

Abstract
VIPP1 protein in photosynthetic organisms is homologous to bacterial PspA, which protects plasma membrane integrity upon stresses. Despite the proposed role of VIPP1 in thylakoid biogenesis, its precise function remains unclear. Recently, our in-depth analysis of Arabidopsis vipp1 mutants revealed VIPP1’s involvement in the maintenance of chloroplast envelopes. Chloroplasts in intact vipp1 leaves exhibited spherical balloon-like morphology, which resulted from osmotic stress across envelopes. In fact, observation of VIPP1 fused to green fluorescence protein in vivo revealed that most VIPP1 is localized as a lattice-like macro complex attached along with the envelope. Because of the proposed function in thylakoids, we examined whether vipp1 also exhibited altered morphologies in thylakoids. Results show that thylakoid morphologies were detected irregularly, but vipp1 chloroplasts retained normal-appearing grana stacks. We infer that VIPP1 might influence thylakoids as well as envelopes, but that it is not involved directly in thylakoid membrane formation.
Keywords: Arabidopsis, PspA, chloroplast, envelope membrane, grana, osmotic stress, thylakoid biogenesis
Text
Biomembranes of chloroplasts have been proposed as a prime site of vulnerability to heat and cold stresses in plants.1,2 In general, such abiotic stresses induce damage in any biomembrane and ultimately engender disturbance in the membrane potential.3,4 Because of the presence of multiple membrane systems (outer and inner envelopes and thylakoid membrane), maintenance of membrane integrity within chloroplasts is critically important for plant growth.5,6 However, little has been documented about the mechanism(s) controlling plastid membrane integrity. Chloroplastic biomembranes, when ruptured, are presumably resealed spontaneously. Membrane maintenance has therefore been regarded as a physical process rather than a biochemical reaction. Nevertheless, accumulated evidence in various organisms implies that resealing of the plasma membrane is conducted via a dynamic and complex mechanism that requires novel cytoplasmic factors.7-9 It is reasonable to assume that organelles have similar mechanisms to protect biomembranes from damage.
Our recent work provided insight into a mechanism of plastid envelope maintenance in which Vesicle-Inducing Protein in Plastids1 (VIPP1) plays a pivotal role.10 Although VIPP1 has been originally identified as a factor involved in thylakoid biogenesis, its precise role remains unclear.11,12 VIPP1 is homologous to Phage-shock Protein A (PspA) in Escherichia coli, which is involved in plasma membrane integrity,11-14 and which was shown to recover the defective proton leakage when expressed in E. coli pspA mutant.10 Results showed that the Arabidopsis vipp1 mutants have globular, swollen, balloon-like chloroplasts representing increased osmotic stress of plastid envelope. A series of experiments, including the expression of VIPP1 fused to green fluorescent protein (VIPP1-GFP) in vipp1 mutants, demonstrated that the lattice-like macro complex of VIPP1 acts as scaffold to facilitate the resealing of osmotically damaged chloroplast envelope.10 It is particularly interesting that the dynamic movement of VIPP1-GFP along with envelopes was detectable, which was correlated with osmotic stress (presented as Supplemental Movies). Based on these observations, we concluded that VIPP1 is necessary for plastid envelope maintenance. The question remains as to whether VIPP1 is also involved in thylakoid membrane as originally proposed. Here, we characterize the morphological alteration of thylakoids in vipp1 knock-down mutants (vipp1-kd).
First, the thylakoid ultrastructure in fixed leaf tissues from wild-type Columbia (Col) and vipp1-kd was examined using electron microscopy (Fig. 1). In addition to the balloon-like structure, vipp1-kd chloroplasts exhibited irregular distorted thylakoids. Although thylakoids in Col were abundant in granal stacks that developed uniformly within the chloroplast (Fig. 1A), vipp1-kd showed distorted thylakoids that appeared to segregate into either well-stacked grana or single-membrane regions such as stroma thylakoid (Fig. 1B). In an extreme case, some vipp1-kd chloroplasts contained only several stacks that were tightly clustered (Fig. 1C). It is noteworthy that the clustered granal stacks frequently contained a large lumenal area (Fig. 1B, indicated by an arrow) that was similar to swollen stroma in the balloon-like chloroplasts and which might represent osmotic stress of thylakoid membranes. A similar fluffy structure of thylakoids has also been observed in chloroplasts suffered from water-osmotic stress or high/low temperature,3,15,16 implying that VIPP1 is involved in the maintenance of thylakoid membranes as well as of envelopes.

Figure 1. Chloroplasts in wild-type and vipp1-kd mutant examined using transmission electron microscopy. Chloroplast ultrastructures of Col (A) and vipp1-kd (B and C) mutants were observed using transmission electron microscopy. Extra lumen space (B, red arrow) was detected using electron micrography in vipp1-kd mutants. Furthermore, some larger blocks of mutlilayer membrane structures were found in vipp1-kd chloroplast (C). Bars show 1.0 µm.
In addition to these data obtained from electron microscopy, observation of chlorophyll fluorescence with fresh tissue supported the existence of altered grana-like structures (Fig. 2A). Protoplasts prepared from vipp1-kd contained balloon-like chloroplasts in which chlorophyll fluorescence signals tended to form a larger cluster. To test whether the grana stacks observed in vipp1-kd indeed retained functional thylakoid membranes, we attempted to detect light-harvesting complexes of PSII (LHCII). Blue-native gel electrophoresis of solubilized thylakoid membranes indicated that LHCII trimer had accumulated significantly in both Col and vipp1-kd mutant (Fig. 2B). Accumulation of LHCII was confirmed further by SDS-PAGE and subsequent western blot analysis (Fig. 2C). We concluded that the reduction of VIPP1 engenders altered thylakoid morphology, raising the possibility that VIPP1 is a multifunctional protein that affects both envelope and thylakoid membranes. However, the morphological alteration in thylakoids was not a simple reduction but rather a reorganization of the functional thylakoid membranes. Although further analysis is needed, experimental data obtained from our laboratory do not support the model that VIPP1 is involved in the direct formation of thylakoid membranes. Consistent with this inference, recent observations from other laboratories suggest that VIPP1 affects thylakoid formation indirectly through protein complex formation and maintenance, or through protein import across thylakoid membranes.17,18 These data are favorable to a scenario in which VIPP1 acts to protect membranes.
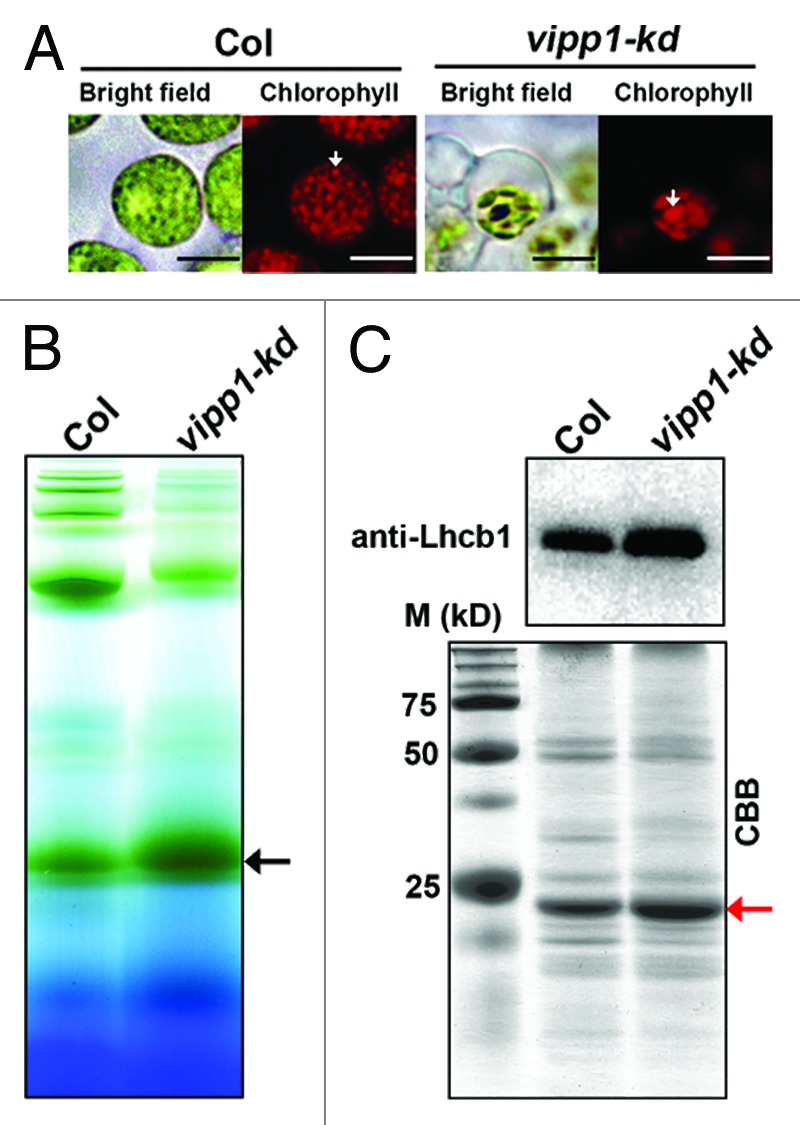
Figure 2. Accumulation of LHCII in vipp-kd mutant. (A) Chloroplast images of Col and vipp-kd protoplast photographed using fluorescence microscopy. White arrows indicate grana thylakoids. Bars show 10 µm. (B) Photosynthetic supercomplexes located on thylakoid membrane were analyzed using blue native gel. The thylakoid was extracted from the leaves of Col and vipp-kd mutant and incubated with 1% n-dodecyl-β-maltoside (DM) for 1.0 h on ice. Samples were normalized by chlorophyll content and were loaded to 0.75-mm-thick 5–13.5% acrylamide gradient gels. The black arrow indicates the LHCII trimer location. (C) The level of Lhcb1 protein in Col and vipp-kd. Upper panel, western blot probed with the antibody against Lhcb1. Lower panel, a Coomassie-stained gel image is shown as loading control. The red arrow indicates the Lhcb1 position in SDS PAGE.
Acknowledgments
This work was supported by the Japan Science and Technology Agency (Core Research for Evolutional Science and Technology, to W.S.) and by Oohara Foundation (to W.S.).
Glossary
Abbreviation:
- Col
wild-type Columbia
- DGDG
digalactosyldiacylglycerol
- GFP
Green Fluorescent Protein
- LHCII
Light Harvesting Complexes of PSII
- MGDG
monogalactosyldiacylglycerol
- PspA
Phage-shock Protein A
- VIPP1
Vesicle-Inducing Protein in Plastids1
- vipp1-kd
vipp1 knock down mutant
Footnotes
Previously published online: www.landesbioscience.com/journals/psb/article/22860
References
- 1.Quinn PJ. Regulation of membrane fluidity in plants. In: Aloia RC, Curtain CC, Gordon LM, ed. Advances in Membrane Fluidity, Physiological Regulation of Membrane Fluidity. Alan R. Liss, Inc., New York. 1988: pp 293-321. [Google Scholar]
- 2.Vigh L, Los DA, Horváth I, Murata N. The primary signal in the biological perception of temperature: Pd-catalyzed hydrogenation of membrane lipids stimulated the expression of the desA gene in Synechocystis PCC6803. Proc Natl Acad Sci U S A. 1993;90:9090–4. doi: 10.1073/pnas.90.19.9090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Heber U. Freezing injury and uncoupling of phosphorylation from electron transport in chloroplasts. Plant Physiol. 1967;42:1343–50. doi: 10.1104/pp.42.10.1343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hincha DK, Oliver AE, Crowe JH. The effects of chloroplast lipids on the stability of liposomes during freezing and drying. Biochim Biophys Acta. 1998;1368:150–60. doi: 10.1016/S0005-2736(97)00204-6. [DOI] [PubMed] [Google Scholar]
- 5.Uemura M, Joseph RA, Steponkus PL. Cold Acclimation of Arabidopsis thaliana (Effect on Plasma Membrane Lipid Composition and Freeze-Induced Lesions) Plant Physiol. 1995;109:15–30. doi: 10.1104/pp.109.1.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Thomashow MF. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu Rev Plant Physiol Plant Mol Biol. 1999;50:571–99. doi: 10.1146/annurev.arplant.50.1.571. [DOI] [PubMed] [Google Scholar]
- 7.McNeil PL, Kirchhausen T. An emergency response team for membrane repair. Nat Rev Mol Cell Biol. 2005;6:499–505. doi: 10.1038/nrm1665. [DOI] [PubMed] [Google Scholar]
- 8.Schapire AL, Voigt B, Jasik J, Rosado A, Lopez-Cobollo R, Menzel D, et al. Arabidopsis synaptotagmin 1 is required for the maintenance of plasma membrane integrity and cell viability. Plant Cell. 2008;20:3374–88. doi: 10.1105/tpc.108.063859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yamazaki T, Kawamura Y, Minami A, Uemura M. Calcium-dependent freezing tolerance in Arabidopsis involves membrane resealing via synaptotagmin SYT1. Plant Cell. 2008;20:3389–404. doi: 10.1105/tpc.108.062679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhang L, Kato Y, Otters S, Vothknecht UC, Sakamoto W. Essential role of VIPP1 in chloroplast envelope maintenance in Arabidopsis. Plant Cell. 2012;24:3695–707. doi: 10.1105/tpc.112.103606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kroll D, Meierhoff K, Bechtold N, Kinoshita M, Westphal S, Vothknecht UC, et al. VIPP1, a nuclear gene of Arabidopsis thaliana essential for thylakoid membrane formation. Proc Natl Acad Sci U S A. 2001;98:4238–42. doi: 10.1073/pnas.061500998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Westphal S, Heins L, Soll J, Vothknecht UC. Vipp1 deletion mutant of Synechocystis: a connection between bacterial phage shock and thylakoid biogenesis? Proc Natl Acad Sci U S A. 2001;98:4243–8. doi: 10.1073/pnas.061501198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Brissette JL, Russel M, Weiner L, Model P. Phage shock protein, a stress protein of Escherichia coli. Proc Natl Acad Sci U S A. 1990;87:862–6. doi: 10.1073/pnas.87.3.862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Otters S, Braun P, Hubner J, Wanner G, Vothknecht UC, Chigri F. The first α-helical domain of the vesicle-inducing protein in plastids 1 promotes oligomerization and lipid binding. Planta. 2012 doi: 10.1007/s00425-012-1772-1. In Press. [DOI] [PubMed] [Google Scholar]
- 15.Ma SF, Lin CY, Chen YM. Comparative studies of chilling stress on alternations of chloroplast ultrastructure and protein synthesis in the leaves of chilling-sensitive (mungbean) and -insensitive (pea) seedlings. Bot Bull Acad Sinica (Taiwan) 1990;31:263–72. [Google Scholar]
- 16.Ristic Z, Cass DD. Chloroplast structure after water and high-temperature stress in two lines of maize that differ in endogenous levels of abscisic acid. Int J Plant Sci. 1992;153:186–96. doi: 10.1086/297021. [DOI] [Google Scholar]
- 17.Nordhues A, Schöttler MA, Unger AK, Geimer S, Schönfelder S, Schmollinger S, et al. Evidence for a role of VIPP1 in the structural organization of the photosynthetic apparatus in Chlamydomonas. Plant Cell. 2012;24:637–59. doi: 10.1105/tpc.111.092692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lo SM, Theg SM. Role of vesicle-inducing protein in plastids 1 in cpTat transport at the thylakoid. Plant J. 2012;71:656–68. doi: 10.1111/j.1365-313X.2012.05020.x. [DOI] [PubMed] [Google Scholar]