

Abstract
Forecasting invasion success remains a fundamental challenge in invasion biology. The effort to identify universal characteristics that predict which species become invasive has faltered in part because of the diversity of taxa and systems considered. Here, we use an alternative approach focused on the spread stage of invasions. FST, a measure of alternative fixation of alleles, is a common proxy for realized dispersal among natural populations, summarizing the combined influences of life history, behaviour, habitat requirements, population size, history and ecology. We test the hypothesis that population structure in the native range (FST) is negatively correlated with the geographical extent of spread of marine species in an introduced range. An analysis of the available data (29 species, nine phyla) revealed a significant negative correlation (R2 = 0.245–0.464) between FST and the extent of spread of non-native species. Mode FST among pairwise comparisons between populations in the native range demonstrated the highest predictive power (R2 = 0.464, p < 0.001). There was significant improvement when marker type was considered, with mtDNA datasets providing the strongest relationship (n = 21, R2 = 0.333–0.516). This study shows that FST can be used to make qualitative predictions concerning the geographical extent to which a non-native marine species will spread once established in a new area.
Keywords: alien species, invasiveness, introduction success, meta-analysis, genetic structure
1. Introduction
The rate of species introductions has increased dramatically in modern times correlating with human population growth, increased international trade and advances in transportation [1–3]. While most introduced species never become established, those that persist can have serious economic impacts [4,5], consequences to human health [6] and pose a threat to biodiversity and ecosystem function [7–9]. Wilcove et al. [10] estimated that 42 per cent of endangered species in the United States are under direct threat from invasive species: an estimate that is much higher in rare taxa [11]. Consequently, biological invasions are regarded as one of the greatest modern threats to global biodiversity [12].
Forecasting which species will become invasive and which ecosystems are most vulnerable is of great scientific and practical interest [3]. Despite considerable efforts, the identification of universal characteristics that predict the success of invasive species remains elusive [13,14]. However, failure to find generalized predictive traits is not surprising given the diversity of taxa and ecosystems subject to invasion. Furthermore, characteristics important to invasion success are likely to vary among the different stages of invasion. The accumulating evidence indicates that taxon specific traits such as reproductive strategy, growth rate, environmental tolerances and diet specificity [13,15,16], combine with introduction dynamics such as habitat match and propagule pressure [17–19], to produce successful invaders [13,14]. Distilling this complexity down to even a few metrics that predict invasion success across taxa would be valuable to ecologists and managers working to control introduced species. One possible metric is FST, a common measure of population structure based on the alternate fixation of alleles between populations (reviewed by Holsinger & Weir [20] and Bird et al. [21]). Because only individuals that survive dispersal events, find suitable habitat and successfully reproduce are contributing to population gene pools, FST is a potential proxy for realized dispersal (but see [22–24]). While F-statistics have been used to estimate genetic differentiation and to infer migration rates among native populations (i.e. [25]), they have not previously been used to predict the outcome of introduction events.
(a). The role of FST in predicting invasion success: a Hawaiian case study
The relationship between FST and invasion success first came to our attention while studying introduced fishes in Hawai‘i. During the 1950s three fishes: the bluestripe snapper, Lutjanus kasmira, the blacktail snapper, Lutjanus fulvus and the peacock hind, Cephalopholis argus were deliberately introduced into Hawaiian waters [26–28]. These three species were introduced during the same time period and in roughly equal numbers (n = 2204–3175), yet they demonstrated contrasting patterns of success [28]. Lutjanus kasmira, with the widest distribution in Hawai‘i, demonstrates little genetic structure (low FST) across nearly 20 000 km of its natural range (figure 1; [29,30]). By contrast, L. fulvus, with the smallest Hawaiian distribution of the three species, showed significant population structure at all geographical scales [30]. Cephalopholis argus demonstrated an intermediate pattern [31]. Given the myriad of factors that influence invader success, we were surprised by the relationship between population structure in the native range and the extent of spread demonstrated by these introduced fishes. This finding prompted the question of whether this relationship is broadly applicable to marine invaders. Here, we present an analysis of the available data, across 29 species and nine phyla, to determine whether there is a significant correlation between FST, as a summary statistic surrogate for realized dispersal, and the extent of spread (invasiveness) in introduced marine species.
Figure 1.
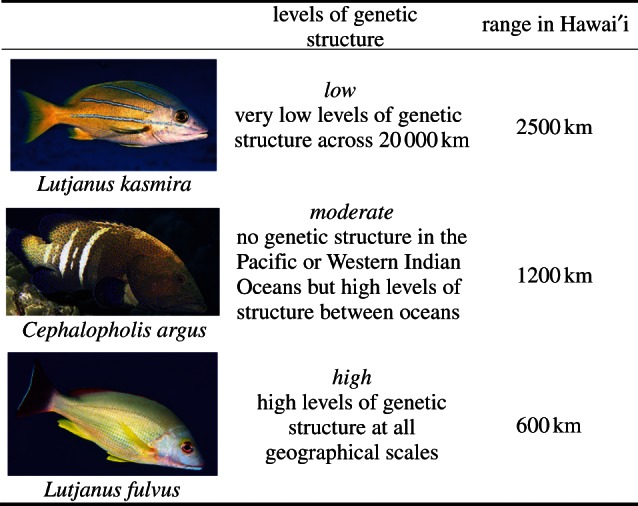
Genetic structure in the native range for each of the three fishes introduced to Hawai‘i. Level of genetic structure is described and the extent of spread within the introduced range is provided.
2. Material and methods
(a). Literature search
We conducted a Web of Science search using the following Boolean combination (phylogeography, genetic, molecular), and (invasive, introduced, alien species) to identify invasive species for which genetic surveys in the native range have been conducted. The initial literature search was conducted in August 2011. The search resulted in 125 species of potential interest (see the electronic supplementary material, table S1). For each of these 125 taxa, an additional search based on the species scientific name was conducted. For inclusion in our study, we set the following criteria: (i) the species must be primarily marine or estuarine (diadromous species were not considered), (ii) a genetic survey of at least three native locations must be available, and (iii) the extent of spread in the introduced region must be documented. The majority of species were omitted from consideration owing to the lack of published population genetic data. Suspected species complexes and species with ambiguous native ranges were omitted from the dataset (i.e. Botryllus schlosseri, Ciona intestinalis and Styela clava). For each case study, we recorded marker type and global and pairwise FST values as reported in the published literature. Because demographic data are rarely available for marine species we recorded expected heterozygosity (HE) as a proxy for native population size. In studies where Mantel tests were conducted to test for isolation by distance (IBD) the slope of the regression was recorded. The extent of native range surveyed (km) was calculated as the shortest straight-line overwater distance between the two most distant sample sites. If desired FST values were not reported, haplotype/allele frequency files were created from the data published in tables in conjunction with sequences downloaded from GenBank (http://www.ncbi.nlm.nih.gov/genbank/). When sufficient data were not available, authors were contacted and the missing data were requested. From these reconstructed datasets, HE was calculated and global and pairwise FST [32] values between sample locations were estimated using the ‘compute pairwise differences’ option and 20 000 permutations in arlequin [33].
In cases such as the European green crab, Carcinus maenas, where multiple surveys of the native range were conducted using the same genetic marker [34,35], we used the study that reported the most complete dataset and/or covered the greatest geographical area (for C. maenas data from Darling et al. [34]). For two species (C. maenas and the sea walnut, Mnemiopsis leidyi), data from two marker types were included (see the electronic supplementary material, table S2). Several species have been introduced to multiple regions around the globe (i.e. Microcosmus squamiger, Caprella mutica, Nematostella vectensism and Mya arenaria). In these cases, we evaluated the extent of spread in the region with the earliest date of detection. The edible alga wakame (Undaria pinnatifida) is cultured in its native Asian range and has since been spread around the globe. We analysed the introduction to Argentina because this was the earliest introduction harbouring haplotypes from natural populations in the native range.
(b). Defining the extent of spread
The maximum extent of spread (MES) in the introduced range was defined as the shortest straight-line overwater distance (km) between the two furthest points in the introduced range. This is a highly conservative estimate of range expansion and does not account for multiple introduction events, secondary introductions or human-mediated dispersal within the introduced range. Disjunct distributions of alien species are common and likely reflect multiple or secondary introductions. To account for multiple introductions, we measured a second metric: the continuous extent of spread (CES). CES is defined as the geographical distance over which there is suitable habitat and no gaps in distribution of greater than 100 km. For example, the Caribbean barnacle Chthamalus proteus, was first recorded on the Hawaiian island of O‘ahu in 1995. This species has since been recorded throughout the Main Hawaiian Islands and at Midway Atoll over 2000 km northwest of O‘ahu. Chthamalus proteus has not been detected at intermediate locations within the archipelago despite suitable habitat. MES for this species was, therefore, recorded as 2489 km (the distance between Hawai‘i Island and Midway Atoll), whereas CES is recorded as 529 km (the distance between the islands of Hawai‘i and Kaua‘i) to reflect the possibility that C. proteus was secondarily transported to Midway Atoll on the hull of a boat or on fishing gear, rather than naturally jumping the intervening 1960 km.
(c). Relationship between FST and extent of spread
Using the pairwise FST values between populations in the native range, we calculated mean, median and mode values. In preliminary analyses, minimum and maximum FST were considered but were later omitted because these measures of genetic structure are heavily influenced by sampling scale and do not correlate well with other measures of FST. All data were log-transformed prior to analyses including FST values (global, mean, median and mode) and geographical distances (km). Because values of FST can be zero or negative, we added 1 to each value prior to transformation [ln(FST + 1)]. Likewise, because the geographical scale over which genetic surveys are conducted (distance between two most distant sample sites) can influence the magnitude of genetic structure, we standardized global FST values by calculating FST per kilometre [ln(global FST + 1)/ln(km)]. We used ordinary least-squares (OLS) regression analyses to determine if FST values (global FST, global FST/km, and mean, median and mode FST values for the pairwise comparisons) are a good predictor of the extent of spread in the introduced range. Simulation studies indicate that OLS regression analyses are preferred if the purpose of the study is not to estimate the parameters of a functional relationship, but instead to simply forecast values of the response variable for given explanatory variables (reviewed in [36]). Genetic structuring of marine taxa is often correlated with geographical distance (IBD). Therefore, in the subset of the studies in which IBD statistics were reported, we tested for a correlation between the IBD slope and extent of spread. To evaluate the influence of alternative variables on invasive success, we used the generalized linear model (GLM) with extent of spread as the response variable, FST as the explanatory variable, and marker type and HE (of native populations) as covariates. Statistical analyses were conducted using SPSS v. 17.0 (IBM, Armonk, New York) except the GLM that was calculated using JMP Pro v. 10.0 (SAS, Cary, North Carolina). Plots of observed versus predicted residuals did not reveal any patterns that would bias the results or interpretation of the regression model (data not shown).
3. Results
(a). Description of the dataset
Searching the literature resulted in 32 cases across 29 species that met our selection criteria: 10 molluscs, four fishes, four crustaceans, three tunicates, two vascular plants and one each of cnidarian, echinoderm, ctenophore, annelid, sponge and an alga (see the electronic supplementary material, table S2). The majority of the studies were based on mitochondrial DNA (21 of 32), including cytochrome oxidase I (n = 16; one of these is a concatenated COI/ND2 dataset), cytochrome b (n = 3), control region (n = 1) and intergenic spacers (n = 1). Nuclear markers were used in eleven studies and included: microsatellites (n = 6), allozymes (n = 2), internal transcribed spacers (n = 2) and amplified fragment length polymorphism (n = 1). Species included in this study are native to regions across the globe including Europe, Asia, America, Australia, South America and Oceania. Introduced regions were also geographically diverse, and included the above continents plus southern Africa. The recording of the green crab, C. maenas, in North America in 1817 makes this the earliest introduction in the dataset, whereas the most recent event involved the detection of the sponge Crambe crambe in the Canary Islands in 1995. The number of native populations surveyed per species ranged from three to 33 (mean = 9.66). Sample sizes per location within the 29 studies ranged from nine to 70 individuals (mean = 25.69) (see the electronic supplementary material, table S2).
Genetic surveys were conducted over a wide range of geographical distances. The shortest distance was 50 km between five populations of the Japanese oyster drill, Ocinebrellus inornatus. The largest native range surveyed was for the bluestripe snapper, L. kasmira, for which 10 populations were sampled across nearly 20 000 km. Global FST values were obtained for 26 of 32 studies and ranged from 0 for the soft-shelled clam M. arenaria to 0.906 for the alga U. pinnatifida. After standardizing for the geographical range over which the genetic survey was conducted, FST/km ranged from 0 to 0.002 with the starlet sea anemone N. vectensism demonstrating the highest value. Pairwise FST values were obtained for 30 of 32 studies. From these pairwise values we calculated mean FST (range = 0.002–0.943), median (range = −0.018 to 0.969) and mode (range = −0.100 to 1.00) (see the electronic supplementary material, table S2). In four studies, although pairwise FST values were available, the number of populations surveyed was low (3 or 4) and precluded the calculation of mode FST. MES varied widely among species ranging from 42 km for the tunicate Pyura praeputialis to 4344 and 4497 km for the ctenophore M. leidyi and the bivalve M. arenaria, respectively. To take secondary introductions into account, we calculated CES for each species which resulted in a narrower range from 8 km for the alga U. pinnatifida to 2583 km for the bluestripe snapper L. kasmira.
(b). Relationship between FST and extent of spread
Weersing & Toonen [37] found that global FST was poorly correlated with geographical study scale in marine organisms, explaining only 2 per cent of the variance among 149 studies. Likewise, we found no correlation between global FST and study scale in our smaller dataset (n = 30, R2 = 0.004, p = 0.754): a finding that may result from the inclusion of different marker types [38]. Similar to other studies, we found that correcting for geographical scale resulted in a slightly higher correlation between FST and the extent of spread (Global FST/km; table 1; [38,39]). Contrary to expectations, we found no correlation between FST and HE.
Table 1.
FST as a predictor of extent of spread of introduced species. Number of studies (n), regression coefficient (R2), and corresponding goodness of fit F-statistic and p-values are reported. R2 values in italics are significant (α = 0.05). Regression equations are reported for significant correlations. MES, maximum extent of spread; and CES, continuous extent of spread.
| all markers |
mtDNA |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| genetic structure | n | R2 | goodness of fit F-statistic | p-value | regression equation | n | R2 | goodness of fit F-statistic | p-value | regression equation |
| MES | ||||||||||
| global FST | 26 | 0.011 | 0.255 | 0.618 | 19 | 0.036 | 0.635 | 0.436 | ||
| global FST/km | 26 | 0.017 | 0.422 | 0.522 | 19 | 0.045 | 0.804 | 0.382 | ||
| mean FST | 30 | 0.001 | 0.026 | 0.873 | 21 | 0.016 | 0.302 | 0.589 | ||
| median FST | 26 | 0.001 | 0.017 | 0.896 | 21 | 0.011 | 0.220 | 0.644 | ||
| mode FST | 26 | 0.001 | 0.032 | 0.859 | 19 | 0.010 | 0.169 | 0.686 | ||
| CES | ||||||||||
| global FST | 26 | 0.245 | 7.801 | 0.010 | Y = −2.62x + 6.68 | 19 | 0.333 | 8.497 | 0.010 | Y = −3.29x + 7.13 |
| global FST/km | 26 | 0.288 | 9.728 | 0.005 | Y = −29.10x + 6.83 | 19 | 0.392 | 10.975 | 0.004 | Y = −37.88x + 7.34 |
| mean FST | 30 | 0.294 | 11.672 | 0.002 | Y = −4.78x + 6.77 | 21 | 0.387 | 12.013 | 0.003 | Y = −5.58x + 7.05 |
| median FST | 26 | 0.394 | 18.201 | <0.001 | Y = −4.85x + 6.69 | 21 | 0.492 | 18.381 | <0.001 | Y = −5.28x + 6.83 |
| mode FST | 26 | 0.464 | 20.814 | <0.001 | Y = −5.25x + 6.47 | 19 | 0.516 | 18.158 | 0.001 | Y = −5.25x + 6.53 |
We found no correlation between genetic structure in the native range and MES. However, when we corrected for secondary introductions and possible human-mediated dispersal within the introduced range, we detected a significant negative correlation between genetic structure and CES. Regardless of the FST value used (global, mean, median or mode), we found significant correlations between population structure in the native range and CES (table 1). Global FST proved to be the weakest predictor of spread (R2 = 0.245, p = 0.010) with a slight improvement in predictive power when global FST was corrected for geographical scale of the study (global FST/km, R2 = 0.288, p = 0.005). The best predictor of CES was the mode of the pairwise FST values (R2 = 0.464, p < 0.001). While not as strongly correlated with spread as mode FST, both mean (R2 = 0.294, p = 0.002) and median (R2 = 0.394, p < 0.001) pairwise FST values were better predictors than global FST.
We did not find a significant effect for marker type, taxon or HE on the extent of spread (table 2). However, previous studies have shown that marker type is a significant covariate when modelling global FST [37–39]. For this reason, we ran regression analyses on the mitochondrial datasets (n = 21) to determine if the correlation between FST and extent of spread improved. In all cases correlations were stronger when analyses were restricted to the mtDNA datasets (table 1 and figure 2). The hierarchy of predictive power between the different measures of FST was consistent between the datasets with global FST, providing the lowest predictive power (R2 = 0.333, p = 0.010) and the mode pairwise FST having the greatest predictive power (R2 = 0.516, p < 0.001).
Table 2.
Results from generalized linear model used to evaluate the influence of alternative variables on invasive success. Extent of spread is the response variable, FST is the explanatory variable and marker type and HE (of native populations) are covariates. p-values in italics are significant (α = 0.05).
| factor | d.f. | χ2 | p-value |
|---|---|---|---|
| CES | |||
| marker type | 1 | 0.075 | 0.784 |
| global FST/km | 1 | 6.626 | 0.010 |
| mean FST | 1 | 7.636 | 0.006 |
| median FST | 1 | 11.607 | <0.001 |
| mode FST | 1 | 16.289 | <0.001 |
| HE | 1 | 0.519 | 0.471 |
Figure 2.
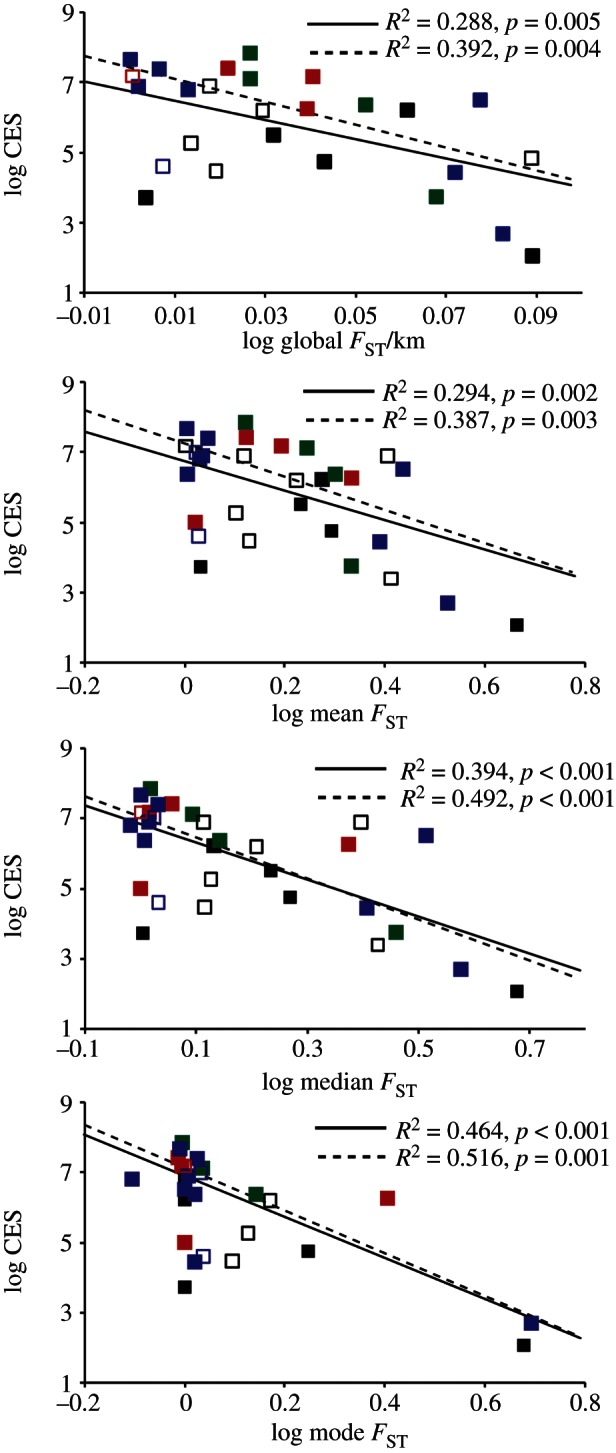
Population genetic structure (FST) versus continuous extent of spread (CES) across nine marine phyla (molluscs, blue; crustaceans, red; fishes, green). Regression lines are plotted for the entire dataset (solid lines) and for just the mtDNA dataset (dashed lines). R2 values and corresponding p-values are shown.
Tests for IBD among native populations were available for only 10 of the 32 studies. We found no significant correlation between IBD slope and MES (R2 = 0.044, p = 0.560). We detected a larger R2 value for the IBD slope versus CES comparison (R2 = 0.190; p = 0.208), however, the power to detect a significant correlation was limited owing to sample size.
4. Discussion
Identifying general characteristics that allow ecologists and managers to predict invaders has proved to be elusive. Here, we offer a metric with considerable power to predict the extent of spread in marine alien species. FST, a measure of alternate fixation of alleles, is a common proxy for realized dispersal among natural populations. Our analysis reveals a strong negative correlation between genetic structure in the native range and the extent of spread of invasive marine species across a diversity of taxa (R2 = 0.245–0.464). An even stronger correlation was detected when marker type was taken into account (R2 = 0.333–0.516). While there are insufficient data to determine if FST can predict the success of initial invasions, our results indicate that this metric is suitable to qualitatively predict the extent of spread once an invasive species is established.
FST is not a simple measure of population differentiation, but instead is influenced by population size and genetic diversity (heterozygosity). There is a predicted relationship between population size, HE and FST [40–42]. Under equilibrium conditions, large effective population sizes are expected to produce high genetic diversity, and low population structure (FST). Consequently, our correlation between FST and invasive spread may indicate that species that sustain large genetically diverse populations make better invaders [43]. Direct population size estimates are not available for most marine species; however, if we assume drift/mutation equilibrium, heterozygosity can be a proxy for population size. We did not find the expected negative correlation between FST and HE in our dataset nor was there a significant relationship between extent of spread and HE, indicating that native population size is not driving the correlations described here.
We restricted our study to marine species because of the strikingly different modes of dispersal between land and sea. Most marine organisms have a biphasic life cycle in which adults are benthic and largely sedentary. Dispersal, over even short distances (1–10 km), is achieved primarily during a pelagic phase (eggs and larvae); the length of which varies greatly among taxa, from 0 days to more than a year in a few taxa (spiny lobsters and eels). As a result, marine larvae can be transported 100's or even 1000's of km before settlement. However, the length of the pelagic larval phase is just one factor effecting dispersal in marine organisms. Timing of spawning, local oceanographic conditions, larval swimming and sensory ability and habitat requirements all play important roles in determining effective dispersal in marine organisms. These myriad of factors probably explain why a consistent correlation between the length of the pelagic phase and FST has not been found except over small geographical scales [37,39]. In the terrestrial realm, perhaps only wind dispersed seeds have a comparable mode of dispersal. Even in fresh water taxa, many of which also have pelagic larvae, dispersal is confined to the lakes and rivers of discrete drainage systems (i.e. zebra mussel in North America) that have no clear analogy in the sea.
(a). FST as a proxy for realized dispersal
Because FST is a summary statistic, significant structure among populations may be a result of differences in effective population size, demographic or colonization history, migration, or some combination of these factors, especially for populations that may not have reached migration drift equilibrium, and thus direct interpretation of population structure in the context of gene flow can sometimes be problematic (reviewed by Hart & Marko [22], Lowe & Allendorf [23] and Marko & Hart [24]). We argue that the use of a summary statistic in this approach is warranted as it represents the cumulative effects of all factors on the genetic structure of the species, and despite the potential confounding variables, we find a significant relationship between FST and CES. Our conclusion, which is somewhat intuitive, is that species that show little or no population structure in their native range (e.g. effective dispersers, habitat or diet generalists, good competitors and broad environmental tolerances) also tend to become the most widespread invaders.
(b). Natural realized dispersal and invasiveness
Our data indicate that up to half of the variation in the extent of spread of marine invasive species can be explained by native population structure as measured by FST. Global FST had the lowest predictive power, explaining 25 per cent of the variation which increased to 39–46% using either median or mode FST. While our dataset was not sufficient to test for the effect of marker type on estimates of genetic structure, measures of FST from mtDNA are generally higher than from nDNA, complicating direct comparisons of FST among marker types ([37–39], but see [44]). Here, we detected higher correlations across estimates of FST (with as much as 52% of the variance explained) when only the mtDNA datasets were considered.
While our findings provide a new measure of invasiveness, caution is indicated in the interpretation of FST values. Our review of the published literature resulted in a moderately sized dataset (29 species) of which the two data points with the highest FST values and correspondingly low CES had a large impact on the relationships revealed here (brown algae, U. pinnatifida; horn snail, Batillaria attramentaria). The former is anchored to substrate and the latter is believed to have a very brief pelagic larval stage. Omitting these two data points results in significant relationship only for median FST (R2 = 0.177, p = 0.026). Examining the data plots (figure 2), it becomes clear that much of the variation in the dataset is at moderate values of FST and CES, indicating that at these values FST has less predictive power. This leads us to conclude that FST can be used to make only qualitative predictions concerning the extent of spread of invasive species [45]. Future studies may provide additional data points to fill in the high end of the FST spectrum and clarify the pattern.
The amount of time that has elapsed since introduction will influence the extent of spread. In our dataset, there is a 12-fold difference in the number of years since the earliest and most recent introductions (the green crab, C. maenas, to North America in 1817 and the sponge C. crambe to the Canary Islands in 1995). However, correcting for this variation is not straightforward. First, the time disparity between first record of occurrence and the actual date of introduction can be vast. Some alien species go undetected for decades, and survey efforts vary considerably across geographical regions [3]. Second, rates of spread vary across taxa and even through time [46]. Therefore, we were not surprised to find no correlation between population structure and invasiveness when we corrected for time since introduction (extent of spread in km per year).
Given the numerous sources of variance in the dataset, the variety of factors that determine invasion success, and the diversity of taxa and systems examined, our finding that FST explains up to 52 per cent of the variance is remarkable. We suspect that the predictive power of FST would increase if analyses could be conducted at the level of individual phyla. However, there is a paucity of population genetic data on alien species. Of the 125 candidate species considered only 23 per cent had genetic data from the native range, and the taxonomic group with the greatest coverage (molluscs, n = 10) was insufficient for a robust analysis.
(c). Continuous versus total extent of spread
The power of FST to predict the geographical spread of alien species is confounded by secondary introductions or human-mediated dispersal within the introduced range [34]. For example, fouling organisms such as tunicates, sponges and oysters can easily be translocated between harbours on boat hulls or fishing gear [47], whereas the larvae of some species can survive in the ballast water of ships [48–50]. The same mechanisms that promote long range introductions can also facilitate spread within the new range. The result is often disjunct distributions in the non-native range that circumvent suitable habitat: a scenario that is less likely with innate (natural) dispersal. Our data indicate that human-mediated secondary introductions are an important means of spread for many alien species.
(d). Terrestrial and freshwater systems
Several reviews have attempted to identify characteristics that predict invader success. However, none have attempted to correlate realized dispersal with invasiveness. Kolar & Lodge [13] review the plant and animal literature, and examine 68 species-level characteristics many of which influence dispersal such as reproductive mode, dispersal mechanisms in plants, fecundity and length of juvenile period. Of those characteristics only reproductive mode in plants was predictive of invasive status (plants with vegetative reproduction were more likely to spread and become abundant). Hayes & Barry [14] examined a larger set of characteristics (115 across seven biological groups) and found that only climate/habitat match was significantly associated with exotic range size across biological groups but not across studies within groups. Here, we add a new quantifiable quality to the array of invasive characteristics.
(e). Conclusions and applications
Using F-statistics to predict the outcome of marine introductions is a novel approach that shows considerable promise. FST as a surrogate for realized dispersal incorporates many of the species-level characteristics that are known to influence invader success: reproductive strategy, habitat specificity and ecology [13]. While our findings show that up to 52 per cent of the variance in the spread of marine invaders can be explained by values of FST, our data do not address the important question of whether a species is likely to become established. Instead, FST would be most useful to wildlife managers when incorporated into specific risk assessment models with success and failure trees for each stage of introduction. In this context, FST could be used to determine which species, once established, are likely to become widespread, providing wildlife officers with a stronger scientific foundation for setting management priorities.
Acknowledgements
This study was supported by University of Hawai‘i Sea Grant no. NA05OAR4171048 (B.W.B.), National Science Foundation (grant no. OIA0554657, OCE-0929031, and OCE-1260169 to R.J.T. and B.W.B.) and NOAA National Marine Sanctuaries Program (MOA grant no. 2005-008/66882 to R.J.T.). We thank the three co-trustees of the Papahānaumokuākea Marine National Monument: NOAA National Marine Sanctuaries, US Fish and Wildlife Service, and the State of Hawai‘i. The views expressed herein are those of the authors and do not necessarily reflect the views of NOAA or any of their sub-agencies. Special thanks to Gail Ashton, Keith Bayha, April Blakeslee, Michael Blum, Sören Bolte, James Carlton, Janie Civille, James Coyer, John Darling, Sharyn Goldstien, Deniz Haydar, Brenden Holland, Shigeru Kojima, Susanna Lopez-Legentil, Angela Mead, Joe Roman, Marc Ruis, Evangelina Schwindt, Benoit Simon-Bouhet, Erik Sotka, Peter Teske, Frédérique Viard and Robert Ward for generously providing datasets and/or constructive comments. We thank editor Gary Carvalho, reviewer Robin Waples, and four anonymous reviewers for comments that greatly improved the manuscript. This is contribution no. 1549 from the Hawai‘i Institute of Marine Biology, no. 8916 from the School of Ocean and Earth Science and Technology, and UNIHI-SEAGRANT-JC-10-35 from the University of Hawai‘i Sea Grant Program.
References
- 1.Carlton JT, Geller JB. 1993. Ecological roulette: the global transport of nonindigenous marine organisms. Science 261, 78–82 10.1126/science.261.5117.78 (doi:10.1126/science.261.5117.78) [DOI] [PubMed] [Google Scholar]
- 2.Cowie RH. 1998. Patterns of introduction of non-indigenous non-marine snails and slugs in the Hawaiian Islands. Biodiv. Conserv. 7, 349–368 10.1023/A:1008881712635 (doi:10.1023/A:1008881712635) [DOI] [Google Scholar]
- 3.Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz FA. 2000. Biotic invasions: causes, epidemiology, global consequences, and control. Ecol. Appl. 10, 689–710 10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2 (doi:10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2) [DOI] [Google Scholar]
- 4.Pimentel D, Lach L, Zuniga R, Morrison D. 2000. Environmental and economic costs of nonindigenous species in the United States. BioScience 50, 53–65 10.1641/0006-3568(2000)050[0053:EAECON]2.3.CO;2 (doi:10.1641/0006-3568(2000)050[0053:EAECON]2.3.CO;2) [DOI] [Google Scholar]
- 5.Pimentel D, Zuniga R, Morrison D. 2005. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 52, 273–288 10.1016/j.ecolecon.2004.10.002 (doi:10.1016/j.ecolecon.2004.10.002) [DOI] [Google Scholar]
- 6.Tatem AJ, Hay SI, Rogers DJ. 2006. Global traffic and disease vector dispersal. Proc. Natl Acad. Sci. USA 103, 6242–6247 10.1073/pnas.0508391103 (doi:10.1073/pnas.0508391103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Willamson M, Fitter A. 1996. The varying success of invaders. Ecology 77, 1661–666 10.2307/2265769 (doi:10.2307/2265769) [DOI] [Google Scholar]
- 8.Cox GW. 1999. Alien species in North America and Hawaii: impacts on natural ecosystems. Washington, DC: Island Press [Google Scholar]
- 9.Simberloff D. 2005. Non-native species do threaten the natural environment. J. Agri. Environ. Ethics 18, 595–607 10.1007/s10806-005-2851-0 (doi:10.1007/s10806-005-2851-0) [DOI] [Google Scholar]
- 10.Wicove DS, Rothstein D, Dubow J, Phillips A, Losos E. 1998. Quantifying threats to imperiled species in the United States. BioScience 48, 607–615 10.2307/1313420 (doi:10.2307/1313420) [DOI] [Google Scholar]
- 11.Armstrong S. 1995. Rare plants protect Cape's water supplies . New Sci. 11, 8 [Google Scholar]
- 12.Elton CS. 1958. The ecology of invasions by animals and plants. London, UK: Methuen [Google Scholar]
- 13.Kolar CS, Lodge DM. 2001. Progress in invasion biology predicting invaders. Trends Ecol. Evol. 16, 199–204 10.1016/S0169-5347(01)02101-2 (doi:10.1016/S0169-5347(01)02101-2) [DOI] [PubMed] [Google Scholar]
- 14.Hayes KR, Barry SC. 2008. Are there any consistent predictors of invasion success? Biol. Invasions 10, 483–506 10.1007/s10530-007-9146-5 (doi:10.1007/s10530-007-9146-5) [DOI] [Google Scholar]
- 15.Rejmanek M, Richardson DM. 1996. What attribute make some plant species more invasive? Ecology 77, 1655–1661 10.2307/2265768 (doi:10.2307/2265768) [DOI] [Google Scholar]
- 16.Reichard SH, Hamilton CW. 1997. Predicting invasions of woody plants introduced into North America. Conserv. Biol. 11, 193–203 10.1046/j.1523-1739.1997.95473.x (doi:10.1046/j.1523-1739.1997.95473.x) [DOI] [Google Scholar]
- 17.Roman J, Darling J. 2007. Paradox lost: genetic variation and the success of aquatic invasions. Trends Ecol. Evol. 22, 454–464 10.1016/j.tree.2007.07.002 (doi:10.1016/j.tree.2007.07.002) [DOI] [PubMed] [Google Scholar]
- 18.Simberloff D. 2009. The role of propagule pressure in biological invasions. Annu. Rev. Ecol. Evol. Syst. 40, 81–102 10.1146/annurev.ecolsys.110308.120304 (doi:10.1146/annurev.ecolsys.110308.120304) [DOI] [Google Scholar]
- 19.Holland BS. 2000. Genetics of marine bioinvasions. Hydrobiologia 420, 63–71 10.1023/A:1003929519809 (doi:10.1023/A:1003929519809) [DOI] [Google Scholar]
- 20.Holsinger KE, Weir BS. 2009. Genetics in geographically structured populations: defining, estimating, and interpreting FST. Nat. Rev. Genet. 10, 639–650 10.1038/nrg2611 (doi:10.1038/nrg2611) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bird CE, Karl SA, Smouse P, Toonen RJ. 2011. Detecting and measuring genetic differentiation. In Crustacean issues: phylogeography and population genetics in crustacea (eds Koenemann S, Held C, Schubart C.), pp. 31–55 Boca Raton, FL: CRC Press [Google Scholar]
- 22.Hart MW, Marko PB. 2010. It's about time: divergence, demography, and the evolution of developmental modes in marine invertebrates. Int. Comp. Biol. 50, 643–661 10.1093/icb/icq068 (doi:10.1093/icb/icq068) [DOI] [PubMed] [Google Scholar]
- 23.Lowe WH, Allendorf FW. 2010. What can genetics tell us about population connectivity? Mol. Ecol. 19, 3038–3051 10.1111/j.1365-294X.2010.04688.x (doi:10.1111/j.1365-294X.2010.04688.x) [DOI] [PubMed] [Google Scholar]
- 24.Marko PB, Hart MW. 2011. Retrospective coalescent methods and the reconstruction of metapopulation histories in the sea. Evol. Ecol. 26, 291–315 10.1007/s10682-011-9467-9 (doi:10.1007/s10682-011-9467-9) [DOI] [Google Scholar]
- 25.Waples R. 1998. Separating the wheat from the chaff: patterns of genetic differentiation in high gene flow species. J. Heredity 89, 438–450 10.1093/jhered/89.5.438 (doi:10.1093/jhered/89.5.438) [DOI] [Google Scholar]
- 26.Oda DK, Parrish JD. 1982. Ecology of commercial snappers and groupers introduced to Hawaiian reefs. In Proc. 4th Int. Coral Reef Symp., pp. 59–67, Manila [Google Scholar]
- 27.Randall JE. 2007. Reef and Shore Fishes of the Hawaiian Islands. Honolulu, HI: Sea Grant College Program, University of Hawaii [Google Scholar]
- 28.Gaither MR, Toonen RJ, Bowen BW. 2012. Coming out of the starting blocks: extended lag time rearranges genetic diversity in introduced marine fishes of Hawai‘i. Proc. R. Soc. B 279, 3948–3957 10.1098/rspb.2012.1481 (doi:10.1098/rspb.2012.1481) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gaither MR, Bowen BW, Toonen RJ, Planes S, Messmer V, Earle J, Robertson DR. 2010. Genetic consequences of introducing two allopatric lineages of Bluestripe Snapper (Lutjanus kasmira) to Hawaii. Mol. Ecol. 19, 1107–1121 10.1111/j.1365-294X.2010.04535.x (doi:10.1111/j.1365-294X.2010.04535.x) [DOI] [PubMed] [Google Scholar]
- 30.Gaither MR, Toonen RJ, Robertson DR, Planes S, Bowen BW. 2010. Genetic evaluation of marine biogeographic barriers: perspectives from two widespread Indo-Pacific snappers (Lutjanus kasmira and Lutjanus fulvus). J. Biogeogr. 37, 133–147 10.1111/j.1365-2699.2009.02188.x (doi:10.1111/j.1365-2699.2009.02188.x) [DOI] [Google Scholar]
- 31.Gaither MR, Bowen BW, Bordenave T, Rocha LA, Newman SJ, Gomez JA, van Herwerden L, Craig MT. 2011. Phylogeography of the reef fish Cephalopholis argus (Epinephelidae) indicates Pleistocene isolation across the Indo-Pacific Barrier with contemporary overlap in the Coral Triangle. BMC Evol. Biol. 11, 189. 10.1186/1471-2148-11-189 (doi:10.1186/1471-2148-11-189) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Weir BS, Cockerham CC. 1984. Estimating F-statistics for the analysis of population structure. Evolution 38, 1358–1370 10.2307/2408641 (doi:10.2307/2408641) [DOI] [PubMed] [Google Scholar]
- 33.Excoffier L, Lischer HEL. 2010. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Res. 10, 564–567 10.1111/j.1755-0998.2010.02847.x (doi:10.1111/j.1755-0998.2010.02847.x) [DOI] [PubMed] [Google Scholar]
- 34.Darling JA, Bagley MJ, Roman J, Tepolt CK, Geller JB. 2008. Genetic patterns across multiple introductions of the globally invasive crab genus Carcinus. Mol. Ecol. 17, 4992–5007 10.1111/j.1365-294X.2008.03978.x (doi:10.1111/j.1365-294X.2008.03978.x) [DOI] [PubMed] [Google Scholar]
- 35.Roman J, Palumbi SR. 2004. A global invader at home: population structure of the green crab, Carcinus maenas, in Europe. Mol. Ecol. 13, 2891–2898 (doi:10.1111/j.1365–294X.2004.02255.x) [DOI] [PubMed] [Google Scholar]
- 36.Legendre P, Legendre L. 1998. Numerical Ecology. Amsterdam, The Netherlands: Elsevier Science BV [Google Scholar]
- 37.Weersing K, Toonen RJ. 2009. Population genetics, larval dispersal, and connectivity in marine systems. Mar. Ecol. Prog. Ser. 393, 1–12 10.3354/meps08287 (doi:10.3354/meps08287) [DOI] [Google Scholar]
- 38.Riginos C, Douglas KE, Jin Y, Shanahan DF, Treml EA. 2011. Effects of geography and life history traits on genetic differentiation in benthic marine fishes. Ecography 34, 566–575 10.1111/j.1600-0587.2010.06511.x (doi:10.1111/j.1600-0587.2010.06511.x) [DOI] [Google Scholar]
- 39.Selkoe KA, Toonen RJ. 2011. Marine connectivity: a new look at pelagic larval duration and genetic metrics of dispersal. Mar. Ecol. Prog. Ser. 436, 291–305 10.3354/meps09238 (doi:10.3354/meps09238) [DOI] [Google Scholar]
- 40.Slatkin M. 1985. Gene flow in natural populations. Annu. Rev. Ecol. Syst. 16, 393–430 10.1146/annurev.ecolsys.16.1.393 (doi:10.1146/annurev.ecolsys.16.1.393) [DOI] [Google Scholar]
- 41.Frankham R. 1996. Relationship of genetic variation to population size in wildlife. Conserv. Biol. 10, 1500–1508 10.1046/j.1523-1739.1996.10061500.x (doi:10.1046/j.1523-1739.1996.10061500.x) [DOI] [Google Scholar]
- 42.Neigel JE. 2002. Is FST obsolete? Conserv. Genet. 3, 167–173 10.1023/A:1015213626922 (doi:10.1023/A:1015213626922) [DOI] [Google Scholar]
- 43.Wares JP, Hughes AR, Grosberg RK. 2005. Mechanism that drive evolutionary change. In Species invasions: insights into ecology, evolution, and biogeography (eds Sax FS, Stachowicz JJ, Gaines SD.), pp. 229–257 Sunderland, UK: Sinauer Associates, Inc [Google Scholar]
- 44.DiBattista JD, Craig MT, Rocha LA, Feldheim KA, Bowen BW. 2012. Phylogeography of the Indo-Pacific butterflyfishes, Chaetodon meyeri and Chaetodon ornatissimus: sister species reveal divergent evolutionary histories and discordant results from mtDNA and microsatellites. J. Heredity 103, 617–629 10.1093/jhered/ess056 (doi:10.1093/jhered/ess056) [DOI] [PubMed] [Google Scholar]
- 45.Fauby S, Barber PH. 2012. Theoretical limits to the correlation between pelagic larval duration and population genetic structure. Mol. Ecol. 21, 3419–3432 10.1111/j.1365-294X.2012.05609.x (doi:10.1111/j.1365-294X.2012.05609.x) [DOI] [PubMed] [Google Scholar]
- 46.Grosholz ED. 1996. Contrasting rates of spread for introduced species in terrestrial and marine systems. Ecology 77, 1680–1686 10.2307/2265773 (doi:10.2307/2265773) [DOI] [Google Scholar]
- 47.Bax N, Williamson A, Aguero M, Gonzalez E, Geeves W. 2003. Marine invasive alien species: a threat to global biodiversity. Mar. Policy 27, 313–323 10.1016/S0308-597X(03)00041-1 (doi:10.1016/S0308-597X(03)00041-1) [DOI] [Google Scholar]
- 48.Medcof JC. 1975. Living marine animals in a ship's ballast water. Proc. Natl. Shellfish Assoc. 65, 54–55 [Google Scholar]
- 49.Carlton JT. 1985. Transoceanic and interoceanic dispersal of coastal marine organisms: the biology of ballast water. Oceanogr. Mar. Biol. Annu. Rev. 23, 313–371 [Google Scholar]
- 50.Gollasch S, Lenz J, Dammer M, Andres H-G. 2000. Survival of tropical ballast water organisms during a cruise from the Indian Ocean to the North Sea. J. Plankton. Res. 22, 923–937 10.1093/plankt/22.5.923 (doi:10.1093/plankt/22.5.923) [DOI] [Google Scholar]