

Abstract
In this study, to study the effect of aging and Apolipoprotein E (ApoE) deficiency on antioxidant ability in mice, we examined whether lipid peroxidation is promoted by aging in ApoE deficient (ApoE−/−) mice, which have a shorter lifespan than normal mice. The levels of thiobarbituric acid-reactive substances (TBARS), a biomarker of lipid peroxidation, were measured in plasma and liver in ApoE−/− mice aged 12 weeks (young) and 52 weeks (early stage of senescence). TBARS in plasma and liver were significantly increased by aging. Next, we examined the reasons why lipid peroxidation was promoted by aging, based on measurement of protein and mRNA levels for antioxidant enzymes (superoxide dismutase, catalase, and glutathione peroxidase) in liver in ApoE−/− mice aged 12 and 52 weeks. The levels of superoxide dismutase 1 and 2 in liver were significantly decreased by aging. The mRNA level of catalase was also significantly decreased and the mRNA levels of superoxide dismutase 1, superoxide dismutase 2 and glutathione peroxidase 1 all showed a tendency to decrease with age. These results suggest that lipid peroxidation is caused by reduction of antioxidant activity with aging and that this promotes senescence and shortens lifespan in ApoE−/− mice.
Keywords: aging, apolipoprotein E, lipid peroxidation, senescence, TBARS
Introduction
The population of elderly persons is increasing worldwide and this trend is expected to continue, with a parallel increase in age-related diseases.(1,2) Aging without disease is important for maintenance of personal QOL (quality of life) and for development of society and the economy. Prevention of age-related diseases requires development of a better understanding of senescence.
The senescence-accelerated mouse (SAM) has a decreased lifespan due to acceleration of senescence caused by an age-related increase in lipid peroxidation.(3,4) Previously, we examined age-dependent changes in lipid peroxidation in SAMP10 mice and found that the levels of lipid peroxides in plasma and liver were significantly increased by aging.(5) We also previously investigated the effect of intake of a large amount of fish oil on intravital lipid peroxidation and lifespan in SAMP8 mice, with the finding that the levels of lipid peroxides in plasma and liver were increased and lifespan was decreased by intake of fish oil.(6)
These findings indicate that lipid peroxidation and senescence are closely related, and that inhibition of accumulation of lipid peroxides will slow down senescence. That is, prevention of lipid peroxidation may delays senescence and increases lifespan. However, it is unclear if age-dependent promotion of lipid peroxidation is observed only in SAM mice. There are mouse strains with shorter lifespans in addition to SAM mice. The lifespan of the normal C57BL/6 mouse is about 2 years, but that of the Apolipoprotein E deficient (ApoE−/−) mouse is only about 1.5 years.(7,8) The ApoE−/− mouse is widely used to study metabolic syndrome as an animal model of atherosclerosis,(9,10) but few studies have examined the effect of ApoE deficiency on senescence and lipid peroxidation. The ApoE−/− mouse has various characteristics associated with aging, but further studies are needed to establish this mouse as an animal model of accelerated senescence.(11)
In this study, we examined whether lipid peroxidation is promoted by aging in ApoE−/− mice. At 12–15 months of age, the ApoE−/− mouse exhibits an age-related decline in presynaptic terminals in the hippocampus.(12) Therefore, we defined the ApoE−/− mouse at 13 months (52 weeks) of age to be in the early stage of senescence. We bred ApoE−/− mice and examined the levels of thiobarbituric acid-reactive substances (TBARS), a biomarker of lipid peroxidation, in plasma and liver at ages 12 weeks (young) and 52 weeks (early stage of senescence). In addition, to examine changes in antioxidant capacity with aging, the protein and mRNA levels of antioxidant enzymes (superoxide dismutase, catalase, and glutathione peroxidase) were measured in liver at ages 12 and 52 weeks.
Materials and Methods
Animals and diets
All procedures were performed in accordance with the Animal Experiment Guidelines of Tohoku University. The animal protocol was approved by the Animal Use Committee at Tohoku University.(13) Female ApoE−/− mice on a C57BL/6J background were obtained from The Jackson Laboratory (Bar Harbor, Maine). C57BL/6J (wild type, WT) mice were obtained from Japan Clea (Tokyo, Japan). Mice had free access to distilled water and fed ad libitum with standard diet (MF; Oriental Yeast, Tokyo, Japan). ApoE−/− mice were sacrificed for analysis at age 12 weeks (12ApoE−/−) and 52 weeks (52ApoE−/−). WT mice were also sacrificed for analysis at age 12 weeks (12WT) and 52 weeks (52WT). The mice were housed in individual cages in a temperature- and humidity-controlled room with light cycles of 12 h on and 12 h off.(14,15) At the appropriate time point, the mice were sacrificed by decapitation, and the liver and plasma were collected and stored at −80°C until performance of assays.
TBARS assay
To examine oxidative stress caused by aging, the levels of TBARS in plasma and liver were determined as described previously.(6,16–18)
Western blot analysis
Western blot analysis was performed using antibodies that specially recognize proteins including catalase (CAT), glutathione peroxidase 1 (GPx1), superoxide dismutase 1 (SOD1), superoxide dismutase 2 (SOD2), and β-actin as described previously.(14) Livers were homogenized, and proteins were extracted. 10 µg of extracted protein was loaded for sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) immunoblot analysis. The protein bands were then transferred to polyvinylidene fluoride membranes (Bio-Rad Laboratories, Hercules, CA). After being blocked of nonspecific sites, the membrane was probed with primary antibodies, followed by a horseradish peroxidase-conjugated secondary antibody (Abcam, Cambridge, MA). The detection of the antibody reactions was performed with ECL Western blotting Detection Reagents (GE Healthcare, Little Chalfont, UK). Each band was normalized by corresponding value of β-actin as an internal control. The antibodies used were anti-CAT (Sigma, St. Louis, MO), anti-GPx1 (Abcam), anti-SOD1 (Abcam), anti-SOD2 (Abcam), and anti-β-actin (Abcam).(19–22)
mRNA expression analysis
For real-time quantitative reverse transcriptase polymerase chain reaction (qRT-PCR), total RNA was isolated from liver using an RNeasy Mini Kit (Qiagen, Valencia, CA),(23) eluted with 30 µl RNase-free water, and stored at –80°C until use. To quantify the expression level of genes, the mRNA levels for catalase (Cat), glutathione peroxidase 1 (Gpx1), superoxide dismutase 1, soluble (Sod1), superoxide dismutase 2, mitochondrial (Sod2), actin, beta (Actb) in liver were determined with a 7300 Real-Time PCR System (Applied Biosystems, Carlsbad, CA). This system allows real-time quantitative detection of PCR products by measuring the increase in fluorescence caused by binding of SYBR green to double-stranded DNA.(13,14) In brief, cDNA was made using a Ready-To-Go T-Primed First-Strand Kit (GE Healthcare) from the total RNA in liver. The cDNA was subjected to PCR amplification using SYBR Premix Ex Taq (Perfect Real Time) (Takara Bio, Otsu, Japan) and gene-specific primers for Cat, Gpx1, Sod1, Sod2, and Actb (Table 1). The PCR conditions were 95°C for 10 s, and then 95°C for 5 s and 60°C for 31 s over 40 cycles for each gene. Melting curve analysis was performed following each reaction to confirm the presence of only a single reaction product. The threshold cycle (CT) represents the PCR cycle at which an increase in reporter fluorescence above a baseline signal can first be detected. The Actb content in each sample was used to normalize the results.(24)
Table 1.
Primer pairs used for the quantitative RT-PCR analysis
| Genbank ID | Target gene | Primer | Primer sequence (5'-3') |
|---|---|---|---|
| NM_007393 | Actb | Forward | GAAATCGTGCGTGACATCAAAG |
| Reverse | TGTAGTTTCATGGATGCCACAG | ||
| NM_009804 | Cat | Forward | AGCGACCAGATGAAGCAGTG |
| Reverse | TCCGCTCTCTGTCAAAGTGTG | ||
| NM_008160 | Gpx1 | Forward | AGTCCACCGTGTATGCCTTCT |
| Reverse | GAGACGCGACATTCTCAATGA | ||
| NM_011434 | Sod1 | Forward | AACCAGTTGTGTTGTCAGGAC |
| Reverse | CCACCATGTTTCTTAGAGTGAGG | ||
| NM_013671 | Sod2 | Forward | CAGACCTGCCTTACGACTATGG |
| Reverse | CTCGGTGGCGTTGAGATTGTT |
Actb, actin, beta; Cat, catalase; Gpx1, glutathione peroxidase 1; Sod1, superoxide dismutase 1, soluble; Sod2, superoxide dismutase 2, mitochondrial.
Biochemical analyses in plasma and liver
The lipid compositions in the plasma and liver were measured as described previously.(25–27) Triacylglycerol (TG) and total cholesterol (TC) levels in plasma and liver, and phospholipid (PL) levels in plasma were measured using commercial enzyme kits (Wako Pure Chem. Ind. Ltd., Osaka, Japan) according to the manufacturer’s protocol. PL levels in liver were determined using the method described by Rouser.(28)
Statistical analysis
Results are expressed as means ± SE. Data were compared between 12ApoE−/− and 52ApoE−/− or 12WT and 52WT. Unpaired two-tailed t test was used to analyze differences between two groups. A difference was considered to be significant at p<0.05.
Results
Effects of aging on lipid peroxidation in plasma and liver of ApoE−/− mice
To investigate the age-dependent changes in lipid peroxidation in ApoE−/− mice, TBARS in plasma and liver were measured at ages 12 weeks (young) and 52 weeks (early stage of senescence) (Fig. 1). As a control, TBARS in plasma and liver of WT mice at ages 12 and 52 weeks were also measured. The plasma TBARS level in the 52ApoE−/− group was 642% of that in the 12ApoE−/− group, showing a significant increase with age. In contrast, there was no significant difference between the 12WT and 52WT groups. The liver TBARS level in the 52ApoE−/− group was 138% of that in the 12ApoE−/− group, also showing a significant increase with age. Again, there was no significant difference between the 12WT and 52WT groups. These results show that TBARS in plasma and liver of ApoE−/− mice were increased by aging, indicating that lipid peroxidation is promoted by aging in ApoE−/− mice.
Fig. 1.

Effects of aging on TBARS concentrations in plasma and liver of ApoE−/− mice (A) and WT mice (B). Values are means ± SE, n = 5–9. *p<0.05 (vs 12-week-old mice).
Effects of aging on antioxidant enzymes in liver of ApoE−/− mice
The promotion of lipid peroxidation in plasma and liver of ApoE−/− mice by aging suggests that the antioxidant capacity in these mice is impaired by aging. Therefore, the expression levels of antioxidant enzymes in liver were measured using Western blot analysis (Fig. 2). SOD catalyzes the dismutation of superoxide into oxygen and hydrogen peroxide. The SOD1 level in the 52ApoE−/− group was 35% of that in the 12ApoE−/− group, showing a significant decrease with aging, whereas there was no significant difference between the 12WT and 52WT groups. Similarly, the SOD2 level in the 52ApoE−/− group was 70% of that in the 12ApoE−/− group, again showing a significant decrease with aging, with no significant difference between the 12WT and 52WT groups. CAT catalyzes the decomposition of hydrogen peroxide to oxygen and water, and GPx1 catalyzes the decomposition of the hydroperoxide group to a hydroxyl group and hydrogen peroxide to water. Changes in CAT and GPx1 levels with aging were not observed in ApoE−/− or WT mice. However, the decreased levels of antioxidant enzymes SOD1 and SOD2 in the liver of ApoE−/− mice with aging suggests that lipid peroxidation in plasma and liver was promoted because of a reduction in antioxidant capacity in older ApoE−/− mice.
Fig. 2.

Effects of aging on expression of SOD1, SOD2, CAT and GPx1 proteins in the liver of ApoE−/− mice (A) and WT mice (B). Expression levels were measured by western blotting. The β-actin content in each sample was used to normalize the results. Upper sides represent quantitative value and bottom sides show graphic data of each enzyme. Values are means ± SE, n = 5–9. *p<0.05 (vs 12-week-old mice).
Effects of aging on mRNA expression of antioxidant-related genes in liver of ApoE−/− mice
To examine the mechanisms underlying the reduced antioxidant ability in liver of ApoE−/− mice with aging, the mRNA levels of enzymes concerned with antioxidant systems in the liver were measured using qRT-PCR (Fig. 3). The level of Sod1 mRNA in the 52ApoE−/− group was 76% of that in the 12ApoE−/− group, showing a tendency for a decrease with age (p = 0.11). Similarly, the Sod2 mRNA level in the 52ApoE−/− group was 74% of that in the 12ApoE−/− group, also showing a tendency to decrease with age (p = 0.07). The Cat mRNA level in the 52ApoE−/− group was 52% of that in the 12ApoE−/− group, showing a significant decrease with age; and the Gpx1 mRNA level in the 52ApoE−/− group was 65% of that in the 12ApoE−/− group, showing a tendency to decrease with age (p = 0.06). Similar changes in these mRNA levels were not observed with aging in WT mice. These results show that the mRNA levels of enzymes concerned with antioxidant systems in the liver of ApoE−/− mice are decreased by aging, which suggests that reduced antioxidant activity is caused by lower gene expression levels in ApoE−/− mice.
Fig. 3.
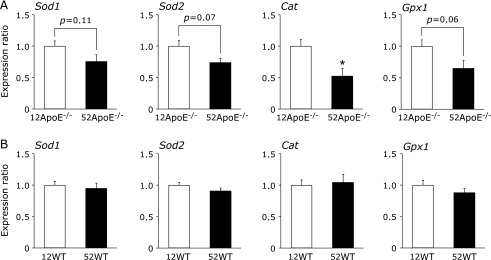
Effects of aging on expression of Sod1, Sod2, Cat and Gpx1 mRNA in the liver of ApoE−/− mice (A) and WT mice (B). Expression levels were measured by quantitative RT-PCR. The Actb content in each sample was used to normalize the results. Values are means ± SE, n = 5–9. *p<0.05 (vs 12-week-old mice).
Effects of aging on the lipid composition in plasma and liver of ApoE−/− mice
To investigate the age-dependent changes of lipid metabolism in ApoE−/− mice, the lipid composition in the plasma and liver were measured at ages 12 and 52 weeks (Table 2). Changes in plasma TG, TC and PL levels with aging were not observed in ApoE−/− or WT mice. Similarly, there were no changes in liver TG, TC and PL levels with aging in ApoE−/− and WT mice. These results indicate that lipid metabolism is unchanged in ApoE−/− mice between 12 and 52 weeks.
Table 2.
Effects of aging on lipid parameters in plasma and liver
| 12ApoE−/− | 52ApoE−/− | 12WT | 52WT | |
|---|---|---|---|---|
| Plasma | ||||
| TG (mM/L) | 0.61 ± 0.06 | 0.77 ± 0.04 | 0.62 ± 0.03 | 0.60 ± 0.04 |
| TC (mM/L) | 9.59 ± 0.49 | 11.17 ± 0.48 | 0.98 ± 0.06 | 1.20 ± 0.09 |
| PL (mM/L) | 0.16 ± 0.02 | 0.18 ± 0.04 | 1.28 ± 0.08 | 1.51 ± 0.08 |
| Liver | ||||
| TG (µM/g) | 46.3 ± 6.1 | 45.8 ± 13.7 | 78.0 ± 7.9 | 57.6 ± 6.2 |
| TC (µM/g) | 8.97 ± 0.28 | 9.94 ± 0.68 | 5.41 ± 0.50 | 5.45 ± 0.39 |
| PL (µM/g) | 30.3 ± 0.5 | 29.8 ± 1.0 | 26.0 ± 1.9 | 24.3 ± 1.2 |
Groups between the same strains were compared. Values are shown as means ± SE, n = 5–9. TG, triacylglycerol; TC, total cholesterol; PL, phospholipids.
Discussion
In this study, we investigated whether lipid peroxidation is promoted by aging in ApoE−/− mice. TBARS in plasma and liver of ApoE−/− mice at 52 weeks of age were significantly higher than those at 12 weeks of age. In contrast, TBARS in plasma and liver of normal (WT) mice were not changed by aging (Fig. 1). Thus, the results revealed that age-dependent promotion of lipid peroxidation in ApoE−/− mice can be attributed to deficiency of ApoE. In addition, the levels of antioxidant enzymes in liver of ApoE−/− mice were decreased by aging, leading to reduced antioxidant capacity (Fig. 2). Overall, these results suggest that lipid peroxidation in ApoE−/− mice is promoted because of reduction of antioxidant capacity by aging. Some previous studies examined relationship between ApoE deficiency and lipid peroxidation. Most of them, however, discussed oxidative stress among experimental groups in ApoE−/− mice at the same age.(29,30) In this study, we first demonstrated ”age-related” reduction of antioxidant activity and ”age-related” promotion of oxidative stress in ApoE−/− mice.
In this study, protein expression levels of SODs that are antioxidant enzymes were significantly reduced by aging in ApoE−/− mice (Fig. 2A). It was reported that SOD1 protect organisms against oxidative stress.(31,32) Therefore, the possibility that the reduction of SOD1 activity promotes lipid peroxidation is high. Since SOD2 protects against oxidative stress and endothelial dysfunction in the carotid artery of ApoE−/− mice, SOD2 deficiency increases endothelial dysfunction.(33) Since the promotion of lipid peroxidation shortens lifespan,(34) it was suggested that long-term depression of SOD activity shortens lifespan. In addition, it was thought that additional aging decreases antioxidant activity and promotes lipid peroxidation further since ApoE−/− mice at 52 weeks of age are early stage of senescence.
ApoE binds directly to 4-hydroxynonenal and has a protective effect against lipid peroxidation.(35) Therefore, lipid peroxidation may be promoted by aging in ApoE−/− mice because of the absence of this protective effect of ApoE. Thus, it was suggested that absence of protective effect of ApoE increases oxidative stress to impair the tissue function, change the cellular function and impair the antioxidant ability. Therefore, these changes of cellular functions due to oxidative stress in ApoE−/− mice might have decreased liver mRNA expression of antioxidant-related genes with aging, and oxidative stress in ApoE−/− mice might further promoted. Another possible reason for reduction of antioxidant enzymes in 52ApoE−/− group is that participation of nuclear factor erythroid 2-related factor-2 (Nrf2) that regulates gene expression of antioxidant relative genes. Nrf2 is negatively regulated by kelch-like ECH associating protein 1 (Keap1) and kept in cytoplasm.(36) To activate Nrf2, it must be dissociated from Keap1. Although cysteine residues in Keap1 play an important role in this regulation,(37) in senescent ApoE−/− mice, the antioxidant system may be impaired by the trouble of activating Nrf2 because of failure in modification of cysteine residues.
In SAM mice, oxidative stress increases with aging and promotes senescence.(3,4) We previously found that TBARS levels in plasma and liver were increased by aging in the SAM mice.(5,6) The finding in this study that lipid peroxidation was promoted by aging in ApoE−/− mice suggests that this effect promotes senescence and shortens lifespan.
In this study, plasma TBARS level in ApoE−/− mice was lower than that in WT mice at 12 weeks of age (Fig. 1). Plasma high-density lipoprotein (HDL) level in ApoE−/− mouse is much lower than that in WT mice.(9) And, HDL has a higher susceptibility to lipid peroxidation than low-density lipoprotein (LDL).(38–41) Therefore, since plasma TBARS level derived from HDL in ApoE−/− mice is lower than that in WT mice, ApoE−/− mice must have indicated lower plasma TBARS level than WT mice at 12 weeks of age. Moreover, liver TBARS level in ApoE−/− mice was also lower than that in WT mice at 12 weeks of age (Fig. 1). It may be because liver lipid level in ApoE−/− mice was lower than that in WT mice. It was reported that ApoE−/− mice exhibit delay of triglyceride clearance from plasma and protective effect against hepatic triglyceride accumulation.(42) Therefore, ApoE−/− mice must have indicated lower liver TBARS level than WT mice at younger age. And, it was thought that oxidative stress was increased because of promotion of lipid peroxidation with aging in ApoE−/− mice.
In plasma, the possibility that the fatty acid that exists as cholesterol ester was oxidized and plasma TBARS was increased was also thought. Further study is needed to clear these points.
In this study, TBARS assay was used to measure lipid peroxidation. TBARS assay is not enough to measure lipid peroxidation completely. However, this assay is widely used as a metrology of lipid peroxidation and many reports discuss lipid peroxidation by this assay.(43–46) In this study, we additionally measured protein and mRNA levels for antioxidant enzymes and indicated promotion of oxidative stress in ApoE−/− mice. In addition, to study about relationship between changes in antioxidant system and lipid peroxidation in detail, it may be effective to examine antioxidant levels (tocopherol, SOD, etc.) in plasma.
We also examined age-dependent changes in lipid metabolism in ApoE−/− mice. Lipid metabolism in plasma and liver was not changed by aging in these mice, which suggests that there is little change in lipid metabolism in the early stage of senescence (Table 2).
In conclusion, our results suggest that antioxidant activity was reduced and lipid peroxidation was caused by aging in ApoE−/− mice. So lipid peroxidation caused by age-dependent reduction of antioxidant activity in ApoE−/− mice may promotes senescence and shortens lifespan. Therefore, inhibition of accumulation of lipid peroxide may delay senescence and increase lifespan. It was reported that long-term intake of vitamin E, one of the major antioxidant, prevents the liver and the brain from lipid peroxidation and improves survival.(47) Therefore, it might be effective to prevent lipid peroxidation by long-term intake of antioxidants to prevent shortening lifespan in ApoE−/− mice. Furthermore, ApoE may have a beneficial role in life extension, and further studies of ApoE may provide useful knowledge for development of methods to delay senescence.
Acknowledgments
This study was supported by a Grant-in-aid for Scientific Research from Japan Society for the Promotion of Science (No.20157639).
Abbreviations
- Actb
actin,beta
- ApoE
apolipoprotein E
- ApoE−/−
apolipoprotein E deficient
- 12ApoE−/−
ApoE−/− mouse at age 12 weeks
- 52ApoE−/−
ApoE−/− mouse at age 52 weeks
- CAT
catalase
- Cat
catalase
- GPx1
glutathione peroxidase 1
- Gpx1
glutathione peroxidase 1
- HDL
high-density lipoprotein
- Keap1
kelch-like ECH-associated protein 1
- Nrf2
nuclear factor erythroid 2-related factor-2
- PL
phospholipids
- qRT-PCR
real-time quantitative reverse transcriptase polymerase chain reaction
- SAM
senescence-accelerated mouse
- SDS-PAGE
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
- SOD1
superoxide dismutase 1
- Sod1
superoxide dismutase 1, soluble
- SOD2
superoxide dismutase 2
- Sod2
superoxide dismutase 2, mitochondrial
- TBARS
thiobarbituric acid reactive substances
- TC
total cholesterol
- TG
triacylglycerol
- WT
wild type
- 12WT
WT mouse at age 12 weeks
- 52WT
WT mouse at age 52 weeks
Conflict of Interest
No potential conflicts of interest were disclosed.
References
- 1.Shannon J, Shikany JM, Barrett-Connor E, et al. Demographic factors associated with the diet quality of older US men: baseline data from the Osteoporotic Fractures in Men (MrOS) study. Public Health Nutr. 2007;10:810–818. doi: 10.1017/S1368980007258604. [DOI] [PubMed] [Google Scholar]
- 2.Mendoza-Núñez VM, Martínez-Maldonado Mde L, Correa-Muñoz E. Implementation of an active aging model in Mexico for prevention and control of chronic diseases in the elderly. BMC Geriatr. 2009;9:40. doi: 10.1186/1471-2318-9-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Matsugo S, Kitagawa T, Minami S, et al. Age-dependent changes in lipid peroxide levels in peripheral organs, but not in brain, in senescence-accelerated mice. Neurosci Lett. 2000;278:105–108. doi: 10.1016/s0304-3940(99)00907-6. [DOI] [PubMed] [Google Scholar]
- 4.Hosokawa M. A higher oxidative status accelerates senescence and aggravates age-dependent disorders in SAMP strains of mice. Mech Ageing Dev. 2002;123:1553–1561. doi: 10.1016/s0047-6374(02)00091-x. [DOI] [PubMed] [Google Scholar]
- 5.Honma T, Yanaka M, Tsuduki T, Ikeda I. Increased lipid accumulation in liver and white adipose tissue in aging in the SAMP10 mouse. J Nutr Sci Vitaminol (Tokyo) 2011;57:123–129. doi: 10.3177/jnsv.57.123. [DOI] [PubMed] [Google Scholar]
- 6.Tsuduki T, Honma T, Nakagawa K, Ikeda I, Miyazawa T. Long-term intake of fish oil increases oxidative stress and decreases lifespan in senescence-accelerated mice. Nutrition. 2011;27:334–337. doi: 10.1016/j.nut.2010.05.017. [DOI] [PubMed] [Google Scholar]
- 7.Singh SP, Niemczyk M, Saini D, Sadovov V, Zimniak L, Zimniak P. Disruption of the mGsta4 gene increases life span of C57BL mice. J Gerontol A Biol Sci Med Sci. 2010;65:14–23. doi: 10.1093/gerona/glp165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Moghadasian MH, McManus BM, Nguyen LB, et al. Pathophysiology of apolipoprotein E deficiency in mice: relevance to apo E-related disorders in humans. FASEB J. 2001;15:2623–2630. doi: 10.1096/fj.01-0463com. [DOI] [PubMed] [Google Scholar]
- 9.Zhang SH, Reddick RL, Piedrahita JA, Maeda N. Spontaneous hypercholesterolemia and arterial lesions in mice lacking apolipoprotein E. Science. 1992;258:468–471. doi: 10.1126/science.1411543. [DOI] [PubMed] [Google Scholar]
- 10.Maizel J, Six I, Slama M, et al. Mechanisms of aortic and cardiac dysfunction in uremic mice with aortic calcification. Circulation. 2009;119:306–313. doi: 10.1161/CIRCULATIONAHA.108.797407. [DOI] [PubMed] [Google Scholar]
- 11.Ang LS, Cruz RP, Hendel A, Granville DJ. Apolipoprotein E, an important player in longevity and age-related diseases. Exp Gerontol. 2008;43:615–622. doi: 10.1016/j.exger.2008.03.010. [DOI] [PubMed] [Google Scholar]
- 12.Buttini M, Yu GQ, Shockley K, et al. Modulation of Alzheimer-like synaptic and cholinergic deficits in transgenic mice by human apolipoprotein E depends on isoform, aging, and overexpression of amyloid beta peptides but not on plaque formation. J Neurosci. 2002;22:10539–10548. doi: 10.1523/JNEUROSCI.22-24-10539.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tsuzuki T, Tokuyama Y, Igarashi M, Miyazawa T. Tumor growth suppression by α-eleostearic acid, a linolenic acid isomer with a conjugated triene system, via lipid peroxidation. Carcinogenesis. 2004;25:1417–1425. doi: 10.1093/carcin/bgh109. [DOI] [PubMed] [Google Scholar]
- 14.Tsuzuki T, Kawakami Y. Tumor angiogenesis suppression by α-eleostearic acid, a linolenic acid isomer with a conjugated triene system, via peroxisome proliferator-activated receptor gamma. Carcinogenesis. 2008;29:797–806. doi: 10.1093/carcin/bgm298. [DOI] [PubMed] [Google Scholar]
- 15.Honma T, Shinohara N, Ito J, et al. High-fat diet intake accelerates aging, increases expression of Hsd11b1, and promotes lipid accumulation in liver of SAMP10 mouse. Biogerontology. 2012;13:93–103. doi: 10.1007/s10522-011-9363-2. [DOI] [PubMed] [Google Scholar]
- 16.Shinohara N, Tsuduki T, Ito J, et al. Jacaric acid, a linolenic acid isomer with a conjugated triene system, has a strong antitumor effect in vitro and in vivo. Biochim Biophys Acta. 2012;1821:980–988. doi: 10.1016/j.bbalip.2012.04.001. [DOI] [PubMed] [Google Scholar]
- 17.Yagi K. A simple fluorometric assay for lipoperoxide in blood plasma. Biochem Med. 1976;15:212–216. doi: 10.1016/0006-2944(76)90049-1. [DOI] [PubMed] [Google Scholar]
- 18.Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 1979;95:351–358. doi: 10.1016/0003-2697(79)90738-3. [DOI] [PubMed] [Google Scholar]
- 19.Qi X, Hauswirth WW, Guy J. Dual gene therapy with extracellular superoxide dismutase and catalase attenuates experimental optic neuritis. Mol Vis. 2007;13:1–11. [PMC free article] [PubMed] [Google Scholar]
- 20.Wirth EK, Conrad M, Winterer J, et al. Neuronal selenoprotein expression is required for interneuron development and prevents seizures and neurodegeneration. FASEB J. 2010;24:844–852. doi: 10.1096/fj.09-143974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kawase-Koga Y, Low R, Otaegi G, et al. RNAase-III enzyme Dicer maintains signaling pathways for differentiation and survival in mouse cortical neural stem cells. J Cell Sci. 2010;123:586–594. doi: 10.1242/jcs.059659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Csiszar A, Labinskyy N, Zhao X, et al. Vascular superoxide and hydrogen peroxide production and oxidative stress resistance in two closely related rodent species with disparate longevity. Aging Cell. 2007;6:783–797. doi: 10.1111/j.1474-9726.2007.00339.x. [DOI] [PubMed] [Google Scholar]
- 23.Tsuzuki T, Kambe T, Shibata A, Kawakami Y, Nakagawa K, Miyazawa T. Conjugated EPA activates mutant p53 via lipid peroxidation and induces p53-dependent apoptosis in DLD-1 colorectal adenocarcinoma human cells. Biochim Biophys Acta. 2007;1771:20–30. doi: 10.1016/j.bbalip.2006.11.006. [DOI] [PubMed] [Google Scholar]
- 24.Yanaka M, Honma T, Sato K, et al. Increased monocytic adhesion by senescence in human umbilical vein endothelial cells. Biosci Biotechnol Biochem. 2011;75:1098–1103. doi: 10.1271/bbb.100909. [DOI] [PubMed] [Google Scholar]
- 25.Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. Can J Biochem Physiol. 1959;37:911–917. doi: 10.1139/o59-099. [DOI] [PubMed] [Google Scholar]
- 26.Tsuzuki T, Kawakami Y, Suzuki Y, Abe R, Nakagawa K, Miyazawa T. Intake of conjugated eicosapentaenoic acid suppresses lipid accumulation in liver and epididymal adipose tissue in rats. Lipids. 2005;40:1117–1123. doi: 10.1007/s11745-005-1475-0. [DOI] [PubMed] [Google Scholar]
- 27.Tsuzuki T, Kawakami Y, Nakagawa K, Miyazawa T. Conjugated docosahexaenoic acid inhibits lipid accumulation in rats. J Nutr Biochem. 2006;17:518–524. doi: 10.1016/j.jnutbio.2005.09.006. [DOI] [PubMed] [Google Scholar]
- 28.Rouser G, Fkeischer S, Yamamoto A. Two dimensional then layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids. 1970;5:494–496. doi: 10.1007/BF02531316. [DOI] [PubMed] [Google Scholar]
- 29.Hayek T, Oiknine J, Brook JG, Aviram M. Increased plasma and lipoprotein lipid peroxidation in apo E-deficient mice. Biochem Biophys Res Commun. 1994;201:1567–1574. doi: 10.1006/bbrc.1994.1883. [DOI] [PubMed] [Google Scholar]
- 30.Ferré N, Camps J, Paul A, et al. Effects of high-fat, low-cholesterol diets on hepatic lipid peroxidation and antioxidants in apolipoprotein E-deficient mice. Mol Cell Biochem. 2001;218:165–169. doi: 10.1023/a:1007296919243. [DOI] [PubMed] [Google Scholar]
- 31.Imamura Y, Noda S, Hashizume K, et al. Drusen, choroidal neovascularization, and retinal pigment epithelium dysfunction in SOD1-deficient mice: a model of age-related macular degeneration. Proc Natl Acad Sci U S A. 2006;103:11282–11287. doi: 10.1073/pnas.0602131103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kojima T, Wakamatsu TH, Dogru M, et al. Age-related dysfunction of the lacrimal gland and oxidative stress: evidence from the Cu,Zn-superoxide dismutase-1 (Sod1) knockout mice. Am J Pathol. 2012;180:1879–1896. doi: 10.1016/j.ajpath.2012.01.019. [DOI] [PubMed] [Google Scholar]
- 33.Ohashi M, Runge MS, Faraci FM, Heistad DD. MnSOD deficiency increases endothelial dysfunction in ApoE-deficient mice. Arterioscler Thromb Vasc Biol. 2006;26:2331–2336. doi: 10.1161/01.ATV.0000238347.77590.c9. [DOI] [PubMed] [Google Scholar]
- 34.Patel BP, Safdar A, Raha S, Tarnopolsky MA, Hamadeh MJ. Caloric restriction shortens lifespan through an increase in lipid peroxidation, inflammation and apoptosis in the G93A mouse, an animal model of ALS. Plos One. 2010;5:e9386. doi: 10.1371/journal.pone.0009386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pedersen WA, Chan SL, Mattson MP. A mechanism for the neuroprotective effect of apolipoprotein E: isoform-specific modification by the lipid peroxidation product 4-hydroxynonenal. J Neurochem. 2000;74:1426–1433. doi: 10.1046/j.1471-4159.2000.0741426.x. [DOI] [PubMed] [Google Scholar]
- 36.Motohashi H, Yamamoto M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends Mol Med. 2004;10:549–557. doi: 10.1016/j.molmed.2004.09.003. [DOI] [PubMed] [Google Scholar]
- 37.Um HC, Jang JH, Kim DH, Lee C, Surh YJ. Nitric oxide activates Nrf2 through S-nitrosylation of Keap1 in PC12 cells. Nitric Oxide. 2011;25:161–168. doi: 10.1016/j.niox.2011.06.001. [DOI] [PubMed] [Google Scholar]
- 38.Bowry VW, Stanley KK, Stocker R. High density lipoprotein is the major carrier of lipid hydroperoxides in human blood-plasma from fasting donors. Proc Natl Acad Sci U S A. 1992;89:10316–10320. doi: 10.1073/pnas.89.21.10316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Viani P, Cazzola R, Cervato G, Gatti P, Cestaro B. Pyrene lipids as markers of peroxidative processes in different regions of low and high density lipoproteins. Biochim Biophys Acta. 1996;1315:78–86. doi: 10.1016/0925-4439(95)00094-1. [DOI] [PubMed] [Google Scholar]
- 40.Hurtado I, Fiol C, Gracia V, Caldu P. In vitro oxidised HDL exerts a cytotoxic effect on macrophages. Atherosclerosis. 1996;125:39–46. doi: 10.1016/0021-9150(96)05840-6. [DOI] [PubMed] [Google Scholar]
- 41.Ohmura H, Watanabe Y, Hatsumi C, et al. Possible role of high susceptibility of high-density lipoprotein to lipid peroxidative modification and oxidized high-density lipoprotein in genesis of coronary artery spasm. Atherosclerosis. 1999;142:179–184. doi: 10.1016/s0021-9150(98)00235-4. [DOI] [PubMed] [Google Scholar]
- 42.Karavia EA, Papachristou DJ, Kotsikogianni I, Giopanou I, Kypreos KE. Deficiency in apolipoprotein E has a protective effect on diet-induced nonalcoholic fatty liver disease in mice. FEBS J. 2011;278:3119–3129. doi: 10.1111/j.1742-4658.2011.08238.x. [DOI] [PubMed] [Google Scholar]
- 43.Bowman TA, Ramakrishnan SK, Kaw M, et al. Caloric restriction reverses hepatic insulin resistance and steatosis in rats with low aerobic capacity. Endocrinology. 2010;151:5157–5164. doi: 10.1210/en.2010-0176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kosinska W, Khmelnitsky M, Kim JH, Zhao ZL, Guttenplan JB. Effects of potential dietary inhibitors of endogenous DNA damage on mutagenesis and lipid peroxidation in lacZ mice. Environ Mol Mutagen. 2011;52:502–509. doi: 10.1002/em.20648. [DOI] [PubMed] [Google Scholar]
- 45.Arukwe A, Mortensen AS. Lipid peroxidation and oxidative stress responses of salmon fed a diet containing perfluorooctane sulfonic- or perfluorooctane carboxylic acids. Comp Biochem Physiol C Toxicol Pharmacol. 2011;154:288–295. doi: 10.1016/j.cbpc.2011.06.012. [DOI] [PubMed] [Google Scholar]
- 46.Kumar M, Verma V, Nagpal R, et al. Anticarcinogenic effect of probiotic fermented milk and chlorophyllin on aflatoxin-B1-induced liver carcinogenesis in rats. Br J Nutr. 2012;107:1006–1016. doi: 10.1017/S0007114511003953. [DOI] [PubMed] [Google Scholar]
- 47.Navarro A, Gómez C, Sánchez-Pino MJ, et al. Vitamin E at high doses improves survival, neurological performance, and brain mitochondrial function in aging male mice. Am J Physiol Regul Integr Comp Physiol. 2005;289:R1392–R1399. doi: 10.1152/ajpregu.00834.2004. [DOI] [PubMed] [Google Scholar]