

Abstract
The melanocortin-3 (MC3) and melanocortin-4 (MC4) receptors regulate energy homeostasis, food intake, and associated physiological conditions. The MC4R has been studied extensively. Less is known about specific physiological roles of the MC3R. A major obstacle to this lack of knowledge is attributed to a limited number of identified MC3R selective ligands. We previously reported a spatial scanning approach of a 10-membered thioether-heterocycle ring incorporated into a chimeric peptide template that identified a lead nM MC4R ligand. Based upon those results, 17 compounds were designed and synthesized that focused upon modification in the pharmacophore domain. Notable results include the identification of a 0.13 nM potent 5800-fold mMC3R selective antagonist/slight partial agonist versus a 760 nM mMC4R full agonist (ligand 11). Biophysical experiments (2D 1H NMR and computer assisted molecular modeling) of this ligand resulted in the identification of an inverse γ-turn secondary structure in the ligand pharmacophore domain.
Keywords: Melanotropin, obesity, feeding behavior, agouti related protein, AGRP, solid phase synthesis, peptide, small molecule, MC3R, MC4R, MC1R, MC5R, GPCR, reverse turn mimetic
Introduction
Peptide ligands for G-protein coupled receptor (GPCR) molecular probes have been well documented as valuable tools for studying ligand-receptor interactions and the differentiation of ligand-based agonist versus antagonist design strategies. The melanocortin GPCRs (consisting of five isoforms1–7) are involved in the regulation of a variety of physiological functions including pigmentation,8 cardiovascular effects,9, 10 food intake,11, 12 and erectile function.13, 14 These receptors are stimulated by the α-, β-,γ-melanocyte stimulating hormones (MSH) and adrenocorticotropin (ACTH) endogenous agonists that are derived from the proopiomelanocortin (POMC) gene.15, 16 Additionally, the melanocortin-1 receptor (MC1R) is endogenously antagonized by the Agouti peptide.17, 18 The Agouti-related protein (AGRP) functions as both a competitive antagonist and an inverse agonist at the melanocortin-4 receptor (MC4R).19, 20 These melanocortin GPCRs stimulate the cAMP signal transduction pathway in response to agonist ligand stimulation.
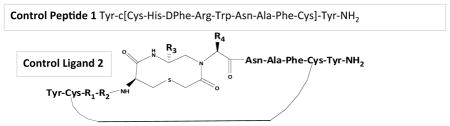
Extensive structure-activity relationship (SAR) studies since the 1960’s,21, 22 have characterized the presence of a reverse turn in the purported His-Phe-Arg-Trp pharmacophore domain of melanocortin receptor agonists. As the endogenous agonist ligands are linear in structure, they possess a plethora of possible “bioactive” conformations depending upon their local changing environment (aqueous or receptor bound). Successful strategies designed to decrease ligand flexible conformations and directed towards identification of the “bioactive” conformation include introduction of cyclic constraints,23–27 peptoids,28 aza peptides,29 D-amino acids,30 constrained amino acids,27, 31, 32 as well as N-methylation33, 34 synthetic modifications. These strategies share a common underlying theme of stabilizing secondary structure(s) and are commonly used to restrict peptide conformations, aid in conformational analysis, and often result in selective and potent ligands. As a complementary strategy, our laboratory examined the pharmacological and structural consequences of incorporating a bioactive heterocyclic moiety35 into a cyclic peptide template36–38 (Figure 1) in attempts to increase structural backbone rigidity and probe the His-Phe-Arg side chain orientations.39 Unexpectedly, the conclusions from that study were that more conformationally flexible ligands resulted [as determined by 2D 1H NMR and computer assisted molecular modeling (CAMM)] compared to the control cyclic peptide devoid of the heterocyclic backbone moiety. That study did provide some interesting SAR insights but also resulted in the identification of a nM full agonist template that could be used for additional SAR experiments to probe the importance of the His-Phe-Arg-Trp side chain orientation. Herein, we have further utilized the lead template (Figure 1, control ligand 2) and performed additional SAR studies by maintaining the heterocyclic template constant and modifying the side chains at the R1 (His), R2 (Phe), R3 (Arg), and R4 (Trp) positions. The initial design strategy was to compare an Ala and Pro amino acid scan at each of the four positions to identify the pharmacological consequences of these modifications at the melanocortin receptors examined. Unfortunately, we were unable to obtain the Pro containing derivative at the Trp position due to steric constraints during the synthesis. The second design strategy incorporated into this study includes the incorporation of amino acid side chain modifications (Figure 2) that have been previously identified as important for melanocortin receptor selectivity, and antagonist versus agonist MC3R and MC4R pharmacological profiles. These amino acid modifications resulted in several compounds with low nM potencies and mixed MC3R/MC4R pharmacological profiles at the mouse melanocortin receptor subtypes. The compounds with DPhe substitutions were selected for biophysical conformational experiments (2D 1H NMR and CAMM) with the goal to correlate structure with mixed MC3R/MC4R pharmacological profiles and attempt to differentiate the structure of the MC3R antagonist/agonist versus MC4R full agonist ligands.
Figure 1.

Illustrates the control ligand 2 incorporating the bioactive heterocycle (pink) into the peptide template. The amino acid substitutions at each of the R positions [R1=His (Green), R2=DPhe (Orange), R3=Arg (Blue), R4=Trp (Red) of the melanocortin ligand pharmacophore domain are indicated. The library design approach was based upon the positional scanning method where one R position is modified while holding the other positions constant.
Figure 2.

Structure of amino acid building blocks used in this study.
Results
Chemistry
The ligands examined herein were either synthesized manually using standard Fmoc synthetic methodology as reported39 or using a microwave synthesizer (Discover SPS™, CEM Corp.), Scheme 1. Microwave synthetic experimental conditions facilitated amino acid coupling and Nα 9-fluorenylmethoxycarbonyl (Fmoc) deprotection reactions in a more time efficient manner (Fmoc removal: 30 min under conventional conditions to 4 min using the microwave; amino acid coupling: 60–120 min to 5 min using the microwave). Use of the microwave versus the manual synthetic approach reduced the overall reaction time significantly from 50 h to 15 h without affecting the yield of the crude molecules. Cleavage of the compounds from the resin and removal of the side chain protecting groups was achieved by a 3 h treatment with the cleavage cocktail (TFA/triisopropylsilane/EDT/water 91:3:3:3) at room temperature. The disulfide bond was formed after cleavage by dissolving the crude product in 20% DMSO/water and mixing at room temperature. Progress of the disulfide cyclization was monitored by analytical reversed-phase high-pressure liquid chromatography (RP-HPLC) at a flow rate of 1.5mL/min (λ = 214 nm) and was generally completed in 24–36 hrs. The peptides were purified to homogeneity using semi-preparative RP-HPLC and a flow rate of 5mL/min. The peptides possessed the correct molecular weights as determined by mass spectrometry. The purity of these peptides (>95%) was assessed by analytical RP-HPLC in two diverse solvent systems (Supplemental materials).
Scheme 1.

Reagents and conditions: a) 20% Piperidine/DMF; b) 3 eq Fmoc-Xaa-OH, 3 eq BOP or HBTU, 6 eq DIEA, DMF; c) Fmoc-Yaa-aldehyde, NaBH3CN, AcOH, DMF; d) Fmoc-Cys(SBut)-OH, BOP or HBTU, DIEA, DMF; e) (ClCH2CO)2O, NEM, DCM; f) Bu3P/H2O/THF; g) NEM, DMF, heat 55–60°C; h) TFA/TIS/EDT/H2O 91:3:3:3
Melanocortin Receptor Pharmacology
All the compounds were tested for agonist or antagonist functional activity at the mouse MC1, MC3, MC4, and MC5 melanocortin receptors using a cAMP based β-galactosidase reporter gene bioassay.40 The MC2R is only stimulated by the endogenous ACTH agonist and was excluded from the present study. The NDP-MSH (Ac-Ser-Tyr-Ser-Nle-Glu-His-DPhe-Arg-Trp-Gly-Lys-Pro-Val-NH2) peptide30 is one of the standard melanocortin agonists used in the study of these receptors and is included herein as a reference control and as an internal control to for maximal ligand efficacy (100%). For the compounds examined in this study, with the exceptions of 9 and 11, which are partial agonists and antagonists at the mMC3R (Figure 3), fully efficacious agonists resulted (Table 1). The super potent MTII peptide [Ac-Nle- c[Asp-His-DPhe-Arg-Trp-Lys]-NH2]41, 42 is used as the agonist in the Schild antagonist bioassays43 to assess the potency of the mMC3R antagonists instead of α-MSH. The reason is that MTII is ca 10–100-fold more potent than α-MSH and when performing antagonist assays on weakly potent compounds (i.e. compound 9), the use of the α-MSH agonist may not be able to generate the required three agonist shifted dose-response curve EC50 values in the presence of increasing concentrations of antagonist peptide. A minimum of three dose-response shifted EC50 value data points are needed to generate a straight line in order to determine the pA2 value according to the Schild method.43 As additional controls, the peptide template 1,38 that does not contain the heterocyclic moiety, as well as the previously reported lead template 239 have been included. All the control values are consistent with previously reported values and are within the inherent 3-fold experimental error associated with this assay in our laboratory over the past decade.
Figure 3.

Melanocortin receptor pharmacologically of the control NDP-MSH peptide and analogues 6–11 that were modified at the DPhe position. Agonist profiles at the mMC1R and mMC3-5Rs are shown in the top four panels. The mMC3R antagonist profiles for molecules 9 and 11 are shown in the bottom two panels. Both ligands 9 and 11 also possess partial mMC3R agonist activity at the higher concentrations examined.
Table 1.
Agonist EC50 (nM) and antagonist pA2 pharmacology of the modified peptides at the mouse melanocortin receptors.
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ligand | mMC1R | Fold | mMC3R | Fold | mMC4R | Fold | mMC5R | Fold | ||||
|
| ||||||||||||
| R1 | R2 | R3 | R4 | EC50 (nM) | Diff | EC50 (nM) | Diff | EC50 (nM) | Diff | EC50 (nM) | Diff | |
| NDP-MSH | 0.05±0.02 | 0.23±0.03 | 0.20±0.02 | 0.28±0.04 | ||||||||
| Control Peptide 1 (AMW3-130) | 0.30±0.18 | 1.70±0.33 | 0.17±0.04 | 1.45±0.63 | ||||||||
| 2 | His | DPhe | Arg | Trp | 2.4±0.5 | 1.0 | 324±44 | 1.0 | 190±20 | 1.0 | 46±11 | 1.0 |
| 3 | Ala | DPhe | Arg | Trp | 52±20 | 22 | 4020±745 | 12 | 705±100 | - | 690±140 | 15 |
| 4 | Phe | DPhe | Arg | Trp | 1.94±0.55 | - | 4500±1950 | 14 | 290±22 | - | 415±65 | 9 |
| 5 | Pro | DPhe | Arg | Trp | 32±17 | 13 | 1900±810 | 6 | 240±6 | - | 270±135 | 6 |
| 6 | His | Pro | Arg | Trp | 5000±2900 | 2080 | 10350±3800 | 32 | 5400±1040 | 28 | 1100±530 | 24 |
| 7 | His | Ala | Arg | Trp | 12500±9900 | 5208 | 7720±3900 | 24 | 20100±4200 | 106 | 7700±2200 | 167 |
| 8 | His | DNal(1′) | Arg | Trp | 7.4±0.9 | - | 190±40 | - | 130±24 | - | 205±100 | - |
| 9 | His | DNal(2′) | Arg | Trp | 8.7±1.3 | - | Antagonist pA2=7.6±0.2 partial agonist (20% maximal efficacy) | (9) | 190±40 | - | 46±7 | - |
| 10 | His | (pI)DPhe | Arg | Trp | 0.50±0.08 | (4) | 49±8 | (6) | 20±3 | (9) | 9.8±4.7 | (4) |
| 11 | His | DBip | Arg | Trp | 1.1±0.8 | - | Antagonist pA2=9.9±0.4 partial agonist (10% maximal efficacy) | (2492) | 760±220 | - | 87±16 | - |
| 12 | His | DPhe | Ala | Trp | 19±7 | 8 | 8070±2400 | 25 | 1300±340 | 6 | 850±74 | 18 |
| 13 | His | DPhe | Lys | Trp | 30±14 | 12 | 2540±360 | 7 | 420±63 | - | 460±60 | 10 |
| 14 | His | DPhe | hArg | Trp | 15±3.9 | 6 | 1830±490 | 5 | 965±442 | 5 | 350±145 | 7 |
| 15 | His | DPhe | Pro | Trp | 16±2 | 6 | 7120±3510 | 22 | 1270±280 | 7 | 630±144 | 14 |
| 16 | His | DPhe | Arg | Ala | 80±20 | 33 | 2230±610 | 7 | 210±57 | - | 125±41 | - |
| 17 | His | DPhe | Arg | Nal(2′) | 6.2±1.0 | - | 200±13 | - | 220±72 | - | 52±19 | - |
| 18 | His | DPhe | Arg | DNal(2′) | 14±4 | 5 | 180±12 | - | 50±9 | (4) | 29±20 | - |
| 19 | His | DPhe | Arg | Bip | 20±6 | 8 | 230±30 | - | 980±170 | 5 | 830±215 | 18 |
The indicated errors represent the standard error of the mean determined from at least three independent experiments.
The potency of the antagonist was determined by using the equation pA2 = −Log Ki where Ki is estimted as the ligand potency in nM.
A parentheses () indicates that an increase in potency has resulted.
Changes less than 3-fold are considered to be within the inherent experimental assay error.
One of the first experiments we performed was to determine the effect of an Ala side chain replacement scan at each of the His-DPhe-Arg-Trp positions in the molecular control template 2.39 Substitution of the His with Ala resulted in no change in potency at the mMC4R with 12- to 22-fold reduced agonist potency at the mMC1R, mMC3R, and mMC5R (Table 1). Replacement of the DPhe side chain with the corresponding methyl of the Ala amino acid in 7 however resulted in 24- to >5000-fold decreased ligand potency at the melanocortin receptors with the most notable decreased potency at the mMC1R. The reduced potency of this ligand may be associated with the side chain modification, but also the L-stereochemistry versus the D- in the control molecule at this position. Incorporation of the Ala side chain at the Arg R3 position in analogue 12 resulted in 6- to 25-fold decreased melanocortin receptor potency. Removal of the Trp indole side chain and replacement by the Ala methyl side chain in analogue 16 resulted in no notable decreased potency at the mMC4 and mMC5 receptors and approximately a 7- to 33-fold reduced potency at the mMC3R and mMC1R, respectively.
The His residue has been previously studied in pentapeptides,44–49 tetrapeptides50 and other peptide templates.21, 51 Herein, the His side chain was substituted with Ala, discussed above, as well as with the Phe (4) and Pro (5) side chains. The Phe substitution was a well-tolerated modification that resulted in equipotent agonist pharmacology at the mMC1R and mMC4R, as compared to the control template 2. At the mMC3R, substitution of the benzyl moiety in 4 resulted in 14-fold decreased potency but only 9-fold decreased mMC5R potency. The Pro substitution at the His position (5) resulted in equipotent mMC4R agonist potency, ca 6-fold decreased agonist potency at both the mMC3R and mMC5R, and 13-fold decreased mMC1R potency as compared to control 2.
The Phe side chain in the melanocortin peptide ligands has been a key feature of SAR experiments and has resulted in the conversion of different agonist scaffolds into compounds possessing antagonist and partial agonist activities at the MC3 and MC4 receptors. Such key modifications include β-(2-naphthyl)-D-alanine [DNal(2′)] in SHU911926 as well as (pI)DPhe.37, 52, 53 Replacement of the Phe side chain with the constrained Pro side chain in 6 resulted in 24- to 32-fold decreased potency at the mMC3-5 receptors with a significant drop in agonist potency at the mMC1R of ca 2000-fold, as compared to control 2. The Ala substitution containing ligand 7, as discussed above, resulted in reduced potency as well at all the melanocortin receptors examined and could be a combination of stereochemical inversion (L- instead of D-) as well as the methyl side chain substitution. Interestingly, the DNal(1′) substitution of compound 8 resulted in equipotent pharmacology at the melanocortin receptors examined as compared to the control 2. Substitution of DPhe with DNal(2′) in 9 resulted in a potent (25nM) mMC3R antagonist that possessed equipotent agonist pharmacology as control 2 at the mMC1, mMC4, and mMC5 receptors. The (pI)DPhe containing molecule 10 resulted in a full agonist ligand at the melanocortin receptors with ca 4 to 9-fold increased potency at the mMC1R, mMC3R, mMC4R, and mMC5R, as compared to control 2. Molecule 11 with the DPhe side chain replaced by the DBip (Figure 2) residue, resulted in equipotent agonist pharmacology at the mMC1R, mMC4R, and mMC5R while unexpectantly resulting in a 0.13 nM mMC3R antagonist/slight partial agonist that is 2400-fold more potent than control 2. Additionally, this ligand is a >5800-fold mMC3R selective antagonist/slight partial agonist (Figure 3) versus the mMC4R.
Modification of the Arg side chain residue by the Ala (12), Lys (13), homoArg (14), and Pro (15) side chains resulted in a range of agonist activities from no changes in ligand potency to ca 25-fold decreased agonist potency depending upon the ligand and melanocortin receptor (Table 1).
Substitution of the Trp residue with Ala (16), Nal(2′) (17), DNal(2′) (18), and Bip (19) amino acids resulted in up to 33-fold decreased agonist potency at the mMC1R as compared to 2. At the mMC3 and mMC4 receptors, these Trp modifications did not significantly affect potency as compared to the control ligand 2. At the mMC5R, 19 resulted in 18-fold decreased potency, but the other side chain modifications possessed equipotent agonist activity as compared to control.
Biophysical studies: 2D 1H NMR and computer assisted molecular modeling (CAMM) structures
Ligands studied in these experiments (2, 6, 8–11) were dissolved in 90% (v/v) acetonitrile-d3 and 10% H2O (pH = ~3.0) for proton NMR experiments. This solvent was selected based upon the criteria that 1) it must completely dissolve all the peptides examined that possessed different biophysical properties, 2) similar solvents have been used previously so that structural comparisons could be performed and discussed, and 3) the amide protons were well resolved in this system versus the other solvents attempted. Other solvents that were attempted for this study but did not result in all the peptides examined being fully dissolved include i) DMSO-d6, ii) CDCl3/DMSO, iii) CD3CN/H2O, iv) MeOH-d3, v) MeOH/DMSO, vi) H2O/D2O]. All two–dimensional (2D) total correlation spectroscopy (TOCSY), nuclear Overhauser effect (NOESY) and rotating-frame Overhauser effect (ROESY) spectra were recorded at 34°C using a 600 MHz Bruker NMR instrument. The watergate 3–9–19 sequence was applied for water suppression. Data were processed using NMR Pipe/NMR Draw software on a Silicon Graphics workstation. Sequential resonance assignment was performed by the combined use of 2D-TOCSY and NOESY spectra (ROESY was used as a complimentary approach to NOESY). Only molecule 7, containing the Ala substitution of DPhe, did not result in experimental spectra possessing resolved peaks necessary for proton assignment and structural information under the conditions described above for the other molecules and therefore was excluded from further biophysical NMR experiments.
The NOE intensities observed for NMR-derived proton assignments for peptide analogues 6, 8–11, control 2 are summarized in the supplemental materials. Despite the fact that only one amino acid at DPhe4 position is changing, the NOE pattern of connectivity is very different in all five experimental compounds. It was observed that dNN (i, i+8) and dNN (i, i+9) NOEs in analogues 6 and 10 are consistent with our earlier study.37 Compounds 9 and 11 have less NOE connectivity than observed for the other compounds examined. Differences between random coil (Δ-RC) shifts54 and experimental chemical shifts values of compounds 6, 8–11, control 2 are presented in the supplemental materials. The Δ-RC shift values in compounds 9 and 11 suggest well-ordered structures in both compounds. Both analogues 9 and 11 have mixed pharmacology at the mMC3R/mMC4R subtypes showing partial agonist/antagonist properties at the mMC3R and full agonist at the mMC4R (Figure 3). The deviations of chemical shifts value for the amino acid residues (His, DBip, Arg for analogue 11 and His, DNal(2′), Arg for analogue 9) from the melanocortin His-Phe-Arg-Trp message sequence are very similar in both compounds. Compounds with full agonist activity at all four melanocortin receptors 6, 8, 10, and control 2 have similar trends at the His, DPhe/Pro/DNal(1′)/(pI)DPhe and Ala residues. These data indicate that the compounds in this study are tending to form more ordered structures rather than random coils and possess subtle differences in structures for antagonist and agonist activity.
Figure 4 depicts the stereoview of sausage representations of the five experimental compounds 6, 8–11, control 2 analyzed by NMR and computer molecular modeling in the present study. The structures are represented as the superposition of the conformers of the major family of compounds aligned on backbone heavy atoms of residues 3–7. Cluster analysis of the molecular dynamics trajectory of compound 6 resulted in one major family with 49% of all the conformers. Analysis of the ϕ- ψ backbone dihedral angles didn’t result in the identification of any classical turn structure. Compound 8, which possesses DNal(1′) in place of DPhe, resulted in one major family with a population of 29% of the conformers. Using a representative structure of the major family 8, we identified a β-turn of type I′ spanning His and DNal(1′) residues. Additionally, we observed a cluster of hydrophobic residues DNal(1′)4, Trp7 and Phe10 at one side and hydrophilic residues Arg6 and Asn8 on the other side of backbone plane in ligand 8. Molecular modeling studies of α-MSH and other ligands previously suggested the stacking of the aromatic rings of Phe7 and Trp9 and orientation of the Arg8 side chains away from the His6, Phe7 and Trp8 side chains.55,56 Cluster analysis of compound 9 resulted in one major family that contains 27% of total conformers and five more families with populations less than 5% that did not possess any classical turn structure. The largest conformational family (25% of total) identified for compound 10 possessed a well-defined structure with residues His3, (pI)DPhe4 and Cys5 participating in a classic γ-turn secondary structure motif (Figure 4).57 Compound 11, which has a DBip residue in the place of DPhe, resulted in one main family possessing a 32% population of the conformers after cluster analysis. Further analysis indicated an inverse γ-turn encompassing Asn8, Ala9, and Phe10 residues and isolated beta-bridges were found at the Cys2 and Cys5 amino acids. The control compound 2 resulted in four families in this study. The major family has 21% of the conformer population and other three families have 13%, 6%, and 5%, respectively. The dihedral angle analysis of the major family indicated a hydrogen bonded turn structure spanning the Arg and Trp residues. Isolated β-bridges were also observed at the His and Phe amino acids. The root mean square superposition deviations of the backbone heavy atoms for the conformers from the major family to the lowest energy member are 0.78, 1.66, 1.67, 1.08, 2.84, and 0.97 Å for compounds 6, 8–11, as compared to control 2, respectively. The ϕ-ψ distribution of the putative pharmacophore residues His-DPhe-Arg-Trp is represented in Figure 5. All the ligand residues were included, except the terminal Tyr residues. The ϕ-ψ distribution patterns are very different in the pharmacophore region spanning the His, DPhe/Pro/DNal(1′)/DNal(2′)/(pI)DPhe/DBip, Arg and Trp residues. In particular, the DPhe substitutions and the Trp residues in all compounds indicate the most dramatic differences. Apart from pharmacophore region, Asn8, Ala9 and Phe10 have prominent differences in ϕ-ψ angle distribution patterns.
Figure 4.


Sausage stereoview representations of the representative major families members of compounds (A) control 2 (RMSD = 0.97 ± 1.01Å) (B) 6 (RMSD = 0.78 ± 0.26 Å), (C) 8 (RMSD = 1.66 ± 0.56 Å), (D) 9 (RMSD = 1.67 ± 0.53 Å), (E) 10 (RMSD = 1.08 ± 0.29 Å), and (F) 11 (RMSD = 2.84 ± 0.53 Å), aligned on the backbone heavy atoms of residues 3–7. The His side chain is indicated in green, the DPhe side chain in orange, the Arg side chain in blue, and the Trp side chain in red. A wide grey backbone indicates greater flexibility in that domain. Following restrained molecular dynamics (RMD) simulations for 10 ns, 200 equally spaced structures were energy minimized with the NMR based restraints. The energy minimized structures were grouped into conformational families by comparison of the back bone dihedral angels within the His-Phe-Arg-Trp domain. For comparative purposes, the energy ranges for each of the representative families illustrated is as follows. (A) control 2 33–110 kCals/mol, (B) 6 157–513 kCals/mol, (C) 8 208–233 kCals/mol, (D) 9 166–190 kCals/mol, (E) 10 173–197 kCals/mol, and (F) 11 196–252 kCals/mol.
Figure 5.

CAMM based ϕ- ψ angle distribution for ligands (A) control 2, (B) 6, (C) 8, (D) 9, (E) 10, and (F) 11 conformational families. The His residue is indicated in green, the DPhe residue in orange, the Arg residue in blue, and the Trp residue in red. All other residues are indicated in purple.
Discussion
The MC4R is expressed centrally in the hypothalamus region of brain and has been shown to regulate food intake upon i.c.v. administration of agonists (decrease food intake) and antagonists (increase food intake).11, 12, 58 The MC3R, which is expressed both centrally (including the hypothalamus) and peripherally,3, 5 has been postulated to play fundamental roles in metabolism and energy homeostasis,59, 60 as well as food intake.12 However, the physiological mechanism(s) of action for the MC3 receptor in the different tissues and the periphery are still unclear. In recent years, the role of the MC3R in other functions like inflammation are becoming more evident.61 Based upon the unclear physiological roles of the melanocortin receptors alone or in synergistic combinations (heterodimerization),62–64 there is a continuing need for the development and identification of selective MC3 versus MC4 receptor agonists, antagonists, and/or mixed functional ligands. It has been established that it is difficult to design MC3R versus MC4R selective ligands based on conventional modifications using the MTII and SHU9119 molecular templates.49, 65–70 Even when it’s possible to get somewhat selective and potent ligands through other novel templates reported in the literature by other laboratories,69, 70 precise structural and conformational features that differentiate SAR between the MC3 and MC4 receptor subtypes is difficult to predict a priori. We continue these efforts in the present study to probe ligand structural features that can differentiate the ligand-receptor pharmacological profiles of the melanocortin receptors (MC1, MC3-5R). In a previous study we reported a novel hybrid peptide-heterocyclic moiety template that resulted in a potent nM MC4R analogue, control 2 herein.39 This compound, however, was not selective towards the melanocortin MC3 and MC4 receptor subtypes. Based on these earlier results, and reports from other laboratories, we designed the present library by replacing the putative His-Phe-Arg-Trp side chain pharmacophores with the natural and unnatural amino acid side chains shown in Figure 2. These selected amino acid side chains have been previously identified to increase melanocortin receptor selectivity, potency, and agonist or antagonist pharmacology.26, 37, 41, 52, 71, 72
Melanocortin-1 Receptor
The MC1R is primarily expressed in melanocytes and leukocytes and is involved in skin and hair pigmentation. Mounting experimental evidence suggests that the MC1R may also be involved in pain modulation, inflammation, and control of the immune system.1, 2, 8, 73–75 Substitution of His with Ala (3) resulted in 22-fold decreased potency at the mMC1R, as compared with the lead control compound 2. Substitution of His with Phe (4) in the cyclic peptide-heterocyclic template gave equipotent agonist activity at this receptor subtype which is in contrast to the AGRP-melanocortin chimeric peptide template KAR Tyr-c[β-Asp-His-DPhe-Arg-Trp-Asn-Ala-Phe-Dpr]-Tyr-NH2,37 which resulted in 27-fold decreased activity when the His residue was replaced with a Phe (Figure 6). The Phe substitution of His in tetrapeptide Ac-His-DPhe-Arg-Trp-NH2 template also resulted in 25-fold decreased mMC1R activity, consistent with the KAR template.37 Incorporation of a Pro residue in place of the His (5) resulted in 13-fold decreased potency as compared to the control template 2, unlike the 100-fold decreased potency observed in the chimeric KAR peptide.37 This SAR at the His position demonstrates that His side chain at position 3 is not critical in the present template for MC1R activity.
Figure 6.

Histogram comparing the relative fold change in EC50 values of the indicated amino acid relative to the appropriate control ligand at the mMC1R. Color coding- the black bar represents the tetrapeptide template Ac-His-DPhe-Arg-Trp-NH2. The blue bar represents the KAR template Tyr-c[β-Asp-His-DPhe-Arg-Trp-Asn-Ala-Phe-Dpr]-Tyr-NH2. The red bar represents the control ligand 2 in this study. ‡ Signifies that this amino acid substitution was not reported in one of the templates used for the comparative analysis in this study. @ Indicates that the modification results in an increase in potency and is shown as bars pointing down.
Replacement of the DPhe side chain at position 4 with Ala (peptide 7) resulted in 5000-fold decreased activity at the MC1R. The Pro substitution (6) resulted in ca 2000-fold decreased potency. These values are consistent with earlier reports for tetrapeptides (Figure 6).37–39, 52 Since the L-configuration in 6 and 7 was used, as opposed to the D- in the parent compound, this may also be a factor for the resulting reduced potency by this modification. Substitution with the “bulkier” side chain DNal(1′) compound 8, DNal(2′) compound 9, and DBip compound 11 resulted in equipotent MC1R agonist activity compared to the control 2. Additionally, compound 10 with the (pI)DPhe side chain instead of just the DPhe phenyl moiety resulted in a modest, ca 4-fold more potent, compound at the mMC1R. These results indicate that an aromatic group at this position is important for activity at the mMC1R, but bulky aromatic residues do not show any additional benefits.
Substitution of the Arg and Trp side chains did not give any dramatic changes with the exception of the Trp to Ala modification (compound 16) that resulted in 33-fold decreased mMC1R activity. Other side chain substitutions resulted in nearly equipotent mMC1R analogues as compared to the control reference molecule. Interestingly, replacement of the Arg side chain didn’t have any effect on mMC1R activity in this template, which is in contrast to some previous reports (Figure 6).71, 76 Results for Trp substitutions in the currently used template are consistent with the chimeric peptide template KAR that does not contain the heterocyclic moiety.
Melanocortin-3 Receptor
Alanine scanning of the His, DPhe, Arg, and Trp residues in the peptide-heterocycle template 2 resulted in 7- to 25-fold decreased potency at the MC3R. All of these compounds, 3, 7, 12, and 16 resulted in full agonists with μM EC50 values (Table 1). Substitution of His with Pro (5) and Phe (4) did increase MC4R versus MC3R agonist ligand selectivity, consistent with previous reports,44, 50 but were different than previous results using the chimeric peptide template KAR, which resulted in mMC3R antagonists (Figure 7).37 Compound 6, with Pro substitution of the DPhe, resulted in a full MC3R agonist with 32-fold decreased activity, with a similar agonist potency of 13200 nM as observed in the KAR template.37 Both compounds 6 and 7 retained full agonist activity at the MC3R, which is in contrast with the studies on melanocortin-based peptide templates where it was concluded that replacement of Phe at the seventh position with anything other than aromatic or “bulky” residue would result in loss of ligand activity (Figure 7). Modification of the DPhe phenyl ring with the with DNal(1′) modification in the chimeric template KAR resulted in 330-fold decreased mMC3R potency,37 whereas compound 8 resulted in equipotent ligand activity at mMC3R. Compound 9, with the DNal(2′) modification of the DPhe side chain in the template used herein resulted in a MC3R antagonist, consistent with previous reports for this substitution in other melanocortin-based peptide templates.26, 77, 78 This compound also had partial MC3R agonist activity (Figure 3). The same Phe-DNal(2′) substitution in the chimeric melanocortin-AGRP template KAR also resulted in an antagonistic pharmacological profile at the MC3R (Figure 7). However, this substitution has a different profile at MC4R than previously observed and will be discussed below. Surprisingly, compound 10 with the addition of the para-I onto the phenyl DPhe side chain, resulted in a full agonist with ca 6-fold increased potency as compared to the control 2 at MC3R. Compound 11, with the DBip residue in place of the DPhe, resulted in mixed pharmacology. At the mMC3R it is an antagonist with slight partial agonist activity and a full agonist at the mMC4R (Figure 3). Additionally, 11 is 5800-fold selective in potency for the mMC3R over the mMC4R (Table 1).
Figure 7.

Histogram comparing the relative fold change in EC50 values of the indicated amino acid relative to the appropriate control ligand at the mMC3R. For the color coding scheme, refer back to the legend in Figure 6. ‡ Signifies that this amino acid substitution was not reported in one of the templates used for the comparative analysis in this study. *Indicates that the amino acid change in the template resulted in an antagonist. @ Indicates that the modification results in an increase in potency and is shown as bars pointing down.
Melanocortin-4 Receptor
All substitutions made at the His, DPhe, Arg, and Trp positions resulted in full agonist activity at the mMC4R, in contrast to previous SAR studies in the KAR and tetrapeptide templates (Figure 8).37, 52 Substitution with Ala (3), Phe (4), or Pro (5) at the His position, DNal(1′) (8), DNal(2′) (9), DBip (11) at the DPhe, Lys (13) at the Arg and Ala (16) or Nal(2′) (17) at the Trp position in the present template resulted in equipotent agonist profiles at the mMC4R, as compare with the control 2. It has been reported previously in the literature that modification at the His side chain position could result in MC4R versus MC3R selectivity.44, 50, 78, 79 In the peptide-heterocyclic template examined herein, replacement of imidazole side chain of His with the amino acids Ala, Phe, and Pro resulted in a modest reduction in MC3R agonist activity, equipotent MC4R, and minor (up to 15-fold) MC4R versus MC3R selectivity. Unexpectedly,26 substitution of DPhe with DNal(2′) in the template examined herein resulted in an equipotent MC4R agonist. The same modification in the peptide template KAR, lacking the heterocyclic moiety, resulted in a partial agonist/antagonist pharmacological profile at the mMC4R (Figure 8).37 In previous studies involving linear or cyclic melanocortin peptide templates, substitution of the Phe to the DNal(2′) resulted in potent MC4R competitive antagonists devoid of any partial agonist activity. Specifically, modification of the DPhe7 of melanocortin ligands (α-MSH numbering) such as MTII (Ac-Nle-c[Asp-His-DPhe-Arg-Trp-Lys]-NH2),23, 41 resulted in the discovery of the MC3R antagonist/partial agonist and MC4R antagonist SHU9119 (Ac-Nle c[Asp-His-DNal(2′)-Arg-Trp-Lys]-NH2).26 The postulated mechanism of MC4R antagonism by incorporation of bulky DNal(2′)-containing analogues was proposed based upon previous reports of SAR, computational molecular modeling, and receptor mutagenesis studies of the MC4R.26, 80 Receptor mutagenesis studies resulted in the hypothesis that the DNal(2′) side chain might hinder side-chain rotation of the conserved MC4R Trp258 amino acid in transmembrane helix 6 and Leu133 in transmembrane helix 3 (hMC4R numbering), thus “locking” the receptor into the “inactive” state.72, 81 Similarly, Chen et al. postulated that in a similar manner, the hMC3R L165 in TM3 interaction upon peptide ligand binding is critical for agonist or antagonist selectivity.82 These hypotheses, that bulky substituents at the ligand DPhe7 (α-MSH numbering) position may be responsible for antagonism at the MC3 and MC4 receptors,26 is applicable in the present template for the MC3R as well. However, the ligands examined herein result in MC4R pharmacological profiles inconsistent with this originally proposed hypothesis (Figure 8). Compound 10, with addition of the p-Iodo moiety to the DPhe amino acid side chain in the control 2 peptide-heterocycle template resulted in a 9-fold more potent agonist at the mMC4R as compared to control 2. Interestingly, in the chimeric peptide template KAR, the linear NDP-MSH, and cyclic MTII templates, this addition of the pI moiety to the DPhe side chain resulted in mixed partial agonist and/or antagonist MC4R pharmacology,26, 80 while the tetrapeptide Ac-His-(pI)DPhe-Arg-Trp-NH2 is a potent MC4R agonist.52, 53 Thus, at the MC4R, compound 10 is consistent with the tetrapeptide melanocortin template that incorporates (pI)DPhe at the seven position (α-MSH numbering) (Figure 8).
Figure 8.

Histogram comparing the relative fold change in EC50 values of the indicated amino acid relative to the appropriate control ligand at the mMC4R. For the color coding scheme, refer back to the legend in Figure 6. ‡ Signifies that this amino acid substitution was not reported in one of the templates used for the comparative analysis in this study. *Indicates that the amino acid change in the template resulted in an antagonist. @ Indicates that the modification results in an increase in potency and is shown as bars pointing down.
Modification at the Arg position of template 2 with Ala (12), hArg (14), and Pro (15) resulted in 5- to 7-fold decreased mMC4R potency. Replacement of the Arg side chain with Lys (13) resulted in an equipotent analogue for mMC4R. Modification at the Arg position in the present template resulted in MC4R pharmacology different than the previously reported melanocortin based templates (Figure 8).37, 71, 79, 83 The functional role of the Trp indole side chain in the control template 2 was explored by substitution with following amino acids: Ala (16), Nal(2′) (17), DNal(2′) (18), Bip (19). All these modifications resulted in nearly equipotent activity at the mMC4R, consistent with previous studies of other peptide templates (Figure 8), with the exception of 19.48, 72, 84
Melanocortin-5 Receptor
The MC5R mRNA has been identified to be expressed in peripheral and central locations and tissues.7, 85, 86 Studies in knockout mice indicated that the MC5R has a role in exocrine gland function,86 but other physiological functions for this receptor in the body are not fully elucidated. In the heterocyclic-peptide template studied herein, replacement of the His side chain with Ala (3), Phe (4), and Pro (5) resulted in 6- to 15-fold decreased activity at the mMC5R, similar to the chimeric peptide template KAR (Figure 9).37 Notably however, a difference between the templates exists for the His to Ala substitution which is equipotent in the peptide KAR template, but is 15-fold less potent for 3 at the mMC5R compared to control 2.37 Studies of the tetrapeptide melanocortin template with similar substitutions resulted in greater reduction in potency (170-fold when His is replaced by Pro).50 These comparisons in pharmacological profiles at the mMC5R suggest that in the chimeric melanocortin-AGRP template KAR, independent of the heterocyclic moiety, the His residue might not be a major potency determining factor at the mMC5R. Substitution of DPhe by Ala (7) resulted in ca 170-fold decreased activity at the mMC5R. Compounds 8; DNal(1′), 9; DNal(2′), 10; (pI)DPhe and 11; (DBip) showed equipotent response at the mMC5R compared with the control 2 template. Substitution at the DPhe with Pro resulted in 24-fold decrease activity at the mMC5R. Replacement of the Arg side chain with Ala (12), Lys (13), hArg (14), and Pro (15) resulted in 8- to 18-fold decreased activity at the mMC5R. In the chimeric peptide template KAR, similar substitutions resulted in 6- to 8600-fold reduced activity (Figure 9).37 Although the present template 2 and chimeric peptide template KAR have similar mMC5R pharmacological patterns with similar amino acid modifications at the Arg side chain, the peptide-heterocycle template 2 has less dramatic changes in potency than the KAR template, but appear to be distinct from other melanocortin templates (Figure 9).37, 71, 79 Specifically, the tetrapeptide Ac-His-DPhe-Arg-Trp-NH2 resulted in 1740-, 11800-, and 80-fold decreased activity upon substitution of Arg with Ala, Pro and Lys respectively.71 Substitution of Trp amino acid side chain with Ala (16), Nal(2′) (17) and DNal(2′) (18) resulted in equipotent agonist potency at the mMC5R, as compared with control template 2, whereas compound 19 is 18-fold less potent. This template has a different pharmacological profile at the mMC5R with Trp substituted by Ala in comparison to chimeric template KAR, which resulted in a peptide with 135-fold decreased mMC5R potency (Figure 9). Substitution of Trp in the heterocyclic-peptide template examined herein with Nal(1′) and DNal(2′) is consistent with similar substitutions at the tetrapeptide template, however the Bip residue substitution resulted in different MC5R pharmacology in the tetrapeptide template.71
Figure 9.

Histogram comparing the relative fold change in EC50 values of the indicated amino acid relative to the appropriate control ligand at the mMC5R. For the color coding scheme, refer back to the legend in Figure 6. ‡ Signifies that this amino acid substitution was not reported in one of the templates used for the comparative analysis in this study. *Indicates that the amino acid change in the template resulted in an antagonist. @ Indicates that the modification results in an increase in potency and is shown as bars pointing down.
In summary, these structure-activity relationship studies presented herein with the peptide-heterocyclic template demonstrate that this is a unique template that possesses distinct mouse melanocortin receptor pharmacological profiles. Some of these ligands may be used to investigate the structural determinants that discriminate between the MC3 and MC4 receptor subtypes.
1H NMR and Computer-Assisted Molecular Modeling Based Structural Studies
Based upon the distinct mMC3R and mMC4R pharmacological profiles of the DPhe ligand modifications in this heterocyclic-peptide template (Table 1), the six DPhe substituted analogues (6–11) and control 2 ligand were selected for further biophysical analysis using 2D 1H NMR and computer assisted molecular modeling studies to attempt to correlate structure with function. Compounds 6, 8, and 10 are full agonists at all four melanocortin receptors MC1R and MC3-5R. Compounds 9 and 11 are mMC3R antagonists with partial agonistic activity (Figure 3) and full mMC4R agonists. These compounds can be compared with similar substitutions in other NMR based structural studies of melanocortin ligands MTII and SHU9119. The template examined herein, 2, consists of a 10–membered heterocycle imbedded into a chimeric peptide 1 postulated to possess different backbone structure in the pharmacophore region, from the typical melanocortin based peptide template. The objective of this study was to compare results of the structural studies presented herein in the context of previous reports to determine any overlapping and/or distinct structural features in attempts to explain differences in ligand-receptor SAR and MC3R/MC4R pharmacological profiles. In our previous study, incorporation of the bioactive turn mimetic into the chimeric AGRP-melanocortin peptide (1) was based upon the hypothesis that incorporating the heterocycle into the peptide backbone would increase structural rigidity.39 The ligands reported in our previous study used a positional scanning approach that examined both the location of the heterocycle turn in relationship to the His-DPhe-Arg side chains as well as a stereochemistry modification of the heterocycle. The control peptide 1 and the most potent ligand identified in that study (referred to as 2 in this study) served as the starting template for the SAR examined in this study. Surprisingly, while the initial hypothesis that incorporating a constrained heterocycle into the backbone template of 1 would produce a more structured biomolecule, 2D 1H NMR and CAMM studies resulted in the observation that the ligand 2 was actually more structurally flexible as compared to 1.39 Consistent with our previous study,39 the lead ligand 2 resulted in multiple observed conformational families that were not highly populated. The most populated family was observed to possess 21% of the sampled conformers, whereas the other three remaining families contained approximately 5% of the sampled structures. In contrast, three of the five DPhe substituted ligands (8, 10, and 11) were observed to possess either a β- or a γ-reverse turn secondary structure. The five analogues 6, 8, 9, 10, and 11, were observed to cluster into one major structural family, in contrast to the control ligand 2. These results are consistent with previous biophysical studies on various potent melanocortin peptide ligands that identified a reverse turn structure in His-DPhe-Arg-Trp region of the ligands.87–90
NMR & Modeling Based Structural Features of Agonist ligands
Compound 8 which contains a bulkier DNal(1′) residue in place of DPhe, is a full agonist at mMC1R and mMC3-5R and is equipotent to the control template 2 at all four melanocortin receptor subtypes. This analogue was characterized as possessing a β-turn involving the His3 and DNal(1′)4 residues. The chimeric peptide KAR compound was reported to possess a β-turn involving the Trp6 and Asn7 residues.37 NMR studies of melanocortin ligands have shown that a similar type of reverse β-turn may also present in the MTII agonist ligand around the His-DPhe-Arg-Trp message sequence.88 Compound 8 was identified to possess an amphiphilic side chain orientation (consistent with observations for other melanocortin analogues), in which the hydrophobic pharmacophore residues are oriented towards one plane which is different for the other hydrophilic amino acids.65, 88 Compound 10 with (pI)DPhe in place of DPhe resulted in ca a 5-fold increased activity at all four of the receptors and was characterized in this study to possesses a classic γ-turn with the (pI)DPhe residue at the center. Classic γ-turns have been found to generally occur at the end of β-hairpin structure where a hydrogen bond is formed between the backbone carbonyl oxygen of one residue (i) the backbone amide NH of the residue two positions further along the chain (i + 2). The classic γ-turn, less common than the abundant inverse γ-turn, and is differentiated only by inversion of the sign of ϕ and ψ angles. For the classic γ-turn, the idealized values are: ϕ+1 ~ 70.0 to 85.0, ψ+1 ~ −60.0 to −70.0, whereas for an inverse γ-turn they are: ϕ+1 ~ −70.0 to −85.0, ψ+1 ~ 60.0 to 70.0.57
Compound 6 with the Pro residue substituted for DPhe, resulted in 24- to 2000-fold decreased agonist potency at all the melanocortin receptors examined in this study. Although the backbone atoms around the pharmacophore (His-Pro-Arg-Trp) region were observed to posses a “bend” conformation, the φ - ψ angles were not consistent with any conventional turn structure. This result is consistent with a previous report where the Pro amino acid was substituted for the His residue in an MTII analogue.79, 91 The backbone heavy atoms for conformers from the major family of 6 are similar, indicating a structured domain, with the root mean square (RMS) superposition deviations for these conformers of 0.78 Å. As a result of the different backbone conformations in all the three compounds (6, 8, and 10), the putative side chains of the reference sequence His-Phe-Arg-Trp were observed to be oriented differently. Specifically, the Arg side chain of compound 6 was characterized to be oriented differently than observed in compounds 8 and 10 (Figure 4).
NMR & Modeling Based Structural Features of mMC3R Mixed Pharmacology (Antagonist/Partial Agonist) Ligands
Compound 11, containing the DBip amino acid in place of the DPhe residue resulted in mixed pharmacology with antagonist (pA2=9.9) and slight partial agonist activity at the mMC3R, while retaining full agonist potency at the mMC4R (~700nM) (Figure 3). This compound is more than 5800-fold selective at the MC3R versus the MC4R and was characterized to possess an inverse γ-turn involving the residues Asn8, Ala9 and Phe10 in this study (Figure 4). A γ-turn was previously reported in the peptide KAR template and postulated to explain the difference between the mMC3R and mMC4R pharmacological profiles.37 In the chimeric KAR peptide template, the Phe substitution for the His residue, resulted in mMC3R antagonist/partial agonist activity and full agonist activity at the mMC4R. Based upon the NMR biophysical studies, the Phe containing KAR derivative was postulated to contain a γ-turn secondary structure.37 Since the present template originates from the same chimeric template, it can be postulated that the γ-turn secondary structure is an important ligand structural feature that may differentiate between mMC3R antagonist and mMC4R agonist pharmacology. However, the second compound 9 with substitution of the DNal(2’) side chain instead of the DPhe side chain also resulted in a similar MC3R/MC4R pharmacological profile but did not result in the characterization of a “classical” turn in the pharmacophore region from these biophysical experiments. Both compounds 9 and 11 were observed to be generally more flexible than the other compounds from the biophysical experiments and are full agonist at MC1R, MC3-5R, consistent with the earlier study.37 The similar substitution in chimeric KAR peptide template, lacking the heterocyclic ring, resulted in a potent antagonist at the mMC3R and a partial agonist and antagonist at mMC4R. The mixed pharmacology was attributed to the equilibrium between two conformer families.37 The solution structure of synthetic MC3R/MC4R antagonist SHU9119 was reported to have type II β-turn, and the orientation of the His6 and DNal(2′)7 residues resulted in no observed aromatic stacking between these two side chains.88 The reduced potency or antagonistic activity of the SHU9119 was postulated to be the difference in the side-chain dispositions of His and DNal(2′).88 In the present template, as anticipated, introduction of the 10-membered heterocyclic ring around the ligand pharmacophore region changed the pharmacology and structure of compound 9. Unexpectantly 9, with the bulky aromatic DNal(2′) side chain in place of the DPhe residue, resulted in a full agonist at the mMC4R. Incorporation of the DNal(2′) residue at the DPhe position in other templates has resulted in competitive antagonists at the MC4R. Thus, these unanticipated data for 9 demonstrate that it has a different molecular interaction with the mMC4R receptor versus other templates possessing the DNal(2′) substitution at the DPhe position. These findings support the previous observation that the substitution of Phe by bulky aromatic groups could be decisive in maintaining the MC4R selectivity by enhancing hydrophobic interactions. The partial agonist/antagonist profile of 9 and 11 can be attributed to a more flexible backbone of these two compounds, as discussed above, that can facilitate binding of the ligands more easily in the mMC3R binding pocket as compared with other mMC3R agonist compounds.88
Conclusions
This study reports structure-activity relationship experiments of a hybrid heterocyclic-peptide template by substituting the putative melanocortin receptor peptide agonist pharmacophore (His-DPhe-Arg-Trp) residues to modify the pharmacological profile of a lead compound. The systematic substitution of amino acids with natural and unnatural amino acid residues (based on knowledge of earlier studies) results in novel pharmacological profile(s) at the mouse melanocortin receptors. The most notable results from this study include the identification of 11, a 5800-fold mMC3R antagonist/slight partial agonist versus the mMC4R full agonist pharmacological profile. This ligand is also a potent nM full agonist at the mMC1R that is expressed in the skin. Biophysical studies identified the presence of an inverse γ-turn in the ligand pharmacophore domain of ligand 11. This SAR presented herein will increase the general knowledge base of ligand-melanocortin receptor interactions as well as provide novel molecules to test in in vivo models. The conformational studies presented herein identify the structural differences between agonist and antagonist activity at the MC3/MC4 receptors.
Experimental Section
Reagents and General Methods
The peptide analogue 1 was synthesized using standard Fmoc methodology92, 93 in a manual reaction vessel. The amino acids Nα 9-fluorenylmethoxycarbonyl (Fmoc)-Cys(Trt), Fmoc-Tyr(tBu), Fmoc-Arg(Pbf), Fmoc-His(Trt), Fmoc-Phe, Fmoc-Asn(Trt), Fmoc-Ala, Fmoc-(1-naphthyl)-D-alanine [DNal(1′)], Fmoc-(2-naphthyl)-D-alanine [DNal(2′)], Fmoc-(2-naphthyl)-L-alanine [Nal(2′)], and the coupling reagents: benzotriazol-1-yl-N-oxy-tris-dimethylamino)phosphonium-hexafluoro-phosphate (BOP), O-Benzotriazole-N,N,N′,N′-tetramethyl-uronium-hexafluoro-phosphate (HBTU) and 1-hydroxybenzotriazole (HOBt) were purchased from Peptides International (Louisville, KY). The amino acids Fmoc-p-iodo-D-phenylalanine [(pI)DPhe], Fmoc-4-phenyl-D-phenylalanine (DBip), Fmoc-4-phenyl-phenylalanine (Bip), were purchased from Synthetech (Albany, OR). The amino acid Fmoc-hArg(Boc)2-OH was purchased from Advanced Chem Tech (Louisville, KY) and Fmoc-Arg(Boc)2-OH was purchased from Bachem (Torrance, CA). Rink amide p-Methylbenzhydrylamine Resin (p-MBHA Resin, 0.43 mequiv/g substitution) was purchased from Peptides International (Louisville, KY). Glacial Acetic acid (HOAc), Dichloromethane (DCM), methanol (MeOH), acetonitrile (MeCN), and anhydrous ethyl ether were purchased from Fisher (Fair Lawn, NJ). N,N Dimethylformamide (DMF) was purchased from Burdick and Jackson (McGaw Park, IL). Trifluoroacetic acid (TFA), piperidine, N,N-Diisopropylethylamine (DIEA), N,O-dimethylhydroxylamine, Lithium aluminum hydride (LiAlH4), 1,2-Ethanedithiol (EDT), Triisopropylsilane (TIS) were purchased from Sigma-Aldrich (St. Louis, MO). All reagents and chemicals were ACS grade or better and were used without further purification.
The modified peptides were synthesized using standard Fmoc-solid phase methodology at room temperature using manual reaction vessel (flat-bottom polyethylene syringes equipped with sintered Teflon filters, Teflon valves for flow control, and suction to drain the syringes from below) at room temperature or in a microwave (Scheme 1). Manual amino acid couplings consisted of the following steps: (i) removal of the Nα-Fmoc group by 20% piperidine in DMF (1 × 2 min, 1 × 18 min) (ii) a single 2 h coupling of Fmoc-amino acid (3 equiv) using BOP (3 equiv), HOBt (3 equiv), and DIEA (6 equiv) in DMF. The presence or absence of the Nα free amino group was monitored using the Kaiser test.94
The thioether ring moiety was assembled on the resin according to the general procedure previously reported.35, 39 The Fmoc-amino aldehydes were used directly without further purification in the next step as previously reported.39 Briefly the Fmoc-amino aldehydes were prepared by using 3 eq of the Fmoc amino acid reacted with 3.3 eq BOP and 4.4 eq DIEA for 30 min at rt. Then 3.3 eq NH(Me)OMe and 3.3 eq DIEA were added and mixed at rt for 3 hrs. Subsequently, 2 eq LiAlH4 in THF was stirred at −78°C for 1–2 hrs until the reaction was complete, followed by quenching with water. The thioether ring closure was performed at elevated temperatures (55–60°C) in an automated synthesizer (Advanced ChemTech 440MOS, Louisville, KY). After the completed synthesis, the peptides were cleaved from the resin and deprotected using a cleavage cocktail consisting of 91% TFA, 3.0% H2O, 3.0% EDT, and 3.0% TIS for 3 hrs at room temperature. After cleavage and side chain deprotection, the solution was concentrated and the peptide was precipitated and washed using cold (4 °C) anhydrous diethyl ether. The crude linear peptides were dissolved in 20% DMSO in water (1.0 mg/mL) and stirred at room temperature. Progress for the disulfide cyclization was monitored by UV-HPLC which was generally completed within 24–36 hrs. The resulting solution was lyophilized to yield the crude cyclic peptide and purified by reversed-phase HPLC [flow rate of 5 mL/min and gradients ranging (10–35mins) from 17%–45% acetonitrile/water 0.1% TFA] using a Shimadzu chromatography system with a photodiode array detector and a semi preparative RP-HPLC C18 bonded silica column (Vydac 218TP1010, 1.0 × 25 cm). The purified peptides were at least >95% pure as determined by RP-HPLC in two diverse solvent systems and had the correct molecular mass (Supplemental materials). Matrix-assisted laser desorption/ionization-Time of Flight (MALDI-TOF) mass spectrometry spectra were recorded on a Voyager instrument (University of Florida Protein Core Facility), using α-cyano-4-hydroxycinnamic acid as the matrix.
Solid Phase Peptide Synthesis using microwave irradiation
The Rink-amide MBHA resin was transferred into a 25 mL polypropylene reaction vessel (CEM) and “swelled” in dichloromethane for 1 h. The bottom cap of the vessel was removed after the 1 h swelling process and the vessel was transferred onto a vacuum filtration manifold. Deprotection of the Fmoc group was achieved in 20% piperidine in DMF by irradiating the sample at 75 °C, 30 W for 4 min in the CEM Discover SPS™ instrument. After cooling and washing the resin with DMF, a ninhydrin “Kaiser” test was performed.94 The amino acid coupling step was performed under the same microwave conditions (75 °C, 30 W) for 5 min using HBTU. The coupling of the Cys and His amino acids were performed at lower temperatures (50 °C, 30 W) for 5 mins. The heterocycle ring was assembled as previously described.39 The reductive alkylation, acylation of chloroacetic anhydride, and the deprotection of the thio tert-Butyl group steps were performed at the room temperature. The thioether bridge was formed using N-ethylmorpholine in DMF and the microwave conditions of (55 °C, 50 W) for 10 mins. The reagents were mixed by bubbling nitrogen gas into the reaction vessel. The iterative process of the Fmoc deprotection/coupling cycle was performed to elongate the peptide chain. After the final deprotection, the peptide was cleaved from the resin using the cleavage cocktail (91% TFA, 3.0% H2O, 3.0% EDT, and 3.0% TIS) at room temperature for 3 hrs. Note, as the peptides containing the heterocyclic moiety were synthesized to completion on resin, if racemization occurred during the isolated heterocycle formation as previously reported,35 it was not identified or purified in the major product peak collected by RP-HPLC.
Functional Bioassay
The HEK-293 cells stably expressing the mouse melanocortin receptors were maintained in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal calf serum and transfected with 4μg of the CRE/β-galactosidase reporter gene as previously described.40 Briefly, 5,000 to 15,000 post-transfection cells were plated into collagen treated 96-well plates (Nunc) and incubated overnight. Forty-eight hours post-transfection the cells were stimulated with 100 μL of peptide (10−4 to 10−12 M) or forskolin (10−4 M) control in assay medium (DMEM containing 0.1 mg/mL BSA and 0.1 mM isobutylmethylxanthine) for 6 h. The assay media was aspirated and 50 μL of lysis buffer (250 mM Tris-HCl pH=8.0 and 0.1% Triton X-100) was added. The plates were stored at −80 °C overnight. The plates containing the cell lysates were thawed the following day. Aliquots of 10 μL were taken from each well and transferred to another 96-well plate for relative protein determination. To the cell lysate plates, 40 μL of phosphate-buffered saline with 0.5% BSA was added to each well. Subsequently, 150 μL of substrate buffer (60 mM sodium phosphate, 1 mM MgCl2, 10 mM KCl, 5 mM β-mercaptoethanol, 2 mg/mL ortho-Nitrophenyl-β-galactoside [ONPG]) was added to each well and the plates were incubated at 37°C. The sample absorbance, OD405, was measured using a 96-well plate reader (Molecular Devices). The relative protein was determined by adding 200 μL of 1:5 dilution Bio Rad G250 protein dye:water to the 10 μ L cell lysate sample taken previously, and the OD595 was measured on a 96-well plate reader (Molecular Devices). Data points were normalized both to the relative protein content and non-receptor dependent forskolin stimulation. Maximal efficacy was compared to that observed for the NDP-MSH control peptide tested simultaneously on each 96-well plate. The antagonistic properties of the compounds examined in this study were evaluated by their ability to competitively displace the MTII agonist (Bachem) in a dose-dependent manner, at up to 10 μM concentrations. The antagonist pA2 values were generated using the Schild analysis method.43
Data Analysis
The agonist EC50 and antagonist pA2 values represent the mean of duplicate wells performed in three or more independent experiments. The EC50 and pA2 value estimates, and their associated standard errors, were determined by fitting the data to a nonlinear least-squares analysis using the PRISM software program (v4.0, GraphPad Inc.). The results are not corrected for peptide content.
NMR Spectroscopy
The ligand NMR samples were prepared (~1mM) by dissolving 1.0 mg of the purified peptides in a 500 μL solution of 90% (v/v) acetonitrile-d3 and 10% H2O, with an adjusted pH of ~3.0. The NMR data were collected at 34°C with a Bruker Avance II spectrometer operating at 600 MHz (using a cryoprobe) at the Advanced Magnetic Resonance and Imaging Spectroscopy (AMRIS) University of Florida facility. Standard proton TOCSY and NOESY 2D 1H NMR data were collected, processed, and analyzed as described previously.37, 39, 95 Acetonitrile was used as an internal standard (1.94 ppm). The chemical shifts of each of the peptides in this study were assigned using standard TOCSY and NOESY 1H-based strategies.96 This approach utilizes TOCSY spectra to identify resonances within a given amino acid and NOESY spectra to correlate one amino acid with the next through interactions of the α and/or β protons of residue i with the amide proton of residue i + 1.
Computer-Assisted Molecular Modeling (CAMM)
Proton-proton distances were calibrated using the well resolved methylene protons of the His residue based on the relationship r= rref × (ηref/η)1/6, where r is the distance between atoms, η is the NOESY cross-peak volume, rref is the known distance, and ηref is the corresponding volume of the NOESY calibration cross-peak. The NOE volumes were categorized as strong (1.8–3.0 Å), medium (1.8–3.5 Å), or weak (1.8–5.0 Å), (Supplemental materials). All conformational molecular modeling experiments were performed using the SYBYL v7.0 software from Tripos Inc. (St. Louis, MO) on a Silicon Graphics workstation. Restrained molecular dynamics (RMD) simulations were run in vacuo with a dielectric constant of 4.0, at a temperature of 500 K, and using the Tripos force field and Gastaiger-Hückel partial atomic charges. The peptides were initially built in a fully extended linear conformation. In the first step of modeling, RMD simulations were run for 1 ns. Following the initial 1 ns RMD trajectory, the cysteine residues were oriented next to each other, and disulfide bonds were manually formed and energy minimized without restraints. Finally, all the NMR based NOE restraints were included, and 10 ns RMD trajectories were collected. Following the RMD simulations, structures from 200 equally spaced points along the dynamics trajectory were energy minimized, analyzed, and grouped into conformational families. Cluster analysis was performed by comparing the backbone phi and psi angles of the His-DPhe-Arg-Trp amino acid ligand domain. This process identified representative structures (lowest energy conformers) of the conformational families that were used for further analysis.
Supplementary Material
Acknowledgments
This work has been supported in part by NIH Grants RO1DK064250 and R01DK091906.
List of Abbreviations
- ACTH
Adrenocorticotropin Hormone
- AGRP
Agouti-Related Protein
- ASIP
agouti-signaling protein
- CAMM
computer-assisted molecular modeling
- cAMP
cyclic 5′-adenosine monophosphate
- DCM
Dichloromethane
- DMF
N,N-Dimethylformamide
- DMS
Dimethylsulfide
- Fmoc
Nα 9-fluorenylmethoxycarbonyl
- GPCR
G Protein Coupled Receptor
- MC1R
Melanocortin-1 Receptor
- MC2R
Melanocortin-2 Receptor
- MC3R
Melanocortin-3 Receptor
- MC4R
Melanocortin-4 Receptor
- MC5R
Melanocortin-5 Receptor
- MCR
Melanocortin Receptor
- MeOH
Methanol
- MSH
Melanocyte Stimulating Hormone
- NOE
Nuclear Overhauser Effect
- POMC
Proopiomelanocortin
- φ
phi
- ψ
psi
- SAR
Structure Activity Relationship
- SEM
Standard Error of the Mean
- TFA
Trifluoroacetic acid
- TM
transmembrane
- α-MSH
Alpha-Melanocyte Stimulating Hormone
- β-MSH
Beta-Melanocyte Stimulating Hormone
- γ-MSH
Gamma-Melanocyte Stimulating Hormone
- μM
Micromolar
- RMD
restrained molecular dynamics
- NDP-MSH (4-Norleucine-7-D-Phenylalanine)
Ac-Ser-Tyr-Ser-Nle-Glu-His-DPhe-Arg-Trp-Gly-Lys-Pro-Val-NH2
- Control Peptide 1 AMW3-130
Tyr-c[Cys-His-DPhe-Arg-Trp-Asn-Ala-Phe-Cys]-Tyr-NH2
- MTII
Ac-Nle-c[Asp-His-DPhe-Arg-Trp-Lys]-NH2
- KAR
Tyr-c[β-Asp-His-DPhe-Arg-Trp-Asn-Ala-Phe-Dpr]-Tyr-NH2
References
- 1.Chhajlani V, Wikberg JES. Molecular Cloning and Expression of the Human Melanocyte Stimulating Hormone Receptor cDNA. FEBS Lett. 1992;309(3):417–420. doi: 10.1016/0014-5793(92)80820-7. [DOI] [PubMed] [Google Scholar]
- 2.Mountjoy KG, Robbins LS, Mortrud MT, Cone RD. The Cloning of a Family of Genes that Encode the Melanocortin Receptors. Science. 1992;257:1248–1251. doi: 10.1126/science.1325670. [DOI] [PubMed] [Google Scholar]
- 3.Roselli-Rehfuss L, Mountjoy KG, Robbins LS, Mortrud MT, Low MJ, Tatro JB, Entwistle ML, Simerly RB, Cone RD. Identification of a Receptor for γ Melanotropin and Other Proopiomelanocortin Peptides in the Hypothalamus and Limbic System. Proc Natl Acad Sci USA. 1993;90:8856–8860. doi: 10.1073/pnas.90.19.8856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mountjoy KG, Mortrud MT, Low MJ, Simerly RB, Cone RD. Localization of the Melanocortin-4 Receptor (MC4-R) in Neuroendocrine and Autonomic Control Circuits in the Brain. Mol Endo. 1994;8:1298–1308. doi: 10.1210/mend.8.10.7854347. [DOI] [PubMed] [Google Scholar]
- 5.Gantz I, Konda Y, Tashiro T, Shimoto Y, Miwa H, Munzert G, Watson SJ, DelValle J, Yamada T. Molecular Cloning of a Novel Melanocortin Receptor. J Biol Chem. 1993;268(11):8246–8250. [PubMed] [Google Scholar]
- 6.Gantz I, Miwa H, Konda Y, Shimoto Y, Tashiro T, Watson SJ, DelValle J, Yamada T. Molecular Cloning, Expression, and Gene Localization of a Fourth Melanocortin Receptor. J Biol Chem. 1993;268(20):15174–15179. [PubMed] [Google Scholar]
- 7.Gantz I, Shimoto Y, Konda Y, Miwa H, Dickinson CJ, Yamada T. Molecular Cloning, Expression, and Characterization of a Fifth Melanocortin Receptor. Biochem Biophys Res Commun. 1994;200(3):1214–1220. doi: 10.1006/bbrc.1994.1580. [DOI] [PubMed] [Google Scholar]
- 8.Lerner AB, McGuire JS. Effect of Alpha- and Beta-Melanocyte Stimulating Hormones on the Skin Colour of Man. Nature. 1961;189:176–179. doi: 10.1038/189176a0. [DOI] [PubMed] [Google Scholar]
- 9.Lindner E, Scholkens B. ACTH and alpha-MSH: cardiovascular and antiarrhythmic properties. Arch Int Pharmacodyn Ther. 1974;208(1):19–23. [PubMed] [Google Scholar]
- 10.Greenfield JR, Miller JW, Keogh JM, Henning E, Satterwhite JH, Cameron GS, Astruc B, Mayer JP, Brage S, See TC, Lomas DJ, O’Rahilly S, Farooqi IS. Modulation of Blood Pressure by Central Melanocortinergic Pathways. N Engl J Med. 2009;360(1):44–52. doi: 10.1056/NEJMoa0803085. [DOI] [PubMed] [Google Scholar]
- 11.Fan W, Boston BA, Kesterson RA, Hruby VJ, Cone RD. Role of Melanocortinergic Neurons in Feeding and the Agouti Obesity Syndrome. Nature. 1997;385:165–168. doi: 10.1038/385165a0. [DOI] [PubMed] [Google Scholar]
- 12.Irani BG, Xiang Z, Yarandi HN, Holder JR, Moore MC, Bauzo RM, Proneth B, Shaw AM, Millard WJ, Chambers JB, Benoit SC, Clegg DJ, Haskell-Luevano C. Implication of the Melanocortin-3 Receptor in the Regulation of Food Intake. Eur J Pharmacol. 2011;660(1):80–87. doi: 10.1016/j.ejphar.2010.10.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bertolini A, Vergoni W, Gessa GL, Ferrari W. Induction of Sexual Excitement by the Action of Adrenocorticotrophic Hormone in Brain. Nature. 1969;221(181):667–669. doi: 10.1038/221667a0. [DOI] [PubMed] [Google Scholar]
- 14.Wessells H, Fuciarelli K, Hansen J, Hadley ME, Hruby VJ, Dorr R, Levine N. Synthetic Melanotropic Peptide Initiates Erections in Men with Psychogenic Erectile Dysfunction: Double-blind, Placebo Controlled Crossover Study. J Urol. 1998;160(2):389–393. [PubMed] [Google Scholar]
- 15.Eipper BA, Mains RE. Structure and Biosynthesis of Pro-ACTH/Endorphin and Related Peptides. Endocrin Rev. 1980;1:1–26. doi: 10.1210/edrv-1-1-1. [DOI] [PubMed] [Google Scholar]
- 16.Smith AI, Funder JW. Proopiomelanocortin Processing in the Pituitary, Central Nervous System and Peripheral Tissues. Endocrin Rev. 1988;9:159–179. doi: 10.1210/edrv-9-1-159. [DOI] [PubMed] [Google Scholar]
- 17.Bultman SJ, Michaud EJ, Woychick RP. Molecular Characterization of the Mouse Agouti Locus. Cell. 1992;71:1195–1204. doi: 10.1016/s0092-8674(05)80067-4. [DOI] [PubMed] [Google Scholar]
- 18.Lu D, Willard D, Patel IR, Kadwell S, Overton L, Kost T, Luther M, Chen W, Yowchik RP, Wilkison WO, Cone RD. Agouti Protein is an Antagonist of the Melanocyte-Stimulating-Hormone Receptor. Nature. 1994;371(6500):799–802. doi: 10.1038/371799a0. [DOI] [PubMed] [Google Scholar]
- 19.Ollmann MM, Wilson BD, Yang YK, Kerns JA, Chen Y, Gantz I, Barsh GS. Antagonism of Central Melanocortin Receptors in Vitro and in Vivo by Agouti-Related Protein. Science. 1997;278:135–138. doi: 10.1126/science.278.5335.135. [DOI] [PubMed] [Google Scholar]
- 20.Haskell-Luevano C, Monck EK. Agouti-related Protein (AGRP) Functions as an Inverse Agonist at a Constitutively Active Brain Melanocortin-4 Receptor. Regulatory Peptides. 2001;99:1–7. doi: 10.1016/s0167-0115(01)00234-8. [DOI] [PubMed] [Google Scholar]
- 21.Hruby VJ, Wilkes BC, Cody WL, Sawyer TK, Hadley ME. Melanotropins: Structural, Conformational and Biological Considerations in the Development of Superpotent and Superprolonged Analogs. Peptide Protein Rev. 1984;3:1–64. [Google Scholar]
- 22.Holder JR, Haskell-Luevano C. Melanocortin Ligands: 30 Years of Structure-Activity Relationship (SAR) Studies. Med Res Rev. 2004;24(3):325–356. doi: 10.1002/med.10064. [DOI] [PubMed] [Google Scholar]
- 23.Al-Obeidi F, Hadley ME, Pettitt BM, Hruby VJ. Design of a New Class of Superpotent Cyclic α-Melanotropins Based on Quenched Dynamic Stimulations. J Am Chem Soc. 1989;111(9):3413–3416. [Google Scholar]
- 24.Sawyer TK, Hruby VJ, Darman PS, Hadley ME. [half-Cys4half-Cys10]-α-Melanocyte-Stimulating Hormone: A Cyclic a-Melanotropin Exhibiting Superagonist Biological Activity. Proc Natl Acad Sci USA. 1982;79:1751–1755. doi: 10.1073/pnas.79.6.1751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hess S, Linde Y, Ovadia O, Safrai E, Shalev DE, Swed A, Halbfinger E, Lapidot T, Winkler I, Gabinet Y, Faier A, Yarden D, Xiang Z, Portillo FP, Haskell-Luevano C, Gilon C, Hoffman A. Backbone Cyclic Peptidomimetic Melanocortin-4 Receptor Agonist as a Novel Orally Administrated Drug Lead for Treating Obesity. J Med Chem. 2008;51(4):1026–1034. doi: 10.1021/jm701093y. [DOI] [PubMed] [Google Scholar]
- 26.Hruby VJ, Lu D, Sharma SD, Castrucci AML, Kesterson RA, Al-Obeidi FA, Hadley ME, Cone RD. Cyclic Lactam α-Melanotropin Analogues of Ac-Nle4-c[Asp5, DPhe7, Lys10]-α-MSH(4–10)-NH2 With Bulky Aromatic Amino Acids at Position 7 Show High Antagonist Potency and Selectivity at Specific Melanocortin Receptors. J Med Chem. 1995;38:3454–3461. doi: 10.1021/jm00018a005. [DOI] [PubMed] [Google Scholar]
- 27.Haskell-Luevano C, Toth K, Boteju L, Job C, Castrucci AML, Hadley ME, Hruby VJ. β-Methylation of Phe7 and Trp9 Melanotropin Side Chain Pharmacophores Affect Ligand-Receptor Interactions and Prolonged Biological Activity. J Med Chem. 1997;40:2740–2749. doi: 10.1021/jm970018t. [DOI] [PubMed] [Google Scholar]
- 28.Holder JR, Bauzo RM, Xiang Z, Scott JW, Haskell-Luevano C. Design and Pharmacology of Peptoids and Peptide-Peptoid Hybrids Based on the Melancortin Agonist Core Tetrapeptide Sequence. Bioorg Med Chem Lett. 2003;13:4505–4509. doi: 10.1016/j.bmcl.2003.08.078. [DOI] [PubMed] [Google Scholar]
- 29.Verhelst SH, Witte MD, Arastu-Kapur S, Fonovic M, Bogyo M. Novel Aza Peptide Inhibitors and Active-Site Probes of Papain-Family Cysteine Proteases. Chembiochem. 2006;7(6):943–950. doi: 10.1002/cbic.200600001. [DOI] [PubMed] [Google Scholar]
- 30.Sawyer TK, Sanfillippo PJ, Hruby VJ, Engel MH, Heward CB, Burnett JB, Hadley ME. 4-Norleucine, 7-D-Phenylalanine-α-Melanocyte-Stimulating Hormone: A Highly Potent α-Melanotropin with Ultra Long Biological Activity. Proc Natl Acad Sci USA. 1980;77:5754–5758. doi: 10.1073/pnas.77.10.5754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Haskell-Luevano C, Boteju LW, Miwa H, Dickinson C, Gantz I, Yamada T, Hadley ME, Hruby VJ. Topographical Modifications of Melanotropin Peptide Analogues with β-Methyltryptophan Isomers at Position 9 Leads to Differential Potencies and Prolonged Biological Activities. J Med Chem. 1995;38:4720–4729. doi: 10.1021/jm00023a012. [DOI] [PubMed] [Google Scholar]
- 32.Hruby VJ, Li G, Haskell-Luevano C, Shenderovich M. Design of Peptides, Proteins, and Peptidomimetics in Chi Space. Biopolymers, Peptide Science. 1997;43:219–266. doi: 10.1002/(SICI)1097-0282(1997)43:3<219::AID-BIP3>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- 33.Todorovic A, Holder JR, Scott JW, Haskell-Luevano C. Synthesis and Activity of the Melanocortin Xaa-DPhe-Arg-Trp-NH2 Tetrapeptides with Amide Bond Modifications. J Pept Res. 2004;63(3):270–278. doi: 10.1111/j.1399-3011.2004.00137.x. [DOI] [PubMed] [Google Scholar]
- 34.Doedens L, Opperer F, Cai M, Beck JG, Dedek M, Palmer E, Hruby VJ, Kessler H. Multiple N-Methylation of MT-II Backbone Amide Bonds Leads to Melanocortin Receptor Subtype hMC1R Selectivity: Pharmacological and Conformational Studies. J Am Chem Soc. 2010;132(23):8115–8128. doi: 10.1021/ja101428m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Bondebjerg J, Xiang Z, Bauzo RM, Haskell-Luevano C, Meldal M. A Solid Phase Approach to Mouse Melanocortin Receptor Agonists Derived From a Novel Thioether Cyclized Peptidomimetic Scaffold. J Am Chem Soc. 2002;124:11046–11055. doi: 10.1021/ja0123913. [DOI] [PubMed] [Google Scholar]
- 36.Wilczynski A, Wang XS, Joseph CG, Xiang Z, Bauzo RM, Scott JW, Sorensen NB, Shaw AM, Millard WJ, Richards NG, Haskell-Luevano C. Identification of Putative Agouti-Related Protein(87–132)-Melanocortin-4 Receptor Interactions by Homology Molecular Modeling and Validation Using Chimeric Peptide Ligands. J Med Chem. 2004;47(9):2194–2207. doi: 10.1021/jm0303608. [DOI] [PubMed] [Google Scholar]
- 37.Wilczynski A, Wilson KR, Scott JW, Edison AS, Haskell-Luevano C. Structure-Activity Relationships of the Unique and Potent Agouti-Related Protein (AGRP)-Melanocortin Chimeric Tyr-c[β-Asp-His-DPhe-Arg-Trp-Asn-Ala-Phe-Dpr]-Tyr-NH2 Peptide Template. J Med Chem. 2005;48(8):3060–3075. doi: 10.1021/jm049010r. [DOI] [PubMed] [Google Scholar]
- 38.Xiang Z, Pogozheva ID, Sorenson NB, Wilczynski AM, Holder JR, Litherland SA, Millard WJ, Mosberg HI, Haskell-Luevano C. Peptide and Small Molecules Rescue the Functional Activity and Agonist Potency of Dysfunctional Human Melanocortin-4 Receptor polymorphisms. Biochemistry. 2007;46(28):8273–8287. doi: 10.1021/bi7007382. [DOI] [PubMed] [Google Scholar]
- 39.Singh A, Wilczynski A, Holder JR, Witek RM, Dirain ML, Xiang Z, Edison AS, Haskell-Luevano C. Incorporation of a Bioactive Reverse-Turn Heterocycle into a Peptide Template Using Solid-Phase Synthesis To Probe Melanocortin Receptor Selectivity and Ligand Conformations by 2D (1)H NMR. J Med Chem. 2011;54(5):1379–1390. doi: 10.1021/jm101425m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chen W, Shields TS, Stork PJS, Cone RD. A Colorimetric Assay for Measuring Activation of Gs- and Gq-Coupled Signaling Pathways. Anal Biochem. 1995;226:349–354. doi: 10.1006/abio.1995.1235. [DOI] [PubMed] [Google Scholar]
- 41.Al-Obeidi F, Castrucci AM, Hadley ME, Hruby VJ. Potent and Prolonged Acting Cyclic Lactam Analogues of α-Melanotropin: Design Based on Molecular Dynamics. J Med Chem. 1989;32:2555–2561. doi: 10.1021/jm00132a010. [DOI] [PubMed] [Google Scholar]
- 42.Al-Obeidi F, Hruby VJ, Castrucci AM, Hadley ME. Design of Potent Linear α-Melanotropin 4–10 Analogues Modified in Positions 5 and 10. J Med Chem. 1989;32(1):174–9. doi: 10.1021/jm00121a032. [DOI] [PubMed] [Google Scholar]
- 43.Schild HO. pA, A New Scale for the Measurement of Drug Antagonism. Brit J Pharmacol. 1947;2:189–206. doi: 10.1111/j.1476-5381.1947.tb00336.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Danho W, Swistok J, Cheung A, Chu Y-J, Wang Y, Chen L, Bartkovitz D, Gore V, Qi L, Fry D, Greeley D, Sun H, Guenot J, Franco L, Kurylko G, Rumennik L, Yagaloff K. Highly Selective Cyclic Peptides for Human Melanocortin-4 Receptor (MC-4 R): Design, Synthesis, Bioactive Conformation, and Pharmacological Evaluation as an Anti-obesity Agent. In: Lebel MHR, editor. Proceedings of the 2nd International/17th American Peptide Symposium. Kluwer Academic Publishers; The Netherlands: 2001. pp. 701–703. [Google Scholar]
- 45.Chen L, Cheung AW-H, Chu X-J, Danho W, Swistok J, Yagaloff KA. WO 01/74844 A2. Selective Linear Peptides with Melanocortin-4 Receptor (MC4-R) Agonist Activity. 2001 Oct 11;2001
- 46.Cheung A, Danho W, Swistok J, Qi L, Kurylko G, Franco L, Yagaloff K, Chen L. Structure-Activity Relationship of Linear Peptide Bu-His-DPhe-Arg-Trp-Gly-NH(2) at the Human Melanocortin-1 and -4 Receptors: Arginine Substitution. Bioorg Med Chem Lett. 2002;12(17):2407–2410. doi: 10.1016/s0960-894x(02)00459-6. [DOI] [PubMed] [Google Scholar]
- 47.Cheung AW, Danho W, Swistok J, Qi L, Kurylko G, Rowan K, Yeon M, Franco L, Chu XJ, Chen L, Yagaloff K. Structure-Activity Relationship of Linear Peptide Bu-His-DPhe-Arg-Trp-Gly-NH(2) at the Human Melanocortin-1 and -4 Receptors: Histidine Substitution. Bioorg Med Chem Lett. 2003;13(1):133–137. doi: 10.1016/s0960-894x(02)00830-2. [DOI] [PubMed] [Google Scholar]
- 48.Danho W, Swistok J, Wai-Hing Cheung A, Kurylko G, Franco L, Chu XJ, Chen L, Yagaloff K. Structure-Activity Relationship of Linear Peptide Bu-His(6)-DPhe(7)-Arg(8)-Trp(9)-Gly(10)-NH(2) at the Human Melanocortin-1 and -4 Receptors: DPhe(7) and Trp(9) Substitution. Bioorg Med Chem Lett. 2003;13(4):649–52. doi: 10.1016/s0960-894x(03)00114-8. [DOI] [PubMed] [Google Scholar]
- 49.Cheung AW, Danho W, Swistok J, Qi L, Kurylko G, Rowan K, Yeon M, Franco L, Chu XJ, Chen L, Yagaloff K. Structure-Activity Relationship of Cyclic Peptide Penta-c[Asp-His(6)-DPhe(7)-Arg(8)-Trp(9)-Lys]-NH(2) at the Human Melanocortin-1 and -4 Receptors: His(6) Substitution. Bioorg Med Chem Lett. 2003;13(7):1307–11. doi: 10.1016/s0960-894x(03)00114-8. [DOI] [PubMed] [Google Scholar]
- 50.Holder JR, Bauzo RM, Xiang Z, Haskell-Luevano C. Structure-Activity Relationships of the Melanocortin Tetrapeptide Ac-His-DPhe-Arg-Trp-NH2 at the Mouse Melanocortin Receptors: I Modifications at the His Position. J Med Chem. 2002;45:2801–2810. doi: 10.1021/jm0104872. [DOI] [PubMed] [Google Scholar]
- 51.Hruby VJ, Wilkes BC, Hadley ME, Al-Obeidi F, Sawyer TK, Staples DJ, DeVaux A, Dym O, Castrucci AM, Hintz MF, Riehm JP, Rao KR. α-Melanotropin: The Minimal Active Sequence in the Frog Skin Bioassay. J Med Chem. 1987;30:2126–2130. doi: 10.1021/jm00394a033. [DOI] [PubMed] [Google Scholar]
- 52.Holder JR, Bauzo RM, Xiang Z, Haskell-Luevano C. Structure-Activity Relationships of the Melanocortin Tetrapeptide Ac-His-DPhe-Arg-Trp-NH2 at the Mouse Melanocortin Receptors: Part 2 Modifications at the Phe Position. J Med Chem. 2002;45:3073–3081. doi: 10.1021/jm010524p. [DOI] [PubMed] [Google Scholar]
- 53.Proneth B, Pogozheva ID, Portillo FP, Mosberg HI, Haskell-Luevano C. Melanocortin Tetrapeptide Ac-His-DPhe-Arg-Trp-NH2 Modified at the Para Position of the Benzyl Side Chain (DPhe): Importance for Mouse Melanocortin-3 Receptor Agonist Versus Antagonist Activity. J Med Chem. 2008;51(18):5585–5593. doi: 10.1021/jm800291b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Wishart DS, Sykes BD. Chemical Shifts as a Tool for Structure Determination. Methods Enzymol. 1994;239:363–92. doi: 10.1016/s0076-6879(94)39014-2. [DOI] [PubMed] [Google Scholar]
- 55.Prabhu NV, Perkyns JS, Pettitt BM. Modeling of α-MSH Conformations with Implicit Solvent. J Peptide Res. 1999;(54):394–407. doi: 10.1034/j.1399-3011.1999.00113.x. [DOI] [PubMed] [Google Scholar]
- 56.Haskell-Luevano C, Sawyer TK, Trumpp-Kallmeyer S, Bikker J, Humblet C, Gantz I, Hruby VJ. Three-Dimensional Molecular Models of the hMC1R Melanocortin Receptor: Complexes with Melanotropin Peptide Agonists. Drug Design & Discovery. 1996;14:197–211. [PubMed] [Google Scholar]
- 57.Rose GD, Gierash LM, Smith JA. Turns in Peptides and Proteins. Adv in Protein Chem. 1985;37:1–109. doi: 10.1016/s0065-3233(08)60063-7. [DOI] [PubMed] [Google Scholar]
- 58.Rowland NE, Schaub JW, Robertson KL, Andreasen A, Haskell-Luevano C. Effect of MTII on Food Intake and Brain c-Fos in Melanocortin-3, Melanocortin-4, and Double MC3 and MC4 Receptor Knockout Mice. Peptides. 2010;31(12):2314–2317. doi: 10.1016/j.peptides.2010.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Butler AA, Marks DL, Fan W, Kuhn CM, Bartolome M, Cone RD. Melanocortin-4 Receptor is Required for Acute Homeostatic Responses to Increased Dietary Fat. Nat Neurosci. 2001;4(6):605–611. doi: 10.1038/88423. [DOI] [PubMed] [Google Scholar]
- 60.Chen AS, Marsh DJ, Trumbauer ME, Frazier EG, Guan XM, Yu H, Rosenblum CI, Vongs A, Feng Y, Cao L, Metzger JM, Strack AM, Camacho RE, Mellin TN, Nunes CN, Min W, Fisher J, Gopal-Truter S, MacIntyre DE, Chen HY, Van Der Ploeg LH. Inactivation of the Mouse Melanocortin-3 Receptor Results in Increased Fat Mass and Reduced Lean Body Mass. Nat Genet. 2000;26(1):97–102. doi: 10.1038/79254. [DOI] [PubMed] [Google Scholar]
- 61.Henagan TM, Phillips MD, Cheek DJ, Kirk KM, Barbee JJ, Stewart LK. The Melanocortin 3 Receptor: A Novel Mediator of Exercise-Induced Inflammation Reduction in Postmenopausal Women? J Aging Res. 2011;2011:512593, 1–7. doi: 10.4061/2011/512593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Mandrika I, Petrovska R, Wikberg J. Melanocortin Receptors Form Constitutive Homo- and Heterodimers. Biochem Biophys Res Commun. 2005;326(2):349–354. doi: 10.1016/j.bbrc.2004.11.036. [DOI] [PubMed] [Google Scholar]
- 63.Rediger A, Piechowski CL, Yi CX, Tarnow P, Strotmann R, Gruters A, Krude H, Schoneberg T, Tschop MH, Kleinau G, Biebermann H. Mutually Opposite Signal Modulation by Hypothalamic Heterodimerization of Ghrelin and Melanocortin-3 receptors. J Biol Chem. 2011;286(45):39623–39631. doi: 10.1074/jbc.M111.287607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Rediger A, Piechowski CL, Habegger K, Gruters A, Krude H, Tschop MH, Kleinau G, Biebermann H. MC4R dimerization in the Paraventricular Nucleus and GHSR/MC3R Heterodimerization in the Arcuate Nucleus: is there Relevance for Body Weight Regulation? Neuroendocrinology. 2012;95(4):277–288. doi: 10.1159/000334903. [DOI] [PubMed] [Google Scholar]
- 65.Sun H, Greeley DN, Chu XJ, Cheung A, Danho W, Swistok J, Wang Y, Zhao C, Chen L, Fry DC. A Predictive Pharmacophore Model of Human Melanocortin-4 Receptor as Derived from the Solution Structures of Cyclic Peptides. Bioorg Med Chem. 2004;12(10):2671–2677. doi: 10.1016/j.bmc.2004.03.017. [DOI] [PubMed] [Google Scholar]
- 66.Schioth HB, Muceniece R, Mutulis F, Bouifrouri AA, Mutule I, Wikberg JE. Further Pharmacological Characterization of the Selective Melanocortin 4 Receptor Antagonist HS014: Comparison with SHU9119. Neuropeptides. 1999;33(3):191–196. doi: 10.1054/npep.1999.0760. [DOI] [PubMed] [Google Scholar]
- 67.Grieco P, Balse-Srinivasan P, Han G, Weinberg D, MacNeil T, Van Der Ploeg LH, Hruby VJ. Extensive Structure-Activity Studies of Lactam Derivatives of MT-II and SHU9119: Their Activity and Selectivity at Human Melanocortin Receptors 3, 4, and 5. J Pept Res. 2003;62(5):199–206. doi: 10.1034/j.1399-3011.2003.00087.x. [DOI] [PubMed] [Google Scholar]
- 68.Grieco P, Han G, Weinberg D, Van der Ploeg LH, Hruby VJ. Design and Synthesis of Highly Potent and Selective Melanotropin Analogues of SHU9119 Modified at Position 6. Biochem Biophys Res Commun. 2002;292(4):1075–1080. doi: 10.1006/bbrc.2002.6739. [DOI] [PubMed] [Google Scholar]
- 69.Bednarek MA, MacNeil T, Kalyani RN, Tang R, Van Der Ploeg LH, Weinberg DH. Selective, High Affinity Peptide Antagonists of α-Melanotropin Action at Human Melanocortin Receptor 4: Their Synthesis and Biological Evaluation in Vitro. J Med Chem. 2001;44:3665–3672. doi: 10.1021/jm010165y. [DOI] [PubMed] [Google Scholar]
- 70.Ying J, Gu X, Cai M, Dedek M, Vagner J, Trivedi DB, Hruby VJ. Design, Synthesis, and Biological Evaluation of New Cyclic Melanotropin Peptide Analogues Selective for the Human Melanocortin-4 Receptor. J Med Chem. 2006;49(23):6888–6896. doi: 10.1021/jm060768f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Holder JR, Xiang Z, Bauzo RM, Haskell-Luevano C. Structure-Activity Relationships of the Melanocortin Tetrapeptide Ac-His-DPhe-Arg-Trp-NH2 at the Mouse Melanocortin Receptors: Part 3 Modifications at the Arg Position. Peptides. 2003;24:73–82. doi: 10.1016/s0196-9781(02)00278-4. [DOI] [PubMed] [Google Scholar]
- 72.Holder JR, Xiang Z, Bauzo RM, Haskell-Luevano C. Structure-Activity Relationships of the Melanocortin Tetrapeptide Ac-His-DPhe-Arg-Trp-NH2 at the Mouse Melanocortin Receptors: Part 4 Modifications at the Trp Position. J Med Chem. 2002;45:5736–5744. doi: 10.1021/jm020296e. [DOI] [PubMed] [Google Scholar]
- 73.Mogil JS, Ritchie J, Smith SB, Strasburg K, Kaplan L, Wallace MR, Romberg RR, Bijl H, Sarton EY, Fillingim RB, Dahan A. Melanocortin-1 Receptor Gene Variants Affect Pain and Mu-Opioid Analgesia in Mice and Humans. J Med Genet. 2005;42(7):583–587. doi: 10.1136/jmg.2004.027698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Maaser C, Kannengiesser K, Kucharzik T. Role of the Melanocortin System in Inflammation. Ann N Y Acad Sci. 2006;1072:123–134. doi: 10.1196/annals.1326.016. [DOI] [PubMed] [Google Scholar]
- 75.Catania A. The Melanocortin System in Leukocyte Biology. J Leukoc Biol. 2007;81(2):383–392. doi: 10.1189/jlb.0706426. [DOI] [PubMed] [Google Scholar]
- 76.Sahm UG, Olivier GWJ, Branch SK, Moss SH, Pouton CW. Synthesis and Biological Evaluation of α-MSH Analogs Substituted with Alanine. Peptides. 1994;15(7):1297–1302. doi: 10.1016/0196-9781(94)90157-0. [DOI] [PubMed] [Google Scholar]
- 77.Kask A, Mutulis F, Muceniece R, Pahkla R, Mutule I, Wikberg JE, Rago L, Schioth HB. Discovery of a Novel Superpotent and Selective Melanocortin-4 Receptor Antagonist (HS024): Evaluation in vitro and in vivo. Endocrinology. 1998;139(12):5006–5014. doi: 10.1210/endo.139.12.6352. [DOI] [PubMed] [Google Scholar]
- 78.Kavarana MJ, Trivedi D, Cai M, Ying J, Hammer M, Cabello C, Grieco P, Han G, Hruby VJ. Novel Cyclic Templates of α-MSH Give Highly Selective and Potent Antagonists/Agonists for Human Melanocortin-3/4 Receptors. J Med Chem. 2002;45(12):2644–2650. doi: 10.1021/jm020021z. [DOI] [PubMed] [Google Scholar]
- 79.Bednarek MA, Silva MV, Arison B, MacNeil T, Kalyani RN, Huang RR, Weinberg DH. Structure-Function Studies on the Cyclic Peptide MT-II, Lactam Derivative of α-melanotropin. Peptides. 1999;20(3):401–409. doi: 10.1016/s0196-9781(99)00048-0. [DOI] [PubMed] [Google Scholar]
- 80.Haskell-Luevano C, Cone RD, Monck EK, Wan Y-P. Structure Activity Studies of the Melanocortin-4 Receptor by In Vitro Mutagenesis: Identification of Agouti-Related Protein (AGRP), Melanocortin Agonist and Synthetic Peptide Antagonist Interaction Determinants. Biochemistry. 2001;40(20):6164–6179. doi: 10.1021/bi010025q. [DOI] [PubMed] [Google Scholar]
- 81.Yang Y, Fong TM, Dickinson CJ, Mao C, Li JY, Tota MR, Mosley R, Van Der Ploeg LH, Gantz I. Molecular Determinants of Ligand Binding to the Human Melanocortin-4 Receptor. Biochemistry. 2000;39(48):14900–14911. doi: 10.1021/bi001684q. [DOI] [PubMed] [Google Scholar]
- 82.Chen M, Aprahamian CJ, Celik A, Georgeson KE, Garvey WT, Harmon CM, Yang Y. Molecular Characterization of Human Melanocortin-3 Receptor Ligand-Receptor Interaction. Biochemistry. 2006;45(4):1128–1137. doi: 10.1021/bi0521792. [DOI] [PubMed] [Google Scholar]
- 83.Bednarek MA, Macneil T, Kalyani RN, Tang R, Van der Ploeg LH, Weinberg DH. Analogs of MTII, Lactam Derivatives of α-Melanotropin, Modified at the N-terminus, and their Selectivity at Human Melanocortin Receptors 3, 4, and 5. Biochem Biophys Res Commun. 1999;261(1):209–213. doi: 10.1006/bbrc.1999.0981. [DOI] [PubMed] [Google Scholar]
- 84.Bednarek MA, MacNeil T, Tang R, Kalyani RN, Van der Ploeg LH, Weinberg DH. Potent and Selective Peptide Agonists of alpha-Melanotropin Action at Human Melanocortin Receptor 4: Their Synthesis and Biological Evaluation in Vitro. Biochem Biophys Res Commun. 2001;286(3):641–645. doi: 10.1006/bbrc.2001.5444. [DOI] [PubMed] [Google Scholar]
- 85.Chhajlani V, Muceniece R, Wikberg JES. Molecular Cloning of a Novel Human Melanocortin Receptor. Biochem Biophys Res Commun. 1993;195(2):866–873. doi: 10.1006/bbrc.1993.2125. [DOI] [PubMed] [Google Scholar]
- 86.Chen W, Kelly MA, Opitz-Araya X, Thomas RE, Low MJ, Cone RD. Exocrine Gland Dysfunction In MC5-R Deficient Mice: Evidence For Coordinated Regulation Of Exocrine Gland Functions By Melanocortin Peptides. Cell. 1997;91:789–798. doi: 10.1016/s0092-8674(00)80467-5. [DOI] [PubMed] [Google Scholar]
- 87.Victoria Silva Elipe M, Mosley RT, Bednarek MA, Arison BH. 1H-NMR Studies on a Potent and Selective Antagonist at Human Melanocortin Receptor 4 (hMC-4R) Biopolymers. 2003;68(4):512–527. doi: 10.1002/bip.10340. [DOI] [PubMed] [Google Scholar]
- 88.Ying J, Kover KE, Gu X, Han G, Trivedi DB, Kavarana MJ, Hruby VJ. Solution Structures of Cyclic Melanocortin Agonists and Antagonists by NMR. Biopolymers. 2003;71(6):696–716. doi: 10.1002/bip.10596. [DOI] [PubMed] [Google Scholar]
- 89.Li SZ, Lee JH, Lee W, Yoon CJ, Baik JH, Lim SK. Type I Beta-Turn Conformation is Important for Biological Activity of the Melanocyte-Stimulating Hormone Analogues. Eur J Biochem. 1999;265(1):430–440. doi: 10.1046/j.1432-1327.1999.00763.x. [DOI] [PubMed] [Google Scholar]
- 90.Al-Obeidi FA, O’Connor S, Hruby VJ, Pettitt M. NMR and Quenched Molecular Dynamics Studies of Superpotent Linear and Cyclic α-Melanotropins. J Peptide Res. 1998;51:420–431. doi: 10.1111/j.1399-3011.1998.tb00640.x. [DOI] [PubMed] [Google Scholar]
- 91.Fotsch C, Smith DM, Adams JA, Cheetham J, Croghan M, Doherty EM, Hale C, Jarosinski MA, Kelly MG, Norman MH, Tamayo NA, Xi N, Baumgartner JW. Design of a New Peptidomimetic Agonist for the Melanocortin Receptors Based on the Solution Structure of the Peptide Ligand, Ac-Nle-cyclo[Asp-Pro-DPhe-Arg-Trp-Lys]-NH(2) Bioorg Med Chem Lett. 2003;13(14):2337–2340. doi: 10.1016/s0960-894x(03)00412-8. [DOI] [PubMed] [Google Scholar]
- 92.Carpino LA, Han GY. The 9-Fluorenylmethoxycarbonyl Function, a New Base-Sensitive Amino-Protecting Group. J Am Chem Soc. 1970;92(19):5748–5749. [Google Scholar]
- 93.Carpino LA, Han GY. The 9-Fluorenylmethyloxycarbonyl Amino-Protecting Group. J Org Chem. 1972;37:3404–3409. [Google Scholar]
- 94.Kaiser E, Colescott RL, Bossinger CD, Cook PI. Color Test for Detection of Free Terminal Amino Groups in the Solid-Phase Synthesis of Peptides. Anal Biochem. 1970;34:595–598. doi: 10.1016/0003-2697(70)90146-6. [DOI] [PubMed] [Google Scholar]
- 95.Thirumoorthy R, Holder JR, Bauzo RM, Richards NGJ, Edison AS, Haskell-Luevano C. Novel Agouti-related Protein (AGRP) Based Melanocortin-1 Receptor Antagonist. J Med Chem. 2001;44:4114–4124. doi: 10.1021/jm010215z. [DOI] [PubMed] [Google Scholar]
- 96.Wüthrich K. NMR of Proteins and Nucleic Acids. John Wiley & Sons; New York: 1986. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.