

Abstract
Orthographic differences across languages impose differential weighting on distinct component processes, and consequently on different pathways during word-reading tasks. Readers of transparent orthographies such as Italian and Hindi are thought to rely on spelling-to-sound assembly and show increased activation in phonologically-tuned areas along the dorsal pathway, whereas reading an opaque orthography such as English is thought to rely more on lexically-mediated processing associated with increased activation of semantically-tuned regions along the ventral pathway. To test if biliterate Hindi/English readers exhibit orthography-specific reading pathways, we used behavioural measures and functional neuroimaging. Reaction times and activation patterns of monolingual English and Hindi readers were compared to two groups of adult biliterates; 14 simultaneous readers who learnt to read both languages at age 5 and 10 sequential readers who learnt Hindi at 5 and English at 10.
Simultaneous, but not sequential readers demonstrated relative activation differences of dorsal and ventral areas in the two languages. Similar to native counterparts, simultaneous readers preferentially activated the left inferior temporal gyrus for English and left inferior parietal lobule (L-IPL) for Hindi, whereas, sequential readers showed higher activation along the L-IPL for reading both languages. We suggest that early simultaneous exposure to reading distinct orthographies results in orthography-specific plasticity that persists through adulthood.
Keywords: biliteracy, reading, simultaneous, sequential, orthography
Introduction
Language and reading are powerful cognitive abilities, uniquely human. Whereas spoken language is biologically specialized and naturally acquired, reading is a cultural byproduct of the former that requires extensive instruction to master. Despite the fact that writing systems vary in how visual symbols represent units of spoken language, principles governing written representations of languages are largely universal (Perfetti, 2003). With regard to the neural basis of reading, a meta-analysis of studies on word reading across various languages has supported the existence of a universal reading network consisting of the left superior temporal gyrus (L-STG), the left inferior frontal gyrus (L-IFG), the left occipitotemporal region, and the mid-fusiform gyrus (Bolger et al., 2005).
Despite general overlap of word reading networks, neuroimaging studies have also reported cortical differences across languages due to orthographic depth (Paulesu et al., 2000). Orthographic depth is the transparency of spelling to sound mapping, and writing systems of the world are classified on a multivalent to univalent continuum (Ziegler and Goswami, 2005). Reading in orthographies with multivalent spelling-to-sound mappings (hereafter referred to as opaque orthographies) such as English is believed to rely, to a large extent, on previously acquired word patterns to resolve ambiguities in pronunciation, and readers preferentially activate the L-IFG and left inferior temporal gyrus (L-ITG) in the ventral pathway (Paulesu et al., 2000). Multivalent spelling-to-sound mapping refers to letter/letter string mappings to distinct sounds in different words, e.g., cough and bough. It also refers to “many-to-many mappings” (e.g., “ch” to /t / as in “church”), “many-to-one mappings” (e.g., “sh” to / / as in “sheep”), “one-to-many mappings” (e.g., “x” to /ks/ as in “Max”), and “one-to-one” (e.g., “c” to /k/ in “cat”). In contrast, univalent orthography means unique letter-to-sound mapping that is one-to-one. Hindi has a unique orthography, wherein, one letter
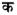 maps to one sound, for example, letter maps to sound /k/ only. Reading in orthographies with largely univalent spelling-to-sound mappings (hereafter referred to as transparent orthographies) such as Italian and Spanish, is thought to rely more exclusively on grapheme to phoneme mapping (given the stability of this process in yielding the correct pronunciation) and showed increased activation in L-STG in the dorsal pathway (Paulesu et al., 2000, Meschyan and Hernandez, 2006). These studies suggest that transparent orthographies appear to elicit greater activation in dorsal areas of the left hemisphere (L-STG) whereas opaque orthographies preferentially activate ventrally-tuned areas (L-ITG) while reading words. Comparatively little is known about the cortical reading networks employed by readers of two languages belonging to two different orthographies (hereafter also called biliterates) and whether they exhibit orthography-specific weighting of reading pathways. Research on second language reading is mixed. Although some studies point towards a largely universal neurocircuitry for reading, showing little difference between activation patterns for bilinguals reading their first language (L1) and second language (L2) (Chee et al., 1999; Crinion et al., 2006; Klein at al., 1999), other studies report differences for L2 reading, such as increased activation of language prefrontal and cerebellar regions (Klein et al., 2006; Marian et al., 2003; Meschyan and Hernandez, 2006; Xue et al., 2004). These differences in L2 reading have been attributed largely to reduced proficiency in reading the second language. For example, Tan and colleagues (2003) compared activation patterns for Chinese-English late bilinguals (Tan et al., 2003) with English monolinguals. They showed that for reading English, the monolinguals had increased participation of L-IFG and L-STG, while the bilinguals showed strong activation in left middle frontal gyrus (L-MFG) for reading both English and Chinese. L-MFG is strongly activated while reading Chinese (Perfetti et al., 2007) and these findings suggested that the late bilinguals, who were less capable of processing English, were applying the L1 (Chinese) reading circuitry while reading L2 (English) (Tan et al., 2003). In this study, the bilingual participants had acquired the second language late (> 12years) and were less proficient at reading English as compared to Chinese. This suggests that cortical specificity for reading different orthographies is likely to be influenced by reading proficiency and age of acquisition: reading is understudied to date in this regard. Since the late sequential bilinguals in the study by Tan et al. (2003) did not demonstrate language-specific activation patterns in the second language but used the L1 reading network to read L2, in this study we ask – do proficient simultaneous bilinguals, who learnt to read both L1 and L2 early in life, recruit orthography specific reading networks and not merely follow the L1 network for reading L2?
maps to one sound, for example, letter maps to sound /k/ only. Reading in orthographies with largely univalent spelling-to-sound mappings (hereafter referred to as transparent orthographies) such as Italian and Spanish, is thought to rely more exclusively on grapheme to phoneme mapping (given the stability of this process in yielding the correct pronunciation) and showed increased activation in L-STG in the dorsal pathway (Paulesu et al., 2000, Meschyan and Hernandez, 2006). These studies suggest that transparent orthographies appear to elicit greater activation in dorsal areas of the left hemisphere (L-STG) whereas opaque orthographies preferentially activate ventrally-tuned areas (L-ITG) while reading words. Comparatively little is known about the cortical reading networks employed by readers of two languages belonging to two different orthographies (hereafter also called biliterates) and whether they exhibit orthography-specific weighting of reading pathways. Research on second language reading is mixed. Although some studies point towards a largely universal neurocircuitry for reading, showing little difference between activation patterns for bilinguals reading their first language (L1) and second language (L2) (Chee et al., 1999; Crinion et al., 2006; Klein at al., 1999), other studies report differences for L2 reading, such as increased activation of language prefrontal and cerebellar regions (Klein et al., 2006; Marian et al., 2003; Meschyan and Hernandez, 2006; Xue et al., 2004). These differences in L2 reading have been attributed largely to reduced proficiency in reading the second language. For example, Tan and colleagues (2003) compared activation patterns for Chinese-English late bilinguals (Tan et al., 2003) with English monolinguals. They showed that for reading English, the monolinguals had increased participation of L-IFG and L-STG, while the bilinguals showed strong activation in left middle frontal gyrus (L-MFG) for reading both English and Chinese. L-MFG is strongly activated while reading Chinese (Perfetti et al., 2007) and these findings suggested that the late bilinguals, who were less capable of processing English, were applying the L1 (Chinese) reading circuitry while reading L2 (English) (Tan et al., 2003). In this study, the bilingual participants had acquired the second language late (> 12years) and were less proficient at reading English as compared to Chinese. This suggests that cortical specificity for reading different orthographies is likely to be influenced by reading proficiency and age of acquisition: reading is understudied to date in this regard. Since the late sequential bilinguals in the study by Tan et al. (2003) did not demonstrate language-specific activation patterns in the second language but used the L1 reading network to read L2, in this study we ask – do proficient simultaneous bilinguals, who learnt to read both L1 and L2 early in life, recruit orthography specific reading networks and not merely follow the L1 network for reading L2?
We focus on biliterate readers and study two groups relative to monolingual control groups; one comprising bilingual simultaneous readers, who acquired reading in English and Hindi at age 5 and a second comprised of bilingual sequential readers who acquired reading Hindi at age 5 but English at 10. Apart from Italian, Hindi is another example of a transparent orthography, written in Devanagari script (Bright, 2000). Unlike Italian, which is alphabetic wherein vowels and consonants are arranged linearly, in Hindi vowel markers appear not only in a linear manner but also on top, below or on the sides of consonants*1 (Kumar et al., 2009). Despite gross differences, both Italian and Hindi are highly regular*2 and hence could be thought to rely on orthography to phonology mapping to a similar extent. We tested the following hypotheses: 1) Hindi as a transparent orthography should have relatively greater weighting of dorsal (phonologically tuned) sites, whereas English should have relatively greater weighting of ventral (lexical semantic) sites and 2) the degree language specificity of weighting in both Hindi and English should be greater for those who were early proficient readers (simultaneous readers). Since monolingual English readers recruit areas such as L-ITG, along the ventral pathway, we postulate that early simultaneous bilingual readers would preferentially recruit L-ITG, for reading English, an opaque orthography. For Hindi, a transparent orthography they would recruit sites along the dorsal route. Based on our hypotheses, we predict that, only for early simultaneous proficient readers, reading in a language is more “native-like” with regard to code-utilization.
Materials and Methods
Participants
Forty participants matched on age, handedness and socio-economic status were selected on the basis of a language questionnaire (Table 1) and reading fluency tasks. In the questionnaire, participants answered questions regarding place of residence for longest period, first language, second language and their ages of acquisition, handedness, years of education in Hindi and/or English and hours spent reading one/both languages. In self-assessment, the participants rated themselves on a scale of 0–10 on their conversation, reading and writing skills. Handedness was assessed by the Edinburgh Inventory (Oldfield, 1971). The participants consisted of 14 simultaneous and 10 sequential Hindi–English bilingual readers, 9 monolingual Hindi and 7 monolingual English readers. The bilingual readers were recruited from various universities. Monolingual Hindi readers were recruited from a Hindi university whereas monolingual English readers were students visiting India under an exchange program. The simultaneous readers had longer formal training in English than the sequential readers (t21=4.3, p=0.001). They also read English more than the sequential readers (Mann Whitney rank sum test, p=0.002) (details in Table 1).
Table 1.
Performance in reading tasks in monolingual readers, simultaneous and sequential readers in English and Hindi.
| English readers (n=7) | Hindi readers (n=9) | Simultaneous readers (n=14) | Sequential readers (n=10) | ||||
|---|---|---|---|---|---|---|---|
| English | Hindi | English | Hindi | ||||
| Self assessment (±s.d.) | 9.23±1.35 | 9.09±0.53 | 8.35±0.76 | 7.94±1.31 | 8.60±1.01 | 9.46±0.71 | |
| Years of formal education | 19.1±1.4 | 13.7±2.1 | 21.9±3.5 | 12.5±1.7 | 16.4±1.5 | 16.0±6.4 | |
| Reading habit (hrs spent/day) | 7.4±5.6 | 5.6±3.1 | 6.6±3.5 | 1.1±0.5 | 2.3±2.1 | 1.7±1.0 | |
No significant difference between self-assessment for English and Hindi in early simultaneous bilingual readers is seen. The late sequential bilingual readers show significant difference between English and Hindi self assessment (paired t9 = −2.899, p=0.018). The sequential bilingual readers differ from simultaneous bilingual readers in years of formal education in English (two sample t-test, t21=4.3, p=0.001) and hours spent in reading English per day (Mann Whitney rank sum test, p=0.002). The sequential readers also differ from English monolingual readers in daily hours spent in English reading (Mann Whitney rank sum test, p=0.015). The simultaneous readers however did not differ from English monolingual readers in this aspect.
Behaviour
Passage reading time
Reading fluency for all groups was measured using passage reading time. A number of studies (e.g. Haller et al., 2005) have established time taken to read a novel passage as a measure of reading fluency. Time taken to read aloud a passage in English and/or Hindi was measured for all participants. The simultaneous and sequential readers read passages in both languages. The passages were matched for number of words (230 words in each passage). Reading time was measured by pressing a button upon beginning to read and again upon conclusion. The monolingual readers read either the English or the Hindi passage.
Stimuli
Word stimuli comprised 420 abstract and concrete words, matched for number of phonemes, 210 from each language. Of the 210 words, 105 were low frequency (e.g., fetter, veer;
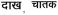 , name of a fruit and a bird respectively) and 105 were high frequency words (e.g., table, water,
, name of a fruit and a bird respectively) and 105 were high frequency words (e.g., table, water,
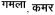 , meaning pot and waist respectively). Regular words were chosen in English and only linear words were used in Hindi (in order to avoid confounding effects of nonlinearity), as shown in the examples above. The choice of regular words in English made it more transparent-like and matched to Hindi orthography. However, the inherent opaqueness of English still remained, for example, the ‘oa’ in ‘toad’ was pronounced as ‘o’ and not /o//a/ separately. It is important to note that even a high frequency word like ‘toad’ requires retrieval of pronunciation of the bigram ‘oa’ from memory. Hindi however only requires knowledge of letters and their corresponding sounds, which remain unchanged. Our paradigm banked upon this difference between the English and Hindi orthographies and we expected to find differences in orthography-specific reading strategies depending upon the levels of proficiency of participant groups across the two orthographies.
, meaning pot and waist respectively). Regular words were chosen in English and only linear words were used in Hindi (in order to avoid confounding effects of nonlinearity), as shown in the examples above. The choice of regular words in English made it more transparent-like and matched to Hindi orthography. However, the inherent opaqueness of English still remained, for example, the ‘oa’ in ‘toad’ was pronounced as ‘o’ and not /o//a/ separately. It is important to note that even a high frequency word like ‘toad’ requires retrieval of pronunciation of the bigram ‘oa’ from memory. Hindi however only requires knowledge of letters and their corresponding sounds, which remain unchanged. Our paradigm banked upon this difference between the English and Hindi orthographies and we expected to find differences in orthography-specific reading strategies depending upon the levels of proficiency of participant groups across the two orthographies.
Of the 420 words used in the behavioural task, 240 words that were concrete imageable nouns were selected for the functional magnetic resonance imaging (fMRI) task (representative words given in Appendix I). These comprised 120 nouns each in English and Hindi. Of these, 60 were high frequency and 60 low frequency nouns as validated by a linguist and rated by ten proficient bilingual Hindi-English readers who read both Hindi and English regularly. The words were rated for high and low frequency on a five-point scale. Words rated above 4 were taken as high frequency and those rated below 2 were used as low frequency words.
Vocal reaction time
Following imaging, the participants performed reaction time tasks outside the scanner. They were instructed to read aloud all 420 words presented on a computer monitor subtending a visual angle of 4°. Participants were recorded while reading the words and time taken to respond to the words (210 each in Hindi and English, total 420) were measured as vocal reaction time. The monolinguals read words (210) in their native language only (English or Hindi) whereas the biliterates read words (420) in both languages. Naming errors were monitored and only correct responses were analysed. Mean vocal reaction times were estimated in both languages.
fMRI
Participants were asked to fixate on a central fixation cross during baseline blocks and read aloud words subtending an angle of 4° during stimuli blocks. Two runs each were administered for English and Hindi. Each run consisted of words in one language only. The monolinguals were administered two runs (in either English or Hindi, depending on whichever was their native language) and biliterates were administered four runs (two in each language).
Each language run was 240 seconds long and consisted of a total of 12 blocks wherein six word-reading blocks (three high and three low frequency blocks) were interspersed with six fixation blocks (Fig. 1). Each run began with a fixation block. Each block was 20 seconds long. Each word reading block consisted of 10 words presented at an interval of 2 seconds each. Thus participants read aloud a total of 30 high frequency and 30 low frequency concrete nouns in each language run. They were recorded using an fMRI-compatible microphone (Optoacoustics FOMRI II dual channel MRI microphone system, version 1.1). The study was approved by the Human Ethics Committee of the National Brain Research Centre.
Figure 1.

Schematic of the paradigm design. Each language run was administered separately; two runs each for English and Hindi. A run was 240 seconds long and consisted of three blocks of high frequency (HF) and three blocks of low frequency (LF) words in a language, interspersed with blocks of baseline fixation. Each block was 20 seconds long and ten words were presented in a block, one every 2 seconds. Task was to read aloud.
Data acquisition and analysis
The participants were scanned in a 3T Philips Achieva scanner. Gradient echo, echo-planar images (TR = 2 sec, TE = 35 msec, flip angle = 90°, FOV = 230 mm, matrix = 64 × 64, slice thickness = 4 mm, gap = 1 mm, 30 axial slices) and a three-dimensional T1 weighted image for anatomical normalization (TR = 8.4 msec, TE = 3.7 msec, flip angle = 8°, FOV = 250 × 230 mm, matrix = 256 × 256) was used. A total of 240 volumes for each language were acquired for each subject and analysed using SPM5 (Friston et al., 1995a). The anatomical image for each participant was co-registered with the T1 weighted image and normalized to the EPI (echo planar imaging) template in the MNI (Montreal Neurological Institute) space. The normalized fMRI images were smoothed with an 8mm FWHM (full width half maximum) Gaussian filter and condition related response was modeled by a boxcar convolved with a canonical hemodynamic response function (Friston et al., 1995b). High pass temporal filtering with 170s was applied. Motion parameters defined by the realignment procedures were added to the model. Statistical analysis was performed using a two-level approach (Holmes and Friston, 1998).
At the first level, contrast images were calculated for the condition of interest for each subject. At the second level, a one-sample t-test was performed using all the contrast images of the subjects to obtain a generalized effect of the population of interest with respect to baseline (for example, low frequency > baseline). The following types of analyses were carried out:
A factorial analysis based on a 2 (orthography: English, Hindi) × 2 (frequency: high, low) × 2 (biliterate type: simultaneous, sequential readers) design was carried out.
-
The effects of frequency and orthography were investigated in detail across groups.
Effects of orthography – Orthography contrasts for reading all words in English and Hindi were obtained. A two sample t-test across the two data acquired while reading each of the orthographies was carried out.
Effects of frequency - Frequency contrasts for reading low and high frequency words in both languages were obtained. Paired t-tests were performed on each population reading high and low frequency words. For example, pairs of high frequency and low frequency contrast images were taken for each simultaneous reader while reading English. Following this, activations specific for high>low frequency word reading and low>high frequency word reading in English were calculated for the simultaneous reader group. This process was repeated for sequential and monolingual groups while reading English and Hindi.
Effects of proficiency - Since the biliterate readers were not matched in proficiency, we further investigated effects of proficiency across the simultaneous and sequential readers. A two-sample t-test across populations was performed by taking contrast images for both populations for the condition of interest. For example, contrast images for simultaneous readers reading low frequency English words were contrasted with those of sequential readers reading low frequency English words.
In all the analyses described above, regions of significant activations were identified using a threshold cluster of p<0.001 (uncorrected).
Results
Based on the information provided by the participants in the language questionnaire, the bilingual readers were categorized into simultaneous and sequential bilinguals. Assessments of reading fluency and performance in the behavioral task are described below.
Behaviour
Passage reading time
Fig. 2a shows the results of time taken to read English and Hindi passages by monolingual and bilingual readers. Sequential bilingual readers differed from simultaneous readers (Mann Whitney test, p=0.014) and English monolinguals (t-test, t13 = 2.536, p=0.025) for reading the English passage Total time to read the passage did not differ for simultaneous readers and English monolinguals. There was no difference across the bilingual groups and the Hindi monolinguals for reading the Hindi passage.
Figure 2.


Figure 2(a). Passage reading time for English and Hindi. The plot shows average passage reading times for English (average±stdev; 106.06±13.84) and Hindi (115.16±10.21) monolingual readers, early simultaneous bilingual readers reading in English (111.08±12.79) and Hindi (115.90±10.21) and late sequential bilingual readers reading in English (129.06±27.8) and Hindi (96.96±15.05). A significant difference is observed between simultaneous and sequential readers in the time taken to read English passage (Mann Whitney test, p=0.014). The sequential readers also differed when compared to English monolingual readers in time taken to read the passage (two sample t-test, t13=2.536, p=0.025).
Figure 2(b). Reaction time for reading English and Hindi words. Top panel shows reaction time to read English high and low frequency words in English readers (high: 0.65±0.02 sec; low: 0.68±0.03 sec), simultaneous (high: 0.71±0.04 sec; low: 0.73±0.05 sec) and sequential readers (high: 0.75±0.06 sec; low: 0.81±0.07 sec). Sequential readers differ from monolingual English readers while reading low frequency words (t-test, t9=− 3.040, p<0.014) and simultaneous readers (t-test, t9=−2.87, p<0.018). The bottom panel shows reaction times for reading high and low frequency Hindi words by monolingual Hindi readers (high: 0.64±0.08 sec; low: 0.66±0.09 sec), simultaneous (high: 0.71±0.05 sec; low: 0.72±0.06 sec) and sequential readers (high: 0.74±0.04 sec; low: 0.76±0.05 sec). No between group differences for reading either high or low frequency Hindi words are observed. The error bars depict the standard error of mean.
Reaction time
The reaction time data (Fig. 2b) showed that all the participants responded faster to high frequency words than to low frequency words (Table 2). No significant differences were seen across groups for reading high frequency words in English. However for low frequency English words, significant differences were seen across groups. The sequential readers took significantly longer to respond to low frequency English words when compared with monolingual English readers (t-test, t9 = −3.04, p<0.014) and simultaneous readers (t-test, t9 =−2.87, p<0.018). However, no differences were obtained when the simultaneous readers were compared to the English readers for reading low frequency words. Response times to Hindi words did not differ significantly across groups (Table 2).
Table 2.
Reaction times and accuracy for English and Hindi words.
| English | Hindi | |||||||
|---|---|---|---|---|---|---|---|---|
| High | Low | High | Low | |||||
| Reaction time (seconds) | Accuracy (%) | Reaction time (seconds) | Accuracy (%) | Reaction time (seconds) | Accuracy (%) | Reaction time (seconds) | Accuracy (%) | |
| Monolingual English readers | 0.65±0.02 | 99 | 0.68±0.03 | 99 | ||||
| Monolingual Hindi readers | 0.64±0.08 | 100 | 0.66±0.09 | 100 | ||||
| Simultaneous bilingual readers | 0.71±0.04 | 98 | 0.73±0.05 | 98 | 0.71±0.05 | 99 | 0.72±0.06 | 99 |
| Sequential bilingual readers | 0.75±0.06 | 96 | *0.81±0.07 | 89 | 0.74±0.04 | 100 | 0.76±0.05 | 100 |
Significant differences seen when sequential readers were compared with monolingual English readers, (t9=−3.040, p<0.014) and simultaneous bilingual readers (t9=−2.87, p<0.018).
fMRI
Factorial analysis (Fig. 3)
Figure 3.

Factorial design. This figure depicts the main effects of orthography and frequency across the simultaneous and sequential readers in the left inferior temporal gyrus. The activations have been projected on a canonical single subject template.
Since significant clusters were not seen at p < 0.001, the threshold was lowered to p < 0.01 (uncorrected) k>0. Main effects of orthography and frequency in biliterates were seen at the border of left inferior temporal gyrus and left fusiform gyrus (−40, −76, −16) (MNI co-ordinate) (Fig. 3). Data from this region was extracted by drawing a sphere of radius 8mm around the peak of activation (Poldrack, 2007). Estimates of hemodynamic responses at this ROI were measured. For the rest of the paper we refer to this as the left inferior temporal gyrus (L-ITG). No effects of interactions were observed for the reader groups.
Effects of orthography (Fig. 4)
Figure 4.

Effects of orthography. This figure depicts effect of reading English>Hindi words in all readers’ groups (biliterates and monolingual readers) at p<0.001, k>0. Top panel shows the activations of English>Hindi contrasts on a brain rendering. The blue line depicts the level at which a section has been cut, at the level of L-ITG activation. The bottom panel depicts slices of activation for L-ITG, encircled.
Effects of orthography were investigated by contrasting activations for all English words with activations for all Hindi words in biliterates and monolingual reader groups. English>Hindi contrast (p<0.001 uncorrected, k > 0) showed activations in ITG (BA 20), calcarine and superior occipital gyrus (BA 17/18) and lingual gyrus (BA 18) in the left hemisphere; calcarine and cuneus (BA 17/18), superior medial frontal gyrus (BA 9), superior frontal gyrus (BA 46) and STG (BA 48) in the right hemisphere (Fig. 4). The top panel shows activations on the brain rendering and the bottom panel highlights the L-ITG activation, which is considerably anterior to that obtained in Fig. 3. Hindi>English contrast did not reveal any significant activations.
Effects of frequency (Fig. 5, Table 3)
Figure 5.

Low>High frequency contrast images for English and Hindi. Regions of increased brain activation in the left hemisphere for reading low frequency as compared to high frequency words in English and Hindi are shown (p < 0.001, k>0). Brain areas are rendered on a standard brain conforming to stereotactic space. English monolingual readers show activations along both dorsal (inferior parietal lobule) (red) and ventral routes (inferior temporal gyrus) (blue). Hindi readers exhibit activation only along the dorsal route. Simultaneous readers show differential recruitment of reading routes, for English both dorsal and ventral routes but for Hindi, only the dorsal route. Sequential readers show increased activation in inferior parietal lobule for reading English whereas for reading Hindi, the low-high contrast shows no activation, suggesting that the areas used for reading high and low frequency words in Hindi overlap.
Table 3.
Functional areas of activation in Low>High frequency word contrasts for English and Hindi monolingual readers, simultaneous and sequential bilingual readers (p<0.001, k>0).
| Brain regions | Coordinates | t | Voxel (mm3) |
|---|---|---|---|
| Monolingual English readers | |||
| Cerebellum | −38 −66 −22 | 6.71 | 672 |
| Occipital Lobe (BA 19) | −44 −78 −12 | 9.20 | 672 |
| Precentral Gyrus (BA 6) | −42 0 58 | 5.36 | 16 |
| Supplementary Motor Area (BA 6) | −6 18 62 | 16.90 | 4384 |
| Inferior Frontal Gyrus (pars opercularis) (BA 44) | −54 12 16 | 10.12 | 4192 |
| Superior frontal gyrus (BA 6) | −14 −6 80 | 5.34 | 8 |
| Inferior Parietal Lobe (BA 40) | −40 −46 46 | 8.45 | 1496 |
| Inferior Temporal Gyrus (BA 20) | −62 −34 −16 | 6.40 | 72 |
| Monolingual Hindi readers | |||
| Cerebellum | −22 −34 −32 | 2.29 | 136 |
| Occipital Lobe (BA 19) | −32 −85 36 | 2.38 | 64 |
| Precentral Gyrus (BA 6) | −32 −16 56 | 2.08 | 40 |
| Inferior Frontal Gyrus (pars opercularis) (BA 44) | −50 10 16 | 4.68 | 34832 |
| Middle Frontal Gyrus (BA 9) | −46 18 44 | 1.97 | 8 |
| Inferior Parietal Lobe (BA 40) | −56 −32 48 | 2.98 | 1976 |
| Simultaneous bilingual readers (English) | |||
| Occipital Lobe (BA 18) | −32 −93 10 | 5.90 | 936 |
| Precentral Gyrus (BA 6) | −54 4 34 | 5.71 | 4200 |
| Inferior Frontal Gyrus (pars triangularis) (BA 45) | −52 30 8 | 5.86 | 1768 |
| Inferior Parietal Lobe (BA 40) | −48 −42 56 | 4.76 | 248 |
| Inferior Temporal Gyrus (BA 20) | −46 −38 −18 | 5.16 | 56 |
| Middle Temporal Gyrus (BA 21) | −64 −32 −8 | 6.54 | 1608 |
| Simultaneous bilingual readers (Hindi) | |||
| Cerebellum | −42 −72 −22 | 4.89 | 304 |
| Occipital Lobe (BA 18) | −18 −100 −11 | 4.52 | 32 |
| Inferior Frontal Gyrus (pars opercularis) (BA 44) | −56 12 24 | 9.68 | 5992 |
| Inferior Frontal Gyrus (pars triangularis) (BA 45) | −56 32 4 | 5.18 | 5992 |
| Middle Frontal Gyrus (BA 45) | −40 46 20 | 3.94 | 8 |
| Supramarginal gyrus (BA 48) | −62 −24 30 | 6.38 | 984 |
| Sequential bilingual readers (English) | |||
| Cerebellum | −4 −36 −12 | 6.77 | 328 |
| Lingual gyrus (BA 17) | −10 −78 4 | 5.19 | 216 |
| Middle occipital gyrus (BA 18) | −26 −88 14 | 7.99 | 3328 |
| Inferior frontal gyrus (pars opercularis) (BA 44) | −50 10 22 | 5.22 | 784 |
| Middle frontal gyrus (BA 6) | −26 2 56 | 4.58 | 64 |
| Superior frontal gyrus (BA 8) | −20 32 58 | 4.43 | 8 |
| Postcentral gyrus (BA 3) | −50 −24 56 | 6.35 | 3432 |
| Middle orbitofrontal gyrus (BA 11) | −24 43 −17 | 5.58 | 200 |
| Inferior parietal lobe (BA 40) | −52 −36 51 | 13.22 | 3432 |
| Superior parietal lobe (BA 7) | −24 −64 48 | 7.25 | 1248 |
| Sequential bilingual readers (Hindi) | |||
| Inferior orbitofrontal gyrus (BA 47) | −44 36 −16 | 4.74 | 24 |
Fig. 5 depicts the contrast images for reading English and Hindi low>high frequency words in monolinguals, simultaneous and sequential biliterates (p<0.001 uncorrected, k>0). The topmost panel shows activations of monolingual English and Hindi readers. The monolingual English readers engage the dorsal reading route via left inferior parietal lobule (L-IPL, BA 40) and ventral route via L-ITG, (BA 20/37). Monolingual Hindi readers recruit only the dorsal reading route through left IPL (BA 40). The middle panel shows that simultaneous readers while reading English engage both dorsal and ventral routes, similar to monolingual English readers. However, while reading Hindi they recruit only the dorsal route through L-IPL, similar to monolingual Hindi readers. The bottom panel shows activations for sequential readers. While reading English low>high frequency words, sequential readers show activations in L-IPL, the dorsal route. No significant activations were seen while comparing activations for reading Hindi low>high frequency words. Results of high > low frequency words showed activations in the ventral route in all populations, namely left middle temporal gyrus (BA 37) and/or L-ITG (BA 37/20) for both languages (Supplementary Fig. 1).
Effects of proficiency for reading English (L2) (Fig. 6)
Figure 6.

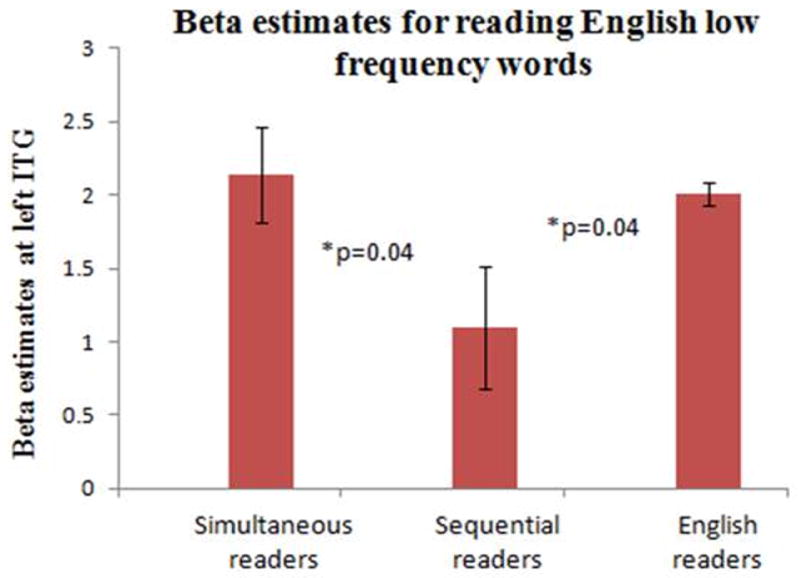
Figure 6(a). Activation in left inferior temporal gyrus for reading low frequency words in English (p<0.001). Simultaneous readers >sequential readers (top panel) shows strong differentially weighted activation cluster in left inferior temporal gyrus (ITG, BA 20), as seen in the rendering. A group ROI (region of interest) analysis, depicted in an axial slice at z = −18mm (shown with a blue line in the rendering) shows the left ITG activation (encircled in blue), rendered on a canonical single subject T1 weighted image. This activation is orthography (English)-specific and shown by simultaneous readers for reading English low frequency words, but not by sequential readers. The control groups (bottom panel) comprising the English readers show differentially weighted activation in left ITG (BA 20) and fusiform gyrus (BA 37) (encircled in blue) at z = −18mm, but the Hindi readers do not show any such activation in this region at any z level when contrasted with English readers. Hence, activation in left ITG is a marker of proficiency for reading English words, but not Hindi words.
Figure 6(b). Beta estimates. The figure shows the average beta estimate of hemodynamic response in left ITG for simultaneous, sequential and English readers. All the groups showed activations in this region, sequential readers’ responses being lower than simultaneous (two sample t-test, p=0.04) and English readers (two sample t-test, p=0.04).
The contrast image of simultaneous readers > sequential readers (Fig. 6) for reading low frequency English words (p<0.001 uncorrected, k>0) is shown in Fig. 6a. The whole brain rendering and axial slice (taken at z=−18mm) shows activation only in the L-ITG (circled in blue, BA 20) at MNI coordinate (−48, −50, −16), with a cluster size of 384mm3. Similar activations in BA 20 at (−58, −20, −20) (p<0.001, uncorrected) were also observed when contrast images between the monolingual English and Hindi readers were compared. We attribute this increased activation in left ITG while reading English to be orthography-specific.
ROI analysis
We used the simultaneous>sequential bilingual readers’ contrast for reading low frequency English words to identify the L-ITG (BA 20) location namely (−48, −50, −16). Incidentally, this co-ordinate closely matched the location described previously in literature (Paulesu et al., 2000) for orthography specific reading in English. The same co-ordinate was also examined in both simultaneous and sequential bilingual readers and beta estimates were extracted. Beta estimates of hemodynamic response (Fig. 6b) revealed significantly lower responses for sequential readers when compared to simultaneous (t-test, p=0.04) and English readers (two sample t-test, p=0.04).
Regression analysis (Fig. 7)
Figure 7.
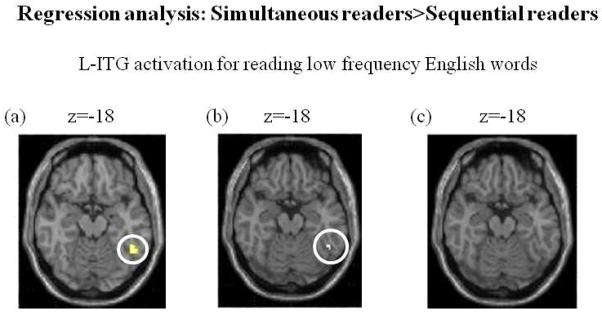
shows axial views of simultaneous>sequential readers’ contrast reading low frequency English words at z=-18, after regressing (a) vocal reaction times for low frequency English words (p<0.01 uncorrected, k=10) (b) English passage reading times (p<0.001, k>0) and (c) age of reading acquisition of English (p<0.001, k>0). L-ITG activations (encircled in white) are obtained only in (a) and (b). No L-ITG cluster was obtained in (c) wherein age of English reading acquisition was regressed. In (c) significant activation can be seen in postcentral gyrus (BA 43/48).
In order to investigate whether L-ITG activation could be predicted with reaction times, we regressed the reaction times for reading low frequency English words in proficient simultaneous and less proficient sequential readers (Fig. 7a). L-ITG (BA 20) activation at (−48, −50, −18) (p<0.01, k=10), for simultaneous>sequential readers’ contrast, was obtained (encircled as shown at z=−18). We also found activations in the frontal lobe, temporal pole and cerebellum. No activations were seen at higher threshold of p<0.001 (uncorrected).
To examine reading fluency in English as a contributing factor towards L-ITG activation for English, passage reading times for English were regressed out (Fig. 7b). Activation was seen in L-ITG (BA 20) at the same locus, (−48, −50, −18) (p<0.001, k>0) for simultaneous>sequential readers.
3) To examine the effect of age of acquisition, we regressed age of reading acquisition in English for simultaneous and sequential bilingual readers. The resultant activation at z=−18 is shown in Fig. 7c. There are no significant clusters in L-ITG at p<0.001, k>0. Clusters are seen in postcentral gyrus (BA 43/48). Even when the threshold was reduced to p<0.01, k>0, L-ITG clusters were not obtained.
Discussion
This study investigates for the first time, cortical reading networks in bilingual readers of two languages, Hindi and English, in two different orthographies, transparent and opaque respectively. Two groups of bilinguals were studied relative to monolingual control groups; one comprising bilingual simultaneous readers, who acquired reading in English and Hindi at age 5 and a second comprised of bilingual sequential readers who acquired reading Hindi at age 5 but English at 10. The language assessment and proficiency tests showed that the two bilingual groups were matched for reading proficiency in Hindi but differed in English. Our study compared the effects of orthography between the monolingual and bilinguals groups and the effects of reading proficiency and age of acquisition in the bilingual groups.
Effects of orthography
Main effects of orthography and frequency were observed in the L-ITG (Fig. 3). When main effects of orthography were further investigated by comparing all English words with all Hindi words in both bilingual and monolingual reader groups (Fig. 4), a significant effect was obtained in the L-ITG (BA 20). This region (−50, 2, −40) is considerably anterior to the L-ITG coordinates obtained for effects of proficiency (Figs. 6 and 7a-b) and needs further investigation. Since no L-ITG was obtained for Hindi > English (lower panel Fig. 6a) it can most likely be attributed to orthography effects of English as illustrated by figures 5 and 6 (lower panel of Fig. 6a, English>Hindi contrast).
(a) Monolingual readers
According to our first hypothesis, Hindi, a transparent orthography would elicit relatively greater weighting of dorsal (phonologically tuned) sites, whereas English, an opaque orthography would have relatively greater weighting of ventral (lexical semantic) sites. This effect was best captured while reading low frequency words in both languages. High frequency words in both languages elicited cortical activations in similar areas (Supplementary material). A comparison of low-frequency word reading, showed that monolingual readers of Hindi primarily elicited activations in L-IPL (BA 40) along the dorsal pathway whereas readers of English primarily employed L-ITG (BA 20/37) (Figs. 5–6) along the ventral route. Our results are in concurrence with other studies comparing reading in transparent and opaque orthographies (Paulesu et al., 2000, Meschyan and Hernandez, 2006). They also support our first hypothesis, that reading Hindi would recruit the dorsal cortical areas like L-IPL and reading in English would recruit ventral regions of the brain like L-ITG.
(b) Bilingual readers
Our second hypothesis concerned the engagement of these specific areas, namely L-IPL for Hindi and L-ITG for English by bilingual readers of both languages. At the behavioural level, for Hindi, which was L1, we expected to find no differences in vocal reaction times between simultaneous and sequential readers. With regard to the loci of cortical activation, we expected to find engagement of L-IPL for both simultaneous and sequential bilingual readers reading low frequency words in Hindi, as seen in the Hindi monolinguals described above. For English, we hypothesized activation in L-ITG for only simultaneous bilingual readers. For the second language English, we expected the late sequential readers to exhibit a network that overlapped with L1, similar to Tan et al. (2003). In early simultaneous bilinguals however, the network recruitment remains an open question. While on one hand, studies by Kim et al. (1997) have shown common networks for L1 and L2 in early bilinguals, we hypothesize that early exposure and high proficiency could exhibit greater language-specific modulation. We therefore hypothesize that the simultaneous readers might possibly show English-specific L-ITG. Our results showed that early simultaneous bilingual readers, proficient in reading both orthographies recruited the dorsal cortical route namely L-IPL for reading Hindi and L-ITG along the ventral route for English (Figs. 5, 6). But, the late sequential readers, less proficient in reading English, engaged areas primarily along the dorsal route (L-IPL) for both Hindi and English (Fig. 5). This finding supports our second hypothesis that the degree of Hindi and English specificity of weighting should be greater for those who learned the two orthographies early (simultaneous readers); whereas sequential readers would activate similar areas for both orthographies.
Comparison of reading pathways in the bilingual group
The sequential and simultaneous readers differed significantly in years of formal English training, number of hours spent in reading English (Table 1). At the behavioral level, differences were reflected in both the behavioral scores, namely passage reading time and vocal reaction time (Fig. 2a and b). The sequential readers were found to be significantly slower than both, monolingual English readers and simultaneous readers (Fig. 2a) for reading the English passage. For vocal reaction times in English, the sequential bilinguals were slower than the simultaneous and monolingual readers; however significant differences between response times were seen only for the low frequency words. A comparison of simultaneous>sequential activations revealed clusters in L-ITG (Fig. 6a). As seen in the beta estimates (Fig. 6b), both the bilingual groups show ITG activation, but the level of activation is significantly higher for the simultaneous readers showing the preferential weighting/usage of this region in proficient versus less proficient readers of English and thereby reiterates the role of reading proficiency in selecting the optimal pathway for a specific orthography.
The simultaneous readers not only showed orthography specific pathways for reading English and Hindi, they also showed native-like reading proficiency in both languages. This was evident from two results. 1) When compared with English monolingual readers, the simultaneous readers showed no significant differences in reading times and vocal reaction times (Figs. 2a-b). Like the English monolinguals they showed recruitment of L-ITG while reading low frequency words in English and also did not show significant differences in the beta estimates of L-ITG (Fig. 6a-b). They also showed closeness to neuroanatomical locus of activation reported in other studies with English monolinguals as discussed below. Neuroimaging studies by Paulesu and colleagues (2000) have showed orthography-specific activation in English in the L-ITG (BA 21/37) at MNI coordinate (−58, −58, −14). When compared with Paulesu et al.’s (2000) results, our study localized the co-ordinate of activation for English at (−48, −50, −16) (MNI coordinate system), very close to the coordinate reported by Paulesu et al. (2000), with a difference of x=+10, y=+8 and z=-2. Our coordinates were slightly more medial, anterior and ventral, but closeness with that of Paulesu and colleagues is quite evident.
Whether the native-like reading proficiency in English, demonstrated by the simultaneous readers, is a consequence of early acquisition or reading proficiency remains to be established. Following is an attempt to shed some light in this direction:
We regressed reaction times for reading low frequency English words as covariates and expected an effect of proficiency to appear in L-ITG. Interestingly, this effect was seen only at a lower threshold of p<0.01 uncorrected (Fig. 7a), thereby suggesting that additional factors might be playing a role in modulating the reading route, not merely the reading proficiency (measured in terms of reaction times). These factors could be age of acquisition and/or reading proficiency. This finding wherein the L-ITG activation is seen only for simultaneous>sequential contrast (Fig. 6) while reading low frequency English words is complementary to our findings shown in Figure 5 where L-ITG was not recruited by the sequential readers while reading low (>high) frequency words in English or Hindi. Therefore, our results suggest a strong association of L-ITG activation with proficient reading in English.
To investigate the contribution of reading proficiency in L-ITG activation, we regressed passage reading time as a covariate (Fig. 7b) in simultaneous versus sequential readers. We obtained localized cluster of activation in L-ITG at the same locus, clearly suggesting a strong role of reading proficiency in English with L-ITG activation.
In an attempt to examine the contribution of age of acquisition in L-ITG activation, we regressed the ages of acquisition of English (Fig. 7c) in bilingual readers. Absence of L-ITG recruitment during English reading suggests that L-ITG activation obtained during reading English would depend upon reading proficiency.
In summary, our results indicate a major role for reading proficiency in cortical reading networks. However, a recent study comparing adult literates who learnt to read early versus those who learnt late, showed structural and connectivity differences suggesting a role for age of acquisition (Carreiras et al., 2009) Clearly an interplay of the two factors along with exposure and practice modulates reading pathways.
In the following section we discuss the roles of L-IPL and L-ITG in reading.
Possible role of L-IPL
In a transparent orthography, word reading relies primarily on the grapheme to phoneme mapping accomplished by areas along the dorsal route (planum temporale, BA 22/42) (Paulesu et al., 2000)/L-STG (BA 22) (Meschyan and Hernandez, 2006). That transparent orthographies like Hindi follow the grapheme to phoneme route to reading is strongly motivated by the dual route theory for reading. Visual words are transformed to their auditory counterparts, based on univalent grapheme to phoneme correspondence. These are sequentially assembled to make available the pronunciation of a word, following which readers can access meaning (Jobard et al., 2003). Readers when confronted with novel or low frequency words in any orthography also adopt such grapheme to phoneme assembly. We therefore posit L-IPL as a candidate for sound assembly while reading.
Possible role of L-ITG
In some theoretical accounts low frequency words in English are believed to require retrieval of pronunciation of letter combinations/letters from previously known words or common words; thus more constraint from previous lexical knowledge is needed (Glushko, 1979). For example, to pronounce the low frequency word ‘goad’, we use a common word like ‘road or toad’ as a template and hence pronounce ‘oa’ as ‘o’ and not /o/ /a/ separately. This may be attributed to the fact that in English, wherein sound letter mapping is inconsistent, in addition to grapheme to phoneme processing, frequent access to orthographic structure dictionary (Paulesu et al., 2000) is also necessary to resolve ambiguities in pronunciation, a process believed to be accomplished by the L-ITG along the ventral route. Thus reading both high and low frequency words in English might require relatively greater reliance on lexico-semantic access, believed to be associated with L-ITG, a finding that has also been suggested by other neuroimaging studies on word retrieval in English (Damasio et al., 1996; Pugh et al., 1996; Price, 2000). While further work is required to ascertain whether same or distinct anatomical regions within L-ITG are responsible for reading low and high frequency words, one expects to find recruitment of this neural site for efficient English-reading to occur.
Whether activation in L-ITG predicts native-like reading in English can be probed further by comparing activation patterns for different groups, such as a) early simultaneous bilingual readers with low proficiency b) late sequential bilingual readers with high proficiency. Such studies would not only provide further insights into the reading process in bilinguals/multilinguals but would also dissect out the specific roles of proficiency and age of acquisition.
Thus early simultaneous bilingual proficient readers of two languages Hindi (L1) and English (L2) exhibit distinct orthography specific routes whereas late sequential readers exhibit a common (L1) reading route for both languages. Advantages of bilingualism in terms of cognitive control and flexibility, as a consequence of early exposure to auditory input from two (or more) languages have been demonstrated in adults and infants alike (Bialystok, 2001; Bialystok et al., 2008; Bialystok and Viswanathan, 2009; Mechelli et al., 2004; Kovacs and Mehler, 2009a; Kovacs and Mehler, 2009b). The findings of the current study demonstrate that early simultaneous adult readers, proficient in reading both orthographies, exhibit orthography-specific plasticity. Whether the proficiency exhibited by simultaneous readers is a consequence of early acquisition or practice remains to be ascertained.
Conclusion
In summary, (1) simultaneous proficient bilingual readers recruited distinct routes for reading different orthographies, Hindi and English. (2) Our behavioural, fMRI and beta estimate results show that the reading routes employed by simultaneous readers were more flexible and similar to monolinguals. For Hindi, they employed L-IPL along the dorsal route and for English they engaged L-ITG along the ventral route, as also recruited by respective native readers. (3) Our results implicate a strong role of reading proficiency could play a major role in determining native-like reading pathways.
Future studies should be addressed at determining if sequential readers matched for behavioral characteristics with monolingual/simultaneous readers also demonstrate such orthographic plasticity in reading networks. What this study does establish for the first time is that, functional plasticity in reading two languages is demonstrated by early simultaneous proficient bilingual readers.
Supplementary Material
Acknowledgments
We would like to thank Steve Frost and Einar Mencl and two anonymous referees for providing excellent and insightful comments that helped in considerable improvements to the manuscript. We would also like to acknowledge Bharat Biswal and Michael Posner for useful comments. We thank J. Ahlawat for technical support, M. K. Singh, T. Sumathi, Anya Chakraborty and T. Padma for assistance in the reaction time analysis and R. S. Bapi for assistance in image analysis. This research was supported by intramural funding from the National Brain Research Centre, India. Kenneth Pugh's contributions were supported by National Institutes of Health grants awarded to Haskins Laboratories [5P01HD001994-430027].
Footnotes
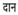 (linear vowel marker),
(linear vowel marker),
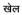 (vowel marker on top of consonant),
(vowel marker on top of consonant),
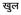 (vowel marker below the consonant).
(vowel marker below the consonant).
Hindi, written in Devanagari (Bright, 2000) is an example of transparent orthography, wherein 45 sounds are sufficient to represent 48 letters; making it more consistent than Italian wherein 33 letters are mapped onto 25 sounds (Lepschy and Lepschy, 1981).
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bialystok E. Bilingualism in development: Language, literacy, and cognition. New York: Cambridge University Press; 2001. [Google Scholar]
- Bialystok E, Craik FIM, Luk G. Cognitive control and lexical access in younger and older bilinguals. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2008;34:859–873. doi: 10.1037/0278-7393.34.4.859. [DOI] [PubMed] [Google Scholar]
- Bialystock E, Viswanathan M. Components of executive control with advantages for bilingual children in two cultures. Cognition. 2009;112:494–500. doi: 10.1016/j.cognition.2009.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolger DJ, Perfetti CA, Schneider W. Cross-cultural effect on brain revisited: Universal structures plus writing system variation. Human Brain Mapping. 2005;25:92–104. doi: 10.1002/hbm.20124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bright W. A matter of typology: Alphasyllabaries and Abugidas. Studies in the Linguistic Sciences. 2000;30:63–71. [Google Scholar]
- Carreiras M, Seghier ML, Baquero S, Estevez A, Lozano A, Devlin JT, Price CJ. An anatomical signature for literacy. Nature. 2009;461:983–986. doi: 10.1038/nature08461. [DOI] [PubMed] [Google Scholar]
- Chee MWL, Tan EWL, Thiel T. Mandarin and English single word processing studied with functional magnetic resonance imaging. The Journal of Neuroscience. 1999;19:3050–3056. doi: 10.1523/JNEUROSCI.19-08-03050.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crinion J, Turner R, Grogan A, Hanakawa T, Noppeney U, Devlin JT, Aso T, Urayama S, Fukuyama H, Stockton K, Usui K, Green DW, Price CJ. Language control in bilingual brain. Science. 2006;312:1537–1540. doi: 10.1126/science.1127761. [DOI] [PubMed] [Google Scholar]
- Damasio H, Grabowski TJ, Tranel D, Hichwa RD, Damasio AR. A neural basis for lexical retrieval. Nature. 1996;380:499–505. doi: 10.1038/380499a0. [DOI] [PubMed] [Google Scholar]
- Friston KJ, Holmes AP, Worsley KJ, Poline JB, Frith CD, Frackowiak RS. Statistical parametric maps in functional imaging: a general linear approach. Human Brain Mapping. 1995a;2:189–210. [Google Scholar]
- Friston KJ, Ashburner J, Frith CD, Poline JB, Heather JD, Frackowiak RS. The spatial registration and normalization of images. Human Brain Mapping. 1995b;2:165–189. [Google Scholar]
- Glushko RJ. The organization and activation of orthographic knowledge in reading aloud. Journal of Experimental Psychology: Human Perception and Performance. 1979;5:674–691. [Google Scholar]
- Haller S, Radue EW, Erb M, Grodd W, Kircher T. Overt sentence production in eventrelated fMRI. Neuropsychologia. 2005;43:807–814. doi: 10.1016/j.neuropsychologia.2004.09.007. [DOI] [PubMed] [Google Scholar]
- Holmes AP, Friston KJ. Generalisability, random effects and population inference. Neuroimage. 1998;7:S754. [Google Scholar]
- Jobard G, Crivello F, Tzourio-Mazoyer N. Evaluation of the dual route theory of reading: a metanalysis of 35 neuroimaging studies. Neuroimage. 2003;20:693–711. doi: 10.1016/S1053-8119(03)00343-4. [DOI] [PubMed] [Google Scholar]
- Kim KHS, Relkin NR, Lee K-M, Hirsch J. Distinct cortical areas associated with native and second languages. Nature. 1997;388:171–174. doi: 10.1038/40623. [DOI] [PubMed] [Google Scholar]
- Klein D, Milner B, Zatorre RJ, Zhao V, Nikelski J. Cerebral organization in bilinguals: A PET study of Chinese-English verb generation. Neuroreport. 1999;10:2841–2846. doi: 10.1097/00001756-199909090-00026. [DOI] [PubMed] [Google Scholar]
- Klein D, Zatorre RJ, Chen J-K, Milner B, Crane J, Belin P, Bouffard M. Bilingual brain organization: A functional magnetic resonance adaptation study. Neuroimage. 2006;31:366–375. doi: 10.1016/j.neuroimage.2005.12.012. [DOI] [PubMed] [Google Scholar]
- Kovacs AM, Mehler J. Cognitive gains in 7-month-old bilingual infants. PNAS. 2009a;106:6556–6560. doi: 10.1073/pnas.0811323106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kovacs AM, Mehler J. Flexible learning of multiple speech structures in bilingual infants. Science. 2009b;325:611–612. doi: 10.1126/science.1173947. [DOI] [PubMed] [Google Scholar]
- Kumar U, Das T, Bapi RS, Padakannaya P, Joshi RM, Singh NC. Reading different orthographies: an fMRI study of phrase reading in Hindi-English bilinguals. Reading and Writing: an interdisciplinary journal. 2009;23:239–255. [Google Scholar]
- Lepschy A, Lepschy G. La Lingua Italiana. Bompiani; Milan: 1981. [Google Scholar]
- Marian V, Spivey M, Hirsch J. Shared and separate system in bilingual language processing: Converging evidence from eyetracking and brain imaging. Brain and Language. 2003;86:70–82. doi: 10.1016/s0093-934x(02)00535-7. [DOI] [PubMed] [Google Scholar]
- Mechelli A, Crinion JT, Nopperney U, O’Doherty J, Ashburner J, Frackowiak RS, Price CJ. Structural plasticity in the bilingual brain. Nature. 2004;431:757. doi: 10.1038/431757a. [DOI] [PubMed] [Google Scholar]
- Meschyan G, Hernandez AE. Impact of language proficiency and orthographic transparency on bilingual word reading: an fMRI investigation. Neuroimage. 2006;29:1135–1140. doi: 10.1016/j.neuroimage.2005.08.055. [DOI] [PubMed] [Google Scholar]
- Oldfield RC. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia. 1971;9:97–113. doi: 10.1016/0028-3932(71)90067-4. [DOI] [PubMed] [Google Scholar]
- Paulesu E, McCrory E, Fazio F, Menoncello L, Brunswick N, Cappa SF, Cotelli M, Cossu G, Corte F, Lorusso M, Pesenti S, Gallagher A, Perani D, Price C, Frith CD, Frith U. A cultural effect on brain function. Nature Neuroscience. 2000;3:91–96. doi: 10.1038/71163. [DOI] [PubMed] [Google Scholar]
- Penny WD, Holmes AP, Friston KJ. Human Brain Function. London: Academic Press; 2003. [Google Scholar]
- Perfetti CA. The universal grammar of reading. Scientific Studies of Reading. 2003;7:3–24. [Google Scholar]
- Perfetti CA, Liu Y, Fiez J, Nelson J, Bolger DJ. Reading in two writing systems: Accomodation and assimilation of the brain's reading network. Bilingualism: Language and Cognition. 2007;10:131–146. [Google Scholar]
- Poldrack RA. Region of interest analysis for fMRI. SCAN. 2007;2:67–70. doi: 10.1093/scan/nsm006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price CJ. The anatomy of language: contributions from functional neuroimaging. Journal of Anatomy. 2000;197:335–359. doi: 10.1046/j.1469-7580.2000.19730335.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pugh KR, Shaywitz BA, Shaywitz SE, Constable RT, Skudlarski P, Fulbright RK, Bronen RA, Shankweiler DP, Katz L, Fletcher JM, Gore JC. Cerebral organization of component processes in reading. Brain. 1996;119:1221–1238. doi: 10.1093/brain/119.4.1221. [DOI] [PubMed] [Google Scholar]
- Tan LH, Spinks JA, Feng C-M, Siok WT, Perfetti CA, Xiong J, Fox PT, Gao J-H. Neural systems of second language reading are shaped by native language. Human Brain Mapping. 2003;18:158–166. doi: 10.1002/hbm.10089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue G, Dong Q, Jin Z, Zhang L, Wang Y. An fMRI study with semantic access in low proficiency second language learners. Neuroreport. 2004;15:791–796. doi: 10.1097/00001756-200404090-00010. [DOI] [PubMed] [Google Scholar]
- Zieler JC, Goswami U. Reading acquisition, developmental dyslexia, and skilled reading across languages: A psycholinguistic grain size theory. Psychological Bulletin. 2004;131:3–29. doi: 10.1037/0033-2909.131.1.3. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.