

Abstract
Sexually antagonistic polymorphisms are polymorphisms in which the allele that is advantageous in one sex is deleterious in the other sex. In an influential 1984 paper, W. Rice hypothesized that such polymorphisms should be relatively common on the X chromosome (or on the W in female-heterogametic species) but relatively rare on the autosomes. Here, I show that there are plausible assumptions under which the reverse is expected to be true, and point out recent studies that give evidence for sexually antagonistic variation on the autosomes. Although more work is needed to resolve the issue, it is premature to conclude that the X chromosome is a “hot spot” for the accumulation of sexually antagonistic variation.
Major components of fitness, such as fecundity or mating success, show appreciable heritable variation in most animal and plant populations (reviewed in Mousseau and Roff 1987; Roff and Mousseau 1987; Houle 1992; Geber and Griffen 2003). This at first seems paradoxical, because natural selection would be expected to eliminate variants with low fitness. Hypotheses for the maintenance of heritable (i.e., additive-genetic) variation for major fitness components can be divided into three broad categories (cf. Houle 1998; Charlesworth and Hughes 1999): high rates of recurrent deleterious mutation; failure of populations to reach equilibrium (e.g., due to a constantly changing environment); and various types of balancing selection, defined as any purely selective mechanism that will maintain genetic variation in an equilibrium population.
A particularly interesting type of balancing selection occurs when opposite alleles are favored in the two sexes. This situation can result in the maintenance of stable polymorphism in a homogenous environment, even without overdominance (Kidwell et al. 1977; Rice 1984). At equilibrium, additive variation for fitness will be present in each sex, with a negative genetic correlation between sexes. Interestingly, several recent studies have given evidence for such a situation (Chippindale et al. 2001; Gibson et al. 2002; Calsbeek and Sinervo 2004; Fedorka and Mousseau 2004; Foerster et al. 2007; Brommer et al. 2007; Delcourt et al. 2009), suggesting that sexually antagonistic polymorphisms are common.
One effect of sexually antagonistic genetic variation (or “intralocus sexual conflict”; Day and Bonduriansky 2004; Bonduriansky and Chenowith 2009) is to reduce the benefit to females of mating with high fitness males, because such males will tend to sire low fitness daughters. The benefit of choosing high-fitness males would be even further reduced if the sexually antagonistic variation is concentrated on the X chromosome in a male heterogametic species. In this case, females choosing high fitness males will have low fitness daughters without the corresponding benefit of high fitness sons, because males transmit the X only to their daughters. This situation could even cause the counterintuitive evolution of female preference for low fitness males (Albert and Otto 2005).
In 1984, W. Rice presented a population-genetic model that showed that under certain assumptions, sexually antagonistic polymorphisms should indeed be expected to accumulate disproportionately on the X. Rice emphasized the situation where the allele that is beneficial in males but deleterious in females is recessive or partly recessive. In this case, when the male-beneficial allele is rare, X-linkage facilitates its maintenance in the population by exposing it to selection in hemizygous males. Conversely, when the dominant, female-beneficial allele is rare, X-linkage facilitates its maintenance compared to the autosomal case because X-linked genes spend 2/3 of their time in females, compared to only one-half of their time for autosomal genes.
Rice’s (1984) paper was influential, having been cited over 230 times. Its most direct prediction is that within populations, X chromosomes, but not autosomes, that give rise to high fitness in one sex should give rise to low fitness in the other sex. This prediction has been elegantly confirmed in a series of experiments on a laboratory population of Drosophila melanogaster (Chippindale et al. 2001; Gibson et al. 2002; Pischedda and Chippindale 2006), and, more tentatively, in a long-term study of a population of red deer (Foerster et al. 2007). Rice’s model has also been invoked in discussions of the genomic location of variation in sexually dimorphic traits, and of genes showing differential expression between the sexes, although its predictions regarding these issues are not always clear (see section on “Empirical Evidence” below).
The purpose of this note is to argue that, although Rice’s (1984) paper has been useful in stimulating discussion and research, it is premature to accept its conclusions, on either theoretical or empirical grounds. First, I review the theoretical conditions for the maintenance of sexually antagonistic polymorphism at X-linked and autosomal loci, and show that there are plausible assumptions under which the conditions are more lenient for autosomal loci. Second, I review the small but growing experimental literature on sexually antagonistic fitness variation, pointing out three studies that give evidence for an autosomal basis to the variation. I conclude with some suggestions for future research.
REEXAMINATION OF THE POLYMORPHISM CONDITIONS
Rice (1984) derived the conditions for the maintenance of polymorphism by sexually antagonistic selection for X-linked and autosomal loci using a fitness scheme equivalent to that in the first three rows of Table 1, which is adapted from Kidwell et al. (1977). In Kidwell et al.’s notation, which has certain advantages over that used by Rice (see Appendix for comparison), the most fit genotype in a given sex has a relative fitness of 1, and sm and sf are the selection coefficients against the less-fit homozygote (or hemizygote) in males and females, respectively. Similarly, hm and hf represent the dominance of the less-fit allele in males and females (because there are no heterozygotes for an X-linked locus in males, hm is not applicable to this case).
Table 1.
Fitness values at a sexually antagonistic locus, after Kidwell et al. (1977). sf and sm (0 < sf,sm ≤ 1) represent the strength of selection in females and males, respectively. hf and hm (0 ≤ hf,hm ≤ 1) are sex-specific dominance parameters; specifically, hf (hm) is the dominance in females (males) of the allele which is deleterious in females (males). Note that hm applies only to the autosomal case. Rice (1984) assumed equal dominance of a given allele in the two sexes; in Kidwell et al.’s notation, this implies hm = 1 − hf. Identities relating Kidwell et al.’s parameters to those used by Rice are given in Appendix Table 1.
| Genotype
|
|||
|---|---|---|---|
| A1A1 or A1Y | A1A2 | A2A2 or A2Y | |
| Females, X-linked or autosomal locus | 1 | 1 − hfsf | 1 − sf |
| Males, X-linked locus | 1 − sm | NA | 1 |
| Males, autosomal locus, Rice’s assumption | 1 − sm | 1 − (1 − hf)sm | 1 |
| Males, autosomal locus, general dominance | 1 − sm | 1 − hmsm | 1 |
NA: not applicable.
Rice assumed that, in the autosomal case, the dominance of a given allele is the same in the two sexes. In Kidwell et al.’s notation, this occurs when hm = 1 − hf (equivalently, hf = 1 − hm). Kidwell et al. showed that, with this assumption, polymorphism at an autosomal locus is guaranteed whenever
| (1) |
independent of dominance. The first inequality gives the condition for “protection” (increase when rare) of the male-beneficial, female-deleterious A2 allele, while the second inequality gives the protection conditions for the female-beneficial, male-deleterious A1 allele. As might be expected, it becomes easier to satisfy the first condition as sm gets larger, thus increasing the strength of selection favoring A2 in males, but this makes it harder to satisfy the second condition. For an X-linked locus, the protection conditions are (cf. Rice 1984; Hedrick 2000):
| (2) |
The autosomal polymorphism conditions can be compared to the X-linked conditions with particular values of hf by viewing the regions on the {sm, sf} plane that satisfy (1) and (2) (Fig. 1). The X-linked case with low hf (partial recessivity of the female-deleterious allele) supports polymorphism over the broadest region, followed by the autosomal case, and then X-linkage with additivity in females (hf = 0.5). When the female-deleterious allele is nearly dominant (hf = 0.9), an X-linked polymorphism can be maintained only with absurdly high selection coefficients in both sexes (Fig. 1A). Nonetheless, for more realistic selection coefficients (sm, sf ≤ 0.1), the only case that supports polymorphism over a relatively large region is X-linkage with low hf (Fig. 1B). In the autosomal case, in contrast, selection coefficients in males and females must be nearly equal to support polymorphism. This is the basis for Rice’s conclusion that the X chromosome should be particularly favorable for the accumulation of sexually antagonistic polymorphisms.
Figure 1.
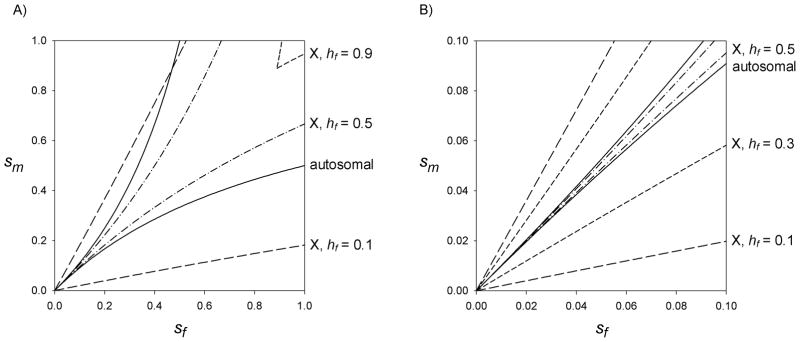
Conditions for the maintenance of sexually antagonistic polymorphism at an X-linked locus for three values of hf, and at an autosomal locus under the assumption of equal dominance in males and females. For each case, polymorphism is maintained in the region between the matching lines. Parameter combinations below the upper lines result in protection (increase when rare) of the female-beneficial, male-deleterious A1 allele, while parameter combinations above the lower lines result in protection of the male-beneficial, female-deleterious A2 allele.
Rice’s conclusion, however, depends heavily on the assumption that the dominance of an allele is the same in the two sexes (cf. Gavrilets and Rice 2006). At first glance, this assumption appears reasonable, because major visible mutations (in domestic animals, Drosophila, etc.), as well as mutations causing Mendelian disorders in humans, are usually dominant or recessive irrespective of sex. It is debatable, however, whether such mutations provide a good model for sexually antagonistic alleles segregating in natural populations, which are likely to have less drastic effects. Moreover, it is important to distinguish between the dominance of an allele with respect to a particular trait and that with respect to fitness itself; the two are not necessarily the same. As a biologically plausible example, consider the situation where the sexes differ with respect to the optimum level of activity of a specific gene product (Fig. 2). Suppose that A1A1 homozygotes have optimum activity in females, and A2A2 homozygotes have optimum activity in males, with heterozygotes exactly intermediate. (The differences in activity could result from differences in amount of gene product, or from an amino acid substitution that changes the biochemical activity of the product). Even though the alleles have additive effects on gene product activity, the concavity of the fitness functions in the vicinity of the optima cause their effects on fitness to be non-additive. In particular, because heterozygotes in each sex are closer in fitness to the more-fit homozygote, whichever allele is favored in a given sex is partly dominant in that sex. A consequence is that the dominance of a given allele’s fitness effect differs between the sexes (e.g., A2 is partly dominant in males but partly recessive in females).
Figure 2.

Illustration of how unequal dominance between the sexes could arise at a sexually-antagonistic autosomal locus. The optimum level of gene product activity is higher in males than in females, and the alleles have additive, sex-independent effects on gene product activity. As a consequence of the concavity of the fitness functions in the vicinity of the optima, whichever allele is beneficial (deleterious) in a given sex is partly dominant (recessive) in that sex. This situation is particularly favorable to the maintenance of polymorphism, because it creates heterozygote superiority when fitnesses are averaged across the sexes.
This example suggests that the possibility of unequal dominance in the two sexes, and particularly partial dominance of the more-fit allele in each sex, needs to be seriously considered. Kidwell et al. (1977) showed that, with general dominance in the two sexes (Table 1, rows 1 and 4), polymorphism at an autosomal locus will be guaranteed whenever:
| (3) |
To compare these conditions with the X-linked conditions, one can visualize the regions of the {hf, sm/sf} plane that satisfy (2) and (3) for given values of sf and (for the autosomal case) hm (Fig. 3). Results are shown for weak selection in females (sf small enough that terms in sf in the denominators of the left- and right-hand sides of (2) and (3) can be neglected; Fig. 3A) and moderately strong selection in females (sf = 0.2; Fig. 3B). Recalling that hm is the dominance of the male-deleterious A1 allele in males, and hf is the dominance of the female-deleterious A2 allele in females (Table 1), partial dominance of the more-fit allele in each sex occurs when both hm and hf < 0.5. In this situation, polymorphism is possible over a broader range of sm/sf in the autosomal case than in the X-linked case (Fig. 3; example shown for hm = 0.2). The reason that low hm and hf make polymorphism relatively easy to maintain is not hard to appreciate intuitively: as long as selection coefficients in males and females are not too dissimilar, partial dominance of the more-fit allele in each sex results in heterozygote superiority when fitnesses are averaged across sexes.
Figure 3.

Conditions for the maintenance of sexually antagonistic polymorphism, relaxing the assumption of equal dominance in males and females in the autosomal case. For each case, polymorphism is maintained in the region between the matching lines (see legend to Fig. 1). (For comparison, with Rice’s dominance assumption, the range of sm/sf supporting an autosomal polymorphism would be vanishingly small in the weak selection case; with sf = 0.2, polymorphism would require that sm/sf lie between 0.8333 and 1.25).
EMPIRICAL EVIDENCE
It is clear from the above that plausible arguments can be made as to why the X chromosome should either be a more favorable, or a less favorable, environment for the accumulation of sexually antagonistic polymorphisms than the autosomes. The only way the matter can be resolved is through empirical evidence. The most relevant evidence is provided by experiments that allow assessment of the chromosomal location of antagonistic variation in sex-specific components of fitness (as opposed to traits that could be under stabilizing selection). I am aware of such experiments on five species.
An elegant experiment on the LHM laboratory population of D. melanogaster revealed a strong negative genetic correlation between male mating success and female fecundity (Chippindale et al. 2001) that was later found to map almost exclusively to the X chromosome (Gibson et al. 2002). X-linkage of sexually antagonistic variation in D. melanogaster has subsequently been confirmed in the same (Pischedda and Chippindale 2006) and a different (Connallon and Jakubowski 2009) laboratory population, so the result is clearly robust. More equivocal evidence for X-linked sexually antagonistic variation comes from a study of a pedigreed wild population of the red deer Cervus elaphus (Foerster et al. 2007). The authors found a significant negative correlation in a measure of total fitness between sires and their daughters; in contrast, there was no significant correlation between sire fitness and son fitness. While this result would be expected from X-linked sexually antagonistic variation (because fathers do not transmit their X to their sons), there were only 30 sires, and three points in the father-son regression appeared to be outliers. The absence of a significant father-son correlation therefore cannot be taken as convincing evidence for X-linkage. The authors’ animal model analysis detected a highly significant negative genetic correlation between female and male reproductive success, but this model assumes an autosomal basis for the inheritance. Thus while Foerster et al.’s (2007) results clearly show that sexually antagonistic variation was present, the evidence for X-linkage is equivocal.
In contrast to the above studies, three studies give evidence for sexually antagonistic genetic variation with an autosomal basis. Studying the cricket Allonemobius socius, Fedorka and Mousseau (2004) compared the fitness of laboratory-reared offspring of field-collected males that had either mated (successful) or not mated (unsuccessful) in the wild. Successful males sired sons with significantly higher mating success, but daughters with significantly lower reproductive success, than unsuccessful males. Because males do not transmit their X chromosome to their sons, these results indicate that the sexually antagonistic variation was autosomally inherited. Similarly, a recent study of Drosophila serrata used a paternal half-sib design to document significant heritable variation in both male and female fitness, with a significant negative genetic correlation between the two (Delcourt et al. 2009). Once again, this is inconsistent with X-linkage, which would have resulted in there being no sire effect on male fitness (and an inestimable genetic correlation). Finally, in a field study of the side-blotched lizard Uta stansburiana, Calsbeek and Sinervo (2004) found that large males sired sons with higher survival, but daughters with lower survival, than small males. Sample sizes were small, but the results are consistent with sexually antagonistic variation with an autosomal basis. While these studies do not rule out the possibility that there is some sexually antagonistic variation on the X in these species, they give evidence against the notion that such variation is not likely to be present on autosomes.
In summary, there is one conclusive and one tentative example of sexually antagonistic variation that maps predominantly to the X, compared to three cases where it appears to map to the autosomes. Moreover, although the D. melanogaster results are striking, it should be kept in mind that the X contains nearly 20% of the D. melanogaster genome. Thus if the sexually antagonistic variation observed in this species is the result of major-effect polymorphisms at one or two genes, it could be coincidence that the gene(s) are X-linked. It is therefore premature to conclude that the X chromosome has properties that make it a “hot spot” for accumulation of sexually antagonistic variation.
It is tempting to also consider evidence on the chromosomal location of variation in sexually dimorphic quantitative traits (e.g., body size), as opposed to variation in male- and female-specific fitness itself, in assessing the predictions of Rice’s (1984) model. Although sexually dimorphic traits may contribute to the X-linked sexually antagonistic variation observed in D. melanogaster (Prasad et al. 2007), however, it cannot be assumed that genetic variation in a sexually dimorphic trait automatically creates a trade-off between the sexes. Sexually dimorphic traits result in sexually antagonistic selection only if there is sex-specific directional selection on breeding values that is in opposition to the sign of the genetic correlation between the sexes (Poissant et al. 2008; Bonduriansky and Chenoweth 2009). If the population means are at their sex-specific optima, or if the genetic correlation between the sexes is zero, selection on the trait in one sex does not oppose selection on the trait in the other sex. Moreover, single-locus models like those considered above are poor predictors of the outcome of stabilizing or disruptive selection on polygenic traits. Polygenic models have complex dynamics, because selection on individual alleles changes in intensity and direction depending on genetic background. For these reasons, Rice’s model does not give a strong basis for predicting that variation in sexually dimorphic polygenic traits should map preferentially to the X.
Another potential line of evidence comes from microarray studies of gene expression in diverse animal taxa showing that genes preferentially expressed in one sex (“sex-biased genes”) tend to be overrepresented or underrepresented on the X chromosome (reviewed in Ellegren and Parsch 2007). Even if one accepts the dominance assumptions of Rice’s model, however, the predictions it makes about the chromosomal distribution of sex-biased genes are not clear-cut (see the excellent discussion of this issue by Vicoso and Charlesworth 2006). Moreover, the unique biological properties of the X chromosome, including meiotic inactivation in the male germ-line and dosage compensation, appear to have a strong influence on the chromosomal distribution of sex-biased genes (Vicoso and Charlesworth 2006, 2009). For these reasons, patterns of sex-biased gene expression do not provide a simple test of Rice’s model.
Rice’s (1984) model, and my comments, apply only to X or W-linked loci in species with heteromorphic, non-recombining sex chromosomes, in which most X or W-linked loci have no homologues on the Y or Z. There is of course no barrier to the accumulation of male-beneficial, female-detrimental alleles on the Y chromosome, which never experiences selection in females. There are also robust theoretical reasons for expecting sexually antagonistic polymorphisms to accumulate linked to the sex-determining locus in species with non-heteromorphic, recombining sex chromosomes (Rice 1987). Thus the extensive “sex-linked” and presumably sexually antagonistic color pattern polymorphism in poeciliid fish is not relevant to Rice’s (1984) model, because the variation is either Y-linked (Poecilia: Lindholm et al. 2004; Hughes et al. 2005), or present on non-heteromorphic, recombining sex chromosomes (Xiphophorus: Basolo 2006).
CONCLUSION
More experiments on diverse taxa will be needed to resolve whether sexually antagonistic variation tends to accumulate disproportionately on the X chromosome (or the W chromosome in female-heterogametic species). The most powerful approach, when possible, is to perform reciprocal crosses between replicated genotypes with divergent male and female fitness (Pischedda and Chippindale 2006). Otherwise, parent-offspring regressions are useful, provided that sample sizes are large enough that a lack of significant correlation between fitness of fathers and fitness of sons can be confidently interpreted as evidence for X-linkage. For wild populations with pedigree data, it should be possible to fit animal models that allow separate estimation of X- or W-linked and autosomal components of variance and covariance (Meyer 2008). Such an approach could in principle resolve whether the sexually antagonistic variation observed by Foerster et al. (2007) in deer and Brommer et al. (2007) in flycatchers is autosomal or sex-linked. Future research should also strive to identify the genes giving rise to sexually antagonistic variation, about which nothing is currently known. Among other benefits, this would allow estimation of the dominance coefficients upon which hinge the question of whether sexually antagonistic polymorphisms are more likely to be maintained on the X chromosome or the autosomes.
Acknowledgments
This work was supported by NSF grant 0623268 and NIH grant R01AA16178 to J.D.F. I thank G. Arnqvist and two anonymous reviewers for helpful comments on earlier drafts.
Appendix
The relationship between the notation used by Rice (1984) and that used by Kidwell et al. (1977) is shown in Table A1. Rice (1984) separately considered the cases of invasion of a male-beneficial/female-detrimental allele, and of a female-beneficial/male-detrimental allele, in each case assigning the “resident” genotype a relative fitness of 1 in both sexes; homozygotes (or hemizygotes) of the sex benefited by the new allele had relative fitness of 1 + S, and homozygotes (or hemizygotes) for the sex harmed by the new allele had relative fitness of 1 − T. Kidwell et al.’s notation has the advantage that the selection coefficients sm and sf always apply to males and females, respectively, in contrast to S and T. Moreover, unlike S and T, sm and sf vary over the same range (0 < sf,sm ≤ 1) and are therefore more easily comparable. An advantage of Rice’s notation is that in the autosomal case, the conditions for invasion of the rare allele become simply S > T, i.e., that the new allele benefits one sex more than it harms the other sex. This result, and the invasion conditions for the X-linked case given by Rice (his expressions 3 and 6), can be reproduced by substituting the appropriate values from Table A1 into inequalities 1–3.
Table A1.
Relationship between parameter definitions of Kidwell et al. (1977) and Rice (1984).
| Kidwell et al. (1977) |
Rice (1984)
|
|
|---|---|---|
| Rare allele favored in males | Rare allele favored in females | |
| sf | T | S/(1 + S) |
| sm | S/(1 + S) | T |
| hf | h | 1 − h |
| hm | 1 − h | h |
LITERATURE CITED
- Albert AYK, Otto SP. Sexual selection can resolve sex-linked sexual antagonism. Science. 2005;310:119–121. doi: 10.1126/science.1115328. [DOI] [PubMed] [Google Scholar]
- Basolo AL. Genetic linkage and color polymorphism in the Southern Platyfish (Xiphophorus maculatus): a model system for studies of color pattern evolution. Zebrafish. 2006;3:65–83. doi: 10.1089/zeb.2006.3.65. [DOI] [PubMed] [Google Scholar]
- Bonduriansky R, Chenoweth C. Intralocus sexual conflict. TREE. 2009;24:280–288. doi: 10.1016/j.tree.2008.12.005. [DOI] [PubMed] [Google Scholar]
- Brommer JE, Kirkpatrick M, Qvarnström A, Gustafsson L. The intersexual genetic correlation for lifetime fitness in the wild and its implications for sexual selection. PLoS ONE. 2007;8:e744. doi: 10.1371/journal.pone.0000744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calsbeek R, Sinervo B. Within-clutch variation in offspring sex determined by differences in sire body size: cryptic mate choice in the wild. J Evol Biol. 2004;17:464–470. doi: 10.1046/j.1420-9101.2003.00665.x. [DOI] [PubMed] [Google Scholar]
- Charlesworth B, Hughes KA. The maintenance of genetic variation in life history traits. In: Singh RS, Krimbas CB, editors. Evolutionary Genetics from Molecules to Morphology. Cambridge University Press; Cambridge, UK: 1999. pp. 369–392. [Google Scholar]
- Chippindale AK, Gibson JR, Rice WR. Negative genetic correlation for adult fitness between sexes reveals ontogenetic conflict in Drosophila. Proc Natl Acad Sci (US) 2001;98:1671–1675. doi: 10.1073/pnas.041378098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Connallon T, Jakubowski E. Association between sex ratio distortion and sexually antagonistic fitness consequences of female choice. Evolution. 2009;63:2179–2183. doi: 10.1111/j.1558-5646.2009.00692.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Day T, Bonduriansky R. Intralocus sexual conflict can drive the evolution of genomic imprinting. Genetics. 2004;167:1537–1546. doi: 10.1534/genetics.103.026211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delcourt M, Blows MW, Rundle HD. Sexually antagonistic genetic variance for fitness in an ancestral and a novel environment. Proc R Soc Lond B. 2009;276:2009–2014. doi: 10.1098/rspb.2008.1459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellegren H, Parsch J. The evolution of sex-biased genes and sex-biased gene expression. Nat Rev Genet. 2007;8:689–698. doi: 10.1038/nrg2167. [DOI] [PubMed] [Google Scholar]
- Fedorka KM, Mousseau TA. Female mating bias results in conflicting sex-specific offspring fitness. Nature. 2004;429:65–67. doi: 10.1038/nature02492. [DOI] [PubMed] [Google Scholar]
- Foerster K, Coulson T, Sheldon BC, Pemberton JM, Clutton-Brock TH, Kruek LEB. Sexually antagonistic genetic variation for fitness in red deer. Nature. 2007;447:1107–1110. doi: 10.1038/nature05912. [DOI] [PubMed] [Google Scholar]
- Gavrilets S, Rice WR. Genetic models of homosexuality: generating testable predictions. Proc Roy Soc Lond B. 2006;273:3031–3038. doi: 10.1098/rspb.2006.3684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geber MA, Griffen LR. Inheritance and natural selection on functional traits. Int J Plant Sci. 2003;164:S21–S42. [Google Scholar]
- Gibson JR, Chippindale AK, Rice WR. The X chromosome is a hot spot for sexually antagonistic fitness variation. Proc R Soc Lond B. 2002;269:499–505. doi: 10.1098/rspb.2001.1863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedrick PW. Genetics of Populations. 2. Jones and Bartlett; Sudbury, MA: 2000. [Google Scholar]
- Houle D. Comparing evolvability and variability of quantitative traits. Genetics. 1992;130:195–204. doi: 10.1093/genetics/130.1.195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Houle D. How should we explain variation in the genetic variance of traits? Genetica. 1998;102/103:241–253. [PubMed] [Google Scholar]
- Hughes KA, Rodd FH, Reznick DN. Genetic and environmental effects on secondary sex traits in guppies (Poecilia reticulata) J Evol Biol. 2005;18:35–45. doi: 10.1111/j.1420-9101.2004.00806.x. [DOI] [PubMed] [Google Scholar]
- Kidwell JF, Clegg MT, Stewart FM, Prout T. Regions of stable equilibria for models of differential selection in the two sexes under random mating. Genetics. 1977;85:171–183. doi: 10.1093/genetics/85.1.171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindholm AK, Brooks R, Breden F. Extreme polymorphism in a Y-linked sexually selected trait. Heredity. 2004;92:156–162. doi: 10.1038/sj.hdy.6800386. [DOI] [PubMed] [Google Scholar]
- Meyer K. Likelihood calculations to evaluate experimental designs to estimate genetic variances. Heredity. 2008;101:212–221. doi: 10.1038/hdy.2008.46. [DOI] [PubMed] [Google Scholar]
- Mousseau TA, Roff DA. Natural selection and the heritability of fitness components. Heredity. 1987;59:181–197. doi: 10.1038/hdy.1987.113. [DOI] [PubMed] [Google Scholar]
- Pischedda A, Chippindale AK. Intralocus sexual conflict diminishes the benefits of sexual selection. PLoS Biology. 2006;4:e356. doi: 10.1371/journal.pbio.0040356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poissant J, Wilson AJ, Festa-Bianchet M, Hogg JT, Coltman DW. Quantitative genetics and sex-specific selection on sexually dimorphic traits in bighorn sheep. Proc R Soc Lond B. 2008;275:623–628. doi: 10.1098/rspb.2007.1361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prasad NG, Bedhomme S, Day T, Chippindale AK. An evolutionary cost of separate genders revealed by male-limited evolution. Am Nat. 2007;169:29–37. doi: 10.1086/509941. [DOI] [PubMed] [Google Scholar]
- Rice WR. Sex chromosomes and the evolution of sexual dimorphism. Evolution. 1984;38:735–742. doi: 10.1111/j.1558-5646.1984.tb00346.x. [DOI] [PubMed] [Google Scholar]
- Rice WR. The accumulation of sexually antagonistic genes as a selective agent promoting the evolution of reduced recombination between primitive sex chromosomes. Evolution. 1987;41:911–914. doi: 10.1111/j.1558-5646.1987.tb05864.x. [DOI] [PubMed] [Google Scholar]
- Roff DA, Mousseau TA. Quantitative genetics and fitness: Lessons from Drosophila. Heredity. 1987;58:103–118. doi: 10.1038/hdy.1987.15. [DOI] [PubMed] [Google Scholar]
- Vicoso B, Charlesworth B. Evolution on the X chromosome: unusual patterns and processes. Nat Rev Genet. 2006;7:645–653. doi: 10.1038/nrg1914. [DOI] [PubMed] [Google Scholar]
- Vicoso B, Charlesworth B. The deficit of male-biased genes on the D. melanogaster X chromosome is expression-dependent: a consequence of dosage compensation? J Mol Evol. 2009;68:576–583. doi: 10.1007/s00239-009-9235-4. [DOI] [PubMed] [Google Scholar]