

Abstract
The transcription factor ΔFosB and the brain-enriched calcium/calmodulin-dependent protein kinase II (CaMKIIα) are induced in the nucleus accumbens (NAc) by chronic exposure to cocaine or other psychostimulant drugs of abuse, in which the two proteins mediate sensitized drug responses. Although ΔFosB and CaMKIIα both regulate AMPA glutamate receptor expression and function in NAc, dendritic spine formation on NAc medium spiny neurons (MSNs), and locomotor sensitization to cocaine, no direct link between these molecules has to date been explored. Here, we demonstrate that ΔFosB is phosphorylated by CaMKIIα at the protein-stabilizing Ser27 and that CaMKII is required for the cocaine-mediated accumulation of ΔFosB in rat NAc. Conversely, we show that ΔFosB is both necessary and sufficient for cocaine induction of CaMKIIα gene expression in vivo, an effect selective for D1-type MSNs in the NAc shell subregion. Furthermore, induction of dendritic spines on NAc MSNs and increased behavioral responsiveness to cocaine after NAc overexpression of ΔFosB are both CaMKII dependent. Importantly, we demonstrate for the first time induction of ΔFosB and CaMKII in the NAc of human cocaine addicts, suggesting possible targets for future therapeutic intervention. These data establish that ΔFosB and CaMKII engage in a cell-type- and brain-region-specific positive feedforward loop as a key mechanism for regulating the reward circuitry of the brain in response to chronic cocaine.
Introduction
Increasing evidence supports the view that changes in gene expression contribute to mechanisms of drug addiction (Robison and Nestler, 2011). One important mediator of these changes is ΔFosB, a Fos family transcription factor (Nestler, 2008). Chronic administration of virtually any drug of abuse induces the long-lasting accumulation of ΔFosB in nucleus accumbens (NAc), a limbic region essential for reward behaviors. Such induction appears specific to the class of NAc medium spiny neuron (MSNs) that expresses D1 dopamine receptors. Inducible overexpression of ΔFosB in these D1-type NAc MSNs increases locomotor and rewarding responses to cocaine and morphine (Kelz et al., 1999; Zachariou et al., 2006), including increased cocaine self-administration (Colby et al., 2003). Furthermore, genetic or viral blockade of ΔFosB transcriptional activity reduces the rewarding effects of these drugs (Zachariou et al., 2006), indicating that this sustained induction of ΔFosB is a critical mediator of the lasting changes induced in NAc by chronic drug administration.
The unusual stability of ΔFosB (relative to all other Fos family proteins) is both an intrinsic property of the molecule, attributable to the truncation of degron domains present in full-length FosB (Carle et al., 2007), and a regulated process. ΔFosB is phosphorylated in vitro and in vivo at Ser27, and this reaction further stabilizes ΔFosB, ∼10-fold, in cell culture and NAc in vivo (Ulery-Reynolds et al., 2009). Although Ser27 ΔFosB has been shown to be a substrate for casein kinase-2 in vitro (Ulery et al., 2006), its mechanism of in vivo phosphorylation remains unknown.
Calcium/calmodulin-dependent protein kinase II (CaMKII) is a highly expressed serine/threonine kinase whose α and β isoforms form dodecameric homo- and hetero-holoenzymes in vivo and are essential for multiple forms of neuroplasticity (Lisman et al., 2002; Colbran and Brown, 2004). CaMKIIα is induced selectively in NAc shell by chronic amphetamine (Loweth et al., 2010), and pharmacological blockade of CaMKII activity in NAc shell reduces behavioral sensitization to amphetamine (Loweth et al., 2008) and cocaine (Pierce et al., 1998), whereas viral overexpression of CaMKIIα in this NAc subregion enhances locomotor sensitization to and self-administration of amphetamine (Loweth et al., 2010). CaMKIIα may affect reward behaviors via modulation of AMPA glutamate receptor subunits (Pierce et al., 1998), because CaMKIIα activity has long been associated with AMPA receptor function and synaptic targeting in several forms of neuroplasticity (Malinow and Malenka, 2002).
This literature demonstrates several parallels between ΔFosB and CaMKII: both are necessary and sufficient for multiple behavioral effects of drugs of abuse, both upregulate dendritic spines in various neuronal cell types in vivo (Jourdain et al., 2003; Maze et al., 2010), and both exert at least some of their behavioral effects through modulation of AMPA receptors (Kelz et al., 1999; Malinow and Malenka, 2002; Vialou et al., 2010). Despite these parallels, no functional link between ΔFosB and CaMKII is known. Here, we establish reciprocal regulation between ΔFosB and CaMKII and demonstrate that the two proteins form a D1-type MSN-specific feedforward loop in NAc shell that is induced by cocaine and regulates a range of cocaine responses in vivo.
Materials and Methods
Experiment 1: isobaric tags for relative and absolute quantitation proteomic analysis of NAc shell and core after cocaine treatment (Fig. 1A).
Figure 1.

Shell-specific induction of CaMKII in NAc by cocaine. A, Depiction of the “skewed donut” method of dissecting NAc core (blue circle) and shell (remaining half-moon between red and blue circles) from a rat coronal brain slice. B, Locomotor activity analysis reveals that chronic (green) but not acute (yellow) preexposure to cocaine sensitizes animals to a cocaine challenge when compared with a saline control group (red) (n = 10; *p < 0.05, one-way ANOVA). C, Western blots of NAc shell and core from rats in B. D, Quantitation of Western blots in C shows significant increases in ΔFosB in both NAc shell and core, whereas significant increases in total CaMKIIα and phospho-Ser831 GluA1 are shell specific (n = 10; *p < 0.05, one-way ANOVA). E, Quantitation of Western blot analysis of rat NAc shell 14 d after the last injection of saline or cocaine either before (14d WD) or 1 h after a challenge dose of cocaine (14d WD Chal) (n = 9–10; *p < 0.05, two-tailed t test vs saline).
Adult (8 weeks) male rats were administered 20 mg/kg cocaine or saline vehicle intraperitoneally once per day for 7 d. At 24 h after the last injection, NAc shell and core were microdissected (Fig. 1A) and flash frozen. Isobaric tags for relative and absolute quantitation (iTRAQ) analyses were performed as described previously (Ross et al., 2004; Dávalos et al., 2010).
Experiment 2: quantifying protein changes in rat NAc core and shell after cocaine treatment (Fig. 1B–D).
Adult (8 weeks) male rats were administered 10 mg/kg cocaine or saline vehicle intraperitoneally once per day for 7 d in locomotor recording chambers. Locomotor responses to a single injection of cocaine (5 mg/kg, i.p.) were recorded in those animals treated previously with cocaine (called chronic) and a portion of those treated with saline (called acute), and locomotor responses to saline alone was recorded in the remaining chronic saline-treated animals (called saline). Locomotor activity assays were performed as described previously (Hiroi et al., 1997). Briefly, adult male rats were placed in 18 × 24 inch photobeam activity system open-field recording boxes (San Diego Instruments) for 30 min to habituate, were given a single intraperitoneal injection of saline and monitored for an additional 30 min, and were given a single intraperitoneal injection of 5 mg/kg cocaine and monitored for 30 min.
At 24 h after this final injection, rats were decapitated without anesthesia to avoid effects of anesthetics on neuronal protein levels and phospho-states. Brains were serially sliced in a 1.2 mm matrix (Braintree Scientific), and target tissue was removed in PBS containing protease (Roche) and phosphatase (Sigma-Aldrich) inhibitors using a 14 gauge punch for NAc core and a 12 gauge punch of the remaining tissue for NAc shell (Fig. 1A) and immediately frozen on dry ice. Samples were homogenized by light sonication in modified RIPA buffer: 10 mm Tris base, 150 mm sodium chloride, 1 mm EDTA, 0.1% SDS, 1% Triton X-100, 1% sodium deoxycholate, pH 7.4, and protease and phosphatase inhibitors as above. After addition of Laemmli's buffer, proteins were separated on 4–15% polyacrylamaide gradient gels (Criterion System; Bio-Rad), and Western blotting was performed using the Odyssey system (Li-Cor) according to the protocols of the manufacturer.
Experiment 3: quantifying protein changes in rat NAc core and shell after cocaine withdrawal (Fig. 1E).
Adult (8 weeks) male rats were administered 10 mg/kg cocaine or saline vehicle intraperitoneally once per day for 7 d. At 14 d after the final injection, animals treated with saline were given another saline injection (called saline), and animals treated with cocaine were given another saline injection (called 14 d withdrawal) or a single injection of cocaine (called 14 d withdrawal challenge). One hour after the final injection, animals were decapitated, and Western blotting performed as in experiment 2.
Experiment 4: quantifying protein changes in rat NAc core and shell after cocaine self-administration (Fig. 2A–C).
Figure 2.

Induction of CaMKII in NAc shell of self-administering rats and human cocaine addicts. A, Lever presses by rats allowed long or short access to cocaine self-administration. B, Immunohistochemical analysis reveals increased ΔFosB in NAc core and shell of long-access rats, whereas increases in CaMKIIα are shell specific; quantified in C (n = 6; *p < 0.05, one-way ANOVA). D, Western blots (below) reveal that cocaine-dependent humans display increased ΔFosB and CaMKIIα levels in shell-enriched NAc samples (n = 18–20; *p < 0.05, two-tailed t test).
Rats were trained to self-administer 0.5 mg/kg per infusion of cocaine in 1 h sessions under a fixed-ratio 1 schedule for 9 d. After nine baseline sessions, the rats were divided into two groups balanced by cocaine intake on the last two sessions. One group of rats was allowed to self-administer cocaine (0.5 mg/kg per infusion) in 1 h sessions (short access), whereas the other group of rats self-administered cocaine in 6 h sessions (long access) for 10 additional days (escalation sessions).
Brain sections were processed for immunohistochemistry as described previously (Perrotti et al., 2004). Brains were perfused 18–24 h after the last exposure to drug, resulting in the degradation of any residual full-length FosB protein such that all remaining immunoreactivity reflects ΔFosB. This degradation was confirmed by Western blotting, which showed no significant staining with an antibody directed against the C terminus of full-length FosB that does not recognize ΔFosB (data not shown). After slicing into 35 μm sections, the number of ΔFosB immunopositive cells was quantified by a blinded observer in two sections through the NAc of each rat, and mean values per 40× field were then calculated by region for each animal. Each animal was considered an individual observation for statistical analysis. Regions of interest were identified according to Paxinos and Watson (2007).
Quantification of CaMKIIα immunoreactivity was performed using a Li-Cor system as described previously (Covington et al., 2009). Integrated intensities of CaMKII and GAPDH were determined with Odyssey software. Results are calculated as integrated intensity values per square millimeter and are presented as means ± SEM (n = 4–10 per group). Values for GAPDH were used as reference to normalize CaMKII intensity for slice thickness and conditions.
Experiment 5: quantifying protein levels in cocaine-dependent humans (Fig. 2D).
Postmortem human brain tissues were obtained from the Quebec Suicide Brain Bank (Douglas Mental Health University Institute, Montreal, Quebec, Canada). The preservation of tissue proceeded essentially as described previously (Quirion et al., 1987). Briefly, once extracted, the brain is placed on wet ice in a Styrofoam box and rushed to the Quebec Suicide Brain Bank facilities. Hemispheres are immediately separated by a sagittal cut in the middle of the brain, brainstem, and cerebellum. Blood vessels, pineal gland, choroid plexus, half cerebellum, and half brainstem are typically dissected from the left hemisphere, which is then cut coronally into 1-cm-thick slices before freezing. The latter half cerebellum is cut sagittally into 1-cm-thick slices before freezing. Tissues are flash frozen in 2-methylbutane at −40°C for ∼60 s. All frozen tissues are kept separately in plastic bags at −80°C for long-term storage. Specific brain regions are dissected from frozen coronal slices on a stainless steel plate with dry ice all around to control the temperature of the environment. Western blotting was performed as described in experiment 2.
The cohort was composed of 37 male and three female subjects, ranging in age between 15 and 66 years. All subjects died suddenly without a prolonged agonal state or protracted medical illness. In each case, the cause of death was ascertained by the Quebec Coroner Office, and a toxicological screen was conducted with tissue samples to obtain information on medication and illicit substance use at the time of death. The subject group consisted of 20 individuals who met the Structured Clinical Interview for DSM-IV (Diagnostic and Statistical Manual of Mental Disorders-IV) Axis I Disorders: Clinician Version (SCID-I) criteria for cocaine dependence. The control group comprised 20 subjects with no history of cocaine dependence and no major psychiatric diagnoses. All subjects died suddenly from causes that had no direct influence on brain tissue. Groups were matched for mean subject age, refrigeration delay, and pH. For all subjects, psychological autopsies were performed as described previously (Dumais et al., 2005), allowing us to have access to detailed case information on psychiatric and medical history, as well as other relevant clinical and sociodemographic data. In brief, a trained interviewer conducted the SCID-I with one or more informants of the deceased. A panel of clinicians reviewed SCID-I assessments, case reports, coroner's notes, and medical records to obtain consensus psychiatric diagnoses.
Experiment 6: chromatin immunoprecipitation for rat NAc (Fig. 3A–C).
Figure 3.

Cell type- and region-specific ΔFosB induction of CaMKIIα in vivo. A, qChIP assays reveal increased ΔFosB binding to the CaMKIIα gene promoter in rat NAc shell but not core after chronic cocaine exposure (n = 6–7; *p < 0.05, two-tailed t test). B, qChIP also reveals decreased H3 acetylation after cocaine exposure in NAc core compared to shell (n = 6–7; p < 0.05, two-tailed t test). C, qChIP data suggesting reduced H3K9 dimethylation in both NAc shell and core after chronic cocaine. D, Quantitative PCR shows that mice overexpressing ΔFosB in D1-type, but not in D2-type, MSNs exhibit increased levels of CaMKIIα mRNA in NAc (n = 8–10; *p < 0.05 two-tailed t test). E, Immunohistochemical analysis shows that the D1- and D2-specific mouse lines overexpress ΔFosB to similar levels in NAc shell and core but that only D1-specific overexpression of ΔFosB increases total CaMKIIα protein; quantified in F (n = 6–8; *p < 0.05 two-tailed t test). G, Western blotting reveals that D1-specific, but not D2-specific, antagonist (Antag) coadministration prevents cocaine-mediated ΔFosB and CaMKIIα induction in rat NAc shell; quantified in H (n = 4–5; *p < 0.05 one-way ANOVA, different from vehicle). Con, Control.
Adult (8 weeks) male rats were administered 10 mg/kg cocaine or saline vehicle intraperitoneally once per day for 7 days. At 24 h after the last injection, NAc shell and core were microdissected. Chromatin immunoprecipitation (ChIP) was performed pooling bilateral NAc punches of shell or core from seven rats per group in 14 total groups (98 animals total, seven cocaine pools, seven saline pools). Tissues were crosslinked, washed, and stored at −80°C until chromatin shearing by sonication. Non-immune IgG was used as a control. After reverse crosslinking and DNA purification, qPCR was used to measure levels of CaMKIIα promoter DNA. Primers were designed to amplify a region containing an AP-1 consensus sequence located ∼450 bp before the transcription start site (forward, ACTGACTCAGGAAGAGGGATA; reverse, TGTGCTCCTCAGAATCCACAA).
Experiment 7: measuring CaMKII transcript and protein expression with cell-type-specific ΔFosB overexpression (Fig. 3D).
Male bitransgenic mice derived from NSE–tTA (line A) × TetOp–ΔfosB (line 11) and NSE–tTA (line B) × TetOp–FLAG–ΔfosB (line 11) mice (Chen et al., 1998; Kelz et al., 1999; Werme et al., 2002; Zachariou et al., 2006) were conceived and raised on 100 μg/ml doxycycline to suppress ΔFosB expression during development. Littermates were divided at weaning: half remained on doxycycline and half were switched to water, and the animals were used 8–11 weeks later when transcriptional effects of ΔFosB are maximal (Kelz et al., 1999; McClung and Nestler, 2003). For transcriptional analyses, mice were rapidly decapitated, and brains were removed and placed on ice. Dissections of NAc were taken with a 14 gauge needle punch and quickly frozen on dry ice until RNA was extracted. RNA isolation, qPCR, and data analysis were performed as described previously (LaPlant et al., 2009). Briefly, RNA was isolated with TriZol reagent (Invitrogen), further purified with the RNAeasy micro kit from Qiagen, and checked for quality with Agilent Bioanalyzer. Reverse transcription was performed using iScript (Bio-Rad). qPCR was performed with an Applied Biosystems 7900HT RT PCR system with the following cycle parameters: 10 min at 95°C; 40 cycles of 95°C for 1 min, 60°C for 30 s, 72°C for 30 s; graded heating to 95°C to generate dissociation curves for confirmation of single PCR products. Immunohistochemical analyses of ΔFosB and CaMKIIα protein expression were performed as described in experiment 4.
Experiment 8: effects of intra-NAc D1 and D2 dopamine receptor antagonists on cocaine-mediated protein changes (Fig. 3H).
Adult (8 weeks) male rats were administered 10 mg/kg cocaine or saline vehicle (vehicle group) intraperitoneally once per day for 7 d. At 30 min before each cocaine injection, rats were intraperitoneally administered either the D1 receptor antagonist SCH 23390 [R(+)-7-chloro-8-hydroxy-3-methyl-1-phenyl-2,3,4,5-tetrahydro-1H-3-benzazepine hydrochloride] (0.5 mg/kg), the D2 receptor antagonist eticlopride (0.5 mg/kg), or a saline control injection (cocaine group). At 24 h after the final injection, animals were decapitated, and proteins were quantified by Western blotting as per experiment 2.
Experiment 9: effects of adeno-associated viral-mediated ΔFosB overexpression on protein expression (Fig. 4A–C).
Figure 4.
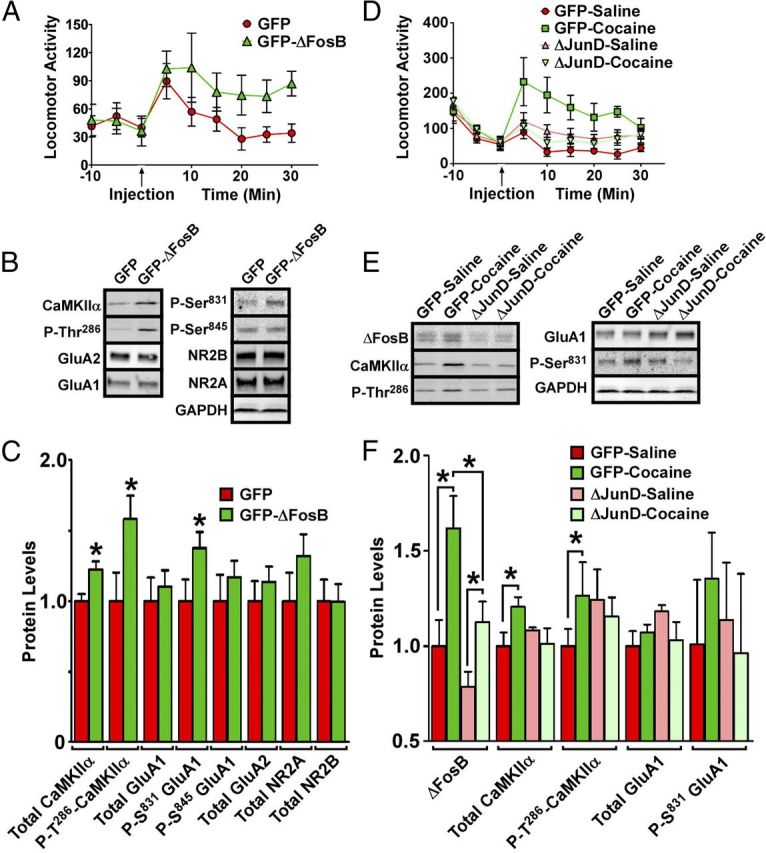
ΔFosB is both necessary and sufficient for cocaine-mediated D1 receptor-dependent CaMKIIα induction in NAc shell. A, AAV-mediated overexpression of ΔFosB in NAc shell promotes locomotor responses to an acute cocaine injection in adult male rats. B, Western blot analysis of NAc shell shows that ΔFosB is sufficient to increase levels of total CaMKIIα and both autophosphorylation of CaMKIIα and Ser831 phosphorylation of GluA1; quantified in C (n = 14–18; *p < 0.05, two-tailed t test). D, AAV-mediated ΔJunD overexpression prevents locomotor sensitization induced by chronic exposure to cocaine. E, ΔJunD overexpression in NAc shell is sufficient to block cocaine-mediated increases in total and Thr286 phospho-CaMKII and to reduce levels of ΔFosB in both saline- and cocaine-treated animals; quantified in F (n = 8–10; *p < 0.05, one-way ANOVA).
Stereotaxic surgery was performed on adult male rats (8 weeks) to inject adeno-associated viral (AAV)–green fluorescent protein (GFP) or AAV–GFP–ΔFosB (Maze et al., 2010). Thirty-three gauge needles (Hamilton) were used for all surgeries, during which 0.5 μl of purified high-titer virus was bilaterally infused over a 5 min period of time, followed by an additional 5 min postinfusion rest period. All distances are measured relative to bregma: 10° angle, +1.7 mm anteroposterior (AP), 2.5 mm lateral (L), −6.7 mm dorsoventral (DV). At 14 d after surgery, animals were given a single intraperitoneal injection of 10 mg/kg cocaine in locomotor monitoring chambers to assess the behavioral effects of ΔFosB overexpression. At 24 h after this final injection, rats were decapitated as per experiment 2, and tissue microdissection was performed under fluorescence microscopic guidance to obtain GFP-positive NAc tissue. Western blotting was then performed as per experiment 2.
Experiment 10: effects of AAV-mediated ΔJunD overexpression on cocaine-dependent protein expression (Fig. 4D–F).
Stereotaxic injection of AAV–GFP or AAV–GFP–ΔJunD was performed as per experiment 8. At 14 d after surgery, animals were administered 10 mg/kg cocaine or saline vehicle intraperitoneally once per day for 7 d in locomotor recording chambers. Locomotor responses to a single injection of cocaine (5 mg/kg, i.p.) or saline was recorded. At 24 h after this final injection, rats were decapitated, tissue was harvested, and Western blots were performed as in experiment 9.
Experiment 11: in vitro protein kinase assays (Fig. 5A–D).
Figure 5.
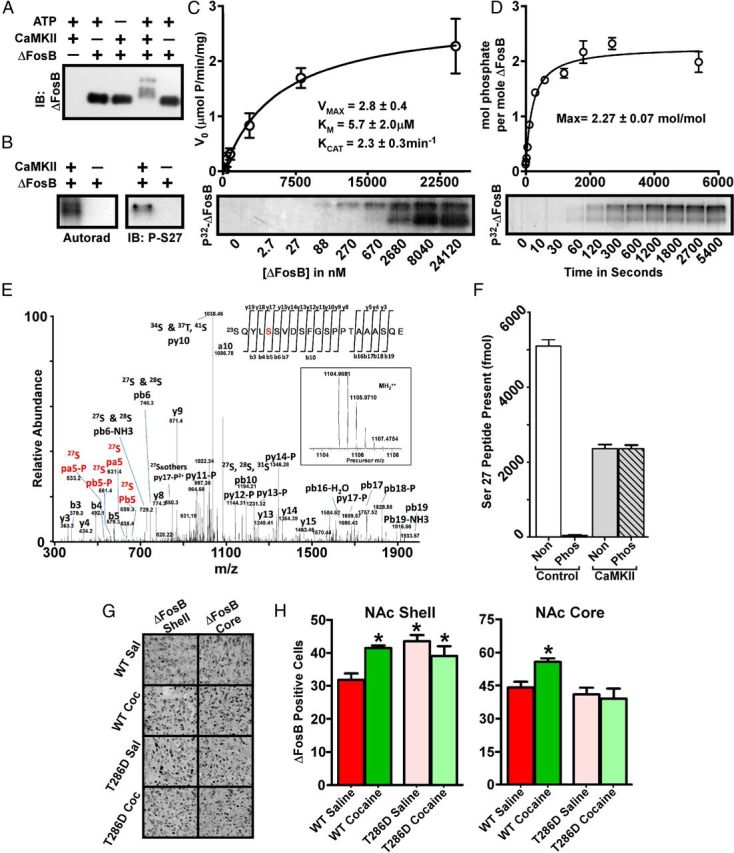
ΔFosB is a potent substrate for CaMKIIα. A, Western blotting shows an ATP-dependent multi-band shift in electrophoretic mobility of ΔFosB after exposure to CaMKIIα. IB, Immunoblot. B, Autoradiogram reveals a CaMKII-dependent incorporation of radiolabeled phosphate into ΔFosB (left), and a ΔFosB Ser27 phospho-specific antibody shows phosphorylation of this site by CaMKII (right). Analyses reveal robust kinase kinetics (C) and incorporation of multiple phosphates into ΔFosB by CaMKII (D). E, The precursor (inset) and fragment spectra of a TiO2 enriched phosphopeptide detected from ΔFosB after in vitro phosphorylation by CaMKII. After using both trypsin and GluC digestion and enrichment of the phosphopeptide samples by TiO2, analysis reveals phosphorylation of Ser27 as well as of several other sites not characterized further here. F, MRM analysis of ΔFosB phosphorylated in vitro by CaMKIIα reveals that Ser27 is a potent CaMKII substrate. Non, Nonphosphorylated peptide; Phos, phosphorylated peptide. G, Immunohistochemical analysis reveals increased ΔFosB in both the NAc shell and core of adult male wild-type (WT) mice exposed to chronic cocaine. Littermates overexpressing a constitutively active form of CaMKIIα show basal elevation in ΔFosB in the NAc shell only and show no effect of cocaine on ΔFosB levels in either region; quantified in H (n = 9–10; *p < 0.05, one-way ANOVA).
Recombinant CaMKIIα and ΔFosB were purified from insect cells (Brickey et al., 1990; Jorissen et al., 2007), and protein kinase assays were performed (Colbran, 1993) as described previously. Briefly, CaMKII was preincubated on ice with 2.5 μm (or indicated concentration) ΔFosB, 1 mm Ca2+, 40 mm Mg2+, 15 μm calmodulin, and 200 mm HEPES, pH 7.5. Phosphorylation was initiated by addition of 200 μm ATP with or without [γ-32P]ATP and allowed to proceed for 10 min at room temperature (Fig. 5A,B) or 2 min on ice (Fig. 5C,D). Products were resolved by Western blotting (Fig. 5A,B) or by autoradiogram and scintillation counting (Fig. 5B–D).
Experiment 12: identification of Ser27 ΔFosB phosphorylation (Fig. 5E).
In vitro kinase assays were performed as per experiment 11, proteins were separated by SDS-PAGE, and bands corresponding to ΔFosB were cut out and subjected to tandem mass spectrometry (MS/MS). The mass/charge (m/z) assignments of the corresponding ion fragments in all of the panels are labeled on top of the ion peaks. Not all fragment ions are labeled because of space limitations. Generally, the text for the fragment ion labels are colored in black except when they directly confirm or add evidence to the presence of the phosphorylation sites of interest, in which case they are marked in red. Evidence for backbone fragmentation products are presented in the sequence readout of the phosphopeptide with the detected site of phosphorylation residue indicated in red with a single amino acid letter designation. The numeric description of the observed fragment ions are also marked on the peptide sequence as b and y ions. The zoom factors for the sections of the m/z axis to show the lower-intensity fragment ions are marked at the top of each fragment mass spectra. The fragment ions shown in Figure 5H confirms the presence of Ser27 phosphorylated isoform, but within a mixture of other phosphorylated isoforms at sites Ser28, Ser31, Ser34, and Thr37. The presence of pa5, pa5-P, pb5, and pb5-P ions uniquely confirm the phosphorylation of the Ser27 residue.
Experiment 13: quantification of Ser27 phosphorylation (Fig. 5F).
Standard peptides were designed mimicking the phospho and non-phospho forms of Ser27 ΔFosB. After synthesis and purification, each “heavy” idiotypic peptide was dissolved in a 50:50 acetonitrile/water buffer and sent for amino acid analysis to determine absolute concentration on the synthetic peptide stock solution. Each heavy peptide was then directly infused into the 4000 QTRAP MS to determine the best collision energy for MS/MS fragmentation and two to four multiple reaction monitoring (MRM) transitions. Next, the neat heavy peptides were subjected to liquid chromatography (LC) MS on the 4000 QTRAP to ensure peptide separation. The instrument was run in the triple quadrupole mode, with Q1 set on the specific precursor m/z value (Q1 is not scanning) and Q3 set to the specific m/z value corresponding to a specific fragment of that peptide. In the MRM mode, a series of single reactions (precursor/fragment ion transitions in which the collision energy is tuned to optimize the intensity of the fragment ions of interest) were measured sequentially, and the cycle (typically 1–2 s) was looped throughout the entire time of the HPLC separation. MRM transitions were determined from the MS/MS spectra of the existing peptides. Two transitions per peptide, corresponding to high-intensity fragment ions, were then selected, and the collision energy was optimized to maximize signal strength of MRM transitions using automation software. Peaks resulting from standard peptides and ΔFosB samples exposed to CaMKII or control were then compared to determine the absolute abundance of each peptide form in the reaction. Data analysis on LC-MRM data is performed using AB Multiquant 1.1 software.
Experiment 14: induction of ΔFosB in CaMKII overexpressing mice Fig. 5G,H).
Transgenic mice overexpressing T286D CaMKII (Mayford et al., 1996; Kourrich et al., 2012) and wild-type littermates were raised in the absence of doxycycline to allow transgene expression. Adult mice were administered 20 mg/kg cocaine or saline intraperitoneally once daily for 14 d. At 24 h after the final injection, animals were decapitated, and immunohistochemistry and quantification of ΔFosB expression was performed as in experiment 4.
Experiment 15: effects of HSV-mediated ΔFosB overexpression and CaMKII inhibition on NAc dendritic spines (Fig. 6A–E).
Figure 6.
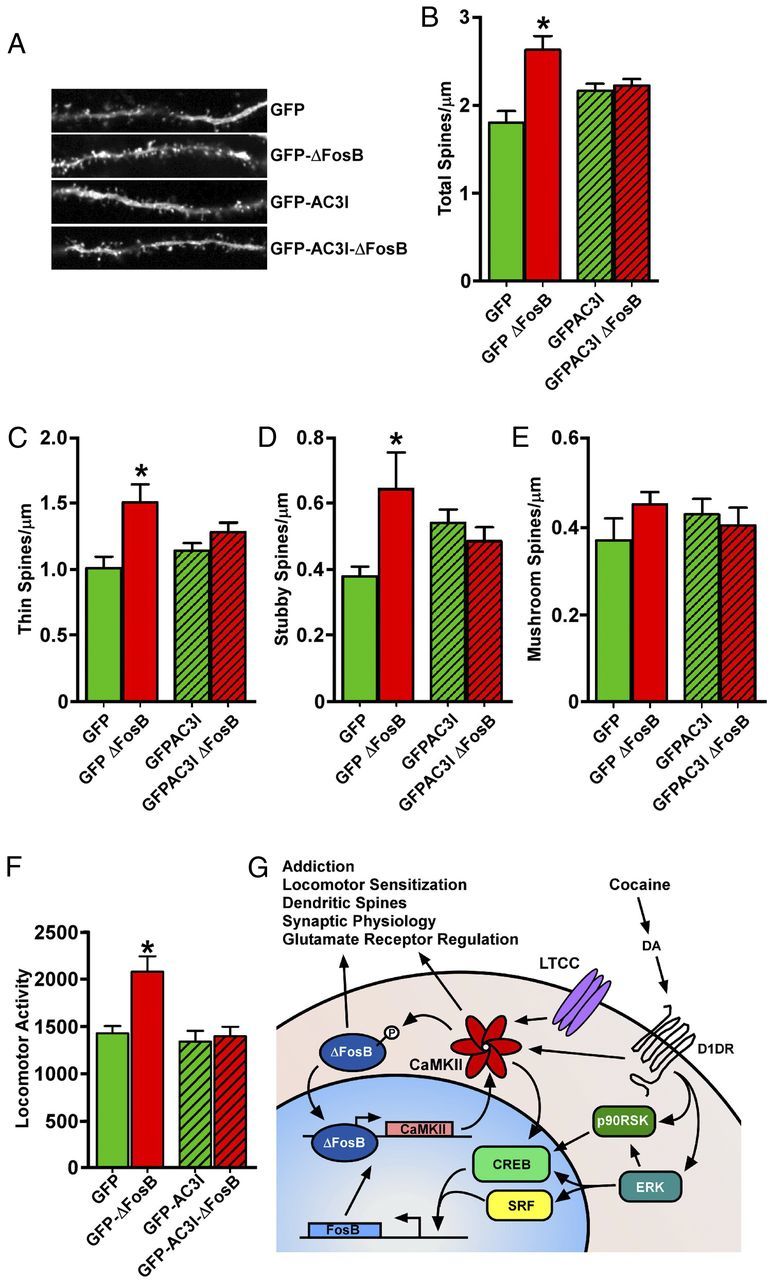
Blockade of CaMKII activity prevents the morphological and behavioral effects of ΔFosB in NAc. A, Increases in the spine density of MSNs in NAc shell induced by HSV-mediated overexpression of ΔFosB are prevented by coexpression of the CaMKII inhibitor peptide AC3I (n = 14–16); quantified in B. C–E, ΔFosB effects on thin and stubby spines are blocked by coexpression of AC3I. F, The ΔFosB-mediated increase in locomotor sensitivity to cocaine is also prevented by AC3I coexpression. G, Model depicting the D1-receptor-dependent induction of a CaMKII/ΔFosB feedforward loop by cocaine, including upstream signaling cascades and physiological processes that may be affected. DA, Dopamine; D1DR, D1 dopamine receptor; LTCC, L-type calcium channel (n = 9–10; *p < 0.05, one-tailed t test).
Adult male mice (8 weeks) were stereotaxically injected in NAc with HSV–GFP, HSV–GFP–ΔFosB (Olausson et al., 2006), HSV–GFPAC3I, or HSV–GFPAC3I–ΔFosB. In these constructs, AC3I, a peptide-based inhibitor of CaMKII activity, is fused to the C terminus of GFP. GFPAC3I was cloned by PCR using the pMM400 vector containing GFPAC3I as a template with the following primers: GFP-AC3I forward, 5′ CC GCTAGC GCCGCCACC ATGGTGAGCAAGGGCGAGGAGCTGT 3′ (clampNheIKozakmet); GFP-AC3I reverse, 5′ CC TCCGGA TTACAGGCAGTCCACGGCCT 3′ (clampBspEIstop). The resulting PCR product was inserted into the p1005+ and p1005+–ΔFosB vectors using NheI and BspEI sites. The construct was validated by sequencing. Stereotaxic coordinates were 10° angle, +1.6 mm AP, +1.5 mm L, −4.4 mm DV (Barrot et al., 2002). Perfusion and brain sectioning was performed as per experiment 4.
Spine analysis was performed as described previously (Christoffel et al., 2011). Briefly, dendritic segments 50–150 μm away from the soma were randomly chosen from HSV-infected cells that express GFP. Images were acquired on a confocal LSM 710 (Carl Zeiss) for morphological analysis using NeuronStudio with the rayburst algorithm. NeuronStudio classifies spines as thin, mushroom, or stubby based on the following values: (1) aspect ratio, (2) head to neck ratio, and (3) head diameter. Spines with a neck can be classified as either thin or mushroom, and those without a significant neck are classified as stubby. Spines with a neck are labeled as thin or mushroom based on head diameter.
Experiment 16: effects of HSV-mediated ΔFosB overexpression and CaMKII inhibition on cocaine responses (Fig. 6F).
Adult male mice were injected with viruses as per experiment 15, and locomotor responses to a single 5 mg/kg injection of cocaine was measured as per experiment 9. Locomotor data are expressed as total beam breaks over 30 min after cocaine injection.
Animal housing.
Male Sprague Dawley rats (250–275 g; Charles River Laboratories) were housed in pairs. Eight-week-old C57BL/6J male mice (The Jackson Laboratory) were group housed with a maximum of five animals per cage. All animals were habituated to the animal facility for ≥1 week before experimental manipulations and housed in climate-controlled rooms (23–25°C) on a 12 h light/dark cycle (lights on at 7:00 A.M.) with access to food and water ad libitum. Experiments were conducted in accordance with guidelines of the Society for Neuroscience and the institutional animal care and use committee at Mount Sinai.
Drugs.
Drugs were administered intraperitoneally and dissolved in sterile saline, including cocaine (5–20 mg/kg per 10 μl for mice, per 1 ml for rats; National Institute on Drug Abuse) and SCH 23390 or eticlopride hydrochloride (0.5 mg/kg per 1 ml; Tocris Bioscience). For stereotaxic surgery, mice were anesthetized with a mixture of ketamine (100 mg/kg) and xylazine (10 mg/kg) (Henry Schein) in sterile saline.
Antibodies.
The following antibodies were used: CaMKIIα (total), Millipore catalog #05-532, at 1:5000; CaMKII phospho-Thr286, Promega catalog #V111A, at 1:1000; ΔFosB (total), Cell Signaling Technology catalog #5G4, at 1:250; ΔFosB phospho-Ser27, Phosphosolutions, at 1:500; GluA1 (total), Abcam catalog #Ab31232, at 1:1000; GluA1 phospho-Ser831, Millipore catalog #N453, at 1:1000; GluA1 phospho-Ser845, Millipore Bioscience Research Reagents catalog #Ab5849, at 1:2000; GluA2, Millipore catalog #07-598, at 1:2000; NR2A, Sigma catalog #HPA004692, at 1:2500; and NR2B, Millipore catalog #Ab1557P, at 1:1000.
Statistical analyses.
All statistical analyses were performed using the Prism 6 software package (GraphPad Software). Student's t tests were used for all pairwise comparisons (indicated in Results when t value is given), and one-way ANOVAs were used for all multiple comparisons (indicated in results section when F value is given).
Results
Chronic cocaine induces CaMKII in the NAc shell
Many studies have indicated that MSNs in the NAc shell and core have different biochemical and physiological responses to chronic exposure to drugs of abuse (Kourrich and Thomas, 2009; Loweth et al., 2010) and that the two subregions differentially regulate drug-seeking behaviors (Ito et al., 2004). To determine the differential effects of cocaine on the protein constituents of NAc shell versus core, we used iTRAQ and MS/MS. Adult male rats were injected intraperitoneally with cocaine (20 mg/kg) or saline daily for 7 d; at 24 h after the last injection, NAc shell and core were microdissected (Fig. 1A) and flash frozen. Proteins in these samples were then quantified using iTRAQ. All four CaMKII isoforms displayed large increases in expression after cocaine treatment that were specific to NAc shell compared with core. Several protein phosphatases, including PP1 catalytic and regulatory subunits and PP2A, which have been associated previously with various CaMKII substrates in other systems (Colbran, 2004), followed a similar pattern. These findings provided novel, unbiased evidence that the CaMKII signaling pathway is prominently regulated by cocaine in NAc in a shell-specific manner.
To validate this finding more quantitatively, we treated rats as above with cocaine (at varying doses) or saline and measured locomotor responses to a cocaine (5 mg/kg) or saline challenge dose. Repeated exposure to 10 mg/kg cocaine resulted in the typical pattern of locomotor sensitization (Fig. 1B). Additional studies with this dosing regimen revealed, by use of Western blotting, that repeated cocaine induces CaMKIIα selectively in NAc shell 24 h after the final injection of cocaine (Fig. 1C,D; p = 0.0019; F = 7.943; df = 29). In addition, phosphorylation of the canonical CaMKII substrate Ser831 of the GluA1 subunit of the AMPA receptor was significantly increased in NAc shell and not core (p = 0.0261; F = 4.208; df = 28), whereas CaMKIIα Thr286 autophosphorylation had a strong but not significant trend toward induction in shell only (Fig. 1D). Several other glutamate receptors were unaffected. In contrast to these measures of CaMKII, the same tissue samples displayed induction of ΔFosB in both shell (p = 0.0260; F = 4.189; df = 29) and core (p = 0.0350; F = 3.807; df = 29) of the NAc (Fig. 1C,D), consistent with previous findings (Perrotti et al., 2008).
Because several previous studies of cocaine regulation of AMPA receptors analyzed animals after ∼14 d of withdrawal from chronic cocaine (Discussion), we repeated these biochemical analyses at this time point. We found that, 14 d after the final injection of cocaine, ΔFosB remains elevated in NAc (p = 0.0288; F = 4.258; df = 22), whereas neither CaMKII nor phosphorylation of GluA1 Ser831 remains increased (Fig. 1E). However, 1 h after a single 10 mg/kg challenge dose of cocaine, levels of total CaMKII (p = 0.0330; F = 3.947; df = 26) and GluA1 Ser831 (p = 0.0213; F = 4.509; df = 27) phosphorylation are both elevated to a degree similar to that found after initial chronic cocaine exposure (Fig. 1E). These data indicate that NAc shell neurons are primed for CaMKII induction during extended periods of abstinence, perhaps via direct priming of the CaMKII gene promoter (Discussion). Moreover, the fact that ΔFosB induction is more persistent than CaMKII induction suggests the existence of additional mechanisms, whether chromatin-based or otherwise, that exert a “brake” on CaMKII regulation, as covered in Discussion.
To further strengthen these observations, we explored models of cocaine self-administration, which involve volitional drug intake. Adult male rats were given either short or long access to cocaine; as expected (Ahmed and Koob, 1998), only long access conditions led to escalating self-administration of the drug (Fig. 2A). ΔFosB was induced to a greater extent by long versus short access to cocaine in both NAc shell (p = 0.0011; F = 11.12; df = 17) and core (p = 0.0004; F = 13.86; df = 17). In contrast, CaMKIIα was induced in NAc shell only by long access to cocaine (Fig. 2B,C; p = 0.0236; F = 4.957; df = 16). It is interesting to compare the average daily cocaine intake across short-access animals (∼12 mg/kg, i.v.), long-access animals (∼70 mg/kg, i.v.), and experimenter-administered animals (10 mg/kg) and ask why the latter elicits robust induction of ΔFosB and CaMKII whereas short-access does not. This discrepancy is likely attributable to differences in peak cocaine levels (experimenter-administered cocaine is given as a single bolus intraperitoneally, whereas self-administered cocaine is delivered via multiple intraperitoneal doses), or differences in length of drug exposure (7 d for experimenter administration, 19 d for self-administration).
Despite the large literature on ΔFosB and CaMKII in cocaine action, there are no studies of these proteins in human cocaine users. Here, we present the first evidence that levels of both ΔFosB (p = 0.0316; t = 1.921; df = 34) and CaMKII (p = 0.0444; t = 1.755; df = 32) are increased in NAc of cocaine-dependent humans (Fig. 2D, Table 1). These data indicate that our examination of ΔFosB and CaMKII induction by cocaine in rodent NAc is clinically relevant to human cocaine addiction.
Table 1.
Characterization of samples from human cocaine addicts and matched control group
| Group | Percentage suicide | Age (years) | Gender (% male) | Brain weight (g) | pH | Refrigeration delay (h) |
|---|---|---|---|---|---|---|
| Control | 32% | 33.05 ± 3.193 | 95% | 1508 ± 37.08 | 6.569 ± 0.062 | 5.934 ± 1.038 |
| Cocaine dependent | 80% | 39.80 ± 2.153 | 95% | 1434 ± 26.32 | 6.546 ± 0.072 | 8.850 ± 1.812 |
| p value | 0.11 | 0.18 | 0.48 | 0.016 |
The control group contains 19 individuals, and the cocaine-dependent group contains 20 individuals. All values are mean ± SE. p values calculated using two-tailed Student's t test.
ΔFosB regulates CaMKII transcription selectively in D1-type MSNs of NAc shell
The finding that both CaMKII and ΔFosB are upregulated by cocaine in the rodent NAc led us to determine whether ΔFosB might regulate transcription of the CaMKII gene. We previously reported CaMKIIα as a possible target for ΔFosB in an unbiased microarray analysis of NAc (McClung and Nestler, 2003), but this finding was not further validated in that study. We first used quantitative ChIP (qChIP; ChIP followed by quantitative PCR) to determine whether ΔFosB binds to the CaMKIIα gene promoter in NAc of adult male rats and found strikingly that this binding is significantly increased, by chronic cocaine administration, in the shell (p = 0.0133; t = 2.901; df = 12) but not the core subregion (Fig. 3A). To further understand the mechanisms related to this subregion-specific difference in ΔFosB binding to the CaMKIIα promoter, we used qChIP to characterize the state of histone modifications at this genomic region. Previous studies demonstrated cocaine induction of H3 acetylation at the CaMKIIα promoter in total mouse NAc (Wang et al., 2010). In contrast, we found that cocaine decreases H3 acetylation at the CaMKIIα promoter selectively in NAc core (Fig. 3B; p = 0.0213; t = 2.726; df = 10), with no change apparent in shell, consistent with subregion-specific chromatin alterations beyond ΔFosB binding. qChIP for the repressive mark, dimethylated H3 lysine 9, revealed trends for decreases in both the shell and core subregions (Fig. 3C).
To determine whether ΔFosB regulates CaMKIIα transcription in vivo, we used two bitransgenic mouse lines that inducibly overexpress ΔFosB specifically in D1- versus D2-type MSNs in a manner controlled by doxycycline administration in drinking water (Chen et al., 1998; Kelz et al., 1999; Werme et al., 2002). Adult male mice overexpressing ΔFosB solely in D1-type MSNs had significantly increased levels of CaMKIIα mRNA in NAc (p = 0.0337; t = 1.996; df = 13), an effect not seen in mice overexpressing ΔFosB predominantly in D2-type MSNs (Fig. 3D). The increase in CaMKIIα mRNA, induced by ΔFosB expression in D1-type MSNs, was accompanied by a concomitant increase in CaMKIIα protein in both NAc shell (p = 0.0030; t = 3.578; df = 14) and core (p = 0.0392; t = 2.275; df = 14; Fig. 3E,F). These data demonstrate that ΔFosB is capable of driving CaMKIIα gene expression in D1-type MSNs in both subregions, although Figure 3B suggests that cocaine-mediated chromatin changes at the CaMKIIα promoter (e.g., reduced acetylation) prevent ΔFosB from upregulating CaMKII in the core subregion after cocaine.
Because our transgenic mouse data indicated that ΔFosB induction of CaMKII gene expression is specific to D1-type MSNs in NAc, we next sought to determine whether cocaine-dependent upregulation of CaMKII requires activation of the D1 dopamine receptor. Adult male rats were administered chronic cocaine or saline as before, but 30 min before each injection, rats in the cocaine group were given an intraperitoneal injection of saline, the D1 antagonist SCH 23390 (0.5 mg/kg), or the D2 receptor antagonist eticlopride (0.5 mg/kg). Animals were analyzed 24 h after the last injection of cocaine. Western blotting revealed that the D1, but not the D2, antagonist completely blocked the cocaine-mediated increase in ΔFosB (p < 0.0001; F = 18.96; df = 18), as reported previously (Nye et al., 1995), as well as in CaMKII (p = 0.0005; F = 10.99; df = 18; Fig. 3G,H). These data support the hypothesis that cocaine engages a ΔFosB-mediated increase in CaMKII gene expression specifically in D1-type MSNs of NAc shell. It would be important in future studies to demonstrate directly this cell-type-specific effect of cocaine on CaMKII expression within this brain region.
ΔFosB is both necessary and sufficient for cocaine induction of CaMKII in NAc shell
To complement the use of bitransgenic mice, we next studied the role of ΔFosB in mediating cocaine induction of CaMKIIα by use of viral-mediated gene transfer in rats. We bilaterally injected AAV particles into NAc shell of adult male rats (in which shell can be selectively targeted) to overexpress ΔFosB plus GFP or GFP alone. The animals were then given a single intraperitoneal injection of 10 mg/kg cocaine. The animals overexpressing ΔFosB/GFP exhibited an increased locomotor response compared with animals overexpressing GFP alone (Fig. 4A). At 24 h after the single cocaine injection, GFP-positive NAc tissue was excised from these animals by dissection under a fluorescent light source. Western blotting of this tissue (Fig. 4B,C) revealed strong ΔFosB overexpression as well as a significant increase in total CaMKIIα protein compared with GFP animals (p = 0.0070; t = 2.894; df = 30), similar to the induction seen with chronic cocaine administration. In addition, CaMKIIα autophosphorylation at Thr286 (indicative of enzyme activation) was increased by ΔFosB overexpression (p = 0.0330; t = 2.243; df = 28), as was phosphorylation of the CaMKII substrate, Ser831 of GluA1 (p = 0.0540; t = 2.012; df = 28), again mimicking the actions of chronic cocaine (Fig. 1C,D). Together, these data provide additional evidence that ΔFosB expression in NAc shell is sufficient for locomotor sensitization to cocaine and for CaMKII induction and activation in this subregion.
We used a similar approach to determine whether ΔFosB is also necessary for cocaine-mediated induction of CaMKIIα in the NAc shell. AAV was used to overexpress a truncated JunD protein, termed ΔJunD, which is a negative regulator of ΔFosB transcriptional activation (Winstanley et al., 2007) plus GFP or GFP alone. Two weeks later, when transgene expression is maximal, animals were given cocaine (10 mg/kg) or saline daily for 7 d, and tested for locomotor responses to a cocaine challenge (5 mg/kg) 24 h after the last chronic injection (Fig. 4D). ΔJunD overexpression prevented locomotor sensitization to cocaine and also prevented CaMKIIα induction and activation in NAc shell (Fig. 4E,F; p = 0.0437; F = 2.997; total df = 38), indicating that ΔFosB transcriptional activity is necessary for cocaine-mediated induction of CaMKIIα in this subregion. Interestingly, we found that ΔJunD reduced levels of ΔFosB under both saline- and cocaine-treated conditions (p = 0.0004; F = 8.110; df = 35), raising the novel possibility that ΔFosB depends on AP-1 activity for its own expression levels.
CaMKII phosphorylates ΔFosB at Ser27
Using in vitro protein kinase assays, we determined that purified ΔFosB is a robust substrate for CaMKIIα. Incubation of His6–ΔFosB with CaMKIIα and ATP caused an upward shift in electrophoretic mobility of ΔFosB (Fig. 5A); the several resulting bands suggested multiple sites of phosphorylation. Similar in vitro kinase assays using [γ-32P]ATP showed incorporation of radiolabeled phosphate into the shifted ΔFosB bands (Fig. 5B), demonstrating direct phosphorylation of the protein. We generated a phospho-specific antibody to the previously characterized Ser27 of ΔFosB (Ulery et al., 2006). Although this antibody does not produce a signal against brain extracts that contain Ser27-phosphorylated ΔFosB (data not shown), we were able to detect Ser27 phosphorylation in the in vitro kinase assay using CaMKII (Fig. 5B). Kinetic analyses of the CaMKII phosphorylation of ΔFosB indicate that it is a potent substrate for the kinase (Fig. 5C), with an apparent KM of 5.7 ± 2.0 μm and KCAT of 2.3 ± 0.3 min−1. These results are comparable with many well-characterized in vivo substrates of CaMKII (Colbran and Brown, 2004). In addition, we determined that CaMKII phosphorylates ΔFosB with a stoichiometry of 2.27 ± 0.07 mol/mol (Fig. 5D), indicating that there are at least three sites of CaMKII phosphorylation within the His6–ΔFosB protein, in agreement with Figure 5A.
To investigate individual sites of phosphorylation, we used MS analyses of samples from our in vitro kinase assays. Figure 5E demonstrates ΔFosB phosphorylation at the previously characterized Ser27 and at several additional sites (data not shown). Given the previous functional characterization of Ser27, we focused on this site by generating labeled synthetic peptides mimicking the phospho- and non-phospho-states of Ser27 and then used known quantities of these peptides as standards in MRM analyses of ΔFosB before and after in vitro phosphorylation by CaMKII. Subsequent quantitation (Fig. 5F) confirms that Ser27 is a potent substrate for CaMKII. These results indicate that, among multiple phosphorylated residues within ΔFosB, Ser27 is a particularly effective substrate for CaMKII.
CaMKII mediates cocaine accumulation of ΔFosB in the NAc shell
Because CaMKII can phosphorylate ΔFosB in vitro at a site that dramatically enhances its stability in vitro and in vivo (Ulery et al., 2006; Ulery-Reynolds et al., 2009), we determined whether CaMKII activity controls ΔFosB levels in NAc in vivo. To address this question, we first used a mouse line overexpressing a calcium-independent mutant of CaMKIIα (T286D) in multiple brain regions including NAc (Mayford et al., 1996; Kourrich et al., 2012). We injected age-matched adult male mutant and wild-type littermates with 20 mg/kg cocaine or saline once daily for 14 d and then analyzed the animals 1 d after the final injection. We found that basal levels of ΔFosB were increased in the mutant animals in NAc shell (p = 0.0001; F = 9.207; df = 37) but not core (Fig. 5G,H). Surprisingly, cocaine-dependent induction of ΔFosB was blocked in the mutant animals in both shell and core, suggesting that, although CaMKII may directly regulate ΔFosB stability in NAc shell, it may also lie upstream of ΔFosB in cocaine-activated pathways in both NAc subregions.
CaMKII activity is required for ΔFosB-mediated structural and behavioral plasticity
Cocaine induction of dendritic spines on NAc MSNs is one of the best established drug-induced adaptations in this brain region, and such spine induction has been correlated with sensitized behavioral responses to the drug (Robinson and Kolb, 2004; Russo et al., 2010) and reported to be selective for D1-type MSNs (Lee et al., 2006). We demonstrated recently that cocaine induction of dendritic spines in NAc is dependent on ΔFosB and its downstream transcriptional program (Maze et al., 2010). Although there is an extensive literature concerning the involvement of CaMKII in dendritic spine morphology and induction in other brain regions and experimental systems (Jourdain et al., 2003; Penzes et al., 2008; Okamoto et al., 2009), its role in NAc MSN spine formation has not been studied. Therefore, we determined whether CaMKII activity is required for ΔFosB-mediated induction of MSN dendritic spines by using HSV-mediated overexpression of the CaMKII inhibitor peptide AC3I fused to GFP, a construct shown previously to inhibit CaMKII activity in vivo (Zhang et al., 2005; Klug et al., 2012). Viral overexpression of ΔFosB in NAc shell of adult mice induced a significant increase in MSN dendritic spine density (p < 0.0001; F = 8.558; df = 59; Fig. 6A,B) as reported previously (Maze et al., 2010), and this increase was driven primarily by thin (p = 0.0027; F = 5.319; df = 59) and stubby (p = 0.0378; F = 2.988; df = 59) spine types (both thought to be immature spines) (Fig. 6C–E). No effect was seen on more mature, mushroom-shaped spines. However, when GFP–AC3I was coexpressed, ΔFosB induction of spines was completely abrogated (Fig. 6A–E), indicating that CaMKII activity is required for ΔFosB induction of dendritic spines in NAc shell.
We next used the same viral tools to determine whether CaMKII activity is required for the effects of ΔFosB on behavioral sensitivity to cocaine. At 72 h after viral injection into NAc shell, animals were given a single injection of 5 mg/kg cocaine and their locomotor activity was recorded. As shown previously with more extended AAV overexpression of ΔFosB (Fig. 4A), HSV-mediated overexpression of ΔFosB increased locomotor sensitivity to cocaine (p = 0.0002; F = 8.823; df = 37; Fig. 6F). As with induction of dendritic spines, inhibition of CaMKII activity by coexpression of GFP–AC3I completely blocked the ΔFosB-mediated increase in cocaine sensitivity, indicating that CaMKII activity is required for ΔFosB-induced alterations in the behavioral effects of cocaine.
Discussion
The present study delineates a novel feedforward mechanism in which cocaine induces ΔFosB in NAc, which upregulates transcription of the CaMKIIα gene selectively in NAc shell. CaMKIIα subsequently phosphorylates and stabilizes ΔFosB, leading to greater ΔFosB accumulation and to further CaMKIIα induction (Fig. 6G). The co-escalating levels of the two proteins during chronic exposure to cocaine then contribute in essential ways to sensitized behavioral responses to the drug. This is a particularly appealing hypothesis because both ΔFosB and CaMKII have each been demonstrated previously to be required for increased behavioral responses to cocaine (Pierce et al., 1998; Peakman et al., 2003), and we replicate this finding for ΔFosB in NAc shell specifically using a viral approach (Figs. 4, 6).
Although transgenic ΔFosB overexpression in D1-type MSNs can drive CaMKII induction in both NAc shell and core of cocaine-naive animals, in the context of cocaine, accumulation of endogenous ΔFosB, which occurs in both subregions, drives induction of CaMKII specifically in NAc shell. This difference could relate to the higher levels of ΔFosB induced in our bitransgenic model, but it might also reflect the ability of cocaine to differentially alter the CaMKIIα promoter in shell versus core MSNs to either promote ΔFosB binding in the former or exclude it in the latter subregion. In fact, our ChIP data, which reveal a cocaine-mediated deacetylation of histones at the CaMKIIα gene promoter in NAc core only, support the possible involvement of a chromatin mechanism. In keeping with this hypothesis, ΔFosB overexpression in D1-type MSNs was able to drive CaMKIIα induction in NAc core in the absence of cocaine (Fig. 3F), suggesting that there are active modifications of the CaMKIIα promoter that prevent this induction during chronic cocaine exposure. Regulation of the chromatin landscape at the CaMKII promoter might also explain why CaMKII is induced by a challenge dose of cocaine in NAc shell of chronic cocaine-withdrawing rats (Fig. 1E) but not of drug-naive animals (Fig. 1D). This could represent an epigenetic “gene priming” effect of ΔFosB (Robison and Nestler, 2011) and might thus be one molecular mechanism of the incubation of cocaine craving (Pickens et al., 2011). However, for this chromatin change to be causally linked to incubation of craving, it would have to increase over time. It will be interesting to determine whether this is the case and to study whether other genes show ΔFosB-dependent, subregion-specific regulation by cocaine. It is also important to note that the feedforward loop we describe does not lead to an endless accumulation of CaMKII or ΔFosB (Fig. 1E); uncovering the molecular brake responsible for this is an important goal of future studies.
The known functions of ΔFosB and CaMKII in several experimental systems and brain regions converge at many levels (Fig. 6F). Both molecules are intimately linked to dendritic spine growth: CaMKII interacts with the actin cytoskeleton (Okamoto et al., 2009), regulates spine head size (Matsuzaki et al., 2004), and is both necessary and sufficient for plasticity-induced increases in filopodia and synapse number in hippocampal organotypic slice cultures (Jourdain et al., 2003), whereas ΔFosB is both necessary and sufficient for cocaine-induced dendritic spine formation in NAc MSNs (Maze et al., 2010). Additionally, both molecules have been associated with regulation of AMPA glutamate receptors. CaMKII does not regulate total levels of AMPA receptor subunits but drives the insertion of AMPA receptors into synapses and increases AMPA channel conductance by phosphorylating GluA1 at Ser831 in hippocampal pyramidal neurons in culture and in vivo (for review, see Malinow and Malenka, 2002; Colbran and Brown, 2004). Such increased trafficking of GluA1 to the synapse has been implicated in chronic cocaine action as well (Boudreau and Wolf, 2005). Moreover, behavioral responses to AMPA receptor activation in NAc are enhanced by CaMKIIα overexpression in a D1 dopamine receptor-dependent manner (Singer et al., 2010). Long-term D1-specific overexpression of ΔFosB has been shown to induce GluA2 transcription in NAc (Kelz et al., 1999), which dampens AMPA responses mediated via GluA1, whereas we show here that shorter-term ΔFosB overexpression—as well as shorter-term cocaine exposure—have no effect on this subunit (Fig. 1). Nevertheless, we have found recently that short-term ΔFosB overexpression nevertheless reduces AMPA responses in D1-type MSNs in NAc (Grueter et al., 2013). These data suggest temporally distinct mechanisms that might constitute a time-dependent series of neuroadaptations to cocaine that underlie different aspects of addiction progression not yet well understood. At the behavioral level, both CaMKII and ΔFosB are required for locomotor sensitization to cocaine (above), and both are required for sustained cocaine self-administration in rodents (Colby et al., 2003; Wang et al., 2010), suggesting that the two proteins are important for both short- and long-term behavioral adaptations to drug exposure, albeit via partly distinct underlying mechanisms. Presumably, ΔFosB and CaMKII regulate such complex behavioral adaptations through changes in NAc synaptic function, although much additional work is needed to directly link synaptic phenomena to behavioral change.
The CaMKII holoenzyme simultaneously interacts with a variety of synapse-associated proteins (Robison et al., 2005) that are thought to regulate its targeting to the postsynaptic density, a phenomenon suggested to be important for synaptic plasticity. In particular, the interaction of CaMKII with the GluN2B subunit of the NMDA-type glutamate receptor was shown recently to regulate both synaptic plasticity and learning (Halt et al., 2012). Although the AC3I peptide mimics the autoinhibitory domain of CaMKII, and thus inhibits enzyme catalytic activity, it also blocks multiple protein–protein interactions (Strack et al., 2000; Robison et al., 2005). Thus, the behavioral and morphological effects of HSV–GFP–AC3I reported here could occur through reduced phosphorylation of CaMKII target proteins, changes in CaMKII targeting, or a change in the proposed structural role of CaMKII at synapses (Lisman et al., 2002).
The restriction of the proposed ΔFosB–CaMKII loop to the NAc shell is of special note, because recent work has demonstrated several physiological differences between the NAc shell and core in response to cocaine administration, a notion confirmed by our unbiased iTRAQ data (Notes). MSNs in NAc shell show a depression in firing capacity after chronic cocaine that is sustained for weeks, whereas core MSNs from the same animals display a transient (1–3 d) increase in firing capacity that returns to basal levels within 2 weeks (Kourrich and Thomas, 2009). In addition, numerous synaptic proteins are differentially regulated in NAc shell versus core of animals exposed to chronic cocaine, including GluA2 (Knackstedt et al., 2010). Because chronic amphetamine induces CaMKIIα specifically in NAc shell (Loweth et al., 2010), it is not surprising that we find a similar effect with cocaine. However, because ΔFosB is induced in both the NAc shell and core by chronic cocaine (Perrotti et al., 2008) and because we show that CaMKIIα induction in shell is ΔFosB dependent, our findings provide new evidence for distinct transcriptional mechanisms at the CaMKIIα promoter between these two subregions, which are responsible for the selective induction of CaMKIIα in shell.
A great deal of recent work has focused on delineating differences between D1- and D2-type NAc MSNs. Although both D1 and D2 receptors are involved in the rewarding effects of cocaine (Self, 2010), recent work demonstrates that optogenetic activation of D1-type MSNs increases behavioral responses to cocaine, whereas D2-type MSN activation has the opposite effect (Lobo et al., 2010). In line with these findings, D1 receptor knock-out mice are deficient in acquisition of cocaine self-administration (Caine et al., 2007), whereas D2 knock-outs are not (Caine et al., 2002). D1 agonist administration directly into NAc triggers cocaine-seeking behavior in reinstatement paradigms (Self, 2010). Interestingly, this effect requires D1-receptor-dependent increases in CaMKII activity in the NAc shell but not core (Anderson et al., 2008), a result that dovetails nicely with the D1- and shell-specific ΔFosB–CaMKII loop proposed here.
We reported previously that Ser27 in ΔFosB can be phosphorylated by casein kinase-2 (Ulery et al., 2006), but we establish here that CaMKII phosphorylates ΔFosB at this and other sites with far greater kinetics and stoichiometry and can replicate the higher apparent Mr observed for ΔFosB (Fig. 5A) with cocaine exposure in vivo (Nestler, 2008). We already know that Ser27 phosphorylation increases ΔFosB stability and transcriptional activity (Ulery et al., 2006; Ulery and Nestler, 2007; Ulery-Reynolds et al., 2009). Future work will now focus on the identification and the functional consequences of novel sites of ΔFosB phosphorylation indicated by the present study.
The feedforward loop described here provides a plausible new mechanism by which repeated administration of cocaine drives progressive abnormalities in the NAc. As such, this biochemical pathway may provide an important target for future therapeutic intervention in addictive disorders. Because CaMKII is ubiquitous and required for many basal neuronal and behavioral functions, direct use of CaMKII inhibitors has been avoided as an addiction treatment. Our data suggest that more subtle targeting of the mechanism of CaMKII induction, which is specific to an individual cell type and subregion of the reward circuitry of the brain, could provide a therapeutic target that would avoid the complications of systemic CaMKII inhibition.
Notes
Supplemental material for this article is available at http://transmitter.neuro.mssm.edu/nestler/nidappg/chromatingenedatabase.html. A table of iTRAQ results is shown. This material has not been peer reviewed.
Footnotes
This work was supported by the National Institute on Drug Abuse (NIDA) (E.J.N.), NIDA–Yale Proteomics Center Grant DA018343 (A.J.R. and E.J.N.), and the Hartwell Foundation (A.J.R.). We thank Gabby Rundenko for the generous gift of purified ΔFosB and Roger Colbran for the generous gift of purified CaMKIIα.
References
- Ahmed SH, Koob GF. Transition from moderate to excessive drug intake: change in hedonic set point. Science. 1998;282:298–300. doi: 10.1126/science.282.5387.298. [DOI] [PubMed] [Google Scholar]
- Anderson SM, Famous KR, Sadri-Vakili G, Kumaresan V, Schmidt HD, Bass CE, Terwilliger EF, Cha JH, Pierce RC. CaMKII: a biochemical bridge linking accumbens dopamine and glutamate systems in cocaine seeking. Nat Neurosci. 2008;11:344–353. doi: 10.1038/nn2054. [DOI] [PubMed] [Google Scholar]
- Barrot M, Olivier JD, Perrotti LI, DiLeone RJ, Berton O, Eisch AJ, Impey S, Storm DR, Neve RL, Yin JC, Zachariou V, Nestler EJ. CREB activity in the nucleus accumbens shell controls gating of behavioral responses to emotional stimuli. Proc Natl Acad Sci U S A. 2002;99:11435–11440. doi: 10.1073/pnas.172091899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boudreau AC, Wolf ME. Behavioral sensitization to cocaine is associated with increased AMPA receptor surface expression in the nucleus accumbens. J Neurosci. 2005;25:9144–9151. doi: 10.1523/JNEUROSCI.2252-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brickey DA, Colbran RJ, Fong YL, Soderling TR. Expression and characterization of the alpha-subunit of Ca2+/calmodulin-dependent protein kinase II using the baculovirus expression system. Biochem Biophys Res Commun. 1990;173:578–584. doi: 10.1016/s0006-291x(05)80074-9. [DOI] [PubMed] [Google Scholar]
- Caine SB, Negus SS, Mello NK, Patel S, Bristow L, Kulagowski J, Vallone D, Saiardi A, Borrelli E. Role of dopamine D2-like receptors in cocaine self-administration: studies with D2 receptor mutant mice and novel D2 receptor antagonists. J Neurosci. 2002;22:2977–2988. doi: 10.1523/JNEUROSCI.22-07-02977.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caine SB, Thomsen M, Gabriel KI, Berkowitz JS, Gold LH, Koob GF, Tonegawa S, Zhang J, Xu M. Lack of self-administration of cocaine in dopamine D1 receptor knock-out mice. J Neurosci. 2007;27:13140–13150. doi: 10.1523/JNEUROSCI.2284-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carle TL, Ohnishi YN, Ohnishi YH, Alibhai IN, Wilkinson MB, Kumar A, Nestler EJ. Proteasome-dependent and -independent mechanisms for FosB destabilization: identification of FosB degron domains and implications for DeltaFosB stability. Eur J Neurosci. 2007;25:3009–3019. doi: 10.1111/j.1460-9568.2007.05575.x. [DOI] [PubMed] [Google Scholar]
- Chen J, Kelz MB, Zeng G, Sakai N, Steffen C, Shockett PE, Picciotto MR, Duman RS, Nestler EJ. Transgenic animals with inducible, targeted gene expression in brain. Mol Pharmacol. 1998;54:495–503. doi: 10.1124/mol.54.3.495. [DOI] [PubMed] [Google Scholar]
- Christoffel DJ, Golden SA, Dumitriu D, Robison AJ, Janssen WG, Ahn HF, Krishnan V, Reyes CM, Han MH, Ables JL, Eisch AJ, Dietz DM, Ferguson D, Neve RL, Greengard P, Kim Y, Morrison JH, Russo SJ. IκB kinase regulates social defeat stress-induced synaptic and behavioral plasticity. J Neurosci. 2011;31:314–321. doi: 10.1523/JNEUROSCI.4763-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colbran RJ. Inactivation of Ca2+/calmodulin-dependent protein kinase II by basal autophosphorylation. J Biol Chem. 1993;268:7163–7170. [PubMed] [Google Scholar]
- Colbran RJ. Protein phosphatases and calcium/calmodulin-dependent protein kinase II-dependent synaptic plasticity. J Neurosci. 2004;24:8404–8409. doi: 10.1523/JNEUROSCI.3602-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colbran RJ, Brown AM. Calcium/calmodulin-dependent protein kinase II and synaptic plasticity. Curr Opin Neurobiol. 2004;14:318–327. doi: 10.1016/j.conb.2004.05.008. [DOI] [PubMed] [Google Scholar]
- Colby CR, Whisler K, Steffen C, Nestler EJ, Self DW. Striatal cell type-specific overexpression of ΔFosB enhances incentive for cocaine. J Neurosci. 2003;23:2488–2493. doi: 10.1523/JNEUROSCI.23-06-02488.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Covington HE, 3rd, Maze I, LaPlant QC, Vialou VF, Ohnishi YN, Berton O, Fass DM, Renthal W, Rush AJ, 3rd, Wu EY, Ghose S, Krishnan V, Russo SJ, Tamminga C, Haggarty SJ, Nestler EJ. Antidepressant actions of histone deacetylase inhibitors. J Neurosci. 2009;29:11451–11460. doi: 10.1523/JNEUROSCI.1758-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dávalos A, Fernández-Hernando C, Sowa G, Derakhshan B, Lin MI, Lee JY, Zhao H, Luo R, Colangelo C, Sessa WC. Quantitative proteomics of caveolin-1-regulated proteins: characterization of polymerase i and transcript release factor/CAVIN-1 IN endothelial cells. Mol Cell Proteomics. 2010;9:2109–2124. doi: 10.1074/mcp.M110.001289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dumais A, Lesage AD, Alda M, Rouleau G, Dumont M, Chawky N, Roy M, Mann JJ, Benkelfat C, Turecki G. Risk factors for suicide completion in major depression: a case-control study of impulsive and aggressive behaviors in men. Am J Psychiatry. 2005;162:2116–2124. doi: 10.1176/appi.ajp.162.11.2116. [DOI] [PubMed] [Google Scholar]
- Grueter BA, Robison AJ, Neve RL, Nestler EJ, Malenka RC. ΔFosB differentially modulates nucleus accumbens direct and indirect pathway function. Proc Natl Acad Sci U S A. 2013;110:1923–1928. doi: 10.1073/pnas.1221742110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halt AR, Dallapiazza RF, Zhou Y, Stein IS, Qian H, Juntti S, Wojcik S, Brose N, Silva AJ, Hell JW. CaMKII binding to GluN2B is critical during memory consolidation. EMBO J. 2012;31:1203–1216. doi: 10.1038/emboj.2011.482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hiroi N, Brown JR, Haile CN, Ye H, Greenberg ME, Nestler EJ. FosB mutant mice: loss of chronic cocaine induction of Fos-related proteins and heightened sensitivity to cocaine's psychomotor and rewarding effects. Proc Natl Acad Sci U S A. 1997;94:10397–10402. doi: 10.1073/pnas.94.19.10397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito R, Robbins TW, Everitt BJ. Differential control over cocaine-seeking behavior by nucleus accumbens core and shell. Nat Neurosci. 2004;7:389–397. doi: 10.1038/nn1217. [DOI] [PubMed] [Google Scholar]
- Jorissen HJ, Ulery PG, Henry L, Gourneni S, Nestler EJ, Rudenko G. Dimerization and DNA-binding properties of the transcription factor DeltaFosB. Biochemistry. 2007;46:8360–8372. doi: 10.1021/bi700494v. [DOI] [PubMed] [Google Scholar]
- Jourdain P, Fukunaga K, Muller D. Calcium/calmodulin-dependent protein kinase II contributes to activity-dependent filopodia growth and spine formation. J Neurosci. 2003;23:10645–10649. doi: 10.1523/JNEUROSCI.23-33-10645.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelz MB, Chen J, Carlezon WA, Jr, Whisler K, Gilden L, Beckmann AM, Steffen C, Zhang YJ, Marotti L, Self DW, Tkatch T, Baranauskas G, Surmeier DJ, Neve RL, Duman RS, Picciotto MR, Nestler EJ. Expression of the transcription factor deltaFosB in the brain controls sensitivity to cocaine. Nature. 1999;401:272–276. doi: 10.1038/45790. [DOI] [PubMed] [Google Scholar]
- Klug JR, Mathur BN, Kash TL, Wang HD, Matthews RT, Robison AJ, Anderson ME, Deutch AY, Lovinger DM, Colbran RJ, Winder DG. Genetic inhibition of CaMKII in dorsal striatal medium spiny neurons reduces functional excitatory synapses and enhances intrinsic excitability. PLoS One. 2012;7:e45323. doi: 10.1371/journal.pone.0045323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knackstedt LA, Moussawi K, Lalumiere R, Schwendt M, Klugmann M, Kalivas PW. Extinction training after cocaine self-administration induces glutamatergic plasticity to inhibit cocaine seeking. J Neurosci. 2010;30:7984–7992. doi: 10.1523/JNEUROSCI.1244-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kourrich S, Thomas MJ. Similar neurons, opposite adaptations: psychostimulant experience differentially alters firing properties in accumbens core versus shell. J Neurosci. 2009;29:12275–12283. doi: 10.1523/JNEUROSCI.3028-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kourrich S, Klug JR, Mayford M, Thomas MJ. AMPAR-independent effect of striatal αCaMKII promotes the sensitization of cocaine reward. J Neurosci. 2012;32:6578–6586. doi: 10.1523/JNEUROSCI.6391-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaPlant Q, Chakravarty S, Vialou V, Mukherjee S, Koo JW, Kalahasti G, Bradbury KR, Taylor SV, Maze I, Kumar A, Graham A, Birnbaum SG, Krishnan V, Truong HT, Neve RL, Nestler EJ, Russo SJ. Role of nuclear factor kappaB in ovarian hormone-mediated stress hypersensitivity in female mice. Biol Psychiatry. 2009;65:874–880. doi: 10.1016/j.biopsych.2009.01.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee KW, Kim Y, Kim AM, Helmin K, Nairn AC, Greengard P. Cocaine-induced dendritic spine formation in D1 and D2 dopamine receptor-containing medium spiny neurons in nucleus accumbens. Proc Natl Acad Sci U S A. 2006;103:3399–3404. doi: 10.1073/pnas.0511244103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lisman J, Schulman H, Cline H. The molecular basis of CaMKII function in synaptic and behavioural memory. Nat Rev Neurosci. 2002;3:175–190. doi: 10.1038/nrn753. [DOI] [PubMed] [Google Scholar]
- Lobo MK, Covington HE, 3rd, Chaudhury D, Friedman AK, Sun H, Damez-Werno D, Dietz DM, Zaman S, Koo JW, Kennedy PJ, Mouzon E, Mogri M, Neve RL, Deisseroth K, Han MH, Nestler EJ. Cell type-specific loss of BDNF signaling mimics optogenetic control of cocaine reward. Science. 2010;330:385–390. doi: 10.1126/science.1188472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loweth JA, Baker LK, Guptaa T, Guillory AM, Vezina P. Inhibition of CaMKII in the nucleus accumbens shell decreases enhanced amphetamine intake in sensitized rats. Neurosci Lett. 2008;444:157–160. doi: 10.1016/j.neulet.2008.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loweth JA, Singer BF, Baker LK, Wilke G, Inamine H, Bubula N, Alexander JK, Carlezon WA, Jr, Neve RL, Vezina P. Transient overexpression of α-Ca2+/calmodulin-dependent protein kinase II in the nucleus accumbens shell enhances behavioral responding to amphetamine. J Neurosci. 2010;30:939–949. doi: 10.1523/JNEUROSCI.4383-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malinow R, Malenka RC. AMPA receptor trafficking and synaptic plasticity. Annu Rev Neurosci. 2002;25:103–126. doi: 10.1146/annurev.neuro.25.112701.142758. [DOI] [PubMed] [Google Scholar]
- Matsuzaki M, Honkura N, Ellis-Davies GC, Kasai H. Structural basis of long-term potentiation in single dendritic spines. Nature. 2004;429:761–766. doi: 10.1038/nature02617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayford M, Bach ME, Huang YY, Wang L, Hawkins RD, Kandel ER. Control of memory formation through regulated expression of a CaMKII transgene. Science. 1996;274:1678–1683. doi: 10.1126/science.274.5293.1678. [DOI] [PubMed] [Google Scholar]
- Maze I, Covington HE, 3rd, Dietz DM, LaPlant Q, Renthal W, Russo SJ, Mechanic M, Mouzon E, Neve RL, Haggarty SJ, Ren Y, Sampath SC, Hurd YL, Greengard P, Tarakhovsky A, Schaefer A, Nestler EJ. Essential role of the histone methyltransferase G9a in cocaine-induced plasticity. Science. 2010;327:213–216. doi: 10.1126/science.1179438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McClung CA, Nestler EJ. Regulation of gene expression and cocaine reward by CREB and DeltaFosB. Nat Neurosci. 2003;6:1208–1215. doi: 10.1038/nn1143. [DOI] [PubMed] [Google Scholar]
- Nestler EJ. Review. Transcriptional mechanisms of addiction: role of DeltaFosB. Philos Trans R Soc Lond B Biol Sci. 2008;363:3245–3255. doi: 10.1098/rstb.2008.0067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nye HE, Hope BT, Kelz MB, Iadarola M, Nestler EJ. Pharmacological studies of the regulation of chronic FOS-related antigen induction by cocaine in the striatum and nucleus accumbens. J Pharmacol Exp Ther. 1995;275:1671–1680. [PubMed] [Google Scholar]
- Okamoto K, Bosch M, Hayashi Y. The roles of CaMKII and F-actin in the structural plasticity of dendritic spines: a potential molecular identity of a synaptic tag? Physiology (Bethesda) 2009;24:357–366. doi: 10.1152/physiol.00029.2009. [DOI] [PubMed] [Google Scholar]
- Olausson P, Jentsch JD, Tronson N, Neve RL, Nestler EJ, Taylor JR. ΔFosB in the nucleus accumbens regulates food-reinforced instrumental behavior and motivation. J Neurosci. 2006;26:9196–9204. doi: 10.1523/JNEUROSCI.1124-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paxinos G, Watson C. The rat brain in stereotaxic coordinates. Ed 6. Amsterdam: Academic/Elsevier; 2007. [Google Scholar]
- Peakman MC, Colby C, Perrotti LI, Tekumalla P, Carle T, Ulery P, Chao J, Duman C, Steffen C, Monteggia L, Allen MR, Stock JL, Duman RS, McNeish JD, Barrot M, Self DW, Nestler EJ, Schaeffer E. Inducible, brain region-specific expression of a dominant negative mutant of c-Jun in transgenic mice decreases sensitivity to cocaine. Brain Res. 2003;970:73–86. doi: 10.1016/s0006-8993(03)02230-3. [DOI] [PubMed] [Google Scholar]
- Penzes P, Cahill ME, Jones KA, Srivastava DP. Convergent CaMK and RacGEF signals control dendritic structure and function. Trends Cell Biol. 2008;18:405–413. doi: 10.1016/j.tcb.2008.07.002. [DOI] [PubMed] [Google Scholar]
- Perrotti LI, Hadeishi Y, Ulery PG, Barrot M, Monteggia L, Duman RS, Nestler EJ. Induction of ΔFosB in reward-related brain structures after chronic stress. J Neurosci. 2004;24:10594–10602. doi: 10.1523/JNEUROSCI.2542-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perrotti LI, Weaver RR, Robison B, Renthal W, Maze I, Yazdani S, Elmore RG, Knapp DJ, Selley DE, Martin BR, Sim-Selley L, Bachtell RK, Self DW, Nestler EJ. Distinct patterns of DeltaFosB induction in brain by drugs of abuse. Synapse. 2008;62:358–369. doi: 10.1002/syn.20500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pickens CL, Airavaara M, Theberge F, Fanous S, Hope BT, Shaham Y. Neurobiology of the incubation of drug craving. Trends Neurosci. 2011;34:411–420. doi: 10.1016/j.tins.2011.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pierce RC, Quick EA, Reeder DC, Morgan ZR, Kalivas PW. Calcium-mediated second messengers modulate the expression of behavioral sensitization to cocaine. J Pharmacol Exp Ther. 1998;286:1171–1176. [PubMed] [Google Scholar]
- Quirion R, Robitaille Y, Martial J, Chabot JG, Lemoine P, Pilapil C, Dalpé M. Human brain receptor autoradiography using whole hemisphere sections: a general method that minimizes tissue artefacts. Synapse. 1987;1:446–454. doi: 10.1002/syn.890010508. [DOI] [PubMed] [Google Scholar]
- Robinson TE, Kolb B. Structural plasticity associated with exposure to drugs of abuse. Neuropharmacology. 2004;47(Suppl 1):33–46. doi: 10.1016/j.neuropharm.2004.06.025. [DOI] [PubMed] [Google Scholar]
- Robison AJ, Nestler EJ. Transcriptional and epigenetic mechanisms of addiction. Nat Rev Neurosci. 2011;12:623–637. doi: 10.1038/nrn3111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robison AJ, Bass MA, Jiao Y, MacMillan LB, Carmody LC, Bartlett RK, Colbran RJ. Multivalent interactions of calcium/calmodulin-dependent protein kinase II with the postsynaptic density proteins NR2B, densin-180, and alpha-actinin-2. J Biol Chem. 2005;280:35329–35336. doi: 10.1074/jbc.M502191200. [DOI] [PubMed] [Google Scholar]
- Ross PL, Huang YN, Marchese JN, Williamson B, Parker K, Hattan S, Khainovski N, Pillai S, Dey S, Daniels S, Purkayastha S, Juhasz P, Martin S, Bartlet-Jones M, He F, Jacobson A, Pappin DJ. Multiplexed protein quantitation in Saccharomyces cerevisiae using amine-reactive isobaric tagging reagents. Mol Cell Proteomics. 2004;3:1154–1169. doi: 10.1074/mcp.M400129-MCP200. [DOI] [PubMed] [Google Scholar]
- Russo SJ, Dietz DM, Dumitriu D, Morrison JH, Malenka RC, Nestler EJ. The addicted synapse: mechanisms of synaptic and structural plasticity in nucleus accumbens. Trends Neurosci. 2010;33:267–276. doi: 10.1016/j.tins.2010.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Self DW. The dopamine receptors. In: Neve KA, editor. New York: Humana; 2010. pp. 479–524. [Google Scholar]
- Singer BF, Loweth JA, Neve RL, Vezina P. Transient viral-mediated overexpression of alpha-calcium/calmodulin-dependent protein kinase II in the nucleus accumbens shell leads to long-lasting functional upregulation of alpha-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate receptors: dopamine type-1 receptor and protein kinase A dependence. Eur J Neurosci. 2010;31:1243–1251. doi: 10.1111/j.1460-9568.2010.07155.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strack S, McNeill RB, Colbran RJ. Mechanism and regulation of calcium/calmodulin-dependent protein kinase II targeting to the NR2B subunit of the N-methyl-d-aspartate receptor. J Biol Chem. 2000;275:23798–23806. doi: 10.1074/jbc.M001471200. [DOI] [PubMed] [Google Scholar]
- Ulery PG, Nestler EJ. Regulation of DeltaFosB transcriptional activity by Ser27 phosphorylation. Eur J Neurosci. 2007;25:224–230. doi: 10.1111/j.1460-9568.2006.05262.x. [DOI] [PubMed] [Google Scholar]
- Ulery PG, Rudenko G, Nestler EJ. Regulation of ΔFosB stability by phosphorylation. J Neurosci. 2006;26:5131–5142. doi: 10.1523/JNEUROSCI.4970-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulery-Reynolds PG, Castillo MA, Vialou V, Russo SJ, Nestler EJ. Phosphorylation of DeltaFosB mediates its stability in vivo. Neuroscience. 2009;158:369–372. doi: 10.1016/j.neuroscience.2008.10.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vialou V, Robison AJ, Laplant QC, Covington HE, 3rd, Dietz DM, Ohnishi YN, Mouzon E, Rush AJ, 3rd, Watts EL, Wallace DL, Iñiguez SD, Ohnishi YH, Steiner MA, Warren BL, Krishnan V, Bolaños CA, Neve RL, Ghose S, Berton O, Tamminga CA, Nestler EJ. DeltaFosB in brain reward circuits mediates resilience to stress and antidepressant responses. Nat Neurosci. 2010;13:745–752. doi: 10.1038/nn.2551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L, Lv Z, Hu Z, Sheng J, Hui B, Sun J, Ma L. Chronic cocaine-induced H3 acetylation and transcriptional activation of CaMKIIalpha in the nucleus accumbens is critical for motivation for drug reinforcement. Neuropsychopharmacology. 2010;35:913–928. doi: 10.1038/npp.2009.193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werme M, Messer C, Olson L, Gilden L, Thorén P, Nestler EJ, Brené S. ΔFosB regulates wheel running. J Neurosci. 2002;22:8133–8138. doi: 10.1523/JNEUROSCI.22-18-08133.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winstanley CA, LaPlant Q, Theobald DE, Green TA, Bachtell RK, Perrotti LI, DiLeone RJ, Russo SJ, Garth WJ, Self DW, Nestler EJ. ΔFosB induction in orbitofrontal cortex mediates tolerance to cocaine-induced cognitive dysfunction. J Neurosci. 2007;27:10497–10507. doi: 10.1523/JNEUROSCI.2566-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zachariou V, Bolanos CA, Selley DE, Theobald D, Cassidy MP, Kelz MB, Shaw-Lutchman T, Berton O, Sim-Selley LJ, Dileone RJ, Kumar A, Nestler EJ. An essential role for DeltaFosB in the nucleus accumbens in morphine action. Nat Neurosci. 2006;9:205–211. doi: 10.1038/nn1636. [DOI] [PubMed] [Google Scholar]
- Zhang R, Khoo MS, Wu Y, Yang Y, Grueter CE, Ni G, Price EE, Jr, Thiel W, Guatimosim S, Song LS, Madu EC, Shah AN, Vishnivetskaya TA, Atkinson JB, Gurevich VV, Salama G, Lederer WJ, Colbran RJ, Anderson ME. Calmodulin kinase II inhibition protects against structural heart disease. Nat Med. 2005;11:409–417. doi: 10.1038/nm1215. [DOI] [PubMed] [Google Scholar]