poor visual search in children remain unclear. In the present paper, we investigate the development of children's ability to organize their visual search of the environment (search organization skills) and assess whether developmental changes in search organization skills contribute to developmental limitations in children's visual search accuracy.
1.1 Visual search
Visual searches can be directed at targets that have distinct features or are made of conjunctions of features (e.g., Duncan & Humphreys, 1989; Treisman & Gelade, 1980; Trick & Enns, 1998). In feature search, distinct low-level object features “pop-out” when a unique object is amidst distractors (e.g., Figure 1a; Treisman & Gelade, 1980). The distinct low-level perceptual features of the target, relative to distractors, can be registered, coded, and processed in parallel across the visual field, resulting in quick location of the target (Treisman & Gelade, 1980). In contrast, when targets and distractors share common features (e.g., Figure 1b), parallel processing of the environment is insufficient (for an alternative view, Guided Search, see Wolfe, Cave, & Franzel, 1989). Rather, conjunction search requires participants to search serially (i.e., from object to object) the visual environment for the target containing the conjunction of features distinguishing it from distractors (Duncan & Humphreys, 1989).
Figure 1.

Behavioral tasks. a) An example of the feature search task, b) an example of the conjunction search task, c) the apple cancellation test. Participants were instructed to search for the red circle in both Feature and Conjunction Search Tasks. Participants were instructed to cancel (or mark out) each of the apples in the apple cancellation test.
Behaviorally, feature searches are quicker and more accurate than conjunction search (e.g., Carrasco, et al., 2006; Gibson & Jiang, 1998; Treisman & Gelade, 1980; Treisman & Sato, 1990; etc.). Feature search relies on feature-based mechanisms of attention to quickly cue attention to unique features in the visual environment (for a review see Carrasco, 2011). In contrast, conjunction search relies upon moving spatial attention between locations and perceptually binding two or more features, as well as working memory processes that assist in remembering locations previously searched (Bernstein & Robertson, 1998; Humphreys et al., 2000; Luria & Vogel, 2011; Robertson, 2003; Treisman & Gelade, 1980).
Although both forms of visual search can fail, the ability to accurately search the visual environment serially significantly improves from childhood to adolescence, peaking in young adulthood (Donnelly et al., 2007; Thompson & Massaro, 1989; Trick & Enns, 1998). In contrast, most studies do not report changes from childhood to adolescence in feature search performance (Gerhardstein & Rovee-Collier; Thompson & Massaro, 1989; Trick & Enns, 1998, also see Donnelly et al., 2007 for an alternative discussion). Consistent with our `lost shoe' example, Donnelly and colleagues (2007) found that 6–7 year olds performed more poorly than adults on a conjunction search task. Furthermore, in a cross sectional study across the lifespan (ages: 6, 8, 10, 22, & 72), Trick and Enns (1998) also demonstrated that young children perform conjunction search tasks less accurately than young adults and seniors. Both Donnelly et al (2007) and Trick and Enns (1998) propose that age-related effects on conjunction search tasks result from children's' inability to appropriately plan and execute an organized serial search of the environment. Unfortunately, the cognitive processes underlying children's ability to plan and execute organized visual search remain unclear. Furthermore, the pattern of development for this skill, organizing visual search, remains uncharted.
1.2 Executive function and search organization
The ability to plan and execute an organized pattern of behavior is most often associated with “executive functions.” The term `executive function' refers to complex cognitive processing that requires coordination of several sub-processes to adapt behavior to the demands of the environment (Elliot, 2003; Funahashi, 2001). These sub-processes include motor/action planning, working memory, inhibitory control, and mental flexibility. The dorsolateral prefrontal cortex (DLPFC) is a critical brain structure affiliated with maturation of executive functions (Baird et al., 2002; Diamond & Goldman-Rakic, 1989; Elliot, 2003; Funahashi, 2001; Moriguchi & Hiraki, 2009). Executive functions instantiated in the DLPFC continue developing well into young adulthood, fully maturating around the mid-twenties (e.g., Baird et al., 2002; Diamond & Goldman-Rakic, 1989; Geidd, 2004; Geidd, et al., 1999; Lenroot & Geidd, 2006; Moriguchi & Hiraki, 2009; Sowell et al., 2001).
Executive processes play important roles in several aspects of conjunction search. For instance, working memory is important for preventing return to a previously searched location, and plays a key role in guiding spatial attention in conjunction search tasks (Boot et al., 2004; Emrich et al., 2009; Peterson et al., 2001 & 2007). Furthermore, processes like motor/action planning and inhibitory control contribute to the generation, initiation, and inhibition of goal-directed behaviors, an important aspect of serial search (e.g., Muggleton et al., 2010). Developmental limitations in working memory and other executive processes do not influence feature search performance, a task relying on lower-level mechanisms of vision and feature-based attention. In contrast, limitations in executive processes, like working memory, may severely hamper children's abilities to plan and execute an organized serial search of the environment (Donnelley et al., 2007; Han & Kim, 2004; Luria & Vogel, 2011; Trick & Enns, 1998). However, the development of children's abilities to organize serial visual search behavior, a skill we will refer to as `search organization,' remains relatively unexplored.
Studies typically infer search organization based on the relationship between reaction time and the number of distractors in the visual environment (i.e., search slope) – as the number of distractors in a conjunction search task increases, reaction time also increases (i.e., a steep search slope; Duncan & Humphreys, 1989; Treisman & Gelade, 1980; Wolfe, Cave, & Franzel, 1989). In contrast, the number of distractors does not significantly influence reaction time on feature search tasks (i.e., a flat search slope; e.g., Treisman & Gelade, 1980). Steeper conjunction search slopes signify more disorganized, or inefficient, search as seen in children compared to adults (Donnelly et al, 2007). Search slope methods in visual search, however, do not necessarily tap exclusively into search organization skills (Trick & Enns, 1998), as reaction time is an indirect marker of information processing speed associated with many aspects of human behavior (e.g., general cognitive function, aging, quality of living, etc.; Deary & Der, 2005; Jakobsen et al., 2011). Furthermore, general age-related improvements in processing speed from childhood to late adolescence make the interpretation of reaction time data unclear for cross-sectional developmental studies (e.g., Anderson et al., 1984; Coyle, et al., 2011; Philip, 1934). Visual search organization measurements unrelated to reaction time would add to understanding of the development of children's search organization skills and how search organization influences visual search performance.
Accuracy is a measure of visual search performance, but is uninformative in adults because of near ceiling performance (i.e., 100%). However, visual search accuracy, particularly for conjunction search, is variable in children and provides an informative measure of visual search performance. Mark and colleagues recently demonstrated a valid non-reaction time based measure of search organization skills in healthy adults and adult patients with brain injury (Mark & Woods, 2003; Mark et al., 2004; Woods & Mark, 2005 & 2007; Woods et al., 2004). Mark and Woods administered a cancellation task that required participants to locate and mark out (or cancel) 56 identical target objects among a field of 127 pseudo-randomly arrayed distractors. While participants cancelled targets, the investigators recorded the order that targets were marked (the cancellation path) and calculated three objective measures of participants' visual search organization skills. In brief, the three measures assess the quality of search organization by quantifying the length of the cancellation path, the number of times the cancellation path intersects itself, and the net orthogonal movement pattern of the path (see Methods for detailed description). Whereas, healthy adults demonstrate highly organized search for all three measures, patients with executive dysfunction demonstrate disorganized search performance on all three measures (Mark & Woods, 2003; Mark et al., 2004; Woods & Mark, 2005 & 2007; Woods et al., 2004). These data demonstrate that search organization measures on cancellation quantify participants' ability to organize visual searches.
Evaluating measures of search organization in children from early childhood to late adolescence will provide novel evidence of the pattern of development for search organization skills. We hypothesize that maturation of search organization skills will correspond with improvement in children's conjunction search accuracy, but not feature search accuracy. We further posit that children will demonstrate developmental inflection points in their search organization skills that coincide with maturation of executive processes and neurodevelopmental milestones in associated brain regions. Alternatively, there will be no relationship between organization and accuracy measures if maturation of search organization is unrelated to improvement in children's search accuracy. These data will provide insight into the development of search organization and its role in developmental changes in visual search performance.
1.3 Spatial orientation and visual search
Many studies use visual search to probe spatial attention (e.g., Treisman & Gelade, 1980). In conjunction searches, spatial attention is crucial for shifting visual search between locations in the environment (e.g., Robertson & Brooks, 2006). However, an initial spatial location in the environment must first be chosen, or oriented to, to serve as the starting point for serial search. Where we orient spatial attention to the visual environment to start a serial search, or search orientation, can influence the organization of search behavior. For example, starting search at or near a perceived edge or corner of the environment provides an effective foundation for planning and completing a more organized search pattern, compared to starting in the middle (Wilson et al., 1998). Although spatial attention has been the focus of many visual search studies, the influence of search orientation on visual search performance remains unexplored.
The orientation of spatial attention shifts leftward over the course of elementary school years as children learn to read in a left-right reading culture (Berch, Foley, Hill, & Ryan, 1999; Chokron & De Agostini, 1995; van Galen & Reitsma, 2008; but see Opfer et al., 2010; Opfer & Furlong, 2011). The opposite pattern is true for right-left reading cultures (e.g., Arabic; Chokron & De Agostini, 1995). For instance, eight-year-old children's attentional orientation was significantly biased more leftward for French children (left-right) compared to Israeli children (right-left; Chokron & De Agostini, 1995) on a task requiring them to mark the midpoint of physical lines of varied length (i.e., line bisection). Furthermore, these directional biases were less pronounced in younger children at age five, before formal reading education. If true, age-related change in search orientation may play a role in conjunction search performance differences between children and adults.
The present paper aimed to 1) investigate the development of search organization skills from early childhood to late adolescence and 2) evaluate the contribution of children's search organization skills to their visual search performance. We also investigated the feasibility of age and reading-related biases in children's initial orientation of spatial attention for serial search (i.e., search orientation). To achieve these aims, 134 children between the ages of two and seventeen were administered a cancellation test designed for children, a feature search task, and a conjunction search task. If search organization is an important component of younger children's difficulty with conjunction search, we should find that 1) search organization improves with age, 2) this improvement contributes to conjunction search accuracy, and 3) this improvement does not contribute to feature search accuracy. We expect children to start serial search more often on the left-side of the cancellation page as they get older, with the most predominant changes in orientation occurring after exposure to formal reading education. Furthermore, if search orientation effects reflect changes related to reading, children will demonstrate a propensity for initially orienting spatial attention to the upper-left portion of the cancellation page, consistent with the starting location for reading, rather than simply orienting more leftward.
2. Methods
2.1 Subjects
Participants were 134 children between the ages of 2.42 and 17.82 years (Mean ± Standard Deviation (SD) = 8.58 ± 3.86; 60 females; 13 left handed; 2 ambidextrous). The sample was recruited from a local pediatric office, day camp and high school, and from personal contacts. Most children were African-American (50.7%) and White (31.3%), with the rest (18%) of Asian American and Hispanic ethnicity. Children received a gift card for participation.
2.2 Cancellation design and procedure
Participants were given an apple cancellation test. The cancellation test presented 16 target apples (16 × 20 mm) against a white background intermixed with 16 panda bear distractors (20 × 18 mm) and 16 wrapped gift distractors (14 × 13 mm) in a pseudorandom arrangement (Figure 1c). The cancellation test was reproduced on white photocopy paper that was 216 mm wide by 279 mm long and placed on an empty horizontal table. The page was centered at the midsagittal plane and placed comfortably within arm's reach. Participants were not permitted to reposition the page. Participants were instructed to draw an individual line only through each apple once. Regardless of handedness, a near equal number of participants used either their left or right hand to perform cancellation (right = 66, left = 68). This procedure was done to focus findings on effects related to visual search, rather than handedness.
Participants were instructed to tell the examiner when they completed crossing out all of the apples. They were neither told to work quickly nor given a time limit. The experimenter sat across from the participant with an apple cancellation test oriented upside down, to mimic the patient's viewpoint. As the participant cancelled targets, the experimenter recorded the order of cancellation on their cancellation test. The examiner did not cue or inform participants about their progress. Participants who ceased exploring the page for about 30 seconds were asked a neutral question such as “Done?” or “Got them all?” Participants were allowed to resume canceling if they restarted exploring the page immediately. Otherwise, the test was terminated if the participants did not communicate that the test was complete. Experimenters were trained for accuracy of manual cancellation recording with mock participants of adult age. Unlike manual recording on more complex cancellation displays (e.g., modified star cancellation; Woods et al., 2007), experimenters were able to maintain 100% accuracy rates consistently on the apple cancellation task.
2.3 Cancellation measurement procedures
All test stimuli were identified by the Cartesian (x, y) coordinates of their geometric centers in reference to a consistent origin at page center. We assumed that participants treated individual targets or non-targets as whole figures and thus were not concerned with marking a specific part of a stimulus. Accordingly, when calculating spatial exploratory measures, we represented the locations of markings on targets or non-targets by the designated coordinates of the stimuli rather than by the precise points of initial pen contact.
2.3.1 Search Organization
We used three cancellation organization measures applied in healthy adults and adult patients with brain injury. These techniques demonstrate both sensitivity and convergent validity as proxies for the organization of visual search in a target and distractor rich environment (Mark et al., 2004; Woods et al., 2007).
1) Mean inter-target distance. The Euclidean distances between pen markings at sequentially different locations were averaged. Well-organized cancellation involves moving to the nearest unmarked targets in succession, thereby minimizing the distances between markings. We calculated the average marking distance for each participant rather than the total cancellation path length because the sum of inter-stimulus marking distances would be affected by failure to mark any of the 16 targets.
2) Number of intersections in the reconstructed cancellation path. The cancellation paths were reconstructed from experimenter paper records by digitally plotting them in Microsoft Excel. Well-organized search does not typically involve revisiting parts of the page that were already marked. Thus, well-organized search minimizes path intersections. The greater the area on the page that is marked, the greater the potential for the reconstructed cancellation path to intersect itself, regardless of how well participants were actually organized. Consequently, the total number of intersections was divided by the number of sequentially different locations marked.
3) Overall path structure - `best-r'. Healthy adult participants typically cancel by rows or columns (Donnelly et al., 1999; Gauthier et al., 1989; Mark et al., 2004; Weintraub & Mesulam, 1988), their movement is generally either horizontal (e.g., left to right) or radial (e.g., far to near) across the page. To capture this net orthogonal movement pattern, we calculated the Pearson correlation coefficient (r) from the linear regression of the x-values of all marked locations relative to the order in which they were marked. The y-values of marked locations were analyzed in the same way. From the two linear regressions calculated for each participant, we selected the one with the higher (“best”) r-value to represent the degree to which cancellations were pursued orthogonally. For example, starting on the left side of the page and marking by columns progressively rightward would yield a higher r-value on the x-coordinate regression than on the y-coordinate regression, because the cancellation progress would be consistently horizontal (left-to-right) but inconsistently radial. In general, a highly organized approach would be reflected by a high best-r.1 Figures 2a and b illustrate examples from our participants of organized and disorganized cancellation, respectively.
Figure 2.

Examples of visual search performance. a) an example of organized visual search with short inter-target distances, no path intersections, and high best-r (age: 13.1, Mean inter-target distance: 4.59, Intersections: 0, best-r: 0.96); b) an example of disorganized visual search with long inter-target distances, numerous intersections, and low best-r (age: 2.46, Mean inter-target distance: 8.45, Intersections: 8, best-r : 0.29).
2.3.2 Search Orientation
Part of efficient visual search involves determining where in the environment to begin searching. Starting at or near the edge of the page provides a foundation for more organized search than starting in the middle of the page (Wilson et al., 1998). Starting at the edges or in the corners of the page could decrease the likelihood of path intersections and facilitate either orthogonal or radial search. To evaluate whether age influences search orientation in visual search, we assessed 1) the frequency that the 16 unique target locations were selected as the first target cancelled and 2) the quadrant of initial cancellation. Using the center page x-, y-coordinate center (0,0), we divided the page into equal quadrants containing 4 targets each (top-left, bottom-left, top-right, bottom-right).
2.4 Feature & Conjunction Search design and procedures
Following completion of the apple cancellation test, participants were given separate computerized feature and conjunction search tasks. Search tasks were presented on a laptop fitted with a button box. Participants used the same hand to make button responses as was used to perform cancellation. Feature Search always preceded the Conjunction Search task.
Both tasks displayed 25 objects evenly distributed in a 5 × 5 rectangular grid (Figure 1a & b; Display size: 330 × 205 mm; Total grid dimensions: 275 × 180 mm). The Feature Search task contained 24 uniform distractor objects (blue squares with smiley faces, 20 × 20 mm; r = 105, g = 186, b = 99) and one target object (red circle with a smiley face, diameter: 20 mm; r = 226, g = 129, b = 112). The Conjunction Search task contained 24 distractor objects comprised of either red squares with smiley faces (n = 11) or blue circles with smiley faces (n = 13). The target was also a red circle with a smiley face. Conjunction search distractors were pseudo-randomly distributed (Figure 1b). For both tasks, targets were located in one of the 16 unique locations that were not in the center row or column. Both tasks also included catch trials where no target was present in the display (i.e., trials that contained only distractors). Catch trials were used to verify that participants were actually performing the requested task.
Participants were told: “In this task, you are looking for the red circle. Sometimes there will be one and sometimes there will not.” If a red circle was present, participants were instructed to press one button, if it was not, they were instructed to press a second button. They were instructed to respond as quickly as possible, but to also answer correctly. At the beginning of each task, participants were given six practice trials and provided with feedback during practice. Practice trial blocks contained two catch trials and four test trials. If accuracy was less than 50%, practice trials were repeated. If participants demonstrated difficulty pressing the response keys, experimenters were allowed to enter responses for them based on the child's verbal response to the trials.
Both practice and test blocks for each task began with instructions, followed by a fixation asterisk (Arial 40 point font; Feature: 500 ms; Conjunction: 1000 ms), and finally a test stimulus. Duration of stimulus presentation was eight seconds. Once participants responded, the next fixation screen was presented, followed by the next stimulus. Each test block contained 16 test trials and 8 catch trials (ntrials = 24). Test trials were visual arrays containing a target at one of the 16 unique target locations. Search and catch trial accuracy were calculated as the percent of trials accurately identified as containing or not containing a target, respectively. Participants' search accuracy data were excluded from analysis if they incorrectly responded to catch trials at chance levels (50%) or worse (i.e., they were randomly pressing buttons or pressing the same button for all trials).
3. Analyses
3.1 Search Organization & Orientation Measures
Separate univariate analyses of covariance (ANCOVA) were used to evaluate the effect of age on dependent measures of average marking distance, intersections, and best-r. Age was modeled as a covariate, while gender and hand-used were modeled as fixed independent factors. Frequency data were plotted to assess meaningful patterns in search orientation and multinomial logistic regression was used to evaluate significant effects of age, gender, and hand-used on the quadrant of search orientation. Locally weighted scatterplot smoothing (Loess or Lowess, also referred to as locally-weighted regression) trendlines, with a 60% Epanechnikov kernel function, were plotted on scatter plots of each variable (y-axis) versus age (x-axis) to evaluate the best fitting locally weighted regression line associated with age related changes. Loess does not require an assumption of linear or non-linear patterns within the data. Instead Loess fits simple models to localized subsets (or clusters) of the data and computes a function describing the deterministic part of the variation in the data on a point-by-point basis. This approach can provide a useful method for characterizing the overall pattern of data when variability is high, data are densely clustered, it is inappropriate to simplistically classify data as linear, or it is difficult to assign a specific non-linear function to the data (e.g., cubic, quadratic, etc.; examples of Loess used in other cognitive fields: Druker & Anderson, 2010; Kuperman & Van Dyke, 2011). One subject's intersection data were classified as an outlier, as the number of intersections was more than three standard deviations above the group mean (age = 2.4).
3.2 Feature versus Conjunction Search Accuracy
A paired t-test was used to compare accuracy in feature versus conjunction search. ANCOVA's were used to evaluate for previously reported effects of age on feature and conjunction search accuracy. Age was modeled as a covariate, while gender and hand-used were modeled as fixed independent factors. Separate ANCOVA's modeled search organization measures (average marking distance, intersections, best-r, and search orientation) and age × search organization interactions to evaluate for relationships between 1) search organization and search accuracy and 2) age-related improvements in search accuracy and development of improved search organization skills. Six children did not complete the conjunction search task (mean age±SD = 6.8±6.3), and were thus excluded from the conjunction search analyses. One participant failed to meet catch trial criteria for the Feature Search task (age = 2.4) and six participants for the Conjunction Search task (mean age±SD = 3.9±1).
4. Results
4.1 Mean Inter-target Distance
An analysis of covariance demonstrated that age was a significant covariate for mean inter-target distance (F1,129 = 8.4, p = .004, ηp2 = .06), with younger children moving further between targets than older (Figure 3a). Gender, Hand-used, and the Gender × Hand-used interaction were not significant (F1,129's < 1.5, p's > .22). According to the Loess trendline, participants continue improving in their ability to choose the next closest target on cancellation even at age 17. The largest degree of change appears to occur before age eleven (see Figure 3a).
Figure 3.

The effects of age on search organization and efficiency. a) The mean inter-target distance between targets marked; b) the number of intersections in the cancellation pathway, controlled for by the number of targets cancelled; c) the overall radial or right-to-left organization of the cancellation pathway; d) the frequency of participants starting in each quadrant stratified by age (top-left: n = 79, bottom-left: n = 24, bottom-right: n = 19, top-right: n = 9). Dark gray lines (a, b, & c) represent locally-weighted regression (Loess or Lowess) trendlines with a 60% Epanechnikov kernel function.
4.2 Intersections
An analysis of covariance demonstrated that age was a significant covariate for the number of path intersections committed during cancellation (F1, 128= 36.9, p < .001, ηp2 = .22), with younger children committing more intersections than older (Figure 3b). Gender and Hand-used were not significant (F1, 128's < 2.1, p's > .14). However, there was a significant interaction between Gender and Hand-Used (F1, 128 = 4.3, p = .04, ηp2 = .03). This significant interaction appears to arise from a tendency for participants using their right hand (right-hand: .17±.02; left-hand: .09±.02) and females (female: .10±.02; male: .89±.02) to commit a slightly higher number of intersections. According to the Loess trendline, participants' performance improves the most before age twelve, at which point improvement in their ability to avoid intersecting their previous path continues at a much smaller rate of improvement.
4.3 Best-r
An analysis of covariance demonstrated that age was a significant covariate for children's tendency to use either a predominantly radial or left-right pattern of cancellation (F1, 129 = 59.9, p < .001, ηp2 = .32), with younger children less likely to use an organized pattern of cancellation than older (Figure 3c). Gender, Hand-used, and the Gender × Hand-used interaction were not significant (F1, 129's < 3.4, p's > .07). According to the Loess trendline, participants' performance improves most before age 13, at which point improvement continues at a much slower rate (see Figure 3c).
4.4 Search Orientation
Frequency analysis demonstrated that the most frequently marked first target was at the extreme top-left of the cancellation page (36.6%, n=49/134; Figure 4 – position 1). The next most frequent target was also in the top-left area of the page, immediately to the right of the most frequently marked target (13.4%, n = 18/134; Figure 4 – position 8). All other targets were marked by at least one participant as an initial target cancelled, but no target was marked by more than 9 of 134 participants (<6.7%). As frequently marked first targets appeared to load onto the top left-quadrant, we divided the cancellation display into four equal quadrants containing 4 targets each (see Figure 4 for graph of quadrant division). Frequency analyses of starting quadrant demonstrated that 59 percent (n = 79) of participants began cancellation on one of the four targets in the top-left quadrant of the cancellation page. In comparison, only 17.9 (n = 24), 9 (n = 12), and 14.2 percent (n = 19) started cancellation on the four targets in either the bottom-left, top-right, or bottom-right of the cancellation page, respectively.
Figure 4.
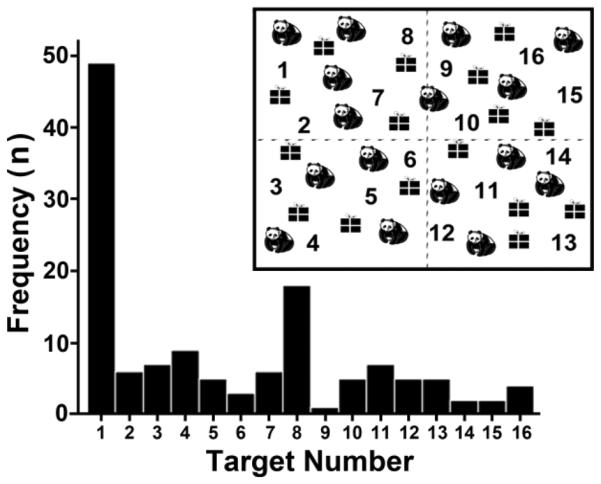
Spatial orientation. The frequency of children (n = 134) choosing a target as the first location of cancellation. Target number has been substituted for the apples in the cancellation display (top right corner of figure). Dashed line represents the quadrant division for quadrant-based analyses. Significantly more children (n = 49) began cancelation with the top-leftmost target compared to all other target locations.
Multinomial logistic regression evaluated effects of age, gender, and hand-used on search orientation. Multinomial logistic regression (Model Fit: χ2 = 38.0, DF = 12, p<.001; Cox & Snell Pseudo R-Square = .247) demonstrated that age (χ2 = 24.1, DF = 3, p<.001) was a significant predictor of search orientation. In contrast Gender, Hand-used, and the Gender × Hand-used interaction did not significantly predict search orientation (χ2's < 3.2, DF's = 3, p's>.35). Parameter estimates demonstrated that for every one-year increase in age, there was a decrease in log-odds of starting in the bottom-left (β = −.287, Wald χ2 = 10.6, DF = 1, p = .001), top-right (β = −.216, Wald χ2 = 4.0, DF = 1, p = .04), and bottom-right (β = −.334, Wald χ2 = 9.3, DF = 1, p = .002) of the cancellation display. As children grow older, they are more likely to orient to the top-left quadrant of the cancellation display when starting serial search (Figure 3d).
Subsequent evaluation of the distribution of search orientation across age demonstrated that the percent of participants' orienting search to either the left (58%) or the right (42%) side of the cancellation page was only slightly above chance levels before age seven (n = 54). These children demonstrated no preference for the upper (51.6%) versus lower (48.4%) left quadrant of the cancellation page. After age seven (n = 80), 89% of participants oriented to the left side of the cancellation page when starting serial search, versus 11% who oriented to the right. These children demonstrated a clear preference for the upper (87.5%) versus lower (12.5%) left quadrant of the cancellation page. For children age six (n = 14), the side of orientation was random (50%). For children age seven (n = 18), 83% of participants oriented search to the left when starting serial search of the cancellation page. 73.3% of these children demonstrated a preference for the upper left versus lower left quadrant.
4.5 Feature versus Conjunction Search Accuracy
Participants were less accurate on conjunction searches (mean accuracy±SE = 84±1.5%) versus feature searches (96.6±.004%; t = 8.0, DF = 122, p < .001).
4.5.1 Feature Search
Feature Search accuracy was near ceiling (96.6%). Feature Search accuracy did not demonstrate a significant relationship with age (F1,129 = 3.1, p = .08). Separate ANCOVA's demonstrated that none of the four search organization measures (distance, intersections, best-r, search orientation) were significantly related to feature search accuracy (F1,117's < 1.7, p's > .16). Gender, Hand-Used, Gender × Hand-used interactions, and Age × search organization interactions were not significant (F's < 1.3, p's > .26).
4.5.2 Conjunction Search
Age was a significant covariate for Conjunction Search accuracy (F1, 122 = 5.8, p = .01, ηp2 = .05), with younger children being less accurate than older children. Separate ANCOVA's demonstrated that best-r (F1,117 = 8.1, p = .005, ηp2 = .06) and search orientation (as defined by quadrant; F3,113 = 3.5, p = .01, ηp2 = .08) were significantly related to conjunction search accuracy. Search organization × age interactions demonstrated that age-related changes in best-r (F1,117 = 6.3, p = .01, ηp2 = .05) and search orientation (F3,113 = 4.6, p = .004, ηp2 = .11) were significantly related to improvement in conjunction search accuracy. Distance (F1,117 = 0.1, p = .75) or intersections (F1,117 = 0.1, p = .74) were not related to conjunction search accuracy. Gender, Hand-Used, and the Gender × Hand-used interactions were not significant (F1,117's < 1.8, p's > .20).
Factor analysis was used to determine whether differences in the underlying components reflected by the three search organization measures explain why best-r demonstrates a significant relationship with age-related change in conjunction search accuracy, but not distance and intersections. Analyses demonstrated that two underlying components explain 89.2% (component 1 = 58%, component 2 = 31.2%) of the variance in distance, best-r, and intersections. The correlation matrix from the factor analysis demonstrated that while intersection was significantly correlated with both distance (r = .46, p < .01) and best-r (r = −.53, p < .01), distance and best-r were not correlated (r = −.06, p = .22). Thus, the two latent components assessed by the three search organization measures load primarily onto distance and best-r, while intersections shares common variance with each of the other variables.
5. Discussion
From finding a lost shoe in a cluttered room to locating a car in a busy parking lot, we frequently search within the visual environment. Our results demonstrate that children's search organization skills improve with age. Children's level of ability to organize serial search is an important component of younger children's difficulty with conjunction search tasks. Our results suggest that children's conjunction search accuracy improves as their ability to organize serial search develops. In contrast, feature search performance was near ceiling (96.6%). Our findings also provide evidence that visual search orientation, like other visuospatial tasks, is influenced by age.
5.1 Development of search organization skills
All three search organization measures (distance, intersections, best-r) demonstrated a significant relationship with age. The most extensive improvements in search organization occurred by age twelve (see Figure 3a–c). After age twelve, the rate of improvement was incremental for distance, intersections, and best-r, but did not fully plateau even at age seventeen. Continued, albeit reduced, improvement in distance, intersections, and best-r through age seventeen is consistent with the development of executive functions (Baird et al., 2002; Diamond & Goldman-Rakic, 1989; Elliot, 2003; Funahashi, 2001; Moriguchi & Hiraki, 2009). Executive processes, including action/motor planning, working memory, inhibitory control, and mental flexibility, are particularly relevant to our distance, intersections, and best-r measures of search organization. These processes are important for shifting spatial attention to new locations in space (distance), preventing return to a previously visited location (distance, intersections, best- r), keeping track of overall progress across the cancellation page (distance, intersections, best- r), and planning a path that searches all possible spatial locations (intersections, best-r).
The developmental inflection point for distance, intersections, and best-r also corresponds with a significant neurodevelopment milestone for the DLFPC. The DLPFC reaches peak levels of gray matter density between the ages of 11 and 12. After this time, the process of neuronal pruning reduces gray matter density and improves neural efficiency by eliminating redundant neuronal connections, until final maturation of the DLPFC occurs (Geidd, 2004; Geidd, et al., 1999; Lenroot & Geidd, 2006). We posit that age-related results from distance, intersections, and best-r reflect maturational processes affiliated with the development of the DLPFC and related executive processes. Thus, late development of prefrontal structures important for executive functions, relative to early development of areas in the visual cortex important for feature search (e.g., V4; Zhou & Desimone, 2011), could explain, at least in part, children's difficulty performing conjunction but not feature search. Future studies evaluating children with lesions to the DLPFC or healthy children in functional neuroimaging designs will help determine the validity of this hypothesis.
5.2 Development of search orientation
Where we choose to first orient our spatial attention to the visual environment provides an important foundation for executive processes to plan and execute organized and efficient serial search (Wilson et al., 1998). By age seven, almost all children oriented to the top-left quadrant when starting their serial search of the cancellation page. Our data suggest that children's search orientation changes most significantly between the ages of six and seven, after approximately two years exposure to formal reading education. Prior research suggests that children in left-right reading cultures, over the course of elementary school years, develop a leftward attentional orientation bias for some visuospatial tasks (Berch, Foley, Hill, & Ryan, 1999; Chokron & De Agostini, 1995; van Galen & Reitsma, 2008). In contrast, the opposite effect is found for children learning Arabic – reading from right-to-left (Zebian, 2005). For example, children from left-right reading cultures typically demonstrate a leftward orientation bias on line bisection, and a more rightward bias is seen for right-left reading cultures.
After approximately two years of formal reading education (i.e., age 7 and older), children had an eighty percent or greater chance of orienting to the left versus right side of the cancellation page. Before age seven, the left versus right direction of orientation was near chance levels (58% left, 42% right). Furthermore, younger children who did orient to the left were at chance levels for choosing the upper (51.6%) versus lower (48.4%) left quadrant of the cancellation page. In contrast, children age seven demonstrated a clear preference for the upper left (73.3%) versus lower left (26.7%) quadrant – a location similar to the starting position of reading. Although the present study did not acquire measures of reading proficiency and cannot make a statistical association between changes in search orientation and reading education, our results are consistent with other studies demonstrating such effects (Berch, Foley, Hill, & Ryan, 1999; Chokron & De Agostini, 1995; van Galen & Reitsma, 2008). More importantly, the present findings indicate that spatial attention orientation biases play a role in an aspect of visual search behavior. Future studies evaluating the relationship between children's reading abilities in both left-right and right-left reading cultures would be helpful in elucidating the degree that reading influences the spatial orientation of attention in serial visual search and its specific impact on visual search performance.
5.3 The role of search organization in visual search accuracy
The primary goal of visual search is to locate a target object amongst a field of distractors in the environment. However, visual search is not always successful and conjunction search, in particular, is prone to higher rates of failure in young children. Trick & Enns (1998) argued that a variety of cognitive skills underlie developmental differences in conjunction search. Our data suggest that Trick & Enns (1998) and Donnelly et al (2007) were correct in positing that developmental limitations in conjunction search performance play a role in children's difficulty to efficiently plan and execute serial visual search. Our results demonstrate that the degree of search organization skills possessed by children plays a significant role in age-related errors in conjunction searches.
Best-r is a measurement of participants' ability to structure their search of the environment. In conjunction search, this would facilitate searching all possible areas in the visual display where the target might be located - facilitating accurate location of the target. As children get better at structuring their visual search of the environment, age-related deficits in conjunction search accuracy improve. Similar to the role of best-r, search orientation provides a foundation for planning and executing an organized search of the environment. As children shift their orientation to the upper-left corner of the display, age-related difficulties with conjunction search accuracy improve. Our results demonstrate that aspects of search organization related to executive function (best-r) and spatial attention (search orientation) contribute to the maturation of conjunction search abilities. Future research evaluating search organization measures in conjunction with tests of working memory capacity, mental flexibility, and other executive sub-processes will help identify the specific aspects of executive function underlying poor organization of visual search in younger children. Furthermore, experiments evaluating the effects of training-based manipulations on children with high versus low search organization abilities will expand our understanding of the relationship between search organization skills and search accuracy.
In contrast to results for conjunction search, the interpretation of the relationship between maturation of search organization and feature search performance is less clear. Near ceiling performance in the feature search accuracy, but clear developmental maturation in search organization could suggest that maturation of search organization skills are unrelated to feature search. However, this conclusion cannot be definitively drawn from the present data. Future research manipulating factors like cue salience to increase the difficulty of feature search will help evaluate the relationship between feature search and maturation of search organization skills.
6. Conclusion
In summary, children better organize their visual search of the environment, as they get older. As children's skills at organizing visual search improve they become more accurate at locating a target amongst distractors in conjunction search. Thus, developmental limitations in children's abilities to organize their visual search of the environment are an important component of poor conjunction search in young children. Our results suggest that maturation of executive functions instantiated in the dorsolateral prefrontal cortex may play a key role in age-related improvement in search organization. Furthermore, our results demonstrate that, like other visuospatial tasks, left-right reading effects may influence how we orient to the visual environment when required to perform a visual search.
Research Highlights
The ability to organize visual search rapidly develops until age twelve
Search organization skills play a role in conjunction search accuracy in children
Executive and search organization developmental inflection points coincide
Leftward spatial biases influence children's initial spatial orientation
Acknowledgements
The project described was supported by award numbers T32NS007413 and R01 DC008779 of the National Institutes of Health. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Exceptions might occur if targets were marked in other, atypical but highly organized manners, such as marking targets in a single spiral path encompassing the whole page. This would result in a lower best-r than when marking by columns or rows. However, we did not observe such exceptions.
References
- Anderson K, Starck L, Rosin I, Svensson E. The development of simple acoustic reaction time in normal children. Developmental Medicine & Child Neurology. 1984;26:490–494. doi: 10.1111/j.1469-8749.1984.tb04476.x. doi: 10.1111/j.1469-8749.1984.tb04476.x. [DOI] [PubMed] [Google Scholar]
- Ashbridge E, Walsh V, Cowey A. Temporal aspects of visual search studied by transcranial magnetic stimulation. Neuropsychologia. 1997;35:1121–1131. doi: 10.1016/s0028-3932(97)00003-1. [DOI] [PubMed] [Google Scholar]
- Baird AA, Kagan J, Gaudette T, Walz KA, Hershlag N, Boas DA. Frontal lobe activation during object performance: data from near-infrared spectroscopy. Neuroimage. 2002;16:1120–26. doi: 10.1006/nimg.2002.1170. [DOI] [PubMed] [Google Scholar]
- Berch DB, Foley EJ, Hill RJ, Ryan PM. Extracting parity and magnitude from Arabic numerals: Developmental changes in number processing and mental representation. Journal of Experimental Child Psychology. 1999;74:286–308. doi: 10.1006/jecp.1999.2518. [DOI] [PubMed] [Google Scholar]
- Bernstein LJ, Robertson LC. Independence between illusory conjunctions of color and motion with shape following bilateral parietal lesions. Psychological Science. 1998;9:167–175. [Google Scholar]
- Bichot NP, Rossi AF, Desimone R. Parallel and serial neural mechanisms for visual search in macaque area V4. Science. 2005;308:529–534. doi: 10.1126/science.1109676. [DOI] [PubMed] [Google Scholar]
- Boot WR, McCarley JS, Kramer AF, Peterson MS. Automatic and intentional memory processes in visual search. Psychological Bulletin & Review. 2004;11:854–861. doi: 10.3758/bf03196712. [DOI] [PubMed] [Google Scholar]
- Carrasco M. Visual attention: the past 25 years. Vision Research. 2011;51:1484–1525. doi: 10.1016/j.visres.2011.04.012. doi: 10.1016/j.visres.2011.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrasco M, Giordano AM, McElree B. Attention speeds processing across eccentricity: feature and conjunction searches. Vision Research. 2006;46:2028–2040. doi: 10.1016/j.visres.2005.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chokron S, De Agostini M. Reading habits and line bisection: a developmental approach. Cognitive Brain Research. 1995;3:51–58. doi: 10.1016/0926-6410(95)00018-6. [DOI] [PubMed] [Google Scholar]
- Coyle TR, Pillow DR, Snyder AC, Kochunov P. Processing speed mediates the development of general intelligence in adolescence. Psychological Science. 2011;22:1265–1269. doi: 10.1177/0956797611418243. doi: 10.1177/0956797611418243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deary IJ, Der G. Reaction time, age, and cognitive ability: Longitudinal findings from age 16 to 63 years in representative population samples. Aging, Neuropsychology, and Cognition. 2005;12:187–215. [Google Scholar]
- Diamond A, Goldman-Rakic PS. Comoarison of human infants and rhesus monkeys on Piaget's AB task: evidence for dependence on dorsolateral prefrontal cortex. Experimental Brain Research. 1989;74:24–40. doi: 10.1007/BF00248277. [DOI] [PubMed] [Google Scholar]
- Donnelly N, Cave K, Greenway R, Hadwin JA, Stevenson J, Sonuga-Barke E. Visual search in children and adults: top-dwon and bottom-up mechanisms. The Quarterly Journal of Experimental Psychology. 2007;60:120–136. doi: 10.1080/17470210600625362. [DOI] [PubMed] [Google Scholar]
- Donnelly N, Guest R, Fairhurst M, Potter J, Deighton A, Patel M. Developing algorithms to enhance the sensitivity of cancellation tests of visuospatial neglect. Behavioral Research Methods in Instructional Computing. 1999;31:668–673. doi: 10.3758/bf03200743. [DOI] [PubMed] [Google Scholar]
- Druker M, Anderson B. Spatial probability aids stimulus discrimination. Frontiers in Human Neuroscience. 2010;63:1–10. doi: 10.3389/fnhum.2010.00063. doi: 10.3389/fnhum.2010.00063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duncan J, Humphreys GW. Visual search and stimulus similarity. Psychological Review. 1989;96:433–458. doi: 10.1037/0033-295x.96.3.433. [DOI] [PubMed] [Google Scholar]
- Elliot R. Executive functions and their disorders. British Medical Bulletin. 2003;65:49–59. doi: 10.1093/bmb/65.1.49. [DOI] [PubMed] [Google Scholar]
- Ellison A, Walsh V. Perceptual learning in visual search: some evidence of specificities. Vision Research. 1998;38:333–45. doi: 10.1016/s0042-6989(97)00195-8. [DOI] [PubMed] [Google Scholar]
- Emrich SM, Al-Aidroos N, Pratt J, Ferber S. Visual search elicits the electrophysiological marker of visual working memory. PLoS ONE. 2009;4(11):e8042. doi: 10.1371/journal.pone.0008042. doi:10.1371/journal.pone.0008042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman-Hill S, Robertson LC, Treisman A. Parietal contributions to visual feature binding: Evidence from a patient with bilateral lesions. Science. 1995;269:853–855. doi: 10.1126/science.7638604. [DOI] [PubMed] [Google Scholar]
- Funahashi S. Neuronal mechanisms of executive control by the prefrontal cortex. Neuroscience Research. 2001;39:147–65. doi: 10.1016/s0168-0102(00)00224-8. [DOI] [PubMed] [Google Scholar]
- Gauthier L, Dehaut F, Joanette Y. The Bells Test: a quantitative and qualitative test for visual neglect. International Journal of Clinical Neuropsychology. 1989;11:49–54. [Google Scholar]
- Gerhardstein P, Rovee-Collier C. The development of visual search in infants and very young children. Journal of Experimental Child Psychology. 2002;81:194–215. doi: 10.1006/jecp.2001.2649. doi: 10.1006/jecp.2001.2649. [DOI] [PubMed] [Google Scholar]
- Gibson BS, Jiang Y. Surprise! An unexpected color singleton does not capture attention in visual search. Psychological Science. 1998;9:176–182. [Google Scholar]
- Giedd JN. Structural magnetic resonance imaging of the adolescent brain. Annals of the New York Academy of Science. 2004;1021:77–85. doi: 10.1196/annals.1308.009. [DOI] [PubMed] [Google Scholar]
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A, Paus T, Evans AC, Rapoport A. Brain development during childhood and adolescence: a longitudinal MRI study. Nature Neuroscience. 1999;2:861–863. doi: 10.1038/13158. [DOI] [PubMed] [Google Scholar]
- Han S, Kim M. Visual search does not remain efficient when executive working memory is working. Psychological Science. 2004;15:623–628. doi: 10.1111/j.0956-7976.2004.00730.x. [DOI] [PubMed] [Google Scholar]
- Humphreys GW, Cinel C, Wolfe J, Olson A, Klempen N. Fractionating the binding process: Neuropsychological evidence distinguishing binding of form from binding of surface features. Vision Research. 2000;40:1569–1596. doi: 10.1016/s0042-6989(00)00042-0. [DOI] [PubMed] [Google Scholar]
- Jakobsen LH, Sorensen JM, Rask IK, Jensen BS, Kondrup J. Validation of reaction time as a measure of cognitive function and quality of life in healthy subjects and patients. Nutrition. 2011;27:561–70. doi: 10.1016/j.nut.2010.08.003. [DOI] [PubMed] [Google Scholar]
- Kalla R, Muggleton NG, Cowey A, Walsh V. Human dorsolateral prefrontal cortex is involved in visual search for conjunctions but not features: a theta TMS study. Cortex. 2009;45:1085–1090. doi: 10.1016/j.cortex.2009.01.005. [DOI] [PubMed] [Google Scholar]
- Kuperman V, Van Dyke JA. Effects of individual differences in verbal skills on eye-movement patterns during sentence reading. Journal of Memory & Language. 2011;65:42–73. doi: 10.1016/j.jml.2011.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lenroot RK, Geidd JN. Brain development in children and adolescents: Insights from anatomical magnetic resonance imaging. Neuroscience & Biobehavioral Reviews. 2006;30:718–729. doi: 10.1016/j.neubiorev.2006.06.001. [DOI] [PubMed] [Google Scholar]
- Luria R, Vogel EK. Visual search demands disctate reliance on working memory storage. Journal of Neuroscience. 2011;31:6199–6207. doi: 10.1523/JNEUROSCI.6453-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mark VW, Woods AJ, Ball KK, Roth DL, Mennemeier M. Disorganized search on cancellation is not a consequence of neglect. Neurology. 2004;63:78–84. doi: 10.1212/01.wnl.0000131947.08670.d4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mark VW, Woods AJ. Age effects on cancellation speed and organization. Journal of the American Geriatric Society. 2003;51:S388. [Google Scholar]
- Moriguchi Y, Hiraki K. Neural origin of cognitive shifting in young children. Proceedings of the National Academy of Science. 2009;106:6017–21. doi: 10.1073/pnas.0809747106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muggleton NG, Chen CY, Tzeng OJL, Hung DL, Juan CH. Inhibitory control and the frontal eye fields. Journal of Cognitive Neuroscience. 2010;22:2804–2812. doi: 10.1162/jocn.2010.21416. [DOI] [PubMed] [Google Scholar]
- Muggleton NG, Juan C-H, Cowey A, Walsh V. Human frontal eye fields and visual search. Journal of Neurophysiology. 2003;89:3340–343. doi: 10.1152/jn.01086.2002. [DOI] [PubMed] [Google Scholar]
- Opfer JE, Furlong EE. How numbers bias preschoolers' spatial search. Jounral of Cross-Cultural Psychology. 2011;42:682–695. [Google Scholar]
- Opfer JE, Thompson CA, Furlong EE. Early development of spatial-numeric associations: evidence from spatial and quantitative performance of preschoolers. Developmental Science. 2010;13:761–71. doi: 10.1111/j.1467-7687.2009.00934.x. [DOI] [PubMed] [Google Scholar]
- Peterson MS, Kramer AF, Wang RF, Irwin DE, McCarley JS. Visual search has memory. Psychological Science. 2001;12:287–292. doi: 10.1111/1467-9280.00353. [DOI] [PubMed] [Google Scholar]
- Peterson MS, Beck MR, Vomela M. Visual search is guided by prospective and retrospective memory. Perception & Psychophysics. 2007;69:123–135. doi: 10.3758/bf03194459. [DOI] [PubMed] [Google Scholar]
- Philip BR. Reaction-times of children. The American Journal of Psychology. 1934;46:379–396. [Google Scholar]
- Robertson LC. Binding, spatial attention and perceptual awareness. Nature Reviews Neuroscience. 2003;4:93–102. doi: 10.1038/nrn1030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robertson LC, Brooks JL. Visual search and spatial deficits. Visual Cognition. 2006;14:851–862. [Google Scholar]
- Sowell ER, Thompson PM, Tessner KD, Toga AW. Mapping continued brain growth and gray matter density reduction in dorsal frontal cortex: inverse relationships during postadolescent brain maturation. The Journal of Neuroscience. 2001;21:8819–8829. doi: 10.1523/JNEUROSCI.21-22-08819.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson LA, Massaro DW. Before you see it, you see its parts: Evidence for feature encoding and integration in preschool children and adults. Cognitive Psychology. 1989;21:334–362. doi: 10.1016/0010-0285(89)90012-1. [DOI] [PubMed] [Google Scholar]
- Treisman A, Gelade G. A feature-integration theory of attention. Cognitive Psychology. 1980;12:97–136. doi: 10.1016/0010-0285(80)90005-5. [DOI] [PubMed] [Google Scholar]
- Treisman A, Sato S. Conjunction search revisited. Journal of Experimental Psychology: Human Perception and Performance. 1990;16:459–478. doi: 10.1037//0096-1523.16.3.459. [DOI] [PubMed] [Google Scholar]
- Trick LM, Enns JT. Life changes in attention: the visual search task. Cognitive Development. 1998;13:369–386. [Google Scholar]
- van Galen MS, Reitsma P. Developing access to number magnitude: A study of the SNARC effect in 7- to 9-year-olds. Journal of Experimental Child Psychology. 2008;101:99–113. doi: 10.1016/j.jecp.2008.05.001. [DOI] [PubMed] [Google Scholar]
- Walsh V, Ashbridge E, Cowey A. Cortical plasticity in perceptual learning demonstrated by transcranial magnetic stimulation. Neuropsychologia. 1998;36:45–49. doi: 10.1016/s0028-3932(97)00111-5. [DOI] [PubMed] [Google Scholar]
- Weintraub S, Mesulam MM. Visual hemispatial inattention: stimulus parameters and exploratory strategies. Journal of Neurology Neurosurgery & Psychiatry. 1988;51:1481–1488. doi: 10.1136/jnnp.51.12.1481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson BA, Evans JJ, Emslie H, Alderman N, Burgess P. The development of an ecologically valid test for assessing patients with a dysexecutive syndrome. Neuropsychological Rehabilitation. 1998;8:213–228. [Google Scholar]
- Wolfe JM, Cave KR, Franzel SL. Guided Search: An alternative to the Feature Integration model for visual search. Journal of Experimental Psychology: Human Perception and Performance. 1989;15:419–433. doi: 10.1037//0096-1523.15.3.419. [DOI] [PubMed] [Google Scholar]
- Woods AJ, Mark VW. Severe executive dysfunction is a component of left but not right neglect. Journal of the International Neuropsychological Society. 2005;11:45. [Google Scholar]
- Woods AJ, Mark VW. Convergent validity of executive organization measures on cancellation. Journal of Clinical and Experimental Neuropsychology. 2007;29:719–723. doi: 10.1080/13825580600954264. [DOI] [PubMed] [Google Scholar]
- Woods AJ, Mark VW, Mennemeier M. Alterations in stimulus response influence cancellation performance. Journal of the International Neuropsychological Society. 2004;10:91. [Google Scholar]
- Zebian S. Linkages between Number Concepts, Spatial Thinking, and Directionality of Writing: The SNARC Effect and the REVERSE SNARC Effect in English and Arabic Monoliterates, Biliterates, and Illiterate Arabic Speakers. Journal of Cognition & Culture. 2005;5:165–190. [Google Scholar]
- Zhou H, Desimone R. Feature-based attention in the frontal eye fields and area V4 during visual search. Neuron. 2011;70:1205–1217. doi: 10.1016/j.neuron.2011.04.032. [DOI] [PMC free article] [PubMed] [Google Scholar]