

Abstract
Protected area (PA) networks will remain valuable for conservation, as the global environment changes, if they facilitate the colonization of new regions by species that are shifting their geographical ranges. We tested the extent to which wetland bird species colonizing the UK since 1960 have exploited PAs. Colonization commenced in a PA for all six species that established permanent (greater than 10 years) breeding populations in the UK during this period. Subsequently, birds started to breed outside as well as inside PAs: the colonizing species showing declining fractions of breeding within PAs over time, a trend not seen in already-resident species. PAs were valuable as ‘landing pads’ for range-shifting species first arriving in a new region, and then as ‘establishment centres’ from which viable populations spread. Given future projections of range change across a broad range of taxonomic groups, this role for PAs can be expected to become increasingly important.
Keywords: adaptation, climate change, conservation, range expansion, Sites of Special Scientific Interest, wetland
1. Introduction
The population size and long-term conservation status of species needing to shift their geographical ranges, for example in response to climate change, will depend not only on their persistence in the regions where they currently occur, but also on their capacity to colonize new areas [1,2]. Facilitating range expansion, therefore, has the potential to become one of the most effective conservation strategies that can be deployed to minimize climatic and other risks to species [3,4]. However, such strategies still lack robust empirical underpinning: there is limited evidence of the effectiveness of static protected areas (PAs) to protect species per se ([5–7] but see [8]), let alone if species distributions become more dynamic [9–13].
While PAs may lose species that were previously present, and indeed those for which the sites were designated, they could conceivably gain others if they safeguard habitats that are colonized by species spreading beyond their former geographical ranges. There is some modelling support for the continuing value of PAs under climate change [14], on the assumption that PAs will contain the most suitable habitats for colonists. There is also empirical evidence that species disproportionately colonize PAs in areas where they have not previously been recorded [15]. However, we still lack information on (i) the extent that PAs act as ‘landing pads’ for species, enabling them to breed and establish for the first time in new regions, and (ii) how their dependency on PAs varies over time. The latter relates to whether PAs act as ‘establishment centres’ from which viable populations can subsequently spread within the region being colonized.
We address these issues by analysing the PA associations of those wetland bird species that have colonized the UK naturally since 1960. Wetland birds account for six of the eight bird species that have established continuous breeding populations in the UK in this period (see below). They are especially suitable for study because of the intensive scrutiny they receive from ornithologists, as well as from formal surveys [16,17]. The breeding distributions of many birds have shifted polewards in recent decades [18–21] and are projected to continue to do so [22]. In the UK, this phenomenon has already been linked with the recent arrival of Cetti's warblers Cettia cetti [23] and little egrets Egretta garzetta [24], two of our focal species. Expansions in other species have been linked with anthropogenic factors such as reduced persecution (e.g. whooper swan Cygnus cygnus [25]) or habitat creation (e.g. Eurasian bittern Botaurus stellaris [26]). However, most species are likely to have been affected by a combination of climatic and non-climatic changes to the environment.
We identify the PA status of the first breeding locations for all 20 wetland bird species that bred for the first time in the UK since 1960 (table 1). For the six species that then established apparently permanent populations, we also evaluate how the percentage of the population breeding in PAs changed over time after initial breeding. We hypothesized that the proportion of the population breeding in PAs would decline over time, as populations grow in PAs (potentially becoming saturated), resulting in birds starting to breed outside of PAs. By contrast, we predicted that such trends would not be present for a ‘control’ group of long-term resident wetland species that have bred continuously in Britain throughout the same period.
Table 1.
Wetland bird species that first bred in the UK after 1960 (during the period 1949–2012). (Those that have not bred for at least 10 consecutive years, prior to 2012, are bold. Population estimates are derived from the RBBP 2010 report unless otherwise stated.)
| species | year first recorded breeding | site first recorded breeding | status of site | current population estimate (minimum confirmed breeding pairs) |
|---|---|---|---|---|
| little egret Egretta garzetta | 1996 | Brownsea Island, Dorset | SSSI | 718 |
| common crane Grus grus | 1981 | Horsey Mere, Norfolk | SSSI | 13 |
| whooper swan Cygnus cygnus | 1978 | An Fhaodhail, Tiree | SSSI | 14 |
| cetti's warbler Cettia cetti | 1973 | Stodmarsh, Kent | SSSI | 1907a |
| goldeneye Bucephala clangula | 1970 | Loch an Eilein, Highlands | SSSI | 196 |
| Mediterranean gull Larus melanocephalus | 1968 | Needs Oar, Hampshire | SSSI | 1016 |
| great white egret Ardea alba | 2012 | Shapwick Heath, Somerset | SSSI | 2—in 2012 |
| purple heron Ardea purpurea | 2010 | Dungeness, Kent | SSSI | 1 |
| cattle egret Bubulcus ibis | 2008 | Undisclosed, Somerset | SSSI | 0 |
| pectoral sandpiperb Calidris melanotus | 2004 | Loch of Strathbeg, Highland | SSSI | 0 |
| spoonbill Platalea leucorodia | 1998 | Orford Ness, Suffolk | SSSI | 8—in 2011 |
| red-necked grebe Podiceps grisegena | 1988 | Undisclosed, Cambridgeshire | non-SSSI | 0 |
| little bittern Ixobyrchus minutus | 1984 | Potteric Carr, Yorkshire | SSSI | 1 |
| black-winged stilt Himantopus himantopus | 1983 | Nene Washes, Cambridgeshire | SSSI | 0 |
| spotted sandpiper Actitis macularius | 1975 | Uig, Skye | non-SSSI | 0 |
| little gull Larus minutus | 1975 | Ouse Washes, Cambridgeshire/Norfolk | SSSI | 0 |
| bluethroat Luscinia svecica | 1968 | Insh Marshes, Highlands | SSSI | 0 |
| black tern Childonius niger | 1966 | Ouse Washes, Cambridgeshire/Norfolk | SSSI | 0 |
| ruff Philomachus pugnax | 1963 | Ouse Washes, Cambridgeshire/Norfolk | SSSI | 0 |
| Savi's warbler Locustella luscinioides | 1960 | Stodmarsh, Kent | SSSI | 0 |
a‘Singing males’, for Cetti's warbler.
bPresumed breeding.
2. Material and methods
(a). Criteria for selecting species
We considered UK wetland bird species (habitat classification based on Gibbons et al. [17]). We identified the first breeding locations for species that first bred in the UK after 1960 (from British Trust for Ornithology; blx1.bto.org/birdfacts), within the 1949 (first designated of Sites of Special Scientific Interest, SSSIs) to 2012 period. We analysed temporal trends in PA use for the six ‘successful colonists’, defined as breeding in at least 10 successive years up until 2012. We analysed 31 ‘comparator species’ to help control for observer effort on and off PAs. Comparators were native wetland bird species that bred throughout 1900–2009 and that had greater than 30 geo-referenced records (from ‘county bird reports’; see the electronic supplementary material). These included species breeding in ‘lowland wetland’ and ‘upland lakes and streams’ habitats [17], ‘coastal’ species that regularly breed inland (shelduck Tadorna tadorna, ringed plover Charadrius hiaticula and cormorant Phalacrocorax carbo), and ‘unclassified’ but primarily wetland species (black-headed gull Chroicocephalus ridibundus, common tern Sterna hirundo and sand martin Riparia riparia).
(b). Obtaining data
For colonizers, data on breeding locations since 1973 were obtained from the Rare Breeding Birds Panel (RBBP: see ‘species list’; http://www.rbbp.org.uk/rbbp-species-list-full.htm). For pre-1973 data, and instances when RBBP referred only to county totals, we obtained site data from the relevant ‘county bird reports’, which are annual compilations of UK bird records by region. When county bird reports lacked sufficient detail, we consulted county bird recorders, who hold historical records of birds submitted within each region. For goldeneye Bucephala clangula, we searched the nest-box record cards (in this case held by the Royal Society for the Protection of Birds), which contain locations of erected nest-boxes and records of breeding attempts in each box. For comparator species, breeding locations were obtained from county bird reports (1964–2009) of 10 counties (see the electronic supplementary material). Counties were selected based on the availability of county bird reports for each year, while maximizing their latitudinal and longitudinal spread.
(c). Determining protected area status of records
The PAs considered are UK SSSIs (Areas of Special Scientific Interest (ASSIs) in Northern Ireland) that correspond to The International Union for Conservation of Nature level IV of protection [27]. Breeding records were cross-referenced against PA location using the interactive mapping software provided by Natural England (NE; http://www.natureonthemap.naturalengland.org.uk), the Countryside Council for Wales (CCW; http://www.ccw.gov.uk/landscape–wildlife/protecting-our-landscape/protected-sites-map.aspx), Scottish Natural Heritage (SNH; http://gateway.snh.gov.uk/sitelink/searchmap.jsp) and the Northern Ireland Environment Agency (NIEA; http://maps.ehsni.gov.uk/NIEAProtectedAreas/Default.aspx#). Records with grid references were assigned PA status if the entire grid cell (normally 100 × 100 m resolution) was within the boundary of a PA. Records without grid references were assigned PA status only if associated with site/reserve names for which PA status was unambiguous. County bird recorders were consulted for clarification when grid references partially overlapped PAs or location names were vague; if still ambiguous, the records were omitted.
We obtained PA notification dates from SSSI/ASSI citation documents from NE (www.naturalengland.org.uk), CCW (www.ccw.gov.uk), SNH (www.snh.gov.uk) and NIEA (maps.ehsni.gov.uk). To avoid instances of a PA being designated because of the arrival of a colonist, we identified sites that were designated after a colonist had begun breeding there. This situation applied only to Cetti's warbler and whooper swan, for which 20 out of 843 (2.37%) and one out of 21 (4.76%) sites, respectively, were designated after their arrival. Records from these sites were excluded if a breeding population of the relevant colonist was cited as a principal reason for notification. This was only the case for two Cetti's warbler sites. Excluding all records from sites that were designated after arrival did not change the conclusions for either species (see the electronic supplementary material, figure S1).
(d). Calculating protected area associations
For each colonizing species, we calculated the percentage of ‘confirmed’ breeding population (pairs) in PAs each year and the percentage of localities in PAs for colonial breeders [28]. ‘Confirmed breeding’ pairs follows the European Bird Census Council definition [29] for each species, apart from the elusive Cetti's warbler, for which we used the number of singing males (as reported by the RBBP [30]).
For each comparator species, we estimated the percentage breeding in PAs in every fifth year between 1964 and 2009. Breeding is rarely ‘confirmed’ for established species, so we used ‘probable breeding’ [29] records for each species, apart from the elusive reed warbler and sedge warbler for which we used numbers of singing males. For each species, we estimated the percentage of breeding pairs present on PA land in a given year, provided that at least 10 breeding pairs were recorded. If fewer pairs were recorded, the interval was binned with the following available year until greater than 10 breeding pairs was achieved. Ambiguous reports of exact numbers (e.g. ‘several’ or ‘breeding was recorded’) at a given location were analysed as ‘two pairs’. This uncertainty only really affected seven of the 31 comparator species (for which greater than 5% of records were ambiguous). Nonetheless, we carried out a complementary analysis of the percentage of sites from which breeding was reported for all 31 comparator species and obtained similar results (see the electronic supplementary material, figure S2).
We also analysed Cetti's warbler in two separate counties which were colonized in different years to distinguish the effects of year-since-colonization from year-per se on PA use. Cetti's warbler was selected because it is by far the most numerous and widespread colonist, and the counties were chosen as they provided the most complete continuous set of records for this species: Norfolk between the initial establishment of a population (10 singing males) in 1975 and a temporary population crash in 1991, and Hampshire between initial establishment in 1980 and 2008.
For each colonizing and comparator species, we calculated Spearman rank values for the correlation between year and percentage breeding in PAs to determine the temporal trend. Mean ‘PA usage’ for each species was estimated as the average percentage of population breeding in PAs at each time interval.
3. Results
(a). Coverage
Six wetland bird species have established persistent (greater than 10 years) breeding populations in the UK since 1960. For four of these, whooper swan C. cygnus, little egret, common crane Grus grus and Mediterranean gull Larus melanocephalus, we were able to categorize over 95 per cent of all recognized UK records (based on RBBP data) as in/out of a PA. For Cetti's warblers, the proportion of the population that we could designate as in/out of PA was greater than 90 per cent for the first 10 years, but declined as the population increased. Records were incomplete for goldeneye, and varied year-to-year (see §4).
(b). First breeding records of colonizing species
The first breeding record of each of the six successful colonists was in a PA (table 1). Including species breeding but not (yet) established for greater than 10 years, 18 of 20 species first bred in PAs in the UK (table 1).
(c). Trends in breeding in protected areas
Five of the six colonizing species showed a similar pattern, with most early breeding records in PAs, but the percentage of the population breeding in PAs declining over time (figure 1). This trend was significant for the three species (Spearman's rank correlations: little egrets n = 14, rs = −0.91, p = 0.001; Mediterranean gulls n = 34, rs = −0.51, p = 0.0034; Cetti's warbler n = 37, rs = −0.61, p = 0.0002; all less than critical p = 0.0083 after Bonferroni correction for six tests) that (i) currently have the largest established breeding populations, and (ii) are ‘southerly’ species, whose range expansions have been associated with climatic change (see the electronic supplementary material, table S1). The sixth species, goldeneye, showed an idiosyncratic pattern (figure 1), apparently driven by the availability of nest-boxes and frequency with which they were checked and reported (see the electronic supplementary material, table S1).
Figure 1.

The percentage of breeding pairs (or singing males for Cetti's warbler) of each colonizing species in PAs each year since colonization (closed circles). For colonial species, the percentage of breeding localities (open circles) within PAs each year is also shown. The squares for goldeneye denote that some data are known to be missing. Lines represent population estimates based on RBBP reports (see the electronic supplementary material). Spearman's rank values (lower right of each panel) describe correlations between year-since-arrival and percentage breeding in PAs (*p < 0.05 and **p < 0.01).
For comparator species, 13 out of 31 species showed declining temporal trends for percentage breeding within PAs, but 18 species showed positive trends (see the electronic supplementary material, figure S3; figure 2), indicating no overall pattern of increasing or decreasing association with PAs (binomial test: p = 0.47). Nonetheless, five of the 13 negative trends (mallard n = 10, rs = −0.77, p = 0.009; Eurasian teal n = 10, rs = −0.74, p = 0.014; grey heron n = 10, rs = −0.75, p = 0.013; black-headed gull n = 10, rs = −0.68, p = 0.029; common tern n = 10, rs = −0.66, p = 0.038) and two of the 18 positive trends (little grebe n = 8, rs = 0.76, p = 0.028; moorhen n = 10, rs = 0.90, p = 0.002) reached nominal significance at p = 0.05. These would not attain individual significance after Bonferroni correction for 31 tests (critical p = 0.0016), but the probability of obtaining seven or more individually significant trends at less than p = 0.05, out of 31, is itself unlikely (Binomial test: p = 0.0008), suggesting that some of the trends are real.
Figure 2.
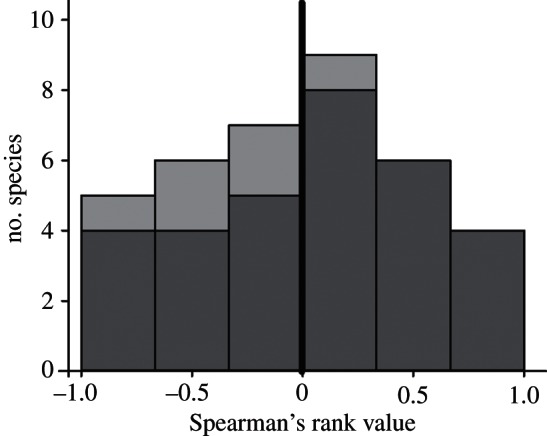
Spearman's rank correlation value describing relationships between year-since-arrival and percentage breeding in PAs for colonizers (light grey) and comparator species (dark grey).
Spearman's rank correlation values for the colonizers were significantly more negative than for the comparators (figure 2; Mann–Whitney, z6,31 = −2, two-tailed p = 0.046; electronic supplementary material, figure S3), indicating that colonists showed declining patterns of association with PAs over time, compared with the long-term resident comparator species.
(d). Overall protected area dependence
PA dependence varied among species for both the colonists and the comparators (figure 3), with riparian birds such as grey wagtails Motacilla cinerea and dippers Cinclus cinclus breeding infrequently in PAs, and reed bed specialists such as bearded tits Panurus biarmicus breeding almost exclusively within PAs (see the electronic supplementary material, figure S3). Although the colonists were clustered at the top end of PA dependency (figure 3), there was no significant difference between the overall PA dependency of colonists and comparator species (Mann–Whitney, z6,31 = 1.59, p = 0.112, two-tailed).
Figure 3.
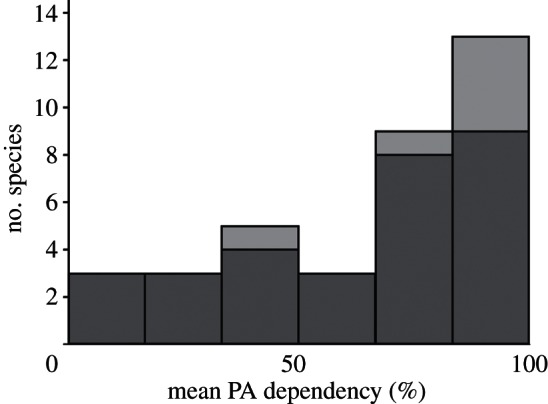
Mean overall PA dependency for colonizers (light grey) and comparator species (dark grey).
(e). Trends at different times
Different species initiated their declining association with PAs at different times (figure 1). Percentages of Cetti's warblers breeding in PAs have been declining since 1975; whooper swans since the early 1990s; Mediterranean gull and little egret since the mid-1990s and common cranes between 2006 and 2008.
Percentages of Cetti's warblers breeding in PAs started to decline earlier in Hampshire (which was colonized earlier) than in Norfolk. The pattern in Norfolk then followed that in Hampshire (figure 4).
Figure 4.

The percentages of Cetti's warblers breeding in PAs each year in (a) Norfolk and (b) Hampshire.
4. Discussion
The conservation value of PAs will be maintained and in some cases enhanced if they can facilitate the colonization of new regions by species whose geographical ranges are expanding. Our results suggest that a PA network can be effective in this context.
Although avian colonizations of new areas are not unique to recent years [31], an apparently increasing number of wetland birds have arrived in the UK over the last half-century and begun to breed. This appears to have happened for a variety of reasons, but primarily as a result of climatic factors and reductions in persecution (see the electronic supplementary material, table S1). Each ‘successful’ colonization started off in a PA, but, as populations became more established, breeding spread into additional sites, not all of which were PAs (figure 1). PAs provided suitable habitat for wetland birds, initially as ‘landing pads’ where they first bred upon arrival, and as ‘establishment centres’, from which populations subsequently spread to other locations in the same region. The goldeneye was a partial exception, perhaps because some of the early data were incomplete or unavailable for this species, and its colonization was affected by the widespread erection of nest-boxes outside as well as inside PAs (see the electronic supplementary material, table S1). Nonetheless, even this species was concentrated in PAs during its initial establishment (figure 1).
Most wetland bird species that have not yet established in the UK also bred for the first time in PAs (table 1), as did the two non-wetland bird species that met our criteria of ‘successful colonists’ since 1960 (firecrests Regulus incapillus on a SSSI in Hampshire [32]; yellow-legged gulls Larus michahellis on a SSSI in Dorset; RBBP data). The geographical ranges of birds are expected to continue to change [22], and thus the trend of wetland birds colonizing the UK could continue. Evidence already exists to show that wetland birds might experience stronger range margin shifts than birds associated with other habitats [33]. Our findings corroborate this by showing that most recent colonizers were wetland birds. A future area of study might address whether this high proportion of wetland birds is a function of PAs being particularly attractive to this group, or whether they are intrinsically more prone to range change and then subsequently use PAs.
Whichever the reason, our results suggest that future breeding populations of these birds in the UK will most probably be centred on protected sites, before expanding into additional undesignated locations. This is concordant with studies which have suggested that PAs will remain important for conservation under climate change [14,15] and provides a contrast to the conclusions of research on alien invasive species, whose colonizing distributions are typically associated with landscapes affected by human activity [34].
The records that we use here to determine the percentages of birds breeding in PAs are a product of casual observations as opposed to systematic surveys, and we are aware that a bias in observer effort towards PAs could lead to a bias in the proportion of records that come from PAs. Resultantly, we took a number of steps to evaluate the robustness of our findings in this context: (i) there was no systematic declining trend in the percentage of comparator species being reported from PAs (figure 2) implying that there was no general shift away from PAs in terms of observer coverage of wetland birds during this period; (ii) individual colonists showed declining trends at different points in time (figure 1), and (iii) the association of one colonist species with PAs declined at different times in different British counties (figure 4), both implying that there was no ‘general’ temporal shift in observer effort; and further (iv) five of the colonizing species are large-bodied, conspicuous birds (the sixth has a loud and characteristic song) that, given the high density of birdwatchers in the UK [19] and novelty of their occurrence in Britain, would be extremely unlikely to be overlooked and not reported regardless of the designation of any particular site. We are confident, therefore, that the results are robust and are not artefacts of changes in the historical distribution of observer effort.
For the comparator species, we are aware that trends in PA use (see the electronic supplementary material, figure S3) are likely to be explicitly related to population trends. Owing to the lack of available population data for each comparator species over the time period covered, an analysis of this relationship was not possible here, but would provide an interesting avenue for further research.
In conclusion, PAs have represented ‘establishment centres’ for wetland bird species colonizing the UK since 1960. Colonizing birds bred in these sites for the first time, establishing populations and then expanding into additional unprotected sites. Hence, PAs enable species to establish in new regions in addition to their benefits to species already established within them.
Acknowledgements
We are grateful to all of the numerous volunteer bird recorders and those involved in the production of county bird reports. We give thanks to Natural England, Scottish Natural Heritage, the Countryside Council for Wales and the Northern Ireland Environment Agency for the use of their online software, and particular thanks to the following individuals for help with the provision of data: Andrew Henderson (Kent Ornithological Society), Keith Betton (Hampshire Ornithological Society), and Pete Moore and Andrew Stanbury (Royal Society for the Protection of Birds). We thank Malcolm Ausden for his support at each stage of the research, and the Royal Society for the Protection of Birds for financial support.
References
- 1.Thomas CD, et al. 2004. Extinction risk from climate change. Nature 427, 145–148 10.1038/nature02121 (doi:10.1038/nature02121) [DOI] [PubMed] [Google Scholar]
- 2.Williams P, Hannah L, Andelman S, Midgley G, Araújo M, Hughes G, Manne L, Martinez-Meyer E, Pearson R. 2005. Planning for climate change: identifying minimum-dispersal corridors for the Cape Proteaceae. Conserv. Biol. 19, 1063–1074 10.1111/j.1523-1739.2005.00080.x (doi:10.1111/j.1523-1739.2005.00080.x) [DOI] [Google Scholar]
- 3.Heller NE, Zavaleta ES. 2009. Biodiversity management in the face of climate change: a review of 22 years of recommendations. Biol. Conserv. 142, 14–32 10.1016/j.biocon.2008.10.006 (doi:10.1016/j.biocon.2008.10.006) [DOI] [Google Scholar]
- 4.Pearce-Higgins JW, Bradbury RB, Chamberlain DE, Drewitt A, Langston RHW, Willis SG. 2011. Targeting research to underpin climate change adaptation for birds. Ibis 153, 207–211 10.1111/j.1474-919X.2010.01086.x (doi:10.1111/j.1474-919X.2010.01086.x) [DOI] [Google Scholar]
- 5.Gaston KJ, Jackson SF, Cantú-Salazar L, Cruz-Piñón G. 2008. The ecological performance of protected areas. Annu. Rev. Ecol. Evol. Syst. 39, 93–113 10.1146/annurev.ecolsys.39.110707.173529 (doi:10.1146/annurev.ecolsys.39.110707.173529) [DOI] [Google Scholar]
- 6.Gaston KJ, Charman K, Jackson SF, Armsworth PR, Bonn A, Briers R. 2006. The ecological effectiveness of protected areas: the United Kingdom. Biol. Conserv. 132, 76–87 10.1016/j.biocon.2006.03.01 (doi:10.1016/j.biocon.2006.03.01) [DOI] [Google Scholar]
- 7.Fuller RA, McDonald-Madden E, Wilson KA, Carwardine J, Grantham HS, Watson JEM, Klein CJ, Green DC, Possingham HP. 2010. Replacing underperforming protected areas achieves better conservation outcomes. Nature 466, 365–367 10.1038/nature09180 (doi:10.1038/nature09180) [DOI] [PubMed] [Google Scholar]
- 8.Donald PF, Sanderson FJ, Burfield IJ, Bierman SM, Gregory RD, Wallczky Z. 2007. International conservation policy delivers benefits for birds in Europe. Science 317, 810–813 10.1126/science.1146002 (doi:10.1126/science.1146002) [DOI] [PubMed] [Google Scholar]
- 9.Hannah L, Midgley G, Hughes G, Bomhard B. 2005. The view from the Cape: extinction risk, protected areas, and climate change. Bioscience 55, 231–242 10.1641/0006-3568(2005)055[0231:TVFTCE]2.0.CO;2 (doi:10.1641/0006-3568(2005)055[0231:TVFTCE]2.0.CO;2) [DOI] [Google Scholar]
- 10.Dockerty T, Lovett A, Watkinson A. 2003. Climate change and nature reserves: examining the potential impacts, with examples from Great Britain. Glob. Environ. Change 13, 125–135 (doi:10.1016/S0959-3780(03)00010-4) [Google Scholar]
- 11.Araújo MB, Cabeza M, Thuiller W, Hannah L, Williams PH. 2004. Would climate change drive species out of reserves? An assessment of existing reserve-selection methods. Glob. Change Biol. 10, 1618–1626 10.1111/j.1365-2486.2004.00828.x (doi:10.1111/j.1365-2486.2004.00828.x) [DOI] [Google Scholar]
- 12.Hannah L, Midgley G, Andelman S, Araújo M, Hughes G, Martinez-Meyer E, Pearson R, Williams P. 2007. Protected area needs in a changing climate. Front. Ecol. Environ. 5, 131–138 10.1890/1540-9295(2007)5[131:PANIAC]2.0.CO;2 (doi:10.1890/1540-9295(2007)5[131:PANIAC]2.0.CO;2) [DOI] [Google Scholar]
- 13.Araújo MB, Alagador D, Cabeza M, Nogués-Bravo D, Thuiller W. 2011. Climate change threatens European conservation areas. Ecol. Lett. 14, 484–492 10.1111/j.1461-0248.2011.01610.x (doi:10.1111/j.1461-0248.2011.01610.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hole DG, Willis SG, Pain DJ, Fishpool LD, Butchart SHM, Collingham YC, Rahbek C, Huntley B. 2009. Projected impacts of climate change on a continent-wide protected area network. Ecol. Lett. 12, 420–431 10.1111/j.1461-0248.2009.01297.x (doi:10.1111/j.1461-0248.2009.01297.x) [DOI] [PubMed] [Google Scholar]
- 15.Thomas CD, et al. 2012. Protected areas facilitate species’ range expansions. Proc. Natl Acad. Sci. USA 149, 14 063–14 068 10.1073/pnas.1210251109 (doi:10.1073/pnas.1210251109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sharrock JTR. 1976. The atlas of breeding birds in Britain and Ireland. London, UK: T & AD Poyser [Google Scholar]
- 17.Gibbons DW, Reid JB, Chapman RA. 1994. The new atlas of breeding birds in Britain and Ireland: 1988–1991. London, UK: T & AD Poyser [Google Scholar]
- 18.Thomas CD, Lennon JJ. 1999. Birds extend their ranges northwards. Nature 399, 213–213 10.1038/20335 (doi:10.1038/20335) [DOI] [Google Scholar]
- 19.Brommer JE. 2004. The range margins of northern birds shift polewards. Ann. Zool. Fenn. 41, 391–397 [Google Scholar]
- 20.Hitch AT, LeBerg PL. 2007. Breeding distributions of north American bird species moving north as a result of climate change. Conserv. Biol. 21, 534–539 10.1111/j.1523-1739.2006.00609.x (doi:10.1111/j.1523-1739.2006.00609.x) [DOI] [PubMed] [Google Scholar]
- 21.Maclean IMD, et al. 2008. Climate change causes rapid changes in the distribution and site abundance of birds in winter. Glob. Change Biol. 14, 2489–2500 10.1111/j.1365-2486.2008.01666.x (doi:10.1111/j.1365-2486.2008.01666.x) [DOI] [Google Scholar]
- 22.Huntley B, Green RE, Collingham YC, Willis SG. 2007. A climatic atlas of European breeding birds. Barcelona, Spain: The RSPB and Lynx Edicions [Google Scholar]
- 23.Bonham PF, Robertson JCM. 1975. The spread of Cetti's warbler in north-west Europe. Br. Birds 68, 393–408 [Google Scholar]
- 24.Lock L, Cook K. 1998. The little egret in Britain: a successful colonist. Br. Birds 9, 273–280 [Google Scholar]
- 25.Boiko D, Kampe-Persson H. 2010. Breeding whooper swans Cygnus cygnus in Latvia, 1973–2009. Wildfowl 60, 168–177 [Google Scholar]
- 26.Gilbert G, Brown AF, Wotton SR. 2010. Current dynamics and predicted vulnerability to sea-level rise of a threatened bittern Botaurus stellaris population. Ibis 152, 580–589 10.1111/j.1474-919X.2010.01022.x (doi:10.1111/j.1474-919X.2010.01022.x) [DOI] [Google Scholar]
- 27.Dudley N. 2008. Guidelines for applying protected area management categories. Gland, Switzerland: IUCN Publication; See http://data.iucn.org/dbtw-wpd/edocs/PAPS-016.pdf [Google Scholar]
- 28.Svensson L. 2009. Collins bird guide second edition. London, UK: Harper Collins [Google Scholar]
- 29.Hagemeijer WJM, Blair MJ. 1997. The EBCC atlas of European breeding birds. London, UK: T & AD Poyser [Google Scholar]
- 30.Holling M. 2009. Rare breeding birds in the United Kingdom in 2006. Br. Birds 102, 158–202 [Google Scholar]
- 31.von Haartman L. 1973. Changes in the breeding bird fauna of northern Europe. In Breeding biology of birds (ed. Farner DS.), pp. 448–481 Washington, DC: National Academy of Sciences [Google Scholar]
- 32.Batten LA. 1973. The colonisation of England by the firecrest. Br. Birds 66, 159–166 [Google Scholar]
- 33.Brommer JE. 2008. Extent of recent polewards range margin shifts in Finnish birds depends on their body mass and feeding ecology. Ornis Fenn. 85, 109–117 [Google Scholar]
- 34.Westphal MI, Browne M, MacKinnon K, Noble I. 2008. The link between international trade and the global distribution of invasive alien species. Biol. Invasions 10, 391–398 10.1007/s10530-007-9138-5 (doi:10.1007/s10530-007-9138-5) [DOI] [Google Scholar]