

Abstract
Background & objectives:
Cytoskeletal proteins are deregulated during oxidative stress and cataract formation. However, estrogen which protects against cataract formation and harmful effects of oxidative stress has not been tested on the cytoskeleton of lens epithelial cells (LECs). The current study was undertaken to assess if the protection rendered to LECs by estrogen was mediated by preserving the cytoskeletal proteins.
Methods:
Oxidative stress was induced by 50 μM of H2O2 in cultured goat LECs (gLECs) and effect of 1 μM 17β-estradiol (E2) was tested. After treatment, morphological analysis of cells was carried out using haematoxylin-eosin staining and cell density was also quantified. Cell viability was determined using Hoechst (Ho), YO-Pro (YP) and propidium iodide (PI). F-actin and vimentin were localized using phalloidin and anti-vimentin antibody, respectively, and viewed under fluorescence microscopy. Vimentin was further analysed at protein level by Western blotting.
Results:
H2O2 led to increased condensation of nucleus, cell death and apoptosis but these were prevented with pre- and co-treatment of E2 with increase in cell viability (P<0.001). E2 also prevented H2O2 mediated depolymerization of cytoskeleton but was not able to reverse the changes when given after induction of oxidative stress.
Interpretation & conclusions:
Our findings showed that E2 helped in preventing deteriorating effect of H2O2, inhibited cell death, apoptosis and depolymerisation of cytoskeletal proteins in LECs. However, the exact mechanism by which estrogen renders this protection to cytoskeleton of lens epithelial cells remains to be determined.
Keywords: Actin, colon, estrogen, lens epithelial cells, oxidative stress, vimentin
Lens epithelial cells (LECs) are metabolically the most active part of the lens and are particularly prone to oxidative damage, which in turn is recognized as a major risk factor for age related cataract1. Age-related cataract associated with oxidative stress exhibits sexual dichotomy and affects more females than males. Increased prevalence of cataract in postmenopausal women compared to age-matched men, leads to the hypothesis that the absence of estrogen probably contributes to the increased risk2. Freeman et al3, have shown a role of estrogen in protecting the lens against cataract. These results are substantiated by tissue culture and animal model studies in which importance of estrogen in diminishing or preventing cataract has been documented4,5. Studies confirming the presence of estrogen receptors in lens, further support these findings6. Using tissue culture of human LECs (HLECs), estrogen has been shown to be a powerful antioxidant7, cytoprotectant that prevents mitochondrial depolarization, maintain cell viability by induction of anti-apoptotic proteins and protective signalling pathways during oxidative stress8,9.
The cytoskeletal network consisting of actin and vimentin, plays an important role in several vital cellular functions and is considered the site where early impact of oxidative stress is seen10. Disruption of cytoskeletal proteins on oxidative damage has been implicated in different cell types11–13. In the ocular tissues like retinal pigment epithelium and trabecular meshwork, oxidative stimulus leads to reorganization of actin and vimentin14,15. Deregulation of vimentin and tubulin has been reported using CD5A HLECs following oxidative stress16. Also, loss or degradation of cytoskeletal proteins during cataract formation has been well documented17,18. Estrogen also regulates the cytoskeleton and has an impact on its remodelling in other cell types19,20. However, the effects of estrogen on cytoskeletal arrangement in LECs during oxidative insult are not studied. The current study was thus designed to elucidate the effect of oxidative stress on cytoskeletal proteins; fibrous actin (F-actin) and vimentin, and to understand if protection afforded to LECs by estrogen during oxidative stress is attained by maintaining the integrity of these cytoskeletal proteins.
Material & Methods
Primary culture of goat LECs: The study was conducted at Iladevi Cataract and Intraocular Lens Research Centre, Ahmedabad, Gujarat. The study protocol was approved by the institutional ethical committee. Goat eyes (n = 4) were collected from slaughter house within 30 min of death. After sterilization with betadine, eyes were dissected and the lens was collected in phosphate buffer saline (PBS) (Sigma, St. Louis, USA). A small incision was made on the lateral side of the lens and the lens capsule with the attached epithelium was excised. The capsule was placed in 1 ml of 0.05 per cent trypsin-EDTA (Sigma, St. Louis, USA) for 15 min and observed under inverted microscope till all cells got detached. Once the cells were detached, 200 μl of foetal bovine serum (FBS) (Gibco, Invitrogen, Eugene, USA) was added and the tube was centrifuged for 10 min at 480xg. Supernatant was discarded and pellets obtained from the capsules were resuspended in 10 ml Eagle's minimal essential media (MEM) (Sigma, St. Louis, USA) supplemented with 10 per cent FBS. Equal number of cell (n=6000), were seeded on coverslips in each well containing 600 μl of MEM-10 per cent FBS. The culture plate was incubated at 37°C and 5 per cent CO2 - 95 per cent air. Medium was changed every third day. The LECs culture was allowed to achieve 70 per cent confluence after which the cells were serum starved. The following day cells were exposed to oxidative stress and estrogen.
Treatment of hydrogen peroxide (H2O2) and estradiol (E2): To study effects of estrogen, 17β-Estradiol (E2) (Sigma, St. Louis, USA) was used as it is the main form of estrogen. Oxidative stress was induced using 30 per cent hydrogen peroxide (H2O2) (MERCK India Ltd, Mumbai, India). For treatment, H2O2 and E2 were used at a final concentration of 50 and 1 μM, respectively8. The wells were divided into six groups as follows: C: control, with no treatment; H: Only H2O2 treatment for the first 24 h, followed by MEM; E: Only E2 treatment for the first 24 h, followed by MEM; H-E: H2O2 treatment for first 24 h, followed by E2 treatment for next 24 h; E-H: E2 treatment for first 24 h, followed by H2O2 treatment for next 24 h; and HE: Co-treatment of H2O2 with E2 for the first 24 h, followed by MEM. Each set of treatment was in duplicates and repeated three times.
Histological studies: For histological analysis, cells were fixed in 2 per cent paraformaldehyde (PFA) (MERCK India Ltd, Mumbai, India), washed with PBS and then stained with haematoxylin for nuclear staining and eosin for cytoplasmic counterstaining.
Analysis of cell density: Cell density was analyzed after fixing the cells in 2 per cent PFA, washing with PBS and then staining the nuclei with 4’,6-diamidino-2-phenylindole (DAPI) (Molecular Probes, Invitrogen, Eugene, USA). Cells were visualized using fluorescent microscope fitted with UV filter and images of cells from 10 randomly selected areas for each group were taken using Cohu cool CCD camera. These images were analyzed for cell density using 4.1 v, Biovis Image software (Expertvision, India).
Analysis of cell viability: Cell viability was assessed on the basis of dye permeability using combination of fluorescent dyes; Hoechst 33342 (Ho), YO-PRO-1 (YP) and propidium iodide (PI) (Molecular Probes, Invitrogen, Eugene, USA). After completion of treatment, cells grown on coverslips were immediately incubated in PBS containing 4 μM Ho, 0.2 μM YP and 0.5 μM PI for 15 min at 37°C. The solution was removed and cells were washed in PBS, mounted and observed under fluorescence microscope (Axioskope II, Carl Zeiss, Germany). Ten random areas were selected from each group. Ho, YP and PI positive nuclei (blue, green and red, respectively) were observed using appropriate filter sets. Number of Ho, YP and PI positive nuclei were counted in each image and represented as nuclei/mm2 area using image analysis software (4.1 v Biovis, Expertvision, India). These counts were used to calculate the relative proportion of dead and apoptotic cells. For quantification of cell mortality, YP and PI positive cells were expressed as a percentage of Ho positive cells.
Immunohistochemical localization of proteins: The cytoskeletal proteins; vimentin and F-actin, were localized using specific antibodies and fluorescence microscopy as previously described21. Briefly, the cells on the coverslips were washed with PBS and then fixed in 2 per cent PFA. After permeabilizing with 0.1 per cent triton-X 100 and 0.1 per cent saponin (Sigma, St Louis, USA), the samples were blocked using 1 per cent bovine serum albumin (BSA) in PBS for 30 min at 37°C. Cells were incubated with 1:200 diluted monoclonal mouse anti-vimentin antibody (Sigma, St Louis, USA) in PBS for one hour at 37°C. Following wash with PBS + tween-20 (PBST), samples were incubated with 1:200 diluted anti-mouse antibody tagged with Alexa Flour 488 (Molecular Probes, Invitrogen, Eugene, USA) and FITC conjugated phalloidin dye (Sigma, St Louis, USA) at 37°C for one hour. DAPI (Molecular Probes, Invitrogen, Eugene, USA) was used to counterstain the nuclei. Fluorescence microscopy (Axioskope II, Carl Zeiss, Germany) was used to visualize the proteins and images were documented using Cohu cool CCD camera.
Western blot analysis: Western blot analysis was carried out to measure protein expression of vimentin21. Cells were washed with PBS and then collected in cell lysis buffer (SDS, Sodium deoxycholate, Tris, NaCl and Triton-x). Total protein concentration was quantified by BCA assay using manufacturer's protocol (Pierce, Thermo Scientific, USA). 10 μg of total protein from each treatment group was electrophoresed in SDS-polyacrylamide gel and transferred onto nitrocellulose membrane. Membrane was blocked in 5 per cent skimmed milk in TBST for 1 h and then incubated with anti-vimentin antibody (Sigma, St Louis, USA) overnight at 4°C. After washing with TBS plus tween-20, the membrane was incubated with horse radish peroxidase conjugated secondary antibody. Protein bands were observed on addition of substrate and chromogen (H2O2 +DAB). The intensity of bands was quantified using UN-SCAN IT gel (v-6.1) (Silk Scientific Corporation, USA).
Statistical analysis: Analysis of variance (ANOVA) was used for the comparison of data sets. All data are reported as mean ± SE and P<0.05 was considered significant.
Results
Effect of H2O2 and E2 on LEC morphology: Oxidative stress can lead to several morphological alterations in LECs. These changes were visualized using haematoxylin-eosin staining. Cells were uniformly arranged in the control group, and had large nuclei with uniform staining. On treatment with H2O2, the nuclei of the cells underwent condensation and some cells even showed nuclear fragmentation, which is a characteristic of apoptotic cells. E2 treated group (E) had cells with morphology similar to that of control cells. Treatment of E2 following H2O2 (H-E) did not improve cell morphology and cells with condensed nuclei were observed. Pre-treatment of E2 (E-H), greatly reduced the number of cells with condensed nuclei. Simultaneous treatment of E2 with H2O2 (HE), showed a heterogeneous group of cells with both large as well as small cells and some with condensed nuclei (Fig. 1).
Fig. 1.
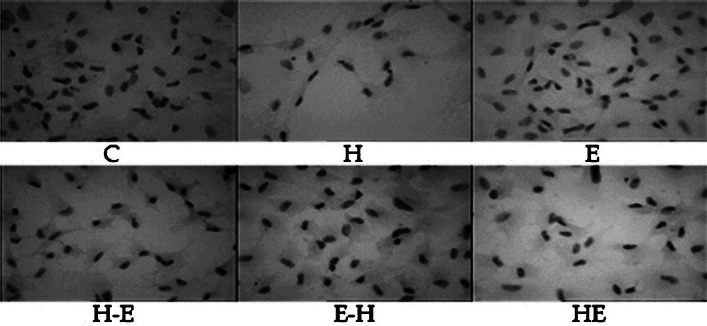
Haematoxylin-eosin stained goat lens epithelial cells (gLECs) of different treatment groups as viewed under light microscope with 10× objective. Groups; C = untreated cells, H = H2O2 treated, E = E2 treated, H-E = H2O2 treatment for the first 24 h followed by 24 h of E2 treatment, E-H = E2 treatment for first 24 h followed by H2O2 treatment, and HE = co-treatment of H2O2 and E2 for the first 24 h followed by MEM.
Effect of H2O2 and E2 on LEC density: After treatment with H2O2 (H), the cell density reduced significantly (P<0.001) compared to control, whereas the cell density was not altered on treatment of E2 alone (E). Although all three groups H-E, E-H and HE showed increase in cell density compared to that of H, increase in only E-H was statistically significant (P<0.05). These three groups showed a decrease in cell density when compared to E2 treatment alone (Fig. 2).
Fig. 2.
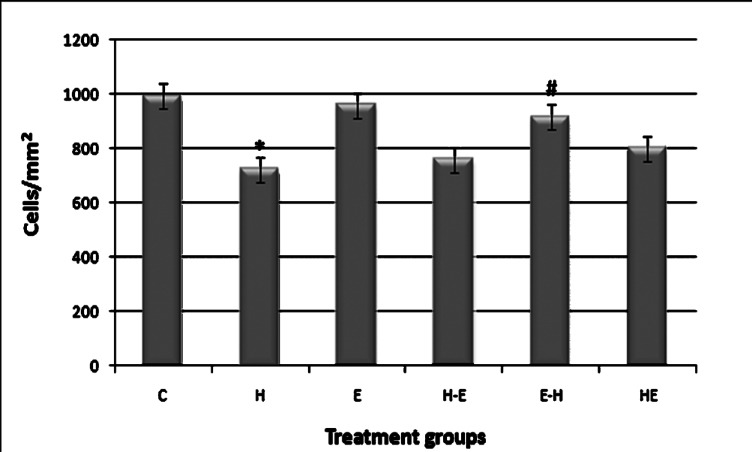
Cell density of gLECs in different treatment groups. Cell density was quantified by Biovis image analysis software. Groups as defined in Fig. 1. NS= non significant. *P<0.001 compared with control; #P <0.05 compared with H (H2O2 group). Values are mean ± SE (n=3).
Effect of H2O2 and E2 on viability of LEC: Cell viability was assessed by dye exclusion assay using Ho, YP and PI (Fig. 3). It should be noted that YP+ and PI+ cells indicate dead cells with damaged plasma membrane, through which these dyes can penetrate. YP- and PI- cells are live cells with intact plasma membranes and apoptotic cells are YP+ and PI- with altered plasma membrane.
Fig. 3.

Cell viability assessed by dye exclusion assay in different treatment groups. Each row depicts same microscopic field of the given treatment group taken with 5× objective. Column I shows Ho stained cells. Column II shows cells positive for YP and cells which labelled for PI are shown in column III. Groups as defined in Fig. 1.
Untreated cells (C) had very few positive nuclei for YP and PI, whereas the per cent of dead cells (YP+ and PI+) increased significantly (P<0.001) on induction of oxidative stress (H) (Fig. 4). but treatment of E2 helped in maintaining cell viability. Pre- or co-supplementation of E2 increased (P<0.001) per cent of viable cells (YP- and PI-), when compared to group H. Post-supplementation of estrogen (H-E) did not have significant protective effect. The number of apoptotic cells (YP+ and PI-) was the highest when only H2O2 treatment was given (H).
Fig. 4.
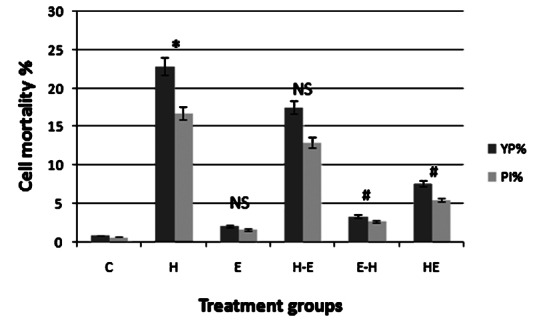
Cell mortality of gLECs in different treatment groups. Number of cells positive for YP and PI are expressed as a percentage of number of cells positive for Ho. Treatment group as defined in Fig. 1. Values are mean ± SE (n=3). NS, non significant. *P <0.001 compared with control; #P<0.001 compared with H.
Effect of H2O2 and E2 on cytoskeleton of LEC: As illustrated in Fig. 5, control group showed vimentin predominantly in filamentous form, usually without a pronounced perinuclear ring. This arrangement was lost when the cells were treated with H2O2; vimentin became diffused and showed a perinuclear arrangement, but control-like distribution was seen on treatment of E2. These cells in the E group showed vimentin filaments traversing throughout the cytoplasm of the cells. Treatment of E2 following oxidative stress (H-E) did not improve vimentin cytoskeleton which looked highly diffused and degraded. Pre-treatment with estrogen (E-H) helped in regaining fine as well as dense mass of vimentin filaments. In the group where co-treatment of E2 was given with H2O2 (HE), cells had heterogeneous vimentin; with some cells showing filamentous reticular network of vimentin while some showing a condensed layout with pronounced perinuclear arrangement.
Fig. 5.
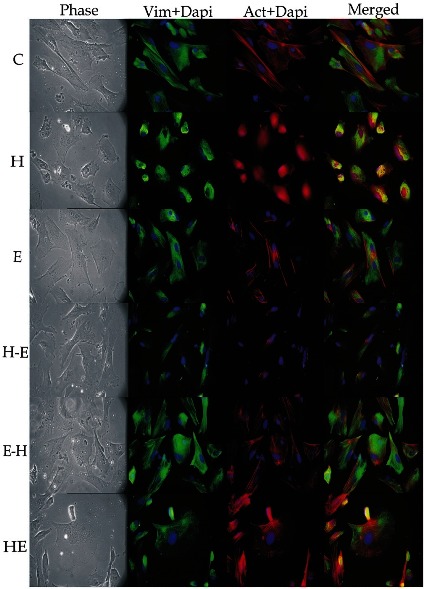
Immunolocalization of cytoskeletal proteins; vimentin and F-actin in different treatment groups with 40 × objective. Each row depicts same microscopic field of the given treatment group. Column I shows phase images of cells. Column II show cells positive for vimentin (green) and cells with F-actin (red) are shown in column III. Column IV shows merged images of column II and III. All the cells are counterstained with DAPI (blue).
Protein analysis by BCA showed no significant difference in quantity of total protein between control and treated groups. Immunoblotting analysis with vimentin specific antibodies revealed that the intensity of bands varied between different treatment groups and was more pronounced in the group where cells were treated with H2O2 alone, but this difference was not statistically significant (Fig. 6).
Fig. 6.
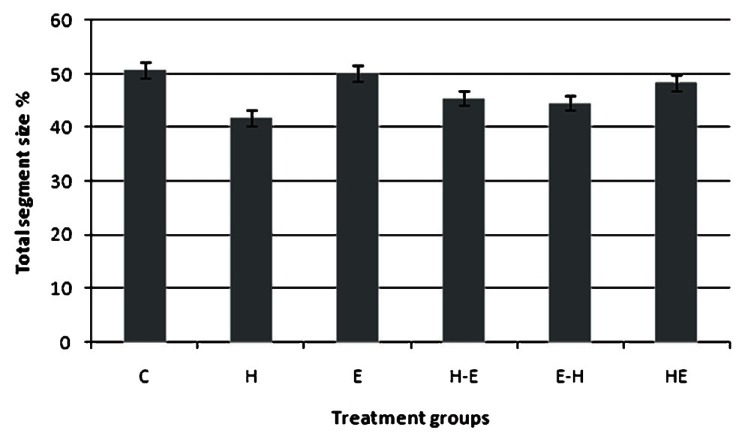
Western blot analysis of vimentin protein expression in gLECs after different treatments. The bands obtained were quantified using UN-SCAN-IT gel and the graph was plotted by comparing the per cent of total segment size in each treatment group. Groups as defined in Fig. 1. Values are mean ± SE (n=3).
In untreated cells, prominent actin stress fibres were present and also showed presence of some lamellipodia and filapodia like structures (protrusions) (Fig. 5). Major reorganization of actin cytoskeleton occurred on treatment of H2O2. During oxidative stress (H), depolymerisation of actin occurs and the protein gets arranged as dense condensation in the cells. In group E, actin localization was almost the same as that in control. Cells showed presence of dorsal stress fibres with cell edges showing strong actin staining. Estrogen treatment after exposure to oxidative stress (H-E) did not help in inhibition of actin depolymerization, save formation of a few stress fibres with overall diffused or disorganized actin filament staining. Pre-treatment with E2 (E-H) protected actin cytoskeleton and led to homogeneous distribution of actin with plenty of cytosolic stress fibres traversing along the length of the cell. In the HE group, heterogeneous distribution of actin was observed with some cells showing normal stress fibre pattern, some with polygonal network of actin stress fibres and some which showed diffused actin cytoskeleton.
Discussion
In this study, the effect of estrogen was assessed on H2O2 induced oxidative stress on two main cytoskeletal proteins; vimentin and fibrous-actin (F-actin), as these two proteins are altered during oxidative stress and in cataract formation11,12,17,18. gLECs were used due to unavailability of clear cadaver lenses. A final concentration of 1 μM E2 was chosen based on a previous report8. H2O2 concentration varies from 10-660 μM in cataract patients with mean concentration of 69 μM22. A physiological concentration of 50 μM H2O2 was employed to induce oxidative stress in this study8.
Oxidative stress leads to profound injury to a number of intracellular macromolecules, and ultimately leads to cell death. On carrying out cell viability assay by Ho-YP-PI, it was found that level of cell death was the least in control, followed by treatment group E. The highest level of cell death and apoptosis was observed on treatment of H2O2 (H) and increased LEC death on treatment of H2O2 is associated with cataract formation1,23. These results corroborated with other studies on LECs showing activation of caspases and induction of apoptosis upon treatment with H2O223,24.
Cystoskeletal protein, vimentin was found in different forms in all the groups, but with no significant difference in the level during oxidative stress. However, on induction of oxidative stress, major rearrangement of vimentin and actin was observed. The depolymerization of cytoskeletal proteins reported here during oxidative stress is well established in endothelial cells12. Loss and disruption of vimentin and actin are reported in in vitro models of oxidative stress using cortical neurons and in vivo cataract models11,17,18; although actin cytoskeleton has not been studied during oxidative stress using cultured LECs. Under oxidative insult the conventional intracellular antioxidant enzymes will be overburdened and this can possibly lead to major remodelling of the cytoskeleton. Our results corroborated with other study on LECs16 and with studies on other cell types using H2O2 induced oxidative stress13, in which cytoskeletal deregulation was observed. This study confirms cytoskeleton depolymerization on induction of oxidative stress and further strengthens the hypothesis that cytoskeleton proteins are directly or indirectly involved in response to oxidative stress. Although contradictory results showing polymerization of these proteins in endothelial cells exist25, but these differences can be ascribed to the basic difference in cells used and culture conditions used for specific cell type.
In the present study, pre-treatment (E-H) and co-treatment (HE) of estrogen inhibited apoptosis in H2O2 treated cells, whereas, post-treatment (H-E) was not able to do the same. The decrease in cell apoptosis observed on treatment of E2 can possibly be attributed to either phosphorylation of HSP27 leading to anti-apoptotic pathway20 or activation of survival pathway MAPK during H2O2 induced oxidative stress9.
Our results showed that on treating with E2 (E), the cells were in similar morphology to those seen in control group (C). Both vimentin and actin were present in the filamentous form. Pre-treatment of E2 before H2O2 (E-H), inhibited depolymerisation of cytoskeleton possibly because prior exposure of cells to E2 would enhance the antioxidant status of the cell. This has also been shown in our previous work7. The other possibility of estrogen binding to its receptors and transmission of protective signals can also be considered26. Further, estrogen is considered to mediate its antioxidative properties both by the genomic mode of action as well as non-genomic pathway26. The non-genomic mode of action is rapid and takes place within minutes7,26. On subjecting the cells simultaneously with E2 and H2O2 (HE), filamentous cytoskeleton was found suggesting that E2 might be acting via the rapid non genomic pathway. Although the possibility of direct binding of the phenolic ring of estrogen to H2O227, which can nullify oxidative effect and thus help in maintaining the cytoskeleton integrity cannot be neglected. However, the exact mechanism is not clear and effect of estrogen is not uniform as some cells show early signs of oxidative damage. Estrogen was not able to reverse the cytoskeletal damage induced by prior treatment of H2O2 (H-E), confirming previous notion that changes induced by oxidative stress are not reversible1.
There is compelling evidence that the cytoskeleton supports cell survival and their disruption will activate caspases and lead to increased apoptosis, which could lead to further cytoskeleton degradation28. These results were replicated in LECs, where loss of actin stress fibres triggered apoptotic pathway and led to cell death29. On the other hand, apoptotic pathways directly triggered by oxidative stress by inducing caspase-8, -9, can cleave vimentin, which in turn promotes apoptosis13,24,30, whereas estrogen will activate anti-apoptotic pathways.
In conclusion, in the present study, induction of oxidative stress by H2O2 led to loss of cytoskeleton fibres and increased apoptosis and cell death, all of which were inhibited by pre- and co- treatment of estradiol. But it is difficult to predict if cytoskeleton deregulation during stress led to increased apoptosis or vice-versa. Estrogen helps in preventing the deteriorating effect of H2O2, inhibits depolymerization of cytoskeletal proteins and apoptosis of lens epithelial cells; which in turn may lead to development of cataract. This study thus highlights importance of estrogen in protecting against cataract. Further studies to identify the signalling pathways by which estrogen mediates this protection to LECs during oxidative stress are in progress.
Acknowledgment
The first author (DAG) thanks Iladevi Cataract and IOL Research Centre Trust, Ahmedabad, for funding the research work. Authors thank Ms. Khusboo Patel for technical assistance.
References
- 1.Spector A. Oxidative stress-induced cataract: mechanism of action. FASEB J. 1995;9:1173–82. [PubMed] [Google Scholar]
- 2.Klein BE, Klein R, Linton KL. Prevalence of age-related lens opacities in a population. The Beaver Dam Eye Study. Ophthalmology. 1992;99:546–52. doi: 10.1016/s0161-6420(92)31934-7. [DOI] [PubMed] [Google Scholar]
- 3.Freeman EE, Munoz B, Schein OD, West SK. Hormone replacement therapy and lens opacities: the Salisbury Eye Evaluation project. Arch Ophthalmol. 2001;119:1687–92. doi: 10.1001/archopht.119.11.1687. [DOI] [PubMed] [Google Scholar]
- 4.Bigsby RM, Cardenas H, Caperell-Grant A, Grubbs CJ. Protective effects of estrogen in a rat model of age-related cataracts. Proc Natl Acad Sci USA. 1999;96:9328–32. doi: 10.1073/pnas.96.16.9328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hales AM, Chamberlain CG, Murphy CR, McAvoy JW. Estrogen protects lenses against cataract induced by transforming growth factor-beta (TGFbeta) J Exp Med. 1997;185:273–80. doi: 10.1084/jem.185.2.273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gupta PD, Johar K, Sr, Nagpal K, Vasavada AR. Sex hormone receptors in the human eye. Surv Ophthalmol. 2005;50:274–84. doi: 10.1016/j.survophthal.2005.02.005. [DOI] [PubMed] [Google Scholar]
- 7.Gajjar D, Patel D, Alapure B, Praveen MR, Patel A, Johar KS, et al. Rapid action of oestradiol against hydrogen peroxide-induced oxidative stress in cataractous lens epithelium: an in vitro study. Eye (Lond) 2009;23:1456–63. doi: 10.1038/eye.2008.284. [DOI] [PubMed] [Google Scholar]
- 8.Moor AN, Gottipati S, Mallet RT, Sun J, Giblin FJ, Roque R, et al. A putative mitochondrial mechanism for antioxidative cytoprotection by 17beta-estradiol. Exp Eye Res. 2004;78:933–44. doi: 10.1016/j.exer.2004.01.001. [DOI] [PubMed] [Google Scholar]
- 9.Moor AN, Flynn JM, Gottipati S, Giblin FJ, Cammarata PR. 17beta-estradiol stimulates MAPK signaling pathway in human lens epithelial cell cultures preventing collapse of mitochondrial membrane potential during acute oxidative stress. Mitochondrion. 2005;5:235–47. doi: 10.1016/j.mito.2005.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Dalle-Donne I, Rossi R, Milzani A, Di Simplicio P, Colombo R. The actin cytoskeleton response to oxidants: from small heat shock protein phosphorylation to changes in the redox state of actin itself. Free Radic Biol Med. 2001;31:1624–32. doi: 10.1016/s0891-5849(01)00749-3. [DOI] [PubMed] [Google Scholar]
- 11.Allani PK, Sum T, Bhansali SG, Mukherjee SK, Sonee M. A comparative study of the effect of oxidative stress on the cytoskeleton in human cortical neurons. Toxicol Appl Pharmacol. 2004;196:29–36. doi: 10.1016/j.taap.2003.12.010. [DOI] [PubMed] [Google Scholar]
- 12.Valen G, Sonden A, Vaage J, Malm E, Kjellstrom BT. Hydrogen peroxide induces endothelial cell atypia and cytoskeleton depolymerization. Free Radic Biol Med. 1999;26:1480–8. doi: 10.1016/s0891-5849(99)00009-x. [DOI] [PubMed] [Google Scholar]
- 13.Chen X, Kang H, Zou F. Low concentration of GA activates a preconditioning response in HepG2 cells during oxidative stress-roles of Hsp90 and vimentin. Cell Stress Chaperones. 2009;14:381–9. doi: 10.1007/s12192-008-0092-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ho TC, Yang YC, Cheng HC, Wu AC, Chen SL, Tsao YP. Pigment epithelium-derived factor protects retinal pigment epithelium from oxidant-mediated barrier dysfunction. Biochem Biophys Res Commun. 2006;342:372–8. doi: 10.1016/j.bbrc.2006.01.164. [DOI] [PubMed] [Google Scholar]
- 15.Zhou L, Li Y, Yue BY. Oxidative stress affects cytoskeletal structure and cell-matrix interactions in cells from an ocular tissue: the trabecular meshwork. J Cell Physiol. 1999;180:182–9. doi: 10.1002/(SICI)1097-4652(199908)180:2<182::AID-JCP6>3.0.CO;2-X. [DOI] [PubMed] [Google Scholar]
- 16.Paron I, D’Elia A, D’Ambrosio C, Scaloni A, D’Aurizio F, Prescott A, et al. A proteomic approach to identify early molecular targets of oxidative stress in human epithelial lens cells. Biochem J. 2004;378:929–37. doi: 10.1042/BJ20031190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Clark JI, Matsushima H, David LL, Clark JM. Lens cytoskeleton and transparency: a model. Eye (Lond) 1999;13:417–24. doi: 10.1038/eye.1999.116. [DOI] [PubMed] [Google Scholar]
- 18.Matsushima H, David LL, Hiraoka T, Clark JI. Loss of cytoskeletal proteins and lens cell opacification in the selenite cataract model. Exp Eye Res. 1997;64:387–95. doi: 10.1006/exer.1996.0220. [DOI] [PubMed] [Google Scholar]
- 19.Kramar EA, Chen LY, Brandon NJ, Rex CS, Liu F, Gall CM, et al. Cytoskeletal changes underlie estrogen's acute effects on synaptic transmission and plasticity. J Neurosci. 2009;29:12982–93. doi: 10.1523/JNEUROSCI.3059-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Razandi M, Pedram A, Levin ER. Estrogen signals to the preservation of endothelial cell form and function. J Biol Chem. 2000;275:38540–6. doi: 10.1074/jbc.M007555200. [DOI] [PubMed] [Google Scholar]
- 21.Lei W, Zhang K, Pan X, Hu Y, Wang D, Yuan X, et al. Histone deacetylase 1 is required for transforming growth factor-beta1-induced epithelial-mesenchymal transition. Int J Biochem Cell Biol. 2010;42:1489–97. doi: 10.1016/j.biocel.2010.05.006. [DOI] [PubMed] [Google Scholar]
- 22.Spector A, Garner WH. Hydrogen peroxide and human cataract. Exp Eye Res. 1981;33:673–81. doi: 10.1016/s0014-4835(81)80107-8. [DOI] [PubMed] [Google Scholar]
- 23.Li WC, Kuszak JR, Dunn K, Wang RR, Ma W, Wang GM, et al. Lens epithelial cell apoptosis appears to be a common cellular basis for non-congenital cataract development in humans and animals. J Cell Biol. 1995;130:169–81. doi: 10.1083/jcb.130.1.169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Choudhary S, Zhang W, Zhou F, Campbell GA, Chan LL, Thompson EB, et al. Cellular lipid peroxidation end-products induce apoptosis in human lens epithelial cells. Free Radic Biol Med. 2002;32:360–9. doi: 10.1016/s0891-5849(01)00810-3. [DOI] [PubMed] [Google Scholar]
- 25.Zhao Y, Davis HW. Hydrogen peroxide-induced cytoskeletal rearrangement in cultured pulmonary endothelial cells. J Cell Physiol. 1998;174:370–9. doi: 10.1002/(SICI)1097-4652(199803)174:3<370::AID-JCP11>3.0.CO;2-D. [DOI] [PubMed] [Google Scholar]
- 26.Levin ER. Integration of the extranuclear and nuclear actions of estrogen. Mol Endocrinol. 2005;19:1951–9. doi: 10.1210/me.2004-0390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Behl C, Skutella T, Lezoualc’h F, Post A, Widmann M, Newton CJ, et al. Neuroprotection against oxidative stress by estrogens: structure-activity relationship. Mol Pharmacol. 1997;51:535–41. [PubMed] [Google Scholar]
- 28.Flusberg DA, Numaguchi Y, Ingber DE. Cooperative control of Akt phosphorylation, bcl-2 expression, and apoptosis by cytoskeletal microfilaments and microtubules in capillary endothelial cells. Mol Biol Cell. 2001;12:3087–94. doi: 10.1091/mbc.12.10.3087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Weber GF, Menko AS. Actin filament organization regulates the induction of lens cell differentiation and survival. Dev Biol. 2006;295:714–29. doi: 10.1016/j.ydbio.2006.03.056. [DOI] [PubMed] [Google Scholar]
- 30.Byun Y, Chen F, Chang R, Trivedi M, Green KJ, Cryns VL. Caspase cleavage of vimentin disrupts intermediate filaments and promotes apoptosis. Cell Death Differ. 2001;8:443–50. doi: 10.1038/sj.cdd.4400840. [DOI] [PubMed] [Google Scholar]