

Abstract
Several brain regions show structural and functional abnormalities in individuals with autism spectrum disorders (ASD), but the developmental trajectory of abnormalities in these structures and how they may relate to social and communicative impairments are still unclear. We assessed the effects of age on cortical thickness in individuals with ASD, between the ages of 7 and 39 years in comparison to typically developing controls. Additionally, we examined differences in cortical thickness in relation to symptomatology in the ASD group, and their association with age. Analyses were conducted using a general linear model, controlling for sex. Social and communication scores from the Autism Diagnostic Interview-Revised (ADI-R) were correlated with the thickness of regions implicated in those functions. Controls showed widespread cortical thinning relative to the ASD group. Within regions-of-interest, increased thickness in the rostral anterior cingulate cortex was associated with poorer social scores. Additionally, a significant interaction between age and social impairment was found in the orbitofrontal cortex, with more impaired younger children having decreased thickness in this region. These results suggest that differential neurodevelopmental trajectories are present in individuals with ASD and some differences are associated with diagnostic behaviours.
Keywords: Autism spectrum disorders, Structural MRI, Cortical thickness, Social impairment, Developmental changes
1. Introduction
Age-related structural abnormalities in the brains of individuals with autism spectrum disorders (ASD) have been explored in a number of longitudinal and cross-sectional studies. Across these studies, grey matter volumes within frontal and temporal lobes were significantly increased in children and adolescents with ASD compared to typically developing individuals (Bloss & Courchesne, 2007; Carper, Moses, Tigue, & Courchesne, 2002; Courchesne, Campbell, & Solso, 2011; Hazlett, Poe, Gerig, Smith, & Piven, 2006; Kates et al., 2004; Palmen et al., 2005; Schumann et al., 2010). However, volumetric measurements of lobular regions do not provide spatially localized information on grey matter abnormalities. Advances in brain morphometric analyses now permit the direct measure of cortical thickness, a determinant of grey matter volume, and can provide detailed information concerning the spatial extent and location of cortical differences between groups.
Large-scale developmental studies in typically developing individuals that include both cross-sectional and longitudinal data show that after a period of growth in early childhood, grey matter thinning occurs across the entire cortex in adolescence (Raznahan, Shaw, et al., 2011; Raznahan, Lerch, et al., 2011; Shaw et al., 2008). To our knowledge, no longitudinal studies in individuals with ASD have examined cortical thickness development across a wide age-range. Cross-sectional studies in ASD suggest atypical cortical thickness changes between childhood and adulthood in individuals with ASD (Mak-Fan, Taylor, Roberts, & Lerch, 2011; Raznahan et al., 2010; Scheel et al., 2011; Wallace, Dankner, Kenworthy, Giedd, & Martin, 2010). In children with ASD aged 6–15 years, Mak-Fan et al. (2011) reported decreased cortical thickness with age across the entire brain, as well as in the frontal and parietal lobes; however group differences were not found. In older samples, studies have observed thinner cortices with increasing age in participants with ASD relative to typically developing controls in the left posterior part of the superior temporal sulcus (Scheel et al., 2011; Wallace et al., 2010), left cuneus (Scheel et al., 2011) and right paracentral lobule (PCL) (Scheel et al., 2011). In contrast, brain regions such as the left supramarginal gyrus (Scheel et al., 2011), left inferior parietal lobule (Scheel et al., 2011), left middle temporal gyrus (Raznahan et al., 2010), right postcentral gyrus (Scheel et al., 2011) and right fusiform (Raznahan et al., 2010), show decreased cortical thickness across age in typically developing individuals, but little to no change in individuals with ASD. These findings suggest a maturational process that is both atypical and complex in ASD.
Maturational brain changes in regions of the brain implicated in sensorimotor and cognitive functions are believed to be associated with improvement in these functions (Greene, Braet, Johnson, & Bellgrove, 2008). Thus, deviation from the typical developmental trajectory of cortical thickness in frontal and temporal brain regions may underlie impairments in social and communication functions in ASD. Few studies have examined the relation between cortical thickness abnormalities and ASD symptom severity (Hadjikhani, Joseph, Snyder, & Tager-Flusberg, 2006; Hardan, Libove, Keshavan, Melhem, & Minshew, 2009), although these studies indicated that cortical thinning over a 2-year period in the frontal lobes (Hardan et al., 2009), and thinner cortices in frontal, temporal and parietal regions in adults with ASD (Hadjikhani et al., 2006), were associated with more severe social impairments. The age-range in both studies was narrow and did not examine morphological changes related to ASD symptomatology in children and adults. As ASD are a group of neurodevelopmental disorders, age-related changes in brain morphology are crucial to understanding the neurobiology and how it relates to changes in behaviour over time.
In the current cross-sectional study, we assessed cortical thickness in a cohort of 53 individuals with and without ASD spanning childhood, adolescence and adulthood. In addition, we assessed the relation between cortical thickness in specific regions implicated in social and communication functions and symptom severity. In this latter analysis, a priori regions of interest (ROIs) included anterior cingulate cortices (ACC), medial prefrontal cortices (mPFC), orbital frontal cortices (OFC), inferior frontal gyri (IFG), the superior temporal gyri (STG) and fusiform gyri because of their involvement in social behaviours and communication (Baron-Cohen & Belmonte, 2005).
2. Materials and methods
2.1. Ethics and consent
This study was approved by the Mount Sinai School of Medicine Institutional Research Board (IRB) and conducted in accordance with its guidelines. Written informed consent was obtained from all participants 18 years of age and older. Younger participants provided written informed assent and parents provided written informed consent in accordance with IRB guidelines.
2.2. Participants
Seventy-one participants between the ages of 7 and 39 years who were enrolled in magnetic resonance imaging (MRI) studies at Mount Sinai School of Medicine were evaluated for inclusion [children: 7–21 years, n = 41; ASD = 25, typically developing children = 16; adults: 22 years and older, n = 30; ASD = 15, typically developing adults = 15]. Participants with ASD were clinically diagnosed according to the Diagnostic and Statistical Manual of Mental Disorders, 4th Edition (DSM-IV) and were recruited from the Seaver Autism Center at Mount Sinai School of Medicine. Their diagnoses were then confirmed using the Autism Diagnostic Observational Schedule (ADOS) (Lord et al., 2000) and the Autism Diagnostic Interview-Revised (ADI-R) (Lord, Rutter, & Le Couteur, 1994). Typically developing individuals were recruited from the community using newspaper advertisements. All participants had a full scale IQ of greater than 70 as estimated by either the Wechsler Intelligence Scale for Children-fourth edition (WISC-IV; Wechsler, 2003) or the Wechsler Adult Intelligence Scale-fourth edition (WAIS-IV; Wechsler, 2008).
Participants were excluded if they had a primary psychiatric diagnosis (in addition to ASD in the ASD group) or a medical history of neurological disease, including but not limited to epilepsy/seizure disorder (except simple febrile seizures), movement disorder, tuberous sclerosis, fragile X, and any other known genetic syndromes, or known abnormal MRI/structural lesion of the brain. Typically developing individuals were also excluded if they had a first-degree relative with ASD. All participants indicated that they were right-handed.
2.3. Imaging
Participants were scanned on a Siemens Allegra 3 T MRI system, which has a maximum gradient strength of 40 mT/m, and a slew-rate of 400 mT/m s. A low-resolution localizer image was first obtained, followed by a series of axial scans. High-resolution structural images with good grey/white matter contrast were obtained using a T1-weighted magnetization-prepared rapid gradient-echo (MP-RAGE) sequence (isotropic resolution of 0.82 mm × 0.82 mm × 0.82 mm, matrix size = 256 × 256 × 208, FOV = 210 mm, 208 slices, TR = 2500 ms, TE = 4.38 ms, TI = 1100 ms and a 8° flipangle, fast low angle shot [FLASH] acquisition). The total imaging time was approximately 10 min.
2.4. Cortical thickness measurements
Structural MRIs were preprocessed using a standard protocol (linear registration into standardized space, RF inhomogeniety correction) within the CIVET processing pipeline as described in Ad-Dab’bagh et al. (Ad-Dab’bagh et al., 2006). This pipeline is highly efficient and has been used widely to study cortical thickness in healthy individuals, special populations and in disease states (Bermudez, Lerch, Evans, & Zatorre, 2009; Bernhardt et al., 2008; Charil et al., 2007; Fahim, Yoon, Sandor, Frey, & Evans, 2009; Giedd et al., 2007; Lenroot et al., 2009; Lerch et al., 2005, 2006, 2008; Meguid et al., 2010; Singh et al., 2006; Zhou et al., 2011). The preprocessing involves linear registration of MRIs into a common stereotactic space followed by correcting for non-uniformity artefacts (Collins, Peters, & Evans, 1994; Sled, Zijdenbos, & Evans, 1998). The processed MRIs were then segmented according to their physiological classification (grey matter, white matter, cerebrospinal fluid) (Zijdenbos, Forghani, & Evans, 2002). The Constrained Laplacian Anatomical Segmentation using Proximities (CLASP) method (Kim et al., 2005) was applied to produce the surfaces of grey and white matter. The white matter surfaces were expanded out to the grey matter/cerebrospinal fluid surface boundary using a surface deformation algorithm (Kim et al., 2005). This procedure permits close matching of grey and white matter boundaries and cortical thickness can be calculated based on the distance between the surfaces. This procedure resulted in 40,962 vertices for each hemisphere. The cortical surfaces were non-linearly aligned to a standardized surface template (Lyttelton, Boucher, Robbins, & Evans, 2007). Cortical thickness data were smoothed following surface curvature using a blurring kernel of 20 mm. This technique enhances the identification of cortical thickness changes among the population.
2.5. Scans and surfaces quality assessment
Structural MRIs were visually assessed for scan quality. Great care was taken to assess each anatomical scan both pre- and post-processing to minimize the total number of excluded scans. First, scans were visually inspected by 2 authors (KDT and EGD) for ghosting of images and blurrinesss, and these attributes, if present, were noted. No scans were excluded at this stage. All images were entered into the cortical thickness processing pipeline and subsequently the surfaces were inspected for any bridging, malformation of gyri, sulci and any features that did not compare with anatomical scans. If any of these characteristics were present those scans were excluded.
2.6. Cortical thickness analyses
Cortical thickness analyses were performed using the Matlab (The Mathworks, Natick VA) based program SurfStat (Worsley et al., 2009). The statistical analysis was performed at each vertex of the cortical surface using linear models to examine: (1) differences in cortical thickness between groups controlling for both sex and age; (2) interaction analyses to assess age-related changes between groups (ASD and controls) by regressing cortical thickness against age, controlling for sex; (3) in the ASD group only, the relation between social and communication scores on the ADI-R (most severe) and changes in cortical thickness; (4) lastly, the interaction effects between age and social scores (most severe on the ADI-R). For the latter analysis, a priori ROIs were manually drawn bilaterally on an average anatomical MRI in MNI space constructed from participant scans by one of the authors (EGD). Fig. 1 shows the location and spatial extent of the ROIs. These regions included those in frontal (rostral and dorsal ACC, mPFC, OFC, IFG) and temporal (STG, fusiform gyri) cortices.
Fig. 1.
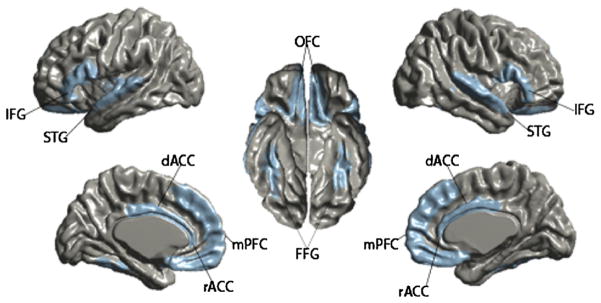
Regions of interest in limbic cortices and social brain network. Depiction of the location and extent of the regions of interest (ROIs) used in the directed search analysis. ROIs were defined around areas implicated in social and communicative functioning that showed significant thickness differences in the whole brain age × group interaction analysis.
The rostral portion of the ACC is anterior and superior to the genu of the corpus callosum. The anterior border was the rostral portion of the cingulate gyrus/marginal segment of the supraorbital sulcus. Posterior boundary was the portion of the gyrus located ~24 mm anterior to the anterior commissure. The superior boundary was the cingulate sulcus and the inferior boundary was the intercommissural plane. The anterior border of the dorsal ACC was just posterior to that of the rostral ACC. The posterior border of the dorsal ACC was the midpoint of the corpus callosum located near the most anterior portion of that thalamus. The superior boundary was the cingulate sulcus and the inferior boundary was the commissural sulcus.
Other frontal lobe structures that were defined included the mPFC, the IFG, and the OFC. The mPFC was comprised of the medial aspect of the superior frontal gyrus that extended from the frontal pole to the anterior commissure. The inferior boundary was the cingulate sulcus. The inferior frontal gyrus was comprised of the most inferior gyri of the frontal lobe and included the pars orbitalis, triangularis, and opercularis. The OFC was comprised of gyrus rectus and the anterior, posterior, lateral and medial oribital gyri extending medially and superiorly to the supraorbital sulcus.
Temporal lobe structures included the STG and fusiform gyrus. The STG extended from the anterior portion of the temporal lobe and posteriorly to the angular gyrus. The superior border was the lateral fissure and the inferior border was the superior temporal sulcus. The fusiform gyrus was defined anteriorly to the medial occipital sulcus and extended posteriorly to the occipital pole. The lateral boundary was the lateral occipitotemporal sulcus and the medial boundary was the collateral sulcus.
The ADI-R scores from the social and communication domain were included in a subsequent linear model with and without age, controlling for sex. The data were corrected for multiple comparisons using the false discovery rate (q < 0.05).
3. Results
3.1. Participants
The scans and surfaces quality assessment resulted in 53 useable scans that allowed us to examine cortical thickness in ASD (n = 28) and typically developing (n = 25) participants between the ages of 7 and 39 years. A significant age difference was found between the excluded group (mean age = 11.05 years) and the included group (mean age = 22.64 years; unpaired t-test, df = 69, t = 5.68, p < 0.05). No significant differences were found between these groups on IQ. ASD participants who were included in the analyses had higher scores in the ADOS social domain compared to the excluded group (included participants: mean score = 7.71; excluded participants: mean score = 5.36, df = 37, t = 2.67, p < 0.05). The ASD groups did not differ on scores in the ADOS communication domain, ADI-R social domain or ADI-R communication domain. The final participant groups were not significantly different in age or full scale IQ based on unpaired t-tests (Table 1). Mean age and IQ scores for ASD and control participants included in the analysis are shown in Table 1.
Table 1.
Demographic characteristics of autism spectrum disorder and typically developing groups.
| ASD (mean, SDa) (95% CIb) | TD (mean, SD) (95% CI) | p value | |
|---|---|---|---|
| Age (years) | Range: 7–39< | Range: 7–37 | t(51) = 0.08, p = 0.93 |
| 22.5 ± 7.9 (19.0, 24.4) | 22.7 ± 9.1 (18.33, 26.0) | ||
| FSIQc | Range: 68–143 | Range: 78–143 | t(51) = 0.72, p = 0.47 |
| 105.9 ± 18.0 (98.03, 112.1) | 109.4 ± 17.0 (102.4, 116.5) | ||
| Gender (M:F)d | 21:7 | 17:8 | Chi-square = 0.6, p = 0.4 |
| ADI-Re social | 17.3 ± 7.3 (14.36, 20.16) |
Standard deviation.
Confidence intervals.
Full scale IQ.
Male:female.
Autism Diagnostic Interview-Revised.
3.2. Whole brain – between group analysis
Based on an analysis of the entire grey matter controlling for sex and age, no significant differences in cortical thickness were observed between groups. In the ASD group compared to controls a trend toward thicker cortices was observed in the rostral ACC, the paracentral lobule, primary somatosensory/motor cortex (M1/S1) and anterior insula (aIns), however these did not survive corrections for multiple comparisons. Typically developing individuals showed a trend towards thicker cortices relative to the ASD group in the IFG, STG and OFC, however these also did not survive corrections for multiple comparisons (all q > 0.05 corrected).
3.3. Whole brain – within group age effects
Within the ASD group, only the inferior frontal gyrus, inferior temporal gyrus (ITG) and the posterior cingulate gyrus (PCG) were thinner with increasing age (all q < 0.05 corrected, Fig. 2A). In controls, cortical thickness reduced with age across the entire grey matter surface (all q < 0.05 corrected, Fig. 2B). All analyses were done controlling for sex.
Fig. 2.
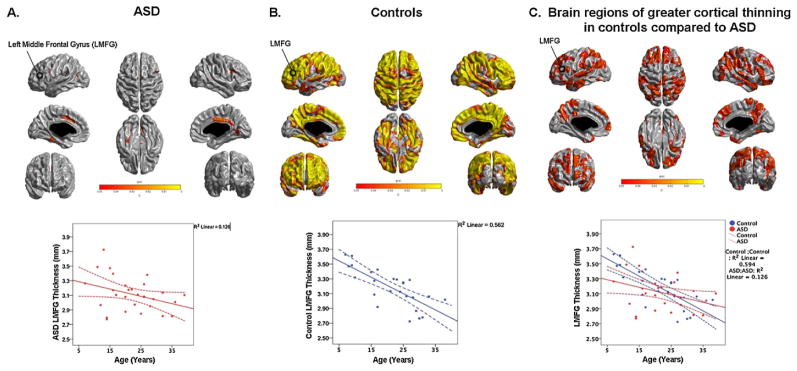
The within group age-effects and age × group interaction effects across the whole brain. The q-map showing regions of the brain where significant developmental changes in cortical thickness were observed in the ASD group (A), control group (B) and where significantly greater cortical thinning with age was observed in the typically developing controls compared to the ASD group (C). An example of thickness changes across age is shown for the left middle frontal gyrus (LMFG) for the ASD and control groups, and for the age × group interaction analysis. The average cortical thickness in a 10 mm radius around the peak in the LMFG is shown. 95% confidence intervals are shown as dotted lines.
3.4. Whole brain – group-by-age interaction analysis
While typically developing children and adults showed thinner cortices with increasing age, significantly less age-related thinning was observed in the ASD group in the superior frontal gyri (SFG), middle frontal gyri (MFG), IFG, medial frontal gyri (MFG), OFC, M1, S1, PCL, superior parietal lobules (SPL), inferior parietal lobules (IPL), middle occipital gyri (MOG), temporal poles, STG, middle temporal gyri (MTG), ITG, medial orbitofrontal gyrus (mOFG), PCG, precuneus and parietoccipital fissure (Fig. 2C and Table 2).
Table 2.
The positive age x group interaction effect across the whole brain.
| Location | Peak vertex MNI coordinates
|
Cluster q-value | ||
|---|---|---|---|---|
| x | y | z | ||
| Superior frontal gyrus | ||||
| Left | −8 | 42 | 54 | 0.03 |
| Right | 8 | 32 | 61 | 0.03 |
| Middle frontal gyrus | ||||
| Left | −29 | 36 | 42 | 0.03 |
| Right | 36 | 27 | 50 | 0.04 |
| Inferior frontal gyrus | ||||
| Left | −57 | 18 | 21 | 0.03 |
| Right | 56 | 25 | 19 | 0.04 |
| Medial frontal gyrus | ||||
| Left | −8 | 60 | 0 | 0.04 |
| Right | 4 | 59 | 14 | 0.05 |
| Orbitofrontal gyrus | ||||
| Left | −16 | 14 | −22 | 0.05 |
| Right | 14 | 40 | −25 | 0.03 |
| Precentral gyrus (M1) | ||||
| Left | −60 | 1 | 13 | 0.03 |
| Right | 33 | −21 | 73 | 0.04 |
| Postcentral gyrus (S1) | ||||
| Left | −50 | −23 | 63 | 0.04 |
| Right | 39 | −30 | 68 | 0.03 |
| Superior parietal lobule | ||||
| Left | −23 | −51 | 70 | 0.03 |
| Right | 16 | −62 | 65 | 0.03 |
| Inferior parietal lobule/middle occipital gyrus | ||||
| Left | −41 | −79 | 35 | 0.03 |
| Inferior parietal lobule | ||||
| Right | 62 | −40 | 41 | 0.04 |
| Posterior/superior temporal gyrus | ||||
| Left | −67 | −26 | 10 | 0.03 |
| Right | 66 | −12 | −18 | 0.03 |
| Middle temporal gyrus | ||||
| Left | −65 | −12 | −18 | 0.03 |
| Right | 66 | −9 | −15 | 0.03 |
| Inferior temporal gyrus | ||||
| Left | −58 | −16 | 29 | 0.03 |
| Right | 60 | −28 | −23 | 0.04 |
| Medial orbitofrontal gyrus | ||||
| Left | −28 | 52 | −13 | 0.04 |
| Posterior cingulate | ||||
| Left | −4 | −32 | 39 | 0.04 |
| Right | 3 | −49 | −28 | 0.03 |
| Precuneus | ||||
| Left | −1 | −73 | 23 | 0.05 |
| Parietoccipital fissure | ||||
| Right | 20 | −68 | 25 | 0.04 |
Brain regions across the entire brain that show significantly greater cortical thinning with age in typically developing controls compared to the ASD group. Coordinates are given in MNI space. Medial-lateral (X), anterior-posterior (Y), and superior-inferior (Z) positive values are right, anterior and superior. All q-values are above the threshold for significance (corrected for multiple comparisons using the False Discovery Rate, q = 0.05).
The age-by-group interaction analysis was repeated after removing the data from the children aged 7–10 years, as this represented a small portion of the sample (n = 6) and may have unduly influenced the results. Results were similar to what was previously found when analysing the data from the entire cohort where the ASD participants showed little age-related thinning in M1, S1, PCL, IPL, STG, mOFG, PCG and parietoccipital fissure. Thus, all participants were included in subsequent analyses.
3.5. Correlation with autism severity measures
3.5.1. Relation between social impairment scores and cortical thickness in ROIs
A directed search in the brain regions (ROIs) implicated in social and communication functions revealed a significant correlation between thicker cortices in the rACC (Montreal Neurological Institute, MNI coordinates; x = 4, y = 28, z = 23; df = 25, q = 0.03) and more severe scores on the ADI-R social domain (t = 2.09, df = 25, p = 0.047, see Fig. 3), but not in the mPFC, IFG or STG. No associations were observed between communication scores on the ADI-R and cortical thickness.
Fig. 3.
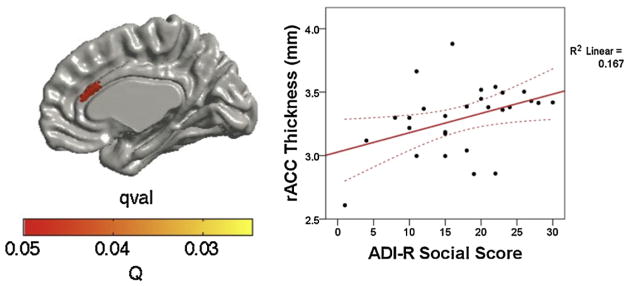
Region of interest analysis correlating cortical thickness against social impairment scores in the rACC. In the ASD group thicker cortices in the rostral anterior cingulate cortex (rACC) were positively associated with greater social impairment as measured by the ADI-R social domain. Data were corrected for multiple comparisons using the false discovery rate of q = 0.05. 95% confidence intervals are shown as dotted lines.
3.5.2. Relation between age, social impairment scores and cortical thickness in ROIs
Significant interaction effects were found in the left orbitofrontal gyrus (MNI coordinates: x = −16, y = 36, z = −26, df = 25, q = 0.014) where reduced cortical thickness was associated with more severe social impairments on the ADI-R in all participants, but was more pronounced in younger participants relative to adults with ASD. No other significant correlations were observed in the selected ROIs between cortical thickness, symptomatology and age.
4. Discussion
The present study provides insight into age-related cortical thickness changes in a well-characterized ASD sample with a wide age range. The results indicate that social impairment is associated with atypical brain morphology in ASD participants.
In this sample of children and adults with and without ASD, when controlling for age, between-group analysis revealed only trends towards thicker cortices in the rACC, PCL, M1/S1 and anterior insula in the ASD group. Some groups have reported increased thickness or density in brain regions in groups of pre-adolescents (Bonilha et al., 2008) and adults (Hyde, Samson, Evans, & Mottron, 2010; Schmitz et al., 2006; Toal et al., 2009) with ASD relative to typically developing individuals. As these prior studies focused on samples with narrower age ranges their results reflect alterations in brain morphology during restricted periods and are not able to examine developmental changes.
However, when examining age-related differences within groups, the ASD group had reduced cortical thinning across age relative to controls, across a large portion of the cortex. To date, few studies have investigated age-related changes in cortical thickness in individuals with ASD and collectively they suggest a complex pattern of cortical thickness maturation in this group. In agreement with the current findings, studies that included participants across a wide age range (Raznahan et al., 2010; Scheel et al., 2011) found more regions with little to no change in cortical thickness in individuals with ASD. In comparison, studies with narrower age ranges report that the cortex thins with age in individuals with ASD. (Mak-Fan et al., 2011; Wallace et al., 2010). This may suggest that the developmental pattern observed in the current study and others with large age-spans is most apparent when examining a wider window of development.
Social impairment was associated with different cortical thickness profiles in the OFC and the rACC in individuals with ASD. The OFC is implicated in social cognitive functions such as affective empathy and arousal (Decety, 2010), social reward processing (Adolphs, 2009) and emotional self-regulation (Bachevalier & Loveland, 2006). The rACC is linked with assessing the salience and motivational value of social stimuli, as well as regulating emotional responses (Bush, Luu, & Posner, 2000). In the OFC, thinner cortices were associated with more severely impacted social function and the rate of thinning was more pronounced in younger participants relative to adults. Conversely, in the rACC, poor social function was related to thicker cortices and did not vary with age, suggesting that this abnormality is present in childhood and persists in a similar form into adulthood.
The observed age-related decrease in cortical thickness in the OFC and its relation to more severe social impairment in ASD is consistent with Hardan et al. (2009), who found similar results across the entire frontal lobe in pre-adolescent boys with ASD over a 2-year period. In the current study, we utilized a higher magnetic field strength (3 T) to acquire the anatomical MRIs which may have improved our ability to detect regional changes (Alvarez-Linera, 2008). In addition, our directed search in the OFC reduced the number of corrections for multiple comparisons, which resulted in a significant correlation between cortical thickness and symptomatology. Our results add to the literature by suggesting that cortical thinning in the OFC may start early in childhood and persist into adulthood.
The results of the present study also show that thicker cortices in the rACC at all ages were associated with more severe social impairment. Increased thickness and other structural atypicalities in the ACC were initially reported in post-mortem studies (Bailey et al., 1998; Kemper & Bauman, 1993). Previous structural MRI studies reported both decreased grey matter volume in the ACC (Abell et al., 1999; Kwon, Ow, Pedatella, Lotspeich, & Reiss, 2004; McAlonan et al., 2002), and increased thickness in this region in individuals with ASD relative to controls (Bonilha et al., 2008; Hyde et al., 2010; Schmitz et al., 2006; Toal et al., 2009), but there were no data on age-related changes in the ACC. The present study is important in its findings of developmental changes in cortical thickness in ASD in the rACC, a region central to the social cognitive functions.
4.1. Limitations
The cross-sectional study design used in the present study is only a proxy for a longitudinal study design, which is better suited for understanding developmental changes. Future studies using a larger sample and a longitudinal study design will further inform our current knowledge of developmental atypicalities in cortical thickness and its relation to social impairment. In the present cohort, a balanced distribution of individuals across the entire age-range would have been most desirable. Unfortunately, there were fewer ASD individuals between the ages of 7 and 10 years than typically developing controls. For the purpose of comparison we removed these extra control participants from the analysis and obtained similar results in many of the regions listed in the full group analysis. Thus developmental differences in cortical thickness across the brain persisted despite this limitation.
In the present study, our focus was on the cortical mantle. Few structural brain morphometric studies have correlated changes in subcortical grey matter volume or cortical density with behavioural impairment in ASD (Hollander et al., 2005; Nacewicz et al., 2006), highlighting the need for more research in this area. Although we observed an association between the morphology of social brain regions, symptomatology and age, it still needs to be determined whether these structural abnormalities are the cause or effect of social impairment in ASD.
5. Conclusions
ASD are neurodevelopmental disorders and therefore age-related changes are crucial to understanding the neurobiology of crucial to understanding the neurobiology of associated behaviour. The present findings underscore the importance of including age interactions when analysing structural properties of the autistic brain, as age effects may contribute to variance in structural indices. Across the age-span of our sample we found significant differences in the development of cortical thickness across the entire brain. In regions implicated in social and communication functions specifically, atypical cortical thickness in the rACC and OFC were associated with impaired social function. These findings raise the question of whether atypical development of cortical thickness in these areas of the brain may be an intermediate phenotype that may be explored further for neurobiological and treatment research.
Acknowledgments
The data for this study were collected at Mount Sinai School of Medicine in New York, New York. The analyses were carried out at Holland Bloorview Kids Rehabilitation Hospital and the Hospital for Sick Children.
Footnotes
Conflict of interest
Dr. Anagnostou has consulted without fees to Neuropharm, Proximagen and Novartis and consulted with fees to Seaside Therapeutics. None of the other authors have any conflicts of interest to disclose.
Contributor Information
Krissy A.R. Doyle-Thomas, Email: kdoylethomas@hollandbloorview.ca.
Emma G. Duerden, Email: emma.duerden@sickkids.ca.
Margot J. Taylor, Email: margot.taylor@sickkids.ca.
Jason P. Lerch, Email: jason@phenogenomics.ca.
Latha V. Soorya, Email: latha.soorya@mssm.edu.
A. Ting Wang, Email: ting.wang@mssm.edu.
Jin Fan, Email: jin.fan@mssm.edu.
Eric Hollander, Email: eholland@montefiore.org.
References
- Abell F, Krams M, Ashburner J, Passingham R, Friston K, Frackowiak R, et al. The neuroanatomy of autism: A voxel-based whole brain analysis of structural scans. Neuroreport. 1999;10:1647–1651. doi: 10.1097/00001756-199906030-00005. [DOI] [PubMed] [Google Scholar]
- Ad-Dab’bagh Y, Lyttelton O, Muehlboeck JS, Lepage C, Einarson D, Mok K, et al. The CIVET image-processing environment: A fully automated comprehensive pipeline for anatomical neuroimaging research. In: Corbetta M, editor. Proceedings of the 12th annual meeting of the organization for human brain mapping; Florence, Italy. 2006. p. 2266. [Google Scholar]
- Adolphs R. The social brain: Neural basis of social knowledge. Annual Review of Psychology. 2009;60:693–716. doi: 10.1146/annurev.psych.60.110707.163514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alvarez-Linera J. 3 T MRI: Advances in brain imaging. European Journal of Radiology. 2008;67:415–426. doi: 10.1016/j.ejrad.2008.02.045. [DOI] [PubMed] [Google Scholar]
- Bachevalier J, Loveland KA. The orbitofrontal-amygdala circuit and self-regulation of social-emotional behavior in autism. Neuroscience & Biobehavioral Reviews. 2006;30:97–117. doi: 10.1016/j.neubiorev.2005.07.002. [DOI] [PubMed] [Google Scholar]
- Bailey A, Luthert P, Dean A, Harding B, Janota I, Montgomery M, et al. A clinicopathological study of autism. Brain. 1998;121:889–905. doi: 10.1093/brain/121.5.889. [DOI] [PubMed] [Google Scholar]
- Baron-Cohen S, Belmonte MK. Autism: A window onto the development of the social and the analytic brain. Annual Review of Neuroscience. 2005;28:109–126. doi: 10.1146/annurev.neuro.27.070203.144137. [DOI] [PubMed] [Google Scholar]
- Bermudez P, Lerch JP, Evans AC, Zatorre RJ. Neuroanatomical correlates of musicianship as revealed by cortical thickness and voxel-based morphometry. Cerebral Cortex. 2009;19:1583–1596. doi: 10.1093/cercor/bhn196. [DOI] [PubMed] [Google Scholar]
- Bernhardt BC, Worsley KJ, Besson P, Concha L, Lerch JP, Evans AC, et al. Mapping limbic network organization in temporal lobe epilepsy using morphometric correlations: insights on the relation between mesiotemporal connectivity and cortical atrophy. Neuroimage. 2008;42:515–524. doi: 10.1016/j.neuroimage.2008.04.261. [DOI] [PubMed] [Google Scholar]
- Bloss CS, Courchesne E. MRI neuroanatomy in young girls with autism: A preliminary study. Journal of the American Academy of Child & Adolescent Psychiatry. 2007;46:515–523. doi: 10.1097/chi.0b013e318030e28b. [DOI] [PubMed] [Google Scholar]
- Bonilha L, Cendes F, Rorden C, Eckert M, Dalgalarrondo P, Li LM, et al. Gray and white matter imbalance—typical structural abnormality underlying classic autism? Brain and Development. 2008;30:396–401. doi: 10.1016/j.braindev.2007.11.006. [DOI] [PubMed] [Google Scholar]
- Bush G, Luu P, Posner MI. Cognitive and emotional influences in anterior cingulate cortex. Trends in Cognitive Sciences. 2000;4:215–222. doi: 10.1016/s1364-6613(00)01483-2. [DOI] [PubMed] [Google Scholar]
- Carper RA, Moses P, Tigue ZD, Courchesne E. Cerebral lobes in autism: Early hyperplasia and abnormal age effects. NeuroImage. 2002;16:1038–1051. doi: 10.1006/nimg.2002.1099. [DOI] [PubMed] [Google Scholar]
- Charil A, Dagher A, Lerch JP, Zijdenbos AP, Worsley KJ, Evans AC. Focal cortical atrophy in multiple sclerosis: relation to lesion load and disability. Neuroimage. 2007;34:509–517. doi: 10.1016/j.neuroimage.2006.10.006. [DOI] [PubMed] [Google Scholar]
- Collins D, Peters T, Evans A. An automated 3D non-linear deformation procedure for determination of gross morphometric variability in the human brain. Proceedings of SPIE. 1994:180–194. 2359. [Google Scholar]
- Courchesne E, Campbell K, Solso S. Brain growth across the life span in autism: Age-specific changes in anatomical pathology. Brain Research. 2011;1380:138–145. doi: 10.1016/j.brainres.2010.09.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Decety J. The neurodevelopment of empathy in humans. Developmental Neuroscience. 2010;32:257–267. doi: 10.1159/000317771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fahim C, Yoon U, Sandor P, Frey K, Evans A. Thinning of the motor-cingulate-insular cortices in siblings concordant for tourette syndrome. Brain Topography. 2009;22:176–184. doi: 10.1007/s10548-009-0105-6. [DOI] [PubMed] [Google Scholar]
- Giedd J, Shaw P, Eckstrand K, Sharp W, Blumenthal J, Lerch JP, et al. Attention deficit/hyperactivity disorder is characterized by a delay in cortical maturation. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:19649–19654. doi: 10.1073/pnas.0707741104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greene CM, Braet W, Johnson KA, Bellgrove MA. Imaging the genetics of executive function. Biological Psychiatry. 2008;79:30–42. doi: 10.1016/j.biopsycho.2007.11.009. [DOI] [PubMed] [Google Scholar]
- Hadjikhani N, Joseph RM, Snyder J, Tager-Flusberg H. Anatomical differences in the mirror neuron system and social cognition network in autism. Cerebral Cortex. 2006;16:1276–1282. doi: 10.1093/cercor/bhj069. [DOI] [PubMed] [Google Scholar]
- Hardan AY, Libove RA, Keshavan MS, Melhem NM, Minshew NJ. A preliminary longitudinal magnetic resonance imaging study of brain volume and cortical thickness in autism. Biological Psychiatry. 2009;66:320–326. doi: 10.1016/j.biopsych.2009.04.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hazlett HC, Poe MD, Gerig G, Smith RG, Piven J. Cortical gray and white brain tissue volume in adolescents and adults with autism. Biological Psychiatry. 2006;59:1–6. doi: 10.1016/j.biopsych.2005.06.015. [DOI] [PubMed] [Google Scholar]
- Hollander E, Anagnostou E, Chaplin E, Esposito K, Haznedar MM, Licalzi E, et al. Striatal volume on magnetic resonance imaging and repetitive behaviors in autism. Biological Psychiatry. 2005;58:226–232. doi: 10.1016/j.biopsych.2005.03.040. [DOI] [PubMed] [Google Scholar]
- Hyde KL, Samson F, Evans AC, Mottron L. Neuroanatomical differences in brain areas implicated in perceptual and other core features of autism revealed by cortical thickness analysis and voxel-based morphometry. Human Brain Mapping. 2010;31:556–566. doi: 10.1002/hbm.20887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kates WR, Burnette CP, Eliez S, Strunge LA, Kaplan D, Landa R, et al. Neuroanatomic variation in monozygotic twin pairs discordant for the narrow phenotype for autism. The American Journal of Psychiatry. 2004;161:539–546. doi: 10.1176/appi.ajp.161.3.539. [DOI] [PubMed] [Google Scholar]
- Kemper TL, Bauman ML. The contribution of neuropathologic studies to the understanding of autism. Neurologic clinics. 1993;11:175–187. [PubMed] [Google Scholar]
- Kim JS, Singh V, Lee JK, Lerch J, Ad-Dab’bagh Y, MacDonald D, et al. Automated 3-D extraction and evaluation of the inner and outer cortical surfaces using a Laplacian map and partial volume effect classification. NeuroImage. 2005;27:210–221. doi: 10.1016/j.neuroimage.2005.03.036. [DOI] [PubMed] [Google Scholar]
- Kwon H, Ow AW, Pedatella KE, Lotspeich LJ, Reiss AL. Voxel-based morphometry elucidates structural neuroanatomy of high-functioning autism and Asperger syndrome. Developmental Medicine and Child Neurology. 2004;46:760–764. doi: 10.1017/s0012162204001306. [DOI] [PubMed] [Google Scholar]
- Lenroot KR, Shmitt JE, Ordaz SJ, Wallace GL, Neale MC, Lerch JP, et al. Differences in genetic and environmental influences on the human cerebral cortex associated with development during childhood and adolescence. Human Brain Mapping. 2009;30:163–174. doi: 10.1002/hbm.20494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerch JP, Pruessner J, Zijdenbos AP, Collins DL, Teipel SJ, Hampel H, et al. Automated cortical thickness measurements from MRI can accurately separate Alzheimer’s patients from normal elderly controls. Neurobiology of Aging. 2008;29:23–30. doi: 10.1016/j.neurobiolaging.2006.09.013. [DOI] [PubMed] [Google Scholar]
- Lerch JP, Worsley K, Shaw WP, Greenstein DK, Lenroot RK, Giedd J, et al. Mapping anatomical correlations across cerebral cortex (MACACC) using cortical thickness from MRI. Neuroimage. 2006;31:993–1003. doi: 10.1016/j.neuroimage.2006.01.042. [DOI] [PubMed] [Google Scholar]
- Lord C, Risi S, Lambrecht L, Cook EH, Leventhal BL, DiLavore PC, et al. The autism diagnostic observation schedule-generic: A standard measure of social and communication deficits associated with the spectrum of autism. Journal of Autism and Developmental Disorders. 2000;30:205–223. [PubMed] [Google Scholar]
- Lord C, Rutter M, Le Couteur A. Autism Diagnostic Interview-Revised: A revised version of a diagnostic interview for caregivers of individuals with possible pervasive developmental disorders. Journal of Autism and Developmental Disorders. 1994;24:659–685. doi: 10.1007/BF02172145. [DOI] [PubMed] [Google Scholar]
- Lyttelton O, Boucher M, Robbins S, Evans A. An unbiased iterative group registration template for cortical surface analysis. NeuroImage. 2007;34:1535–1544. doi: 10.1016/j.neuroimage.2006.10.041. [DOI] [PubMed] [Google Scholar]
- Mak-Fan KM, Taylor MJ, Roberts W, Lerch JP. Measures of cortical grey matter structure and development in children with autism spectrum disorder. Journal of Autism and Developmental Disorders. 2011 doi: 10.1007/s10803-011-1261-6. http://dx.doi.org/10.1007/s10803-011-1261-6. [DOI] [PubMed]
- McAlonan GM, Daly E, Kumari V, Critchley HD, van Amelsvoort T, Suckling J, et al. Brain anatomy and sensorimotor gating in Asperger’s syndrome. Brain. 2002;125:1594–1606. doi: 10.1093/brain/awf150. [DOI] [PubMed] [Google Scholar]
- Meguid N, Fahim C, Yoon U, Nashaat N, Ibrahim A, Mancini-Marie A, et al. Brain morphology in autism and fragile X syndrome correlates with social IQ: First report from the Canadian–Swiss–Egyptian neurodevelopmental study. Journal of Child Neurology. 2010;25:599–608. doi: 10.1177/0883073809341670. [DOI] [PubMed] [Google Scholar]
- Nacewicz BM, Dalton KM, Johnstone J, Long MT, McAuliff EM, Oakes TR, et al. Amygdala volume and nonverbal social impairment in adolescent and adult males with autism. Archives of General Psychiatry. 2006;63:1417–1428. doi: 10.1001/archpsyc.63.12.1417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmen SJ, Hulshoff Pol HE, Kemner C, Schnack HG, Durston S, Lahuis BE, et al. Increased gray-matter volume in medication-naive high-functioning children with autism spectrum disorder. Psychological Medicine. 2005;35:561–570. doi: 10.1017/s0033291704003496. [DOI] [PubMed] [Google Scholar]
- Raznahan A, Lerch JP, Lee N, Greenstein D, Wallace GL, Stockman M, et al. Patterns of coordinated anatomical change in human cortical development: a longitudinal neuroimaging study of maturational coupling. Neuron. 2011;72:873–884. doi: 10.1016/j.neuron.2011.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raznahan A, Shaw P, Lalonde F, Stockman M, Wallace GL, Greenstein D, et al. How does your cortex grow? The Journal of Neuroscience. 2011;31:7174–7177. doi: 10.1523/JNEUROSCI.0054-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raznahan A, Toro R, Daly E, Robertson D, Murphy C, Deeley Q, et al. Cortical anatomy in autism spectrum disorder: An in vivo MRI study on the effect of age. Cerebral Cortex. 2010;20:1332–1340. doi: 10.1093/cercor/bhp198. [DOI] [PubMed] [Google Scholar]
- Scheel C, Rotarska-Jagiela A, Schilbach L, Lehnhardt FG, Krug B, Vogeley K, et al. Imaging derived cortical thickness reduction in high-functioning autism: Key regions and temporal slope. NeuroImage. 2011;58:391–400. doi: 10.1016/j.neuroimage.2011.06.040. [DOI] [PubMed] [Google Scholar]
- Schmitz N, Rubia K, Daly E, Smith A, Williams S, Murphy DGM. Neural correlates of executive function in autistic spectrum disorders. Biological Psychiatry. 2006;59:7–16. doi: 10.1016/j.biopsych.2005.06.007. [DOI] [PubMed] [Google Scholar]
- Schumann CM, Bloss CS, Barnes CC, Wideman GM, Carper RA, Akshoomoff N, et al. Longitudinal magnetic resonance imaging study of cortical development through early childhood in autism. The Journal of Neuroscience. 2010;30:4419–4427. doi: 10.1523/JNEUROSCI.5714-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaw P, Kabani NJ, Lerch JP, Eckstrand K, Lenroot R, Gogtay N, et al. Neurodevelopmental trajectories of the human cerebral cortex. The Journal of Neuroscience. 2008;28:3586–3594. doi: 10.1523/JNEUROSCI.5309-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh V, Chertkow H, Lerch JP, Evans AC, Door AE, Kabani NJ. Spatial patterns of cortical thinning in mild cognitive impairment in Alzheimer’s disease. Brain. 2006;129:2885–2893. doi: 10.1093/brain/awl256. [DOI] [PubMed] [Google Scholar]
- Sled JG, Zijdenbos AP, Evans AC. A nonparametric method for automatic correction of intensity nonuniformity in MRI data. IEEE Transactions on Medical Imaging. 1998;17:87–97. doi: 10.1109/42.668698. [DOI] [PubMed] [Google Scholar]
- Toal F, Bloemen OJN, Deeley Q, Tunstall N, Daly EM, Page L, et al. Psychosis and autism: magnetic resonance imaging study of brain anatomy. The British Journal of Psychiatry. 2009;194:418–425. doi: 10.1192/bjp.bp.107.049007. [DOI] [PubMed] [Google Scholar]
- Wallace GL, Dankner N, Kenworthy L, Giedd JN, Martin A. Age-related temporal and parietal cortical thinning in autism spectrum disorders. Brain. 2010;133:3745–3754. doi: 10.1093/brain/awq279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wechsler D. Manual for the Wechsler Intelligence Scale for Children (WISC-IV) 4. San Antonio, Texas: Psychological Corporation; 2003. [Google Scholar]
- Wechsler D. Wechsler Adult Intelligence Scale. 4. San Antonio, Texas: Pearson; 2008. [Google Scholar]
- Worsley KJ, Taylor JE, Carbonell F, Chung MK, Duerden E, Bernhardt B, et al. SurfStat: A Matlab toolbox for the statistical analysis of univariate and multivariate surface and volumetric data using linear mixed effects models and random field theory. Paper presented at the Human Brain Mapping; San Francisco, California. 2009. [Google Scholar]
- Zhou D, Lebel C, Lepage C, Rasmussen C, Evans A, Wyper K, et al. Developmental cortical thinning in fetal alcohol spectrum disorders. Neuroimage. 2011;58:16–25. doi: 10.1016/j.neuroimage.2011.06.026. [DOI] [PubMed] [Google Scholar]
- Zijdenbos AP, Forghani R, Evans AC. Automatic pipeline analysis of 3-D MRI data for clinical trials: application to multiple sclerosis. IEEE Transactions on Medical Imaging. 2002;21:1280–1291. doi: 10.1109/TMI.2002.806283. [DOI] [PubMed] [Google Scholar]