

Abstract
Asthma is a chronic inflammatory airway disease characterized by allergen-induced airway hyperresponsiveness, airway inflammation, and remodeling. Nitric oxide (NO) derived from constitutive and inducible enzymes affects many aspects of asthma physiopathology. Animal in vivo studies have indicated that inhibition of iNOS may play a central role in the modulation of these features, particularly extracellular matrix remodeling. Additionally, increases in iNOS-derived NO, observed in asthmatic patients, may lead to an increase in peroxynitrite and an imbalance of oxidant and antioxidant pathways. In addition, endogenous nitric oxide produced by constitutive enzymes may protect against the remodeling of the lung. Therefore, nitric oxide donors and/or iNOS inhibitors may have therapeutic potential in asthma treatment and can also be used with corticosteroids to counteract airway remodeling. This paper focuses on the pathophysiological role of nitric oxide, mainly derived from inducible isoforms, in the various pathologic mechanisms of allergic asthma and the importance of nitric oxide and/or arginase inhibitors in asthma treatment.
1. General Aspects Regarding Nitric Oxide
Nitric oxide (NO) has long been considered an atmospheric pollutant released by cigarette smoke and vehicles [1] that contributes to ozone layer destruction [2]. In 1980, Furchgott and Zawadzki [3] observed that the presence of intact endothelial cells is essential to arterial relaxation induced by acetylcholine, because a relaxant factor that was endothelium-dependent was released. Because the nitric oxide half life is extremely short, it was not until seven years later when Palmer et al. [4] and Ignarro et al. [5] showed that this relaxant factor, which was found in veins and arteries, was a form of nitric oxide. Therefore, nitric oxide began to be recognized as an essential molecule to the function of various organs in the human body.
Nitric oxide can be found in abundance in the central and peripheral nervous system [6, 7], acting as a classic neurotransmitter in the regulation of gastrointestinal motility, regional control of blood flux, and neuroendocrine function. In the brain, it affects memory formation and the response intensity to pain stimuli [8]. In addition, Kobzik et al. [9] demonstrated that nitric oxide is present in the skeletal musculature regulating its metabolism and muscle contractility. At high concentrations, it can have cytotoxic and cytostatic effects acting in the host defense against tumors and pathogens [10, 11].
NO has an unpaired electron, which makes it a free radical that can react with other molecules, such as oxygen, superoxide radicals, or transition metals [12]. Nitric oxide production begins when a group of L-arginine amino acid is broken down by oxidative enzymatic activity [13–16], generating nitric oxide and L-citrulline but not D-arginine [2, 10].
The L-arginine is transported inside the cells via a cationic amino acid transporter (CAT) and can be metabolized by two distinct groups of enzymes: nitric oxide synthase and arginase (Figure 1). Classically, arginase is an enzyme that acts in the urea cycle in the liver. However, this enzyme has been discovered in other cells that do not express the complete urea cycle, including the lung [17, 18].
Figure 1.
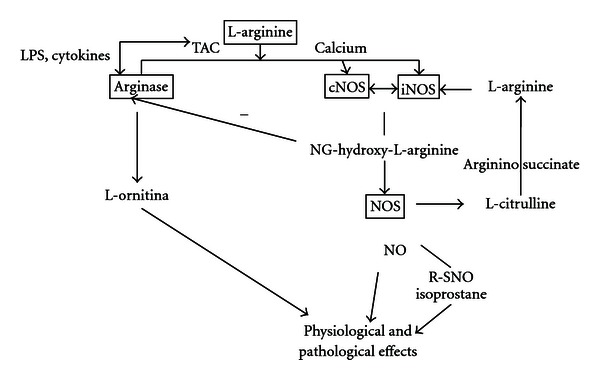
L-arginine, the common substrate to both the nitric oxide synthase and the arginase pathway. L-arginine is catalyzed by both arginase and nitric oxide. Arginase produces L-ornithine and urea that can act in various organs. In contrast, nitric oxide can be produced by both constitutive (cNOS) and inducible (iNOS) nitric oxide synthases and have pathophysiological roles important in health and diseases via the direct or indirect effects on oxidative stress production. Thus, arginase regulates the production of NO, and NO regulates the activity of arginase by substrate competition.
The nitric oxide is generated during the conversion of the amino acid L-arginine to L-citrulline by nitric oxide synthase (NOS) using the NG-hydroxyl-L-arginine as an intermediate that in turn inhibits the arginase activity [19]. The L-citrulline can be converted from the argininosuccinate into L-arginine, and arginase is able to metabolize L-arginine to L-ornithine (Figure 1) [11]. There are three nitric oxide enzyme isoforms, two constitutive and one inducible: the neuronal or type I (nNOS) [7, 20], the inducible isoform or type II (iNOS) [21–23], and the endothelial isoform or type 3 (eNOS) [24]. All the isoforms are flavoproteins that have tetrahydrobiopterin, heme, and an area that is homologous to cytochrome P450 reductase. They act as dioxygenases using an oxygen molecule and nicotinamide adenine dinucleotide phosphate-oxidase (NADPH) to transform the L-arginine into L-citrulline and release the nitric oxide. The tetrahydrobiopterin (BH4) is the main cofactor for all nitric oxide synthases isoforms, and in its absence, these enzymes can produce superoxide instead of nitric oxide [25, 26].
Although the sequence of amino acids of iNOS from mice macrophages was similar to the sequence found in rat cerebellum cNOS (51% of identity), these represent distinct enzymes [21].
The nNOS and eNOS, which are constitutive, are normally present in physiological situations. The constitutive nitric oxide synthase isoforms are activated by the increase in intracellular calcium. However, some authors have found increases of these isoforms in inflammatory situations such as asthma [27, 28]. In addition, these enzymes are calcium-dependent and can produce low quantities of nitric oxide. The eNOS was first discovered in endothelial cells [24], and the nitric oxide formed by this isoform is responsible for maintaining low vascular tone and preventing the vascular wall adherence of leucocytes and platelets [16].
The nNOS isoform is present mainly in neurons [29] and nerves; however, it has also been found in airway smooth muscle cells [30] and in epithelial cells [31]. The nitric oxide derived from this isoform acts as a neuromodulator and/or neuromediator in the central and periphery nerves and in the termination of nerves in the nonadrenergic noncholinergic nervous systems, particularly as an iNANC mediator [32–35], causing bronchodilation in smooth muscle airways cells. Ward et al. [36] studied human lung-tissue and observed that iNANC relaxation depends on nitric oxide release being higher in the trachea and relatively lower in the distal airways.
In contrast, the lipopolysaccharides (LPSs) and other cytokines increase the L-arginine transport, inducing NO production by both inducible NOS (iNOS) and arginase activity. In addition, nitric oxide can react with thiol groups, releasing S-nitrosothiols (R-SNO) that induces a variety of physiologic effects mediated by oxidative/nitrosative stress.
However, the inducible nitric oxide is present mainly in pathophysiological situations, and its stimulation produces high quantities of nitric oxide. The presence of this isoform was demonstrated initially in macrophages; however, it is well known now that iNOS can act in a variety of tissues and organs including the vascular endothelium, inducing multiple functions in the response to inflammation [37–39]. iNOS is inducible by LPS and other cytokines, such as TNF-α, IL-1 and IFN-γ, stimulating the formation of nanomolar quantities of nitric oxide [10, 40].
Many studies have evaluated the presence of iNOS in different organs, and now it is well established that this isoform can be present in different types of cells including epithelial cells [41], macrophages [42, 43], neutrophils [42, 44], eosinophils [45, 46], and mononucleated cells [46].
Among the various functions of nitric oxide derived from iNOS, it is important to note the proinflammatory role of helping in the inflammatory cell recruitment to a local injury site. Another important role that has been intensively discussed is that nitric oxide produced by inflammatory cells can contribute to the amplification of the inflammatory response [12, 47, 48].
The NO production can be inhibited by the blockage of nitric oxide synthase enzyme [10] or by the administration of an alternative substrate that inhibits the production of NO by these enzymes. Recently, many NO inhibitors have become commercially available that block the action of these enzymes. Some examples of these inhibitors are L-NG-monomethyl-arginine citrate (L-NMMA), L-NG-nitroarginine methyl ester (L-NAME), NG-nitro-L-arginine (L-NOARG), NG-iminoethyl-L-ornithine (L-NIO), L-NG-nitroarginine-p-nitroanilide (L-NAPNA), L-canavanin, hydroxycobalamin and aminoguanidine [49].
Aminoguanidine was considered to be a specific inhibitor of iNOS [50]; however, recently, its specificity to block iNOS has been debated [28]. Some years ago, Garvey et al. [51] demonstrated that 1400 W presented a high selectivity to iNOS inhibition both in vitro as in vivo, presenting toxic effects only in high doses (25 mg/Kg). Therefore, the 1400 W has been used as a pharmacological tool to study the role of nitric oxide produced by iNOS in various pathophysiological processes, including bronchial asthma.
As previously demonstrated in Figure 1, the substrate L-arginine is a common substrate for nitric oxide synthase and arginase. The blockage of one of these pathways can affect the activation of the other by substrate competition [11, 52]. Some authors suggested that the blockage of nitric oxide by L-NAME could be considered a pharmacological strategy to study the indirect role of arginases in several diseases [18, 52].
2. NO in Allergic Asthma Physiopathology
During the last few decades, the role of nitric oxide has been studied in the physiopathology of different inflammatory diseases [28, 51, 53–62].
Barnes [63] first suggested that NO produced by nNOS has beneficial effects in asthma, causing bronchodilation by relaxing airway smooth muscle via NANC nerves activation. However, nitric oxide produced by eNOS can induce vasodilation in the arterioles, with plasma extravasation and edema. High quantities of nitric oxide produced by iNOS can result in vasodilation, plasma extravasation, increase in mucus secretion and indirect activation by Th2 cells (mainly due to eosinophilic recruitment) to contribute to asthma physiopathology.
However, the role of nitric oxide in asthma is not to as simple as has been initially described because various in vitro and in vivo studies with animal experimental models and humans showed different results. The nitric oxide effects seem to be dependent on the animal mode and type of NO inhibitor as well as the parameters evaluated.
Some authors suggested that NOS activation can exert both beneficial and detrimental effects [18], such as altering relevant aspects of asthma pathology and participating in inflammatory cell recruitment [46–48, 55], airway responsiveness [52], and airway remodeling [11, 12, 64], as illustrated in Figure 2.
Figure 2.
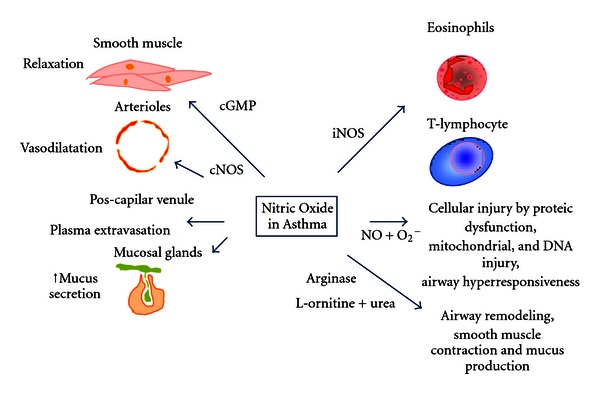
The dual effects of nitric oxide in asthma pathology. In asthma, nitric oxide can have both beneficial and adverse effects. The production of NO by constitutive isoforms can relax the smooth muscle of airways and vessels via cyclic GMP regulation inducing bronchodilation and vasodilation. However, by acting in the postcapillary venule, it can induce plasma extravasation. Nitric oxide can also regulate the mucosal glands, increasing the mucus secretion. High amounts of nitric oxide produced by iNOS in pathological situations induce the inflammatory cell chemotaxis, particularly recruiting eosinophils and T-lymphocytes to the lung. The reaction of nitric oxide with anion superoxide increases the oxidative stress pathway and can induce cellular injury by protein dysfunction or DNA injury and airway hyperresponsiveness. By substrate competition, nitric oxide can control the arginase pathway and induces airway remodeling, smooth muscle contraction and mucus production.
Nitric oxide was detected in the exhaled air in normal humans, but not in animals [65]. Various researchers demonstrated that the exhaled nitric oxide concentrations were increased in asthmatic patients [66–69] and in experimental models of chronic pulmonary inflammation [48, 70, 71]. Some authors suggested that the increase in exhaled NO can be due to increases in iNOS expression [22, 63, 66, 72]. It has been shown that L-NAME or aminoguanidine inhalation was able to reduce the amount of NO exhaled in healthy and asthmatic patients [73, 74]. Yates et al. [75] and Kharitonov et al. [72] noted that treatment with corticosteroids decreased the exhaled NO in asthmatics, but not in normal individuals. In addition, the exhaled NO was unaltered by the use of β 2 agonists [76]. Recently, Dweik et al. [77] have shown that fractional exhaled nitric oxide can be clinical used for the identification of the most reactive asthma phenotype. In this regard, these authors showed that among patients with severe asthma, FeNO can identify the most reactive and the worst asthma phenotype.
In contrast, Lim et al. [78] found no correlation between exhaled NO and the number of eosinophils found in airways, suggesting that NO does not directly modulate the eosinophilic inflammatory response in asthma. Thus, NO measurement should not be the only marker of disease control for asthmatic patients.
The exhaled nitric oxide has been proposed as an indirect marker of inflammation to be used in clinical practice. However, the efficacy of these measurements is a matter of controversy. Some authors showed that exhaled nitric oxide was a good marker of inflammation [79], which could be used to control the treatment of asthma and that it could be correlated with bronchial hyperreactivity in asthmatic children [80]. However, in a recently published study, Petsky et al. [81] showed that the tailoring of asthma treatment based on sputum eosinophils, but not in exhaled nitric oxide, was effective in decreasing asthma exacerbations. Despite these controversies, there is a consensus that exhaled nitric oxide is elevated in asthmatic patients, and repeated noninvasive and standardized monitoring contributes to the efficacy of the treatment of asthma because it is reduced in a dose-dependent manner by the use of inhaled corticosteroids [82].
3. Vascular Permeability
Nitric oxide plays an important role in controlling vascular tone [10]. There is evidence that vasodilation mediated by nitric oxide may promote edema formation via neural stimulation (see Figure 2). This mechanism is sensitive to treatment with tetrodotoxin, a drug that blocks calcium channels that are responsible for voltage-dependent propagation of neuronal action potential.
Nitric oxide modulates increased vascular permeability in the airways [83]. Ialenti et al. [84] showed that the inhibition of nitric oxide synthase by L-NAME, and L-NMMA inhibits the plasma leakage induced by carrageenan in rat skin and edema induced by carrageenan and dextran in the animal paw.
Erjefalt et al. [85] observed that NOS inhibitors decreased edema formation in the airway epithelium of guinea pigs. The NO-dependent changes in permeability may also be due to the release of other mediators, such as histamine, PAF and SP in the airways.
In an acute model, Mehta et al. [86] showed that chronic inhibition of nitric oxide synthase (72 hours) with L-NAME reduced plasma proteins. However, these results were not modified by aminoguanidine treatment, suggesting that vascular effects could be attributed, in particular, to constitutive isoforms.
Kuo et al. [87] suggested a link between substance P (SP) and NO production in airway microvascular permeability. Ziche et al. [88] demonstrated that SP can induce NO production by endothelial cells of postcapillary venules. Nguyen et al. [89] found that L-NAME blocks the increased vascular permeability induced by substance P, which was restored by L-arginine, the precursor of NO.
Our previous results [47, 48] showed that only chronic NO inhibition by L-NAME treatment attenuates the peribronchial edema formation around airways in an animal model of chronic allergic airway inflammation. In addition, when we evaluated the effects of specific iNOS inhibition in the same model, we did not observe any effects on plasma extravasation around airways, and in fact, this inhibition reduced the inflammation in the lung. Corroborating our findings that only cNOS played a role in the airway edema observed in asthma pathology, Bernareggi et al. [90] suggested that NO suppresses plasma leakage only in physiological situations and not when iNOS is highly expressed.
4. Airway Hyperresponsiveness
Donors of nitric oxide (nitrovasodilator) are smooth muscle relaxants, including airway smooth muscle in vitro [91, 92]. NO administration relaxes the smooth muscles of the trachea and airways and reduces methacholine-induced bronchoconstriction in guinea pigs and rabbits [93, 94]. Stimulation of nerve endings induces nNOS activation, and the NO release increases cyclic guanosine monophosphate (cGMP) levels in vascular smooth muscle inducing relaxation [10, 95].
The role of nitric oxide in the modulation of bronchoconstriction has been studied in animal models of both acute and chronic inflammatory responses. In acute models, Persson et al. [96] first showed that pretreatment with L-NAME increased inspiratory pressure, and exogenous nitric oxide administration was able to reverse this effect. Nijkamp et al. [97] showed that bronchoconstriction induced by intravenous administration of histamine in naïve guinea pigs was dose-dependently potentiated by L-NAME treatment. In addition, epithelium removal increased responsiveness to histamine. Other studies also suggested that the airway epithelium secretes nitric oxide [98].
Corroborating this hypothesis, De Sanctis et al. [27] observed that nNOS knockout mice led to a reduction in exhaled NO in both the basal and maximal response to methacholine responsiveness. De Boer et al. [99], studying isolated trachea of guinea pigs sensitized with ovalbumin, showed that endogenous nitric oxide deficiency induced by treatment with L-NAME increased bronchial responsiveness to methacholine after the immediate asthmatic reaction, which was reversed by L-arginine administration.
Although many studies emphasize the role of NO as a bronchodilator of the proximal airways, few studies have evaluated the role of NO as a modulator of distal airway tone. Dupuy et al. [93] found that inhaled NO can diffuse through the barrier of the bronchial epithelium and cause bronchodilation. In this study, the NO effects in the distal airways, which was measured by the complacency effect, was found only with high doses of inhaled nitric oxide, suggesting a larger role for this mediator in the proximal airways.
The importance of the distal lung in the pathophysiology of asthma has been evidenced recently in both in humans and in animal models [100, 101]. It has been suggested that small airway alterations may significantly contribute to the functional impairment in asthma, especially in the most severely affected patients [102, 103]. Our group showed that nitric oxide can act as a constrictor in areas other than the proximal airways. Both L-NAME and 1400 W reduced the lung-tissue mechanics in a model of inflammation induced by chronic challenges with ovalbumin [104, 105], suggesting that iNOS, which was probably produced by inflammatory cells that presented in the lung-tissue, regulated the constriction of the lung-tissue.
In contrast, some studies failed to show an important role of NO as a bronchodilator. Taylor et al. [106] studied patients with moderate persistent asthma and found that nebulization of L-NAME did not alter antigen-induced bronchoconstriction in patients with immediate and delayed responses. In this study, exhaled NO increased during the late response, and the L-NAME reduced exhaled NO levels by approximately 75%.
Another mechanism involved in the NO modulation of pulmonary mechanical alterations in asthma may be the interaction with the cholinergic neurotransmitters. In this regard, Belvisi et al. [34, 107] studied the effect of L-NAME on the NANC constriction response system by means of electrical stimulation in isolated tracheas of guinea pigs and humans. The results showed that L-NAME significantly increased cholinergic neurotransmission in a dose-dependent manner, but it was not effective in modulating constrictor responses of the NANC, suggesting that this effect was specific in cholinergic nerves.
Corroborating the interaction between nitric oxide and neurokinins, our group showed that capsaicin-sensitive sensory nerve fibers and neurokinins modulate nonneuronal nNOS expression by inflammatory and respiratory epithelial cells [108]. This increase in nNOS expression was also maintained for 14 days after capsaicin treatment and correlated with pulmonary mechanical reduction observed after capsaicin treatment when the lung content of neurokinin was already reduced [109].
Moreover, this study demonstrated that the acute inhibition of nitric oxide production with L-NAME treatment in sensitized mice restored the bronchodilator effects induced by neuropeptide depletion in both the proximal and distal airways [46].
Ricciardolo et al. [110] observed that the administration of L-NAME significantly increased the respiratory system resistance in response to the administration of bradykinin, but not to capsaicin in nonsensitized guinea pigs.
5. Leukocytes and Their Mediators
The modulation of the inflammatory process by NO production has been a matter of controversy. Nitric oxide may contribute to the recruitment of neutrophils, monocytes and eosinophils [23]. It is produced by some cells of the body's defense as neutrophils and macrophages [40].
Although the evidence from many studies has suggested that the inducible isoform is the major isoform involved in the inflammatory responses [42–45], recent studies suggest that the neuronal isoform also plays an important role in inflammation [111, 112]. It is known that there is eNOS and/or nNOS immunoreactivity in the various cells involved in inflammation, such as the lymphomononuclear cells [46], neutrophils [113, 114], eosinophils [46, 112, 115], and mast cells [116].
Nitric oxide may contribute to the extravasation of eosinophils from the circulation into the lung-tissue. Nitric oxide inhibition by L-NAME treatment does not influence eosinophilopoiesis [54].
Cardell et al. [117] showed that the inhibition of nitric oxide synthase reduces neutrophil recruitment and nasal secretion induced by LTB4 in the nasal mucosa of dogs. Trifilieff et al. [118] showed that iNOS promotes airway inflammation by increasing the expression of cytokines in an allergic asthma model in mice.
Prado et al. [47] studied the effects of acute inhibition of nitric oxide production in a model of chronic airway inflammation in guinea pigs. The authors demonstrated that only acute L-NAME treatment reduces eosinophils and lymphocytes in airway walls of these animals. The majority of the lymphomononuclear cells were represented by CD4+ cells. The chronic treatment did not alter this response. The same authors also showed that iNOS inhibition, due to treatment with a high selective inhibitor, was able to reduce the eosinophilic and lymphocytic recruitment to airway walls in guinea pigs with chronic allergic pulmonary inflammation.
Regarding this issue, Kobayashi et al. [28] showed that acute treatments with L-NAME, but not 1400 W, attenuated eosinophilic recruitment. These differences could be related to compensatory increases in iNOS, with no effects on eosinophilopoiesis [119] or differences in apoptosis modulation [120]. Taylor et al. [120] suggested that NO could have different effects on inflammatory cell apoptosis (anti- and proapoptotic properties), depending on the concentration, flux, and source of NO.
McCluskie et al. [121] evaluated the effects of NO in an aerosolized LPS-driven animal model of airway inflammation. Their study involved the assessment of exhaled NO and the inflammatory response in mice treated with L-NAME (100 mg/kg) or 1400 W (30 or 100 mg/kg) 2 h before and 1 h after the challenge. Treatment with either drug resulted in a decrease in exhaled NO and airway neutrophilia. Iijima et al. [122] also evaluated the relationship between eosinophil recruitment and NO in sensitized mice using 1400 W (1.0 mg/kg) and L-NAME (10 mg/kg) administered 0.5 h before and 8, 20, 32, and 44 h after OVA challenge. The authors demonstrated that both L-NAME and 1400 W reduced the number of eosinophils in the bronchoalveolar lavage fluid.
In addition, eNOS-derived NO has been shown to inhibit airway inflammation by suppressing the activation of NF-κB, thereby inhibiting both the expression of iNOS and the production of inflammatory cytokines [123–127].
Although there are discrepancies in the literature regarding the effects of nitric oxide in inflammation, there is a consensus that nitric oxide produced in high amounts has proinflammatory effects.
6. Airway Remodeling
Airway remodeling is characterized by structural changes in the airways and lung-tissue and involves hypertrophy and hyperplasia of the smooth muscle and goblet cells, extracellular matrix destruction and repair and neovascularization. Airway remodeling may be clinically important because it contributes to the irreversibility of lung function alterations observed in patients with asthma [128].
The role of nitric oxide in airway remodeling, particularly that associated with chronic inflammatory responses, has been recently emphasized [48, 129]. Gabazza et al. [130] showed a positive correlation between the concentrations of nitrite/nitrate in induced sputum from patients with asthma and bronchial remodeling, suggesting that NO is involved in the process of airway remodeling.
In vitro studies recently showed that DNA synthesis and proliferation of smooth muscle cells from human airways can be reduced by exogenous nitric oxide donors [30]. More recently, it was demonstrated that nitric oxide inhibits proliferation of smooth muscle cells of airways in G1 phase via cGMP-dependent mechanisms [131].
As illustrated in Figure 3, we first demonstrated that nitric oxide derived from constitutive isoforms acts by protecting the extracellular matrix alterations in an experimental model of allergic airway inflammation, whereas iNOS-derived NO contributes to the increase of both collagen and elastic fibers [48]. Interestingly, the effects of nitric oxide counteracting the remodeling can also be observed in lung distal parenchyma [105].
Figure 3.
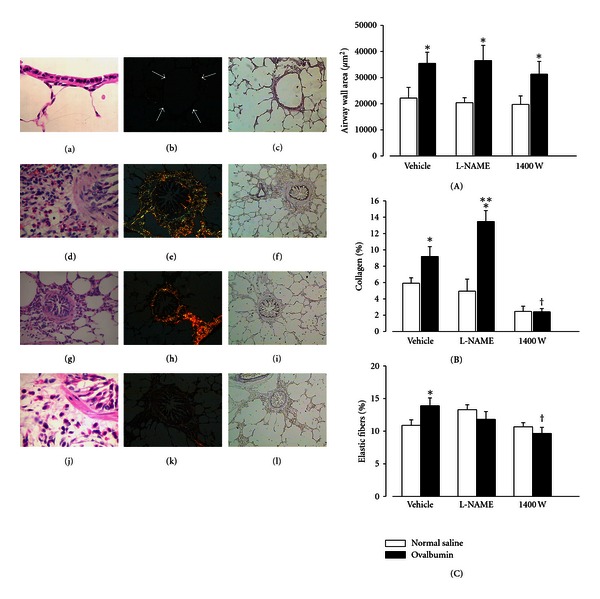
In the left panel, noncartilaginous guinea pig airways obtained from controls (a–c), ovalbumin-exposed (d–f), and ovalbumin-exposed animals treated with L-NAME (g–i) or 1400 W (j–l), a specific and highly selective iNOS inhibitor. We noted a weak yellow-greenish birefringence of the walls in the tissue section from control animals (b), coincident with the maintenance of the histoarchitecture of the extracellular matrix in H&E preparations (arrows) (a) and scant elastic fibers (c). In contrast, the airways of animals with airway inflammations induced by ovalbumin show an intense bronchoconstriction associated with peribronchial edema (d), an increase of birefringence in the airway wall (e) and in the elastic fibers content (f). L-NAME treatment decreased peribronchial edema (g), coincident with the increase of collagen content in the ECM (h), without interference in the elastic content (i). In contrast, 1400 W treatment attenuated the inflammatory cell infiltrate (j), collagen (k), and elastic (l) fiber deposition in airway walls without influencing peribronchial edema (original magnification in (b, c, e, f, h, i, k, l) ×200; (g) ×400; (a, d, j) ×1,000). In the right panel, the mean values of the total area (A), collagen (B), and elastic fibers (C) content in the airway walls of animals exposed to ovalbumin that received treatment with vehicle, L-NAME or 1400 W. *P < .05 compared with controls (open bars); **P < .05 compared with ovalbumin-exposed animals treated with vehicle and with 1400 W; and † P < .05 compared with ovalbumin-exposed animals treated with vehicle and L-NAME (closed bars). Reproduced with permission from Prado et al. [48].
Some authors have argued that arginase activation into myofibroblasts may act by increasing collagen production via production of L-ornithine [12, 18]. As previously described, arginase and nitric oxide synthase use a common substrate. Thus, chronic treatment with L-NAME is an indirect way to evaluate the role of arginase in the changes found in this model, particularly in airway remodeling.
Hogaboam et al. [64] studied a model of nonfibrotic lung granuloma. They showed that L-NAME induces an increase in mRNA levels of the C-C chemokine receptors, CCR2 and CCR3, and reduces macrophage chemoattractants protein-1 and eotaxin in isolated lung fibroblast culture, increasing the collagen content. Previous reports suggested that arginase could interfere with myofibroblast differentiation, increasing collagen production. Meurs et al. [52] also suggested that allergen-induced deficiency of cNOS-derived NO enhanced arginase activity. Finally, NO can have some effects on metalloproteinase activation and expression to modifying collagen degradation [132].
Considering that airway remodeling is not totally controlled by current treatment, the nitric oxide pathway may be important for the therapeutic control of airway remodeling.
7. Activation of Oxidative Stress Pathway
Various enzymatic reactions and chemical processes can generate endogenous reactive oxygen species (ROS). Nitric oxide can interact with reactive oxygen species to form reactive nitrogen species (RNS). The oxygen and nitrogen reactive species as well as nitric oxide are crucial to many physiological functions in the body, mainly as defense mechanisms. However, when cells are exposed to oxidative stress, characterized by an imbalance between pro- and anti-oxidative molecules, they have a variety of deleterious effects.
Some studies have also shown increased generation of superoxide radicals and ROS/RNS by alveolar macrophages (AMs), eosinophils, and polymorphonuclear leucocytes of asthmatic patients [133–135]. Both ROS and RNS produce many of the pathophysiological changes associated with asthma and may contribute to its pathogenesis.
The manner by which the iNOS-derived NO production leads to the potentiation of lung-tissue mechanics is poorly understood, but it may be related to the promotion of peroxynitrite production. This oxidant agent is formed by the interaction of NO and superoxide, which leads to lipid peroxidation and isoprostane (8-epi-PGF2α) generation. PGF-2α, the main member of the family of isoprostanes, was produced via the peroxidation of arachidonic acid [136]. Isoprostanes contribute to smooth muscle contractions acting through tyrosine kinase, Rho and Rho kinase, leading to decreased activity of myosin light-chain phosphatase, which augments the level of phosphorylated myosin light-chain and contraction [136].
We previously showed that there was a significant decrease in 8-epi-PGF2α density in lung-tissue from guinea pigs that were repeatedly exposed to ovalbumin and treated with 1400 W [105]. These findings corroborate the notion that the iNOS-derived NO production increases the oxidative stress pathway, which may have detrimental effects on lung-tissue with functional repercussions.
The induction of chronic airway inflammation in guinea pigs resulted in an increase in oxidative stress. We evaluated the oxidative stress in this study by 8-iso-PGF2α staining [137]. The 8-iso-PGF2α is the most well-characterized isoprostane that may act through a novel receptor closely related to, but distinct from, the thromboxane A2/PGH2 receptor, with a high specificity for 8-iso-PGF2α [138]. Considering the physiological effects of isoprostanes, Quaggiotto and Garg [139] demonstrated that 8-iso-PGE2 produced physiological effects that were similar to 8-iso- PGF2α, but at a reduced potency. In addition, Wood et al. [138] showed that 8-iso-PGF2α was increased three-to-four-fold in patients with persistent asthma compared with the normal group. For these reasons, the evaluation of 8-iso-PGF2α represents a valuable indicator of the oxidative stress pathway [137, 140].
8. Future Directions and Clinical Applications of Nitric Oxide in Asthma
Although many studies using asthma animal models have shown beneficial effects associated with iNOS inhibition and/or maintenance of endogenous NO production by constitutive isoforms, in humans, this therapeutic approach has not been consistently established and must be further investigated.
Hansel et al. [141] showed a potent reduction in exhaled NO over a period of 72 h both in mild asthmatics and in healthy volunteers using the semiselective iNOS inhibitor L-NIL. Singh et al. [142] showed that treatment of asthmatic patients with a selective iNOS inhibitor (GW274150) reduced NO levels in patients, but had no effect on airway reactivity and inflammation in patients with asthma, suggesting that targeting NO may not be an effective intervention in asthma treatment.
Considering that nitric oxide can have dual effects on asthma depending on the quantity, type of enzyme and location of release, it is important to consider the future drugs that may be donors of nitric oxide and/or inhibitors of nitric oxide, particularly the nitric oxide derived from iNOS. In this context, Turner et al. [143] showed in guinea pigs that TPI1020, a novel nitric oxide donating compound potentiates the bronchodilation activity of salbutamol. In addition, Jonasson et al. [144] showed that the association of nitric oxide and corticosteroids improved the protection against bronchoconstriction in an experimental murine model of asthma. The association of nitric oxide donors with corticosteroids and/or other current therapies used in asthma could be tested in clinical studies as airway remodeling therapies.
Regarding the future clinical application of nitric oxide pathway, it is also important to consider the clinical use of arginase inhibitors. Arginases are enzymes that use a common substrate together nitric oxide, and various studies have shown that their activity is involved in airway remodeling. Maarsingh et al. [145] have shown that arginase inhibition attenuated chronic inflammation induced by antigen, particularly reducing the increase in smooth muscle mass, the airway bronchial reactivity, and the hydroxyproline content that is involved in lung fibrosis. The authors clearly suggested the emerging effectiveness of arginase inhibitors in the treatment of asthma.
9. Conclusions
Based on the results described above, we conclude that several studies of respected research groups around the world have suggested an important role of nitric oxide in the various mechanisms involved in the pathogenesis of asthma. They suggested that endogenous nitric oxide, produced mainly by constitutive isoforms, has protective effects on bronchoconstriction and on remodeling, whereas nitric oxide produced by iNOS in high amounts is involved in the constriction, inflammation and remodeling observed in asthma. Collectively, the results discussed in this paper suggest that iNOS inhibition or NO donors were promising therapeutic strategies for asthma treatment, particularly for counteracting the remodeling response. However, presently, commercially available drugs for asthma treatment do not affect this aspect of asthma pathophysiology.
Acknowledgments
This work was supported by Fundação de Amparo de Pesquisa do Estado de São Paulo (FAPESP), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and Laboratório de Investigação Médica (LIM-20-FMUSP).
References
- 1.Norman V, Keith CH. Nitrogen oxides in tobacco smoke. Nature. 1965;205(4974):915–916. [Google Scholar]
- 2.Culotta E, Koshland DE. No news is good news. Science. 1992;258(5090):1862–1865. doi: 10.1126/science.1361684. [DOI] [PubMed] [Google Scholar]
- 3.Furchgott RF, Zawadzki JV. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature. 1980;288(5789):373–376. doi: 10.1038/288373a0. [DOI] [PubMed] [Google Scholar]
- 4.Palmer RMJ, Ferrige AG, Moncada S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature. 1987;327(6122):524–526. doi: 10.1038/327524a0. [DOI] [PubMed] [Google Scholar]
- 5.Ignarro LJ, Buga GM, Wood KS, Byrns RE, Chaudhuri G. Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proceedings of the National Academy of Sciences of the United States of America. 1987;84(24):9265–9269. doi: 10.1073/pnas.84.24.9265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Knowles RG, Palacios M, Palmer RMJ, Moncada S. Formation of nitric oxide from L-arginine in the central nervous system: a transduction mechanism for stimulation of the soluble guanylate cyclase. Proceedings of the National Academy of Sciences of the United States of America. 1989;86(13):5159–5162. doi: 10.1073/pnas.86.13.5159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bredt DS, Snyder SH. Nitric oxide mediates glutamate-linked enhancement of cGMP levels in the cerebellum. Proceedings of the National Academy of Sciences of the United States of America. 1989;86(22):9030–9033. doi: 10.1073/pnas.86.22.9030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Christopherson KS, Bredt DS. Nitric oxide in excitable tissues: physiological roles and disease. Journal of Clinical Investigation. 1997;100(10):2424–2429. doi: 10.1172/JCI119783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kobzik L, Reid MB, Bredt DS, Stamler JS. Nitric oxide in skeletal muscle. Nature. 1994;372(6506):546–548. doi: 10.1038/372546a0. [DOI] [PubMed] [Google Scholar]
- 10.Moncada S, Higgs A. The L-arginine-nitric oxide pathway. The New England Journal of Medicine. 1993;329(27):2002–2012. doi: 10.1056/NEJM199312303292706. [DOI] [PubMed] [Google Scholar]
- 11.Ricciardolo FLM, Sterk PJ, Gaston B, Folkerts G. Nitric oxide in health and disease of the respiratory system. Physiological Reviews. 2004;84(3):731–765. doi: 10.1152/physrev.00034.2003. [DOI] [PubMed] [Google Scholar]
- 12.Ricciardolo FLM. Multiple roles of nitric oxide in the airways. Thorax. 2003;58(2):175–182. doi: 10.1136/thorax.58.2.175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Moncada S, Palmer RMJ, Higgs EA. Biosynthesis of nitric oxide from L-arginine. A pathway for the regulation of cell function and communication. Biochemical Pharmacology. 1989;38(11):1709–1715. doi: 10.1016/0006-2952(89)90403-6. [DOI] [PubMed] [Google Scholar]
- 14.Tayeh MA, Marletta MA. Macrophage oxidation of L-arginine to nitric oxide, nitrite, and nitrate. Tetrahydrobiopterin is required as a cofactor. Journal of Biological Chemistry. 1989;264(33):19654–19658. [PubMed] [Google Scholar]
- 15.Michel T, Feron O. Nitric oxide synthases: which, where, how, and why? Journal of Clinical Investigation. 1997;100(9):2146–2152. doi: 10.1172/JCI119750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ignarro LJ. Nitric oxide as a unique signaling molecule in the vascular system: a historical overview. Journal of Physiology and Pharmacology. 2002;53(4):503–514. [PubMed] [Google Scholar]
- 17.Que LG, Kantrow SP, Jenkinson CP, Piantadosi CA, Huang YCT. Induction of arginase isoforms in the lung during hyperoxia. American Journal of Physiology. 1998;275(1):L96–L102. doi: 10.1152/ajplung.1998.275.1.L96. [DOI] [PubMed] [Google Scholar]
- 18.Meurs H, Maarsingh H, Zaagsma J. Arginase and asthma: novel insights into nitric oxide homeostasis and airway hyperresponsiveness. Trends in Pharmacological Sciences. 2003;24(9):450–455. doi: 10.1016/S0165-6147(03)00227-X. [DOI] [PubMed] [Google Scholar]
- 19.Maarsingh H, Zuidhof AB, Bos IST, et al. Arginase inhibition protects against allergen-induced airway obstruction, hyperresponsiveness, and inflammation. American Journal of Respiratory and Critical Care Medicine. 2008;178(6):565–573. doi: 10.1164/rccm.200710-1588OC. [DOI] [PubMed] [Google Scholar]
- 20.Bredt DS, Hwang PM, Snyder SH. Localization of nitric oxide synthase indicating a neural role for nitric oxide. Nature. 1990;347(6295):768–770. doi: 10.1038/347768a0. [DOI] [PubMed] [Google Scholar]
- 21.Xie Q, Cho HJ, Calaycay J, et al. Cloning and characterization of inducible nitric oxide synthase from mouse macrophages. Science. 1992;256(5054):225–228. doi: 10.1126/science.1373522. [DOI] [PubMed] [Google Scholar]
- 22.Hamid Q, Springall DR, Riveros-Moreno V, et al. Induction of nitric oxide synthase in asthma. Lancet. 1993;342(8886-8887):1510–1513. doi: 10.1016/s0140-6736(05)80083-2. [DOI] [PubMed] [Google Scholar]
- 23.Robbins RA. Nitric oxide. In: Barnes PJ, Grunstein MM, Leff AR, editors. Asthma. Philadelphia, Pa, USA: Lippincott-Raven; 1997. pp. 695–705. [Google Scholar]
- 24.Lamas S, Marsden PA, Li GK, Tempst P, Michel T. Endothelial nitric oxide synthase: molecular cloning and characterization of a distinct constitutive enzyme isoform. Proceedings of the National Academy of Sciences of the United States of America. 1992;89(14):6348–6352. doi: 10.1073/pnas.89.14.6348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Guzik TJ, Korbut R, Adamek-Guzik T. Nitric oxide and superoxide in inflammation and immune regulation. Journal of Physiology and Pharmacology. 2003;54(4):469–487. [PubMed] [Google Scholar]
- 26.Alp NJ, Mussa S, Khoo J, et al. Tetrahydrobiopterin-dependent preservation of nitric oxide-mediated endothelial function in diabetes by targeted transgenic GTP-cyclohydrolase I overexpression. Journal of Clinical Investigation. 2003;112(5):725–735. doi: 10.1172/JCI17786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.De Sanctis GT, Mehta S, Kobzik L, et al. Contribution of type I NOS to expired gas NO and bronchial responsiveness in mice. American Journal of Physiology. 1997;273(4):L883–L888. doi: 10.1152/ajplung.1997.273.4.L883. [DOI] [PubMed] [Google Scholar]
- 28.Kobayashi K, Nishimura Y, Yamashita T, Nishiuma T, Satouchi M, Yokoyama M. The effect of overexpression of endothelial nitric oxide synthase on eosinophilic lung inflammation in a murine model. International Immunopharmacology. 2006;6(7):1040–1052. doi: 10.1016/j.intimp.2005.09.016. [DOI] [PubMed] [Google Scholar]
- 29.Vizzard MA, Erdman SL, de Groat WC. Increased expression of neuronal nitric oxide synthase in dorsal root ganglion neurons after systemic capsaicin administration. Neuroscience. 1995;67(1):1–5. doi: 10.1016/0306-4522(95)00137-8. [DOI] [PubMed] [Google Scholar]
- 30.Patel HJ, Belvisi MG, Donnelly LE, Yacoub MH, Chung KF, Mitchell JA. Constitutive expressions of type I NOS in human airway smooth muscle cells: evidence for an antiproliferative role. FASEB Journal. 1999;13(13):1810–1816. doi: 10.1096/fasebj.13.13.1810. [DOI] [PubMed] [Google Scholar]
- 31.Asano K, Chee CBE, Gaston B, et al. Constitutive and inducible nitric oxide synthase gene expression, regulation, and activity in human lung epithelial cells. Proceedings of the National Academy of Sciences of the United States of America. 1994;91(21):10089–10093. doi: 10.1073/pnas.91.21.10089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bult H, Boeckxstaens GE, Pelckmans PA, Jordaens FH, Van Maercke YM, Herman AG. Nitric oxide as an inhibitory non-adrenergic non-cholinergic neurotransmitter. Nature. 1990;345(6273):346–347. doi: 10.1038/345346a0. [DOI] [PubMed] [Google Scholar]
- 33.Li CG, Rand MJ. Evidence that part of the NANC relaxant response of guinea-pig trachea to electrical field stimulation is mediated by nitric oxide. British Journal of Pharmacology. 1991;102(1):91–94. doi: 10.1111/j.1476-5381.1991.tb12137.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Belvisi MG, Stretton D, Barnes PJ. Nitric oxide as an endogenous modulator of cholinergic neurotransmission in guinea-pig airways. European Journal of Pharmacology. 1991;198(2-3):219–221. doi: 10.1016/0014-2999(91)90626-2. [DOI] [PubMed] [Google Scholar]
- 35.Imasaki T, Kobayashi H, Hataishi R, Hayashi I, Tomita T, Majima M. Nitric oxide is generated in smooth muscle layer by neurokinin A and counteracts constriction in Guinea pig airway. Nitric Oxide. 2001;5(5):465–474. doi: 10.1006/niox.2001.0361. [DOI] [PubMed] [Google Scholar]
- 36.Ward JK, Barnes PJ, Springall DR, et al. Distribution of human iNANC bronchodilator and nitric oxide-immunoreactive nerves. American Journal of Respiratory Cell and Molecular Biology. 1995;13(2):175–184. doi: 10.1165/ajrcmb.13.2.7542897. [DOI] [PubMed] [Google Scholar]
- 37.Meng QH, Polak JM, Edgar AJ, et al. Neutrophils enhance expression of inducible nitric oxide synthase in human normal but not cystic fibrosis bronchial epithelial cells. Journal of Pathology. 2000;190(2):126–132. doi: 10.1002/(SICI)1096-9896(200002)190:2<126::AID-PATH500>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- 38.Birrell MA, McCluskie K, Haddad EB, et al. Pharmacological assessment of the nitric-oxide synthase isoform involved in eosinophilic inflammation in a rat model of sephadex-induced airway inflammation. Journal of Pharmacology and Experimental Therapeutics. 2003;304(3):1285–1291. doi: 10.1124/jpet.102.044339. [DOI] [PubMed] [Google Scholar]
- 39.Offer S, Shoseyov D, Bibi H, Eliraz A, Madar Z. A leukotriene receptor antagonist modulates iNOS in the lung and in a leukotriene-free cell model. Nitric Oxide. 2003;9(1):10–17. doi: 10.1016/s1089-8603(03)00047-8. [DOI] [PubMed] [Google Scholar]
- 40.Moncada S, Palmer RMJ, Higgs EA. Nitric oxide: physiology, pathophysiology, and pharmacology. Pharmacological Reviews. 1991;43(2):109–142. [PubMed] [Google Scholar]
- 41.Guo FH, De Raeve HR, Rice TW, Stuehr DJ, Thunnissen FBJM, Erzurum SC. Continuous nitric oxide synthesis by inducible nitric oxide synthase in normal human airway epithelium in vivo. Proceedings of the National Academy of Sciences of the United States of America. 1995;92(17):7809–7813. doi: 10.1073/pnas.92.17.7809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Iyengar R, Stuehr DJ, Marletta MA. Macrophage synthesis of nitrite, nitrate, and N-nitrosamines: precursors and role of the respiratory burst. Proceedings of the National Academy of Sciences of the United States of America. 1987;84(18):6369–6373. doi: 10.1073/pnas.84.18.6369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Watanabe M, Kakuta F. Expression and localization of the inducible isoform of nitric oxide synthase in polyps. Nippon Jibiinkoka Gakkai Kaiho. 2002;105(8):873–881. doi: 10.3950/jibiinkoka.105.873. [DOI] [PubMed] [Google Scholar]
- 44.McCall TB, Palmer RMJ, Moncada S. Induction of nitric oxide synthase in rat peritoneal neutrophils and its inhibition by dexamethasone. European Journal of Immunology. 1991;21(10):2523–2527. doi: 10.1002/eji.1830211032. [DOI] [PubMed] [Google Scholar]
- 45.Cerchiaro GA, Scavone C, Texeira S, Sannomiya P. Inducible nitric oxide synthase in rat neutrophils: Role of insulin. Biochemical Pharmacology. 2001;62(3):357–362. doi: 10.1016/s0006-2952(01)00672-4. [DOI] [PubMed] [Google Scholar]
- 46.Prado CM, Leick-Maldonado EA, Arata V, Kasahara DI, Martins MA, Tibério IFLC. Neurokinins and inflammatory cell iNOS expression in guinea pigs with chronic allergic airway inflammation. American Journal of Physiology. 2005;288(4):L741–L748. doi: 10.1152/ajplung.00208.2004. [DOI] [PubMed] [Google Scholar]
- 47.Prado CM, Leick-Maldonado EA, Kasahara DI, Capelozzi VL, Martins MA, Tibério IFLC. Effects of acute and chronic nitric oxide inhibition in an experimental model of chronic pulmonary allergic inflammation in guinea pigs. American Journal of Physiology. 2005;289(4):L677–L683. doi: 10.1152/ajplung.00010.2005. [DOI] [PubMed] [Google Scholar]
- 48.Prado CM, Leick-Maldonado EA, Yano L, et al. Effects of nitric oxide synthases in chronic allergic airway inflammation and remodeling. American Journal of Respiratory Cell and Molecular Biology. 2006;35(4):457–465. doi: 10.1165/rcmb.2005-0391OC. [DOI] [PubMed] [Google Scholar]
- 49.Rees DD, Palmer RMJ, Moncada S. Role of endothelium-derived nitric oxide in the regulation of blood pressure. Proceedings of the National Academy of Sciences of the United States of America. 1989;86(9):3375–3378. doi: 10.1073/pnas.86.9.3375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Baddedge RC, Moore PK, Gathen Z, Hart SL. L-NG-nitroarginine p-nitroanilide (L-NAPNA): a selective inhibitor of nitric oxide synthase in the brain. British Journal of Pharmacology. 1993;107:194 pages. [Google Scholar]
- 51.Garvey EP, Oplinger JA, Furfine ES, et al. 1400W is a slow, tight binding, and highly selective inhibitor of inducible nitric-oxide synthase in vitro and in vivo . Journal of Biological Chemistry. 1997;272(8):4959–4963. doi: 10.1074/jbc.272.8.4959. [DOI] [PubMed] [Google Scholar]
- 52.Meurs H, McKay S, Maarsingh H, et al. Increased arginase activity underlies allergen-induced deficiency of cNos-derived nitric oxide and airway hyperresponsiveness. British Journal of Pharmacology. 2002;136(3):391–398. doi: 10.1038/sj.bjp.0704725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Van Camp JR, Yian C, Lupinetti FM. Regulation of pulmonary vascular resistance by endogenous and exogenous nitric oxide. Annals of Thoracic Surgery. 1994;58(4):1025–1030. doi: 10.1016/0003-4975(94)90448-0. [DOI] [PubMed] [Google Scholar]
- 54.Feder LS, Stelts D, Chapman RW, et al. Role of nitric oxide on eosinophilic lung inflammation in allergic mice. American Journal of Respiratory Cell and Molecular Biology. 1997;17(4):436–442. doi: 10.1165/ajrcmb.17.4.2845. [DOI] [PubMed] [Google Scholar]
- 55.Schuiling M, Meurs H, Zuidhof AB, Venema N, Zaagsma J. Dual action of iNOS derived nitric oxide in allergen-induced airway hyperreactivity in conscious, unrestrained guinea pigs. American Journal of Respiratory and Critical Care Medicine. 1998;158(5):1442–1449. doi: 10.1164/ajrccm.158.5.9803027. [DOI] [PubMed] [Google Scholar]
- 56.Ferreira HHA, Bevilacqua E, Gagioti SM, et al. Nitric oxide modulates eosinophil infiltration in antigen-induced airway inflammation in rats. European Journal of Pharmacology. 1998;358(3):253–259. doi: 10.1016/s0014-2999(98)00575-5. [DOI] [PubMed] [Google Scholar]
- 57.Fagan KA, Tyler RC, Sato K, et al. Relative contributions of endothelial, inducible, and neuronal NOS to tone in the murine pulmonary circulation. American Journal of Physiology. 1999;277(3, part 1):L472–L478. doi: 10.1152/ajplung.1999.277.3.L472. [DOI] [PubMed] [Google Scholar]
- 58.De Boer J, Meurs H, Flendrig L, Koopal M, Zaagsma J. Role of nitric oxide and superoxide in allergen-induced airway hyperreactivity after the late asthmatic reaction in guinea-pigs. British Journal of Pharmacology. 2001;133(8):1235–1242. doi: 10.1038/sj.bjp.0704191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Lührs H, Papadopoulos T, Schmidt HHHW, Menzel T. Type I nitric oxide synthase in the human lung is predominantly expressed in capillary endothelial cells. Respiration Physiology. 2002;129(3):367–374. doi: 10.1016/s0034-5687(01)00323-1. [DOI] [PubMed] [Google Scholar]
- 60.Koarai A, Ichinose M, Sugiura H, et al. iNOS depletion completely diminishes reactive nitrogen-species formation after an allergic response. European Respiratory Journal. 2002;20(3):609–616. doi: 10.1183/09031936.02.00274902. [DOI] [PubMed] [Google Scholar]
- 61.Kenyon NJ, Gohil K, Last JA. Susceptibility to ovalbumin-induced airway inflammation and fibrosis in inducible nitric oxide synthetase-deficient mice: Mechanisms and consequences. Toxicology and Applied Pharmacology. 2003;191(1):2–11. doi: 10.1016/s0041-008x(03)00227-8. [DOI] [PubMed] [Google Scholar]
- 62.Poon BY, Raharjo E, Patel KD, Tavener S, Kubes P. Complexity of inducible nitric oxide synthase: cellular source determines benefit versus toxicity. Circulation. 2003;108(9):1107–1112. doi: 10.1161/01.CIR.0000086321.04702.AC. [DOI] [PubMed] [Google Scholar]
- 63.Barnes PJ. NO or no NO in asthma? Thorax. 1996;51:218–220. doi: 10.1136/thx.51.2.218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Hogaboam CM, Gallinat CS, Bone-Larson C, et al. Collagen deposition in a non-fibrotic lung granuloma model after nitric oxide inhibition. The American Journal of Pathology. 1998;153:1861–1872. doi: 10.1016/S0002-9440(10)65700-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Gustafsson LE, Leone AM, Persson MG, Wiklund NP, Moncada S. Endogenous nitric oxide is present in the exhaled air of rabbits, guinea pigs and humans. Biochemical and Biophysical Research Communications. 1991;181(2):852–857. doi: 10.1016/0006-291x(91)91268-h. [DOI] [PubMed] [Google Scholar]
- 66.Kharitonov SA, Yates D, Robbins RA, Logan-Sinclair R, Shinebourne EA, Barnes PJ. Increased nitric oxide in exhaled air of asthmatic patients. Lancet. 1994;343(8890):133–135. doi: 10.1016/s0140-6736(94)90931-8. [DOI] [PubMed] [Google Scholar]
- 67.Persson MG, Zetterström O, Agrenius V, Ihre E, Gustafsson LE. Single-breath nitric oxide measurements in asthmatic patients and smokers. Lancet. 1994;343(8890):146–147. doi: 10.1016/s0140-6736(94)90935-0. [DOI] [PubMed] [Google Scholar]
- 68.Belvisi M, Barnes PJ, Larkin S, et al. Nitric oxide synthase activity is elevated in inflammatory lung disease in humans. European Journal of Pharmacology. 1995;283(1–3):255–258. doi: 10.1016/0014-2999(95)00421-g. [DOI] [PubMed] [Google Scholar]
- 69.Deykin A, Halpern O, Massaro AF, Drazen JM, Israel E. Expired nitric oxide after bronchoprovocation and repeated spirometry in patients with asthma. American Journal of Respiratory and Critical Care Medicine. 1998;157(3, part 1):769–775. doi: 10.1164/ajrccm.157.3.9707114. [DOI] [PubMed] [Google Scholar]
- 70.Mehta S, Lilly CM, Rollenhagen JE, Haley KJ, Asano K, Drazen JM. Acute and chronic effects of allergic airway inflammation on pulmonary nitric oxide production. American Journal of Physiology. 1997;272(1):L124–L131. doi: 10.1152/ajplung.1997.272.1.L124. [DOI] [PubMed] [Google Scholar]
- 71.Leick-Maldonado EA, Kay FU, Leonhardt MC, et al. Comparison of glucocorticoid and cysteinyl leukotriene receptor antagonist treatments in an experimental model of chronic airway inflammation in guinea-pigs. Clinical and Experimental Allergy. 2004;34(1):145–152. doi: 10.1111/j.1365-2222.2004.01854.x. [DOI] [PubMed] [Google Scholar]
- 72.Kharitonov SA, Yates DH, Barnes PJ. Inhaled glucocorticoids decrease nitric oxide in exhaled air of asthmatic patients. American Journal of Respiratory and Critical Care Medicine. 1996;153(1):454–457. doi: 10.1164/ajrccm.153.1.8542158. [DOI] [PubMed] [Google Scholar]
- 73.Gómez FP, Pallí GM, Barberà JA, Roca J, Rodríguez-Roisin R. Measurement of exhaled nitric oxide in healthy subjects. Medicina Clinica. 1998;111(1):1–5. [PubMed] [Google Scholar]
- 74.Brindicci C, Ito K, Barnes PJ, Kharitonov SA. Effect of an inducible nitric oxide synthase inhibitor on differential flow-exhaled nitric oxide in asthmatic patients and healthy volunteers. Chest. 2007;132(2):581–588. doi: 10.1378/chest.06-3046. [DOI] [PubMed] [Google Scholar]
- 75.Yates DH, Kharitonov SA, Thomas PS, Barnes PJ. Endogenous nitric oxide is decreased in asthmatic patients by an inhibitor of inducible nitric oxide synthase. American Journal of Respiratory and Critical Care Medicine. 1996;154(1):247–250. doi: 10.1164/ajrccm.154.1.8680689. [DOI] [PubMed] [Google Scholar]
- 76.Yates DH, Kharitonov SA, Barnes PJ. Effect of short- and long-acting inhaled β2-agonists on exhaled nitric oxide in asthmatic patients. European Respiratory Journal. 1997;10(7):1483–1488. doi: 10.1183/09031936.97.10071483. [DOI] [PubMed] [Google Scholar]
- 77.Dweik RA, Sorkness RL, Wenzel S, et al. Use of exhaled nitric oxide measurement to identify a reactive, at-risk phenotype among patients with asthma. American Journal of Respiratory and Critical Care Medicine. 2010;181(10):1033–1041. doi: 10.1164/rccm.200905-0695OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lim S, Jatakanon A, Meah S, Oates T, Chung KF, Barnes PJ. Relationship between exhaled nitric oxide and mucosal eosinophilic inflammation in mild to moderately severe asthma. Thorax. 2000;55(3):184–188. doi: 10.1136/thorax.55.3.184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Pisi R, Aiello M, Tzani P, Marangio E, Olivieri D, Chetta A. Measurement of fractional exhaled nitric oxide by a new portable device: comparison with the standard technique. Journal of Asthma. 2010;47(7):805–809. doi: 10.3109/02770903.2010.485667. [DOI] [PubMed] [Google Scholar]
- 80.Ciprandi G, Tosca MA, Capasso M. Exhaled nitric oxide in children with allergic rhinitis and/or asthma: a relationship with bronchial hyperreactivity. Journal of Asthma. 2010;47(10):1142–1147. doi: 10.3109/02770903.2010.527026. [DOI] [PubMed] [Google Scholar]
- 81.Petsky HL, Cates CJ, Lasserson TJ, et al. A systematic review and meta-analysis: tailoring asthma treatment on eosinophilic markers (exhaled nitric oxide or sputum eosinophils) doi: 10.1136/thx.2010.135574. Thorax, 2010. In press. [DOI] [PubMed] [Google Scholar]
- 82.Kharitonov SA, Barnes PJ. Clinical aspects of exhaled nitric oxide. European Respiratory Journal. 2000;16(4):781–792. doi: 10.1183/09031936.00.16478100. [DOI] [PubMed] [Google Scholar]
- 83.Alving K, Fornhem C, Weitzberg E, Lundberg JM. Nitric oxide mediates cigarette smoke-induced vasodilatory responses in the lung. Acta Physiologica Scandinavica. 1992;146(3):407–408. doi: 10.1111/j.1748-1716.1992.tb09439.x. [DOI] [PubMed] [Google Scholar]
- 84.Ialenti A, Ianaro A, Moncada S, Di Rosa M. Modulation of acute inflammation by endogenous nitric oxide. European Journal of Pharmacology. 1992;211(2):177–182. doi: 10.1016/0014-2999(92)90526-a. [DOI] [PubMed] [Google Scholar]
- 85.Erjefalt JS, Erjefalt I, Sundler F, Persson CGA. Mucosal nitric oxide may tonically suppress airways plasma exudation. American Journal of Respiratory and Critical Care Medicine. 1994;150(1):227–232. doi: 10.1164/ajrccm.150.1.8025753. [DOI] [PubMed] [Google Scholar]
- 86.Mehta S, Boudreau J, Lilly CM, Drazen JM. Endogenous pulmonary nitric oxide in the regulation of airway microvascular leak. American Journal of Physiology. 1998;275(5):L961–L968. doi: 10.1152/ajplung.1998.275.5.L961. [DOI] [PubMed] [Google Scholar]
- 87.Kuo HP, Liu S, Barnes PJ. The effect of endogenous nitric oxide on neurogenic plasma exudation in guinea-pig airways. European Journal of Pharmacology. 1992;221(2-3):385–388. doi: 10.1016/0014-2999(92)90728-m. [DOI] [PubMed] [Google Scholar]
- 88.Ziche M, Morbidelli L, Parenti A, Amerini S, Granger HJ, Maggi CA. Substance P increases cyclic GMP levels on coronary postcapillary venular endothelial cells. Life Sciences. 1993;53(14):PL229–PL234. doi: 10.1016/0024-3205(93)90556-i. [DOI] [PubMed] [Google Scholar]
- 89.Nguyen LS, Villablanca AC, Rutledge JC. Substance P increases microvascular permeability via nitric oxide-mediated convective pathways. American Journal of Physiology. 1995;268(4):R1060–R1068. doi: 10.1152/ajpregu.1995.268.4.R1060. [DOI] [PubMed] [Google Scholar]
- 90.Bernareggi M, Mitchell JA, Barnes PJ, Belvisi MC. Dual action of nitric oxide on airway plasma leakage. American Journal of Respiratory and Critical Care Medicine. 1997;155(3):869–874. doi: 10.1164/ajrccm.155.3.9117019. [DOI] [PubMed] [Google Scholar]
- 91.Kishen R, Pleuvry BJ. Some actions of sodium nitroprusside and glyceryl trinitrate on guinea-pig isolated trachealis muscle. Journal of Pharmacy and Pharmacology. 1985;37(7):502–504. doi: 10.1111/j.2042-7158.1985.tb03052.x. [DOI] [PubMed] [Google Scholar]
- 92.Nijkamp FP, Folkerts G. Nitric oxide and bronchial reactivity. Clinical and Experimental Allergy. 1994;24(10):905–914. doi: 10.1111/j.1365-2222.1994.tb02721.x. [DOI] [PubMed] [Google Scholar]
- 93.Dupuy PM, Shore SA, Drazen JM, Frostell C, Hill WA, Zapol WM. Bronchodilator action of inhaled nitric oxide in guinea pigs. Journal of Clinical Investigation. 1992;90(2):421–428. doi: 10.1172/JCI115877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Högman M, Frostell C, Arnberg H, Hedenstierna G. Inhalation of nitric oxide modulates methacholine-induced bronchoconstriction in the rabbit. European Respiratory Journal. 1993;6(2):177–180. [PubMed] [Google Scholar]
- 95.Knowles RG, Moncada S. Nitric oxide as a signal in blood vessels. Trends in Biochemical Sciences. 1992;17(10):399–402. doi: 10.1016/0968-0004(92)90008-w. [DOI] [PubMed] [Google Scholar]
- 96.Persson MG, Friberg SG, Hedqvist P, Gustafsson LE. Endogenous nitric oxide counteracts antigen-induced bronchoconstriction. European Journal of Pharmacology. 1993;249(3):R7–R8. doi: 10.1016/0014-2999(93)90532-m. [DOI] [PubMed] [Google Scholar]
- 97.Nijkamp FP, Van der Linde HJ, Folkerts G. Nitric oxide synthesis inhibitors induce airway hyperresponsiveness in the guinea pig in vivo and in vitro: role of the epithelium. American Review of Respiratory Disease. 1993;148(3):727–734. doi: 10.1164/ajrccm/148.3.727. [DOI] [PubMed] [Google Scholar]
- 98.Munakata M, Masaki Y, Sakuma I, et al. Pharmacological differentiation of epithelium-derived relaxing factor from nitric oxide. Journal of Applied Physiology. 1990;69(2):665–670. doi: 10.1152/jappl.1990.69.2.665. [DOI] [PubMed] [Google Scholar]
- 99.De Boer J, Duyvendak M, Schuurman FE, Pouw FMH, Zaagsma J, Meurs H. Role of L-arginine in the deficiency of nitric oxide and airway hyperreactivity after the allergen-induced early asthmatic reaction in guinea-pigs. British Journal of Pharmacology. 1999;128(5):1114–1120. doi: 10.1038/sj.bjp.0702882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Kraft M. Part III: location of asthma inflammation and the distal airways: clinical implications. Current Medical Research and Opinion. 2007;23(supplement 3):S21–S27. doi: 10.1185/030079907X226177. [DOI] [PubMed] [Google Scholar]
- 101.Lanças T, Kasahara DI, Prado CM, Tibério IFLC, Martins MA, Dolhnikoff M. Comparison of early and late responses to antigen of sensitized guinea pig parenchymal lung strips. Journal of Applied Physiology. 2006;100(5):1610–1616. doi: 10.1152/japplphysiol.00828.2005. [DOI] [PubMed] [Google Scholar]
- 102.In ’t Veen JCCM, Beekman AJ, Bel EH, Sterk PJ. Recurrent exacerbations in severe asthma are associated with enhanced airway closure during stable episodes. American Journal of Respiratory and Critical Care Medicine. 2000;161(6):1902–1906. doi: 10.1164/ajrccm.161.6.9906075. [DOI] [PubMed] [Google Scholar]
- 103.Mauad T, Carolina A, Xavier G, Hilário P, Saldiva N, Dolhnikoff M. Elastosis and fragmentation of fibers of the elastic system in fatal asthma. American Journal of Respiratory and Critical Care Medicine. 1999;160(3):968–975. doi: 10.1164/ajrccm.160.3.9809088. [DOI] [PubMed] [Google Scholar]
- 104.Angeli P, Prado CM, Xisto DG, et al. Effects of chronic L-NAME treatment lung tissue mechanics, eosinophilic and extracellular matrix responses induced by chronic pulmonary inflammation. American Journal of Physiology. 2008;294(6):L1197–L1205. doi: 10.1152/ajplung.00199.2007. [DOI] [PubMed] [Google Scholar]
- 105.Starling CM, Prado CM, Leick-Maldonado EA, et al. Inducible nitric oxide synthase inhibition attenuates lung tissue responsiveness and remodeling in a model of chronic pulmonary inflammation in guinea pigs. Respiratory Physiology and Neurobiology. 2009;165(2-3):185–194. doi: 10.1016/j.resp.2008.11.011. [DOI] [PubMed] [Google Scholar]
- 106.Taylor DA, McGrath JL, O’Connor BJ, Barnes PJ. Allergen-induced early and late asthmatic responses are not affected by inhibition of endogenous nitric oxide. American Journal of Respiratory and Critical Care Medicine. 1998;158(1):99–106. doi: 10.1164/ajrccm.158.1.9709091. [DOI] [PubMed] [Google Scholar]
- 107.Belvisi MG, Stretton CD, Yacoub M, Barnes PJ. Nitric oxide is the endogenous neurotransmitter of bronchodilator nerves in humans. European Journal of Pharmacology. 1992;210(2):221–222. doi: 10.1016/0014-2999(92)90676-u. [DOI] [PubMed] [Google Scholar]
- 108.Prado CM, Leick-Maldonado EA, Miyamoto L, et al. Capsaicin-sensitive nerves and neurokinins modulate non-neuronal nNOS expression in lung. Respiratory Physiology and Neurobiology. 2008;160(1):37–44. doi: 10.1016/j.resp.2007.08.004. [DOI] [PubMed] [Google Scholar]
- 109.Martins MA, Shore SA, Drazen JM. Capsaicin-induced release of tachykinins: effects of enzyme inhibitors. Journal of Applied Physiology. 1991;70(5):1950–1956. doi: 10.1152/jappl.1991.70.5.1950. [DOI] [PubMed] [Google Scholar]
- 110.Ricciardolo FLM, Nadel JA, Yoshihara L, Geppetti P. Evidence for reduction of bradykinin-induced bronchoconstriction in guinea-pigs by release of nitric oxide. British Journal of Pharmacology. 1994;113(4):1147–1152. doi: 10.1111/j.1476-5381.1994.tb17117.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Samb A, Pretolani M, Dinh-Xuan A-T, et al. Decreased pulmonary and tracheal smooth muscle expression and activity of type 1 nitric oxide synthase (nNOS) after ovalbumin immunization and a multiple aerosol challenge in guinea pigs. American Journal of Respiratory and Critical Care Medicine. 2001;164:149–154. doi: 10.1164/ajrccm.164.1.2004030. [DOI] [PubMed] [Google Scholar]
- 112.Ferreira HH, Lodo ML, Martins AR, et al. Expression of nitric oxide synthases and in vitro migration of eosinophils from allergic rhinits subjects. European Journal of Pharmacology. 2002;442(1-2):155–162. doi: 10.1016/s0014-2999(02)01507-8. [DOI] [PubMed] [Google Scholar]
- 113.Gatto EM, Riobó NA, Carreras MC, et al. Overexpression of neutrophil neuronal nitric oxide synthase in Parkinson’s disease. Nitric Oxide. 2000;4(5):534–539. doi: 10.1006/niox.2000.0288. [DOI] [PubMed] [Google Scholar]
- 114.Greenberg SS, Ouyang J, Zhao X, Giles TD. Human and rat neutrophils constitutively express neural nitric oxide synthase mRNA. Nitric Oxide. 1998;2(3):203–212. doi: 10.1006/niox.1998.0176. [DOI] [PubMed] [Google Scholar]
- 115.Maruo K, Kayashima KI, Ono T. Expression of neuronal nitric oxide synthase in dermal infiltrated eosinophils in eosinophilic pustular folliculitis. British Journal of Dermatology. 1999;140(3):417–420. doi: 10.1046/j.1365-2133.1999.02701.x. [DOI] [PubMed] [Google Scholar]
- 116.Bridi R, Crossetti FP, Steffen VM, Henriques AT. The antioxidant activity of standardized extract of Ginkgo biloba (EGb 761) in rats. Phytotherapy Research. 2001;15(5):449–451. doi: 10.1002/ptr.814. [DOI] [PubMed] [Google Scholar]
- 117.Cardell LO, Agustí C, Nadel JA. Nitric Oxide-dependent neutrophil recruitment: role in nasal secretion. Clinical and Experimental Allergy. 2000;30(12):1799–1803. doi: 10.1046/j.1365-2222.2000.00984.x. [DOI] [PubMed] [Google Scholar]
- 118.Trifilieff A, Fujitani Y, Mentz F, Dugas B, Fuentes M, Bertrand C. Inducible nitric oxide synthase inhibitors supress airway inflammation in mice through down-regulation of chemokine expression. The Journal of Immunology. 2000;165:1526–1533. doi: 10.4049/jimmunol.165.3.1526. [DOI] [PubMed] [Google Scholar]
- 119.Miller MJS, Thompson JH, Liu X, et al. Failure of L-NAME to cause inhibition of nitric oxide synthesis: role of inducible nitric oxide synthase. Inflammation Research. 1996;45(6):272–276. doi: 10.1007/BF02280990. [DOI] [PubMed] [Google Scholar]
- 120.Taylor EL, Megson IL, Haslett C, Rossi AG. Nitric oxide: a key regulator of myeloid inflammatory cell apoptosis. Cell Death and Differentiation. 2003;10(4):418–430. doi: 10.1038/sj.cdd.4401152. [DOI] [PubMed] [Google Scholar]
- 121.McCluskie K, Birrell MA, Wong S, Belvisi MG. Nitric oxide as a noninvasive biomarker of lipopolysaccharide-induced airway inflammation: possible role in lung neutrophilia. Journal of Pharmacology and Experimental Therapeutics. 2004;311(2):625–633. doi: 10.1124/jpet.104.068890. [DOI] [PubMed] [Google Scholar]
- 122.Iijima H, Duguet A, Eum SY, Hamid Q, Eidelman DH. Nitric oxide and protein nitration are eosinophil dependent in allergen-challenged mice. American Journal of Respiratory and Critical Care Medicine. 2001;163(5):1233–1240. doi: 10.1164/ajrccm.163.5.2003145. [DOI] [PubMed] [Google Scholar]
- 123.Cirino G, Fiorucci S, Sessa WC. Endothelial nitric oxide synthase: the Cinderella of inflammation? Trends in Pharmacological Sciences. 2003;24(2):91–95. doi: 10.1016/S0165-6147(02)00049-4. [DOI] [PubMed] [Google Scholar]
- 124.Cook S, Vollenweider P, Ménard B, Egli M, Nicod P, Scherrer U. Increased eNO and pulmonary iNOS expression in eNOS null mice. European Respiratory Journal. 2003;21(5):770–773. doi: 10.1183/09031936.03.00121203. [DOI] [PubMed] [Google Scholar]
- 125.Marshall HE, Stamler JS. Inhibition of NF-κB by S-nitrosylation. Biochemistry. 2001;40(6):1688–1693. doi: 10.1021/bi002239y. [DOI] [PubMed] [Google Scholar]
- 126.Ten Broeke R, De Crom R, Van Haperen R, et al. Overexpression of endothelial nitric oxide synthase suppresses features of allergic asthma in mice. Respiratory Research. 2006;7, article 58 doi: 10.1186/1465-9921-7-58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Thomassen MJ, Buhrow LT, Connors MJ, Kaneko FT, Erzurum SC, Kavuru MS. Nitric oxide inhibits inflammatory cytokine production by human alveolar macrophages. American Journal of Respiratory Cell and Molecular Biology. 1997;17(3):279–283. doi: 10.1165/ajrcmb.17.3.2998m. [DOI] [PubMed] [Google Scholar]
- 128.Phipps S, Benyahia F, Ou TT, Barkans J, Robinson DS, Kay AB. Acute allergen-induced airway remodeling in atopic asthma. American Journal of Respiratory Cell and Molecular Biology. 2004;31(6):626–632. doi: 10.1165/rcmb.2004-0193OC. [DOI] [PubMed] [Google Scholar]
- 129.Eynott PR, Groneberg DA, Caramori G, et al. Role of nitric oxide in allergic inflammation and bronchial hyperresponsiveness. European Journal of Pharmacology. 2002;452(1):123–133. doi: 10.1016/s0014-2999(02)02237-9. [DOI] [PubMed] [Google Scholar]
- 130.Gabazza EC, Taguchi O, Tamaki S, et al. Role of nitric oxide in airway remodelling. Clinical Science. 2000;98(3):291–294. [PubMed] [Google Scholar]
- 131.Hamad AM, Knox AJ. Mechanisms mediating the antiproliferative effects of nitric oxide in cultured human airway smooth muscle cells. FEBS Letters. 2001;506(2):91–96. doi: 10.1016/s0014-5793(01)02883-6. [DOI] [PubMed] [Google Scholar]
- 132.Ohbayashi H, Shimokata K. Matrix metalloproteinase-9 and airway remodeling in asthma. Current Drug Targets: Inflammation and Allergy. 2005;4(2):177–181. doi: 10.2174/1568010053586246. [DOI] [PubMed] [Google Scholar]
- 133.Cluzel M, Damon M, Chanez P, et al. Enhanced alveolar cell luminol-dependent chemiluminescence in asthma. Journal of Allergy and Clinical Immunology. 1987;80(2):195–201. doi: 10.1016/0091-6749(87)90129-1. [DOI] [PubMed] [Google Scholar]
- 134.Degenhart HJ, Raatgeep HC, Neijens HJ, Kerrebjin KF. Oxygen radicals and their production by leucocytes from children with asthma and bronchial hyperresponsiveness. Clinical Respiratory Physiology. 1986;22(supplement 7):100–103. [PubMed] [Google Scholar]
- 135.Sugiura H, Ichinose M. Oxidative and nitrative stress in bronchial asthma. Antioxidants and Redox Signaling. 2008;10(4):785–797. doi: 10.1089/ars.2007.1937. [DOI] [PubMed] [Google Scholar]
- 136.Janssen LJ, Premji M, Netherton S, Coruzzi J, Lu-Chao H, Cox PG. Vasoconstrictor actions of isoprostanes via tyrosine kinase and Rho kinase in human and canine pulmonary vascular smooth muscles. British Journal of Pharmacology. 2001;132(1):127–134. doi: 10.1038/sj.bjp.0703784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Montuschi P, Barnes PJ, Roberts LJ., II Isoprostanes: markers and mediators of oxidative stress. FASEB Journal. 2004;18(15):1791–1800. doi: 10.1096/fj.04-2330rev. [DOI] [PubMed] [Google Scholar]
- 138.Wood LG, Gibson PG, Garg ML. Biomarkers of lipid peroxidation, airway inflammation and asthma. European Respiratory Journal. 2003;21(1):177–186. doi: 10.1183/09031936.03.00017003a. [DOI] [PubMed] [Google Scholar]
- 139.Quaggiotto P, Garg ML. Isoprostanes: indicators of oxidative stress in vivo and their biological activity. In: Basu TK, Norman T, Garg MD, editors. Antioxidant in Human Health and Diseases. Oxford, UK: CAB; 1999. pp. 393–410. [Google Scholar]
- 140.Mehrabi MR, Serbecic N, Ekmekcioglu C, et al. The isoprostane 8-epi-PGF(2alpha) is a valuable indicator of oxidative injury in human heart valves. Cardiovascular Pathology. 2001;10(5):241–245. doi: 10.1016/s1054-8807(01)00084-9. [DOI] [PubMed] [Google Scholar]
- 141.Hansel TT, Kharitonov SA, Donnelly LE, et al. A selective inhibitor of inducible nitric oxide synthase inhibits exhaled breath nitric oxide in healthy volunteers and asthmatics. The FASEB Journal. 2003;17(10):1298–1300. doi: 10.1096/fj.02-0633fje. [DOI] [PubMed] [Google Scholar]
- 142.Singh D, Richards D, Knowles RG, et al. Selective inducible nitric oxide synthase inhibition has no effect on allergen challenge in asthma. American Journal of Respiratory and Critical Care Medicine. 2007;176(10):988–993. doi: 10.1164/rccm.200704-588OC. [DOI] [PubMed] [Google Scholar]
- 143.Turner DL, Ferrari N, Ford WR, et al. TPI 1020, a novel anti-inflammatory, nitric oxide donating compound, potentiates the bronchodilator effects of salbutamol in conscious guinea-pigs. European Journal of Pharmacology. 2010;641(2-3):213–219. doi: 10.1016/j.ejphar.2010.05.025. [DOI] [PubMed] [Google Scholar]
- 144.Jonasson S, Hedenstierna G, Hjoberg J. Concomitant administration of nitric oxide and glucocorticoids improves protection against bronchoconstriction in a murine model of asthma. Journal of Applied Physiology. 2010;109(2):521–531. doi: 10.1152/japplphysiol.01317.2009. [DOI] [PubMed] [Google Scholar]
- 145.Maarsingh H, Zaagsma J, Meurs H. Arginase: a key enzyme in the pathophysiology of allergic asthma opening novel therapeutic perspectives. British Journal of Pharmacology. 2009;158(3):652–664. doi: 10.1111/j.1476-5381.2009.00374.x. [DOI] [PMC free article] [PubMed] [Google Scholar]