

SUMMARY
The anti-convulsant Stiripentol (Diacomit™) has been shown to have a positive impact on control of seizures for many patients with Dravet Syndrome. As with most anti-epileptic drugs, stiripentol has multiple mechanisms of action. Its direct anti-convulsant activity is likely due to enhancement of inhibitory, GABAergic neurotransmission. Stiripentol was shown to increase the activity of both neuronal and recombinant GABAA receptors at clinically relevant concentrations. At recombinant receptors, stiripentol was found to act through a unique site in a subunit-dependent manner. Positive modulation by stiripentol was most effective at GABAA receptors containing an α3 subunit. The expression of the α3 subunit is developmentally regulated, with highest levels in the immature brain. This subunit selectivity may explain the greater clinical efficacy of stiripentol in childhood-onset epilepsies, including Dravet Syndrome.
Keywords: recombinant, patch-clamp, Dravet Syndrome
Stiripentol (STP) is a novel anti-epileptic drug (AED) structurally unrelated to other anticonvulsants used clinically to treat seizure disorders (Figure 1) (Trojnar et al., 2005). It has received orphan drug status as adjunctive therapy for the treatment of severe myoclonic epilepsy in infancy (SMEI, Dravet Syndrome) and can be effective in reduction of pharmacoresistant seizures and status epilepticus. STP has both direct and indirect anticonvulsant actions.
Figure 1. Structure of Stiripentol.
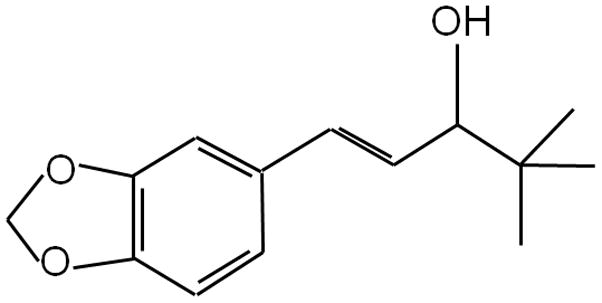
Stiripentol (4,4-dimethyl-1-[3,4-(methylene dioxy)-phenyl]-1-penten-3-ol) is structurally unrelated to other clinically available anti-convulsant drugs.
Stiripentol inhibits metabolism of other anti-convulsant drugs
STP has been tested for decades in combination with other AEDs because of its inhibitory action on metabolic enzymes. STP inhibits a variety of hepatic cytochrome P450 enzymes, with high activity at human CYP1A2, CYP3A4 and CYP2C19 (Tran et al., 1997). As a result, it increases the peak concentrations and duration of action of several commonly used AEDs, including phenobarbital, clobazam, carbamazepine, phenytoin and valproate (Trojnar et al., 2005; Landmark and Johannessen, 2008). For patients with Dravet syndrome, STP is commonly combined with clobazam. Evidence suggests that much of the anti-convulsant activity of clobazam is mediated through its active metabolite, norclobazam (N-desmethylclobazam) (Kinoshita et al., 2007). While co-administration with STP increases the concentration of both clobazam and norclobazam, it has a much greater impact on norclobazam levels (Giraud et al., 2006). Therefore, some of the improved efficacy seen with the combination of STP and clobazam may be due to increased levels of norclobazam. In addition to its effects on drug metabolism, there is strong evidence that STP alone is directly anti-convulsant. STP monotherapy reduces seizure severity in a variety of rodent models and in a primate model of focal epilepsy (Trojnar et al. 2005). This suggested that STP has additional effects on seizure activity independent of alterations in drug metabolism. Subsequent studies have shown that STP acts directly on GABAA receptors (GABAARs) as a positive allosteric modulator.
Stiripentol enhances GABAergic neurotransmission
The GABAARs are ligand-gated chloride channels responsible for fast inhibitory neurotransmission. They are targets for many commonly used anti-convulsants, including benzodiazepines and barbiturates. Experimental evidence for a direct action of STP on the activity of GABAARs was first provided by Quilichini et al. (2006). Using a neonatal rat brain slice preparation they found that STP slowed the decay of GABAergic mIPSCs in hippocampal CA3 pyramidal cells. STP increased both the frequency and the duration of post-synaptic events, suggesting both pre- and post-synaptic effects. A direct modulation of GABAARs was confirmed with single-channel recordings, which showed that STP increased the mean open time of GABA-activated channels.
Stiripentol has subunit-dependent effects on recombinant GABAA receptors
The finding that STP interacts with post-synaptic GABAARs in hippocampal neurons was extended by studies of recombinant receptors (Fisher, 2009). Neuronal GABAARs can have a large degree of structural heterogeneity due to variations in subunit composition. At least 16 different GABAAR subunits have been found in mammalian brain and the composition of the receptor has a significant impact on its functional and pharmacological properties (Korpi et al., 2002).
STP was found to positively modulate all recombinant GABAARs tested, but had its greatest effect when the receptor contained an α3 subunit. Unlike the benzodiazepines, STP did not require presence of a γ subunit and was a strong modulator of δ-containing receptors. STP was shown to enhance GABAAR activity by increasing sensitivity to GABA without increasing the peak current to maximally effective GABA concentrations. This mechanism is similar to that of benzodiazepine-site agonists, but the STP site is clearly distinct from other well-known GABAAR modulators. The activity of STP is not blocked by antagonists of the benzodiazepine or neurosteroid sites (Quilichini et al., 2006; Fisher, 2009) nor is it influenced by mutations that disrupt loreclezole or neurosteroid activity (Fisher, 2009). Co-administration studies suggest that STP acts independently and additively with benzodiazepines and neurosteroids, though it is not able to further enhance modulation by pentobarbital (Quilichini et al., 2006 and Fisher, 2009). STP has a unique pattern of subunit dependence unlike any other GABAAR modulator. Its particularly high activity at α3- and δ-containing receptors could have important implications for its effect on neuronal activity and its clinical usefulness.
Characteristics of the α3 subunit of GABAA receptors
Expression of the α3 subunit is strongly regulated by development. mRNA for the α3 is found at relatively high levels in most brain regions of the embryonic and newborn mammal (Laurie et al., 1992). As maturation progresses, its expression becomes more restricted and it is produced primarily by neurons in the cortex and thalamus of the adult brain. However, higher production of the α3 subunit has been found in adult animals that have a greater susceptibility to seizures (Poulter et al., 1999). GABAARs containing the α3 subunit have been suggested to play an important role in normal cortical development, and in adults may mediate some anxiolytic and anticonvulsant effects of GABA-modulating drugs (Sohal et al., 2003; Yee et al., 2005; Dias et al., 2005).
Characteristics of the δ subunit of GABAA receptors
GABAARs containing the δ subunit represent a unique population of receptors (Belelli et al., 2009). These receptors are exclusively located in extra-synaptic regions, and contribute to the long-lasting tonic current observed in the hippocampus, cerebellum and thalamus in response to the low ambient levels of GABA. The δ subunit confers functional properties such as high agonist sensitivity and minimal desensitization that make them well-suited for this role. The δ-containing receptors also have unique pharmacological properties. In particular, they are insensitive to positive modulation by the benzodiazepine class of AEDs. Therefore, drugs such as STP that enhance the activity of these receptors may retain effectiveness in patient populations that are pharmacoresistant to benzodiazepines.
Summary
The studies of neuronal and recombinant GABAARs confirm that STP can have a direct action on neurotransmission, supporting its anti-convulsant action in animal models of acute seizures. The preferential enhancement of α3-containing receptors provides a plausible mechanism for the observation that STP is very effective in treating childhood seizures, despite its lower clinical utility in adults. These findings reinforce the need to consider the unique characteristics of the immature, developing brain when designing therapies to treat neurological disorders of infants and children.
Acknowledgments
Dr. Fisher’s studies with Stiripentol are supported by a research grants from Biocodex Pharmaceuticals and from NIH-NINDS (RO1-NS045950).
Footnotes
Disclosures: Dr. J. Fisher has received research support from Biocodex Pharmaceuticals
I confirm that I have read the Journal’s position on issues involved in ethical publication and affirm that this report is consistent with those guidelines.
References
- Belelli D, Harrison NL, Maguire J, Macdonald RL, Walker MC, Cope DW. Extrasynaptic GABAA receptors: form, pharmacology and function. J Neurosci. 2009;29:12757–12763. doi: 10.1523/JNEUROSCI.3340-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dias R, Sheppard WFA, Fradley RL, Garrett EM, Stanley JL, Tye SJ, Goodacre S, Lincoln RJ, Cook SM, Conley R, Hallett D, Humphries AC, Thompson SA, Wafford KA, Street LJ, Castro JL, Whiting PJ, Rosahl TW, Atack JR, McKernan RM, Dawson GR, Reynolds DS. Evidence for a significant role of α3-containing GABAA receptors in mediating the anxiolytic effects of benzodiazepines. J Neurosci. 2005;25:10682–10688. doi: 10.1523/JNEUROSCI.1166-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher JL. The anti-convulsant stiripentol acts directly on the GABAA receptor as a positive allosteric modulator. Neuropharm. 2009;56:190–197. doi: 10.1016/j.neuropharm.2008.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giraud C, Treluyer J-M, Rey E, Chiron C, Vincent J, Pons G, Tran A. In vitro and in vivo inhibitory effect of stiripentol on clobazam metabolism. Drug Metab and Dispos. 2006;34:608–611. doi: 10.1124/dmd.105.007237. [DOI] [PubMed] [Google Scholar]
- Kinoshita M, Ikeda A, Begum T, Terada K, Shibasaki H. Efficacy of low-dose, add-on therapy of clobazam (CLB) is produced by its major metabolite N-desmethyl-CLB. J Neurol Sci. 2007;263:44–48. doi: 10.1016/j.jns.2007.05.025. [DOI] [PubMed] [Google Scholar]
- Korpi ER, Gründer G, Lüddens H. Drug interactions at GABAA receptors. Prog Neurobiol. 2002;67:113–159. doi: 10.1016/s0301-0082(02)00013-8. [DOI] [PubMed] [Google Scholar]
- Landmark CJ, Johannessen SI. Pharmacological management of epilepsy. Drugs. 2008;68:1925–1939. doi: 10.2165/00003495-200868140-00001. [DOI] [PubMed] [Google Scholar]
- Laurie DJ, Wisden W, Seeburg PH. The distribution of thirteen GABAA receptor subunit mRNAs in the rat brain. III. Embryonic and postnatal development. J Neurosci. 1992;12:4151–4172. doi: 10.1523/JNEUROSCI.12-11-04151.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poulter MO, Brown LA, Tynan S, Willick G, William R, McIntyre DC. Differential expression of α1, α2, α3, and α5 GABAA receptor subunits in seizure-prone and seizure-resistant rat models of temporal lobe epilepsy. J Neurosci. 1999;19:4654–4661. doi: 10.1523/JNEUROSCI.19-11-04654.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quilichini PP, Chiron C, Ben-Ari Y, Gozlan H. Stiripentol, a putative antiepileptic drug, enhances the duration of opening of GABAA-receptor channels. Epilepsia. 2006;47:704–716. doi: 10.1111/j.1528-1167.2006.00497.x. [DOI] [PubMed] [Google Scholar]
- Sohal VS, Keist R, Rudolph U, Huguenard JR. Dynamic GABAA receptor subtype-specific modulation of the synchrony and duration of thalamic oscillations. J Neurosci. 2003;23:3649–3657. doi: 10.1523/JNEUROSCI.23-09-03649.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tran A, Rey E, Pons G, Rouseau M, d’Athis P, Olive G, Mahter GG, Bishop FE, Wurden CJ, Labroo R, Trager WF, Kunze KL, Thummel KE, Vincent JC, Gillardin J-M, Lepage F, Levy RH. Influence of stiripentol on cytochrome P450-mediated metabolic pathways in humans: In vitro and in vivo comparison and calculation of in vivo inhibition constants. Clin Pharmacol Ther. 1997;62:490–504. doi: 10.1016/S0009-9236(97)90044-8. [DOI] [PubMed] [Google Scholar]
- Trojnar MK, Wojtal K, Trojnar MP, Czuczwar SJ. Stiripentol. A novel antiepileptic drug. Pharm Reports. 2005;57:154–160. [PubMed] [Google Scholar]
- Yee BK, Keist R, von Bohmer L, Studer R, Benke D, Hagenbuch N, Dong Y, Malenka RC, Fritschy J-M, Bluethmann H, Feldon J, Möhler H, Rudolph U. A schizophrenia-related sensorimotor deficit links α3-containing GABAA receptors to a dopamine hyperfunction. Proc Natl Acad Sci. 2005;(102):17154–17159. doi: 10.1073/pnas.0508752102. [DOI] [PMC free article] [PubMed] [Google Scholar]