

Abstract
In this study, we have investigated the effects of total saponin from Korean red ginseng (TSKRG) on thrombin-induced platelet aggregation. TSKRG dose-dependently inhibited thrombin-induced platelet aggregation with IC50 value of about 81.1 μg/mL. In addition, TSKRG dose-dependently decreased thrombin-elevated the level of cytosolic-free Ca2+ ([Ca2+]i), one of aggregation-inducing molecules. Of two Ca2+-antagonistic cyclic nucleotides as aggregation-inhibiting molecules, cyclic adenosine monophosphate (cAMP) and cyclic guanosine monophosphate (cGMP), TSKRG significantly dose-dependently elevated intracellular level of cAMP, but not cGMP. In addition, TSKRG dose-dependently inhibited thrombin-elevated adenosine triphosphate (ATP) release from platelets. These results suggest that the suppression of [Ca2+]i elevation, and of ATP release by TSKRG are associated with upregulation of cAMP. TSKRG elevated the phosphorylation of vasodilator-stimulated phosphoprotein (VASP)-Ser157, a cAMP-dependent protein kinase (A-kinase) substrate, but not the phosphorylation of VASP-Ser239, a cGMPdependent protein kinase substrate, in thrombin-activated platelets. We demonstrate that TSKRG involves in increase of cAMP level and subsequent elevation of VASP-Ser157 phosphorylation through A-kinase activation to inhibit [Ca2+]i mobilization and ATP release in thrombin-induced platelet aggregation. These results strongly indicate that TSKRG is a beneficial herbal substance elevating cAMP level in thrombin-platelet interaction, which may result in preventing of platelet aggregation-mediated thrombotic diseases.
Keywords: Panax ginseng, Cytosolic-free Ca2+, Cyclic adenosine monophosphate, Adenosine triphosphate release, Vasodilator-stimulated phosphoprotein
INTRODUCTION
Platelet aggregation is essential for normal haemostatic process when blood vessels are injured. However, it can also cause cardiovascular disease such as thrombosis, atherosclerosis and myocardial infarction [1]. Hence, inhibition of platelet aggregation might be a promising target to the development of anti-thrombotic drugs and an approach for the prevention of cardiovascular disease. When platelets are activated by agonists such as collagen, thrombin, and adenosine diphosphate, phosphatidylinositol 4, 5-bisphosphate (PIP2) is broken down by phospholipase C which is activated through G-protein coupled receptor or glycoprotein VI. At this time, diacylglycerol and inositol-1,4,5-trisphosphate (IP3) are generated from PIP2 [2,3]. IP3 mobilizes Ca2+ from dense tubular system into cytoplasm. Ca2+/calmodulin complex activates myosin light chain kinase, which in turn phosphorylates the myosin light chain to activate platelets [4,5]. On the other hand, both intracellular cyclic adenosine monophosphate (cAMP) and cyclic guanosine monophosphate (cGMP) as anti-platelet regulators decrease the cytosolic-free Ca2+ ([Ca2+]i), an essential factor for platelet aggregation. Therefore, inhibition of [Ca2+]i or increase of cAMP and cGMP is very useful to evaluate an antiplatelet effect of any substance or compound. It is known that the effects of cAMP and cGMP in platelets are mediated via cAMP- and cGMP-dependent protein kinases (A-kinase and G-kinase), which phosphorylate substrate protein IP3 receptor and vasodilator-stimulated phosphoprotein (VASP) [6-8]. IP3 receptor phosphorylation involves in inhibition of [Ca2+]i mobilization [9,10], and VASP phosphorylation involves in inhibition of VASP affinity for contractile protein filamentous actin [11] to inhibit platelet aggregation. VASP is composed of 46 kDa-dephosphoprotein and 50 kDa-phosphoprotein [7]. If VASP is phosphorylated by A-kinase or G-kinase, phosphorylation of VASP (p-VASP) shifts from 46 kDa-dephosphoprotein to 50 kDa-phosphoprotein [6,7]. Ser157 at 50 kDa of VASP is phosphrylated by cAMP/A-kinase pathway, on the other hand, Ser239 at 50 kDa of VASP is phosphorylated by cGMP/G-kinase pathway [12,13]. Therefore, phosphorylations of Ser157 or Ser239 at 50 kDa of VASP are useful indicators for monitoring cAMP/A-kinase and cGMP/G-kinase pathways.
Ginseng, the root of Panax ginseng Meyer, has been used frequently in traditional oriental medicine, and is known to have various pharmacological activities such as anti-inflammatory action, anti-oxidation, antitumor, anti-diabetes, and anti-hepatotoxicity [14,15]. In recent, it is reported that Korean red ginseng has an effect on cardiovascular disease, which is characterized with regard to reduction of blood pressure and arterial stiffness by inhibition of Rho kinase [16], anti-coagulation by prolong of prothrombin time and activated partial thromboplastin time [17], endothelium relaxation by nitric oxide-cGMP pathway [18], and inhibition of hypercholesterolemia-induced platelet aggregation [19]. In our previous report, we demonstrated that total saponin from Korean red ginseng (TSKRG) is a beneficial traditional oriental medicine in platelet-mediated thrombotic disease via suppression of cyclooxygenase-1 (COX-1) and TXA2 synthase (TXAS) to inhibit production of TXA2 [20]. As described above, since TXA2 is produced when [Ca2+]i level is increased by agonists, TSKRG that decreases TXA2 production must decline [Ca2+]i level to have antiplatelet effect. In this study, we investigated whether TSKRG reduces thrombin-elevated [Ca2+]i level, increases Ca2+-antagonistic cAMP and cGMP level, stimulates the p-VASP, and inhibits the release of ATP, a cAMP precursor, to evaluate antiplatelet effect of TSKRG.
MATERIALS AND METHODS
Materials
TSKRG was obtained from R&D Headquarter, Korea Ginseng Corporation (Daejeon, Korea). Thrombin was purchased from Chrono-Log Corporation (Havertown, PA, USA). cAMP and cGMP enzyme immunoassay (EIA) kits were purchased from GE Healthcare (Buckinghamshire, UK). Fura 2-AM, and other reagents were obtained from Sigma Chemical Co. (St. Louis, MO, USA). ATP assay kit is purchased from Biomedical Research Service Center (Buffalo, NY, USA). Anti-VASP, anti-phosphor-VASP (Ser157), anti-phosphor-VASP (Ser239), and anti-rabbit IgG-horseradish peroxidase conjugate (HRP) or anti-goat IgG-HRP were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Polyvinylidene difluoride (PVDF) membrane was from GE Healthcare (Piseataway, NJ, USA). Enhanced chemiluminesence solution (ECL) was from GE Healthcare.
Preparation of washed rat platelets
Blood was collected from Sprague-Dawley rats (6 to 7 weeks old, male), and anti-coagulated with acid-citrate-dextrose solution (0.8% citric acid, 2.2% sodium citrate, 2.45% glucose). Platelet-rich plasma (PRP) was centrifuged at 125 ×g for 10 min to remove red blood cells, and the platelets were washed twice with washing buffer (138 mM NaCl, 2.7 mM KCl, 12 mM NaHCO3, 0.36 mM NaH2PO4, 5.5 mM glucose, and 1 mM EDTA, pH 6.9). The washed platelets were then resuspended in suspension buffer (138 mM NaCl, 2.7 mM KCl, 12 mM NaHCO3, 0.36 mM NaH2PO4, 0.49 mM MgCl2, 5.5 mM glucose, 0.25% gelatin, pH 7.4) to a final concentration of 5×108/mL. All of the above procedures were carried out at 25℃ to avoid platelet aggregation on cooling. The ethics committee for animal experiments of Inje University (Gimhae, Korea) approved these animal experiments.
Measurement of platelet aggregation
Washed platelets (108/mL) were preincubated for 3 min at 37℃ in the presence of 2 mM exogenous CaCl2 with or without TSKRG, then stimulated with thrombin (0.5 U/mL) for 5 min. Aggregation was monitored using an aggregometer (Chrono-Log Corporation) at a constant stirring speed of 1,000 rpm. Each aggregation rate was calculated as an increase in light transmission. The suspension buffer was used as the reference (transmission 0). TSKRG was dissolved in distilled water.
Determination of cytosolic-free Ca2+
PRP was incubated with 5 μM fura 2-AM at 37℃ for 60 min. Because fura 2-AM is light sensitive, the tube containing the PRP was covered with aluminum foil during loading. The fura 2-loaded washed platelets were prepared using the procedure described above and platelets 108/mL were preincubated for 3 min at 37℃ with or without various concentrations of TSKRG in the presence of 2 mM CaCl2, then stimulated with thrombin (0.5 U/mL) for 5 min for evaluation of [Ca2+]i. Fura 2 fluorescence was measured with a spectrofluorometer (SFM 25; Bio-Teck Instrument, Milan, Italy) with an excitation wavelength that was changed every 0.5 s from 340 to 380 nm; the emission wavelength was set at 510 nm. The [Ca2+]i values were calculated using the method of Schaeffer [21].
Measurement of cyclic adenosine monophosphate and cyclic guanosine monophosphate
Washed platelets (108/mL) were preincubated for 3 min at 37℃ with or without various concentrations of TSKRG in the presence of 2 mM CaCl2, then stimulated with thrombin (0.5 U/mL) for 5 min for platelet aggregation. The aggregation was terminated by the addition of 80% ice-cold ethanol. cAMP and cGMP were measured with Synergy HT Multi-Model Microplate Reader (BioTek Instruments, Winooski, VT, USA) using cAMP and cGMP EIA kits.
Determination of adenosine triphosphate release
Washed platelets (108/mL) were preincubated for 3 min at 37℃ with or without various concentrations of TSKRG in the presence of 2 mM CaCl2, then stimulated with thrombin (0.5 U/mL). The reaction was terminated by the addition of ice-cold 2mM EDTA, the samples were centrifuged and supernatants were used for the assay of ATP release from dense body. ATP release was measured in a luminometer (BioTek Instruments) using an ATP assay kit.
Western blot for analysis of vasodilator-stimulated phosphoprotein phosphorylation
Platelet lysates containing the same protein (15 μg) were used for analysis. Protein concentrations were measured by using bicinchoninic acid protein assay kit (Pierce Biotechnology, Rockford, IL, USA). An 8% to 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) was used for electrophoresis and a PVDF membrane was used for protein transfer from the gel. The dilutions for anti-VASP, anti-phosphor-VASP (Ser157), anti-phosphor-VASP (Ser239), and anti-rabbit IgG-HRP were 1:1,000, 1:1,000, 1:1,000, and 1:10,000, respectively. The membranes were visualized using ECL. Blots were analyzed by using the Quantity One ver. 4.5 (Bio-Rad, Hercules, CA, USA).
Analysis of ginsenosides in total saponin from Korean red ginseng with HPLC-evaporative light scattering detection
TSKRG was dissolved with 100% methanol and then analyzed by HPLC [22,23]. An Dionex-Ultimate 3000 series HPLC apparatus (Dionex, Sunnyvale, CA, USA), equipped with vacuum degaser, quaternary gradient pump, and autosampler was used. A Zorbax ODS C18 column (250 mm×4.6 mm id, 5 μm) and a Zorbax ODS C18 guard column (12.5 mm×4.6 mm id, 5 μm) were used at a column temperature of 35℃. The mobile phase consisted of water (a) and acetonitrile (b) using the following gradient program: 0 to 30 min, 18% to 19% b; 30 to 40 min, 19% to 31% b; 40 to 60 min, 31% to 56% b. The flow rate was at 1.5 mL/min and sample injection volume was 10 μL. Evaporative light scattering detection (ELSD) was set to a probe temperature of 70℃ and nebulizer nitrogen gas flow rate was at 1.4 L/min [24]. Due to the distinct variation on contents of ginsenosides in TSKRG, the two methanol stock solutions of standards, one containing Rg1, Rb1, Rg2, and Rd; the other containing Re, Rc, Rb2, and Rg3, were prepared and diluted with methanol in appropriate concentration for calibration curves. Three concentrations of the 8 ginsenosides solutions were injected in duplicate, then the calibration curves were constructed by plotting the peak area against the concentration of each analyte.
Statistical analysis
The experimental results are expressed as means±SEM accompanied by the number of observations. Data were assessed by ANOVA. If this analysis indicated significant differences among the group means, then each group was compared by the Newman-Keuls method. A p-value less than 0.05 was considered statistically significant.
RESULTS
Effects of total saponin from Korean red ginseng on thrombin-induced platelet aggregation
In our previous report [20], thrombin concentration that maximally generates platelet aggregation is observed to 0.25 U/mL. However, we used the 0.5 U/mL of thrombin to aggregate platelets. When washed platelets (108/mL) were stimulated by thrombin in the presence of 2 mM CaCl2, the aggregation rate was increased up to 83.5±2.1%. However, various concentrations of TSKRG (25 to 150 μg/mL) significantly reduced thrombin-stimulated platelet aggregation in a dose-dependent manner (Fig. 1A). IC50 value of TSKRG was approximately 81.1 μg/mL (Fig. 1B).
Fig. 1. Effects of total saponin from Korean red ginseng (TSKRG) on thrombin-stimulated platelet aggregation. (A) Effects of TSKRG pretreatment on thrombin-stimulated platelet aggregation. (B) IC50 value of TSKRG on thrombin-stimulated platelet aggregation. Washed platelets (108/mL) were preincubated with or without various concentrations of TSKRG (25 to 150 μg/mL) in the presence of 2 mM CaCl2 for 3 min at 37℃, and then stimulated with thrombin (0.5 U/mL) for 5 min. Platelet aggregation (%) was recorded as an increase in light transmission. Inhibition rate by TSKRG was recorded as percentage of the thrombin-induced aggregation rate. IC50 value of TSKRG was calculated by 4-parameter log fit method [25]. Data are expressed as means±SEM (n=4). **p<0.001.
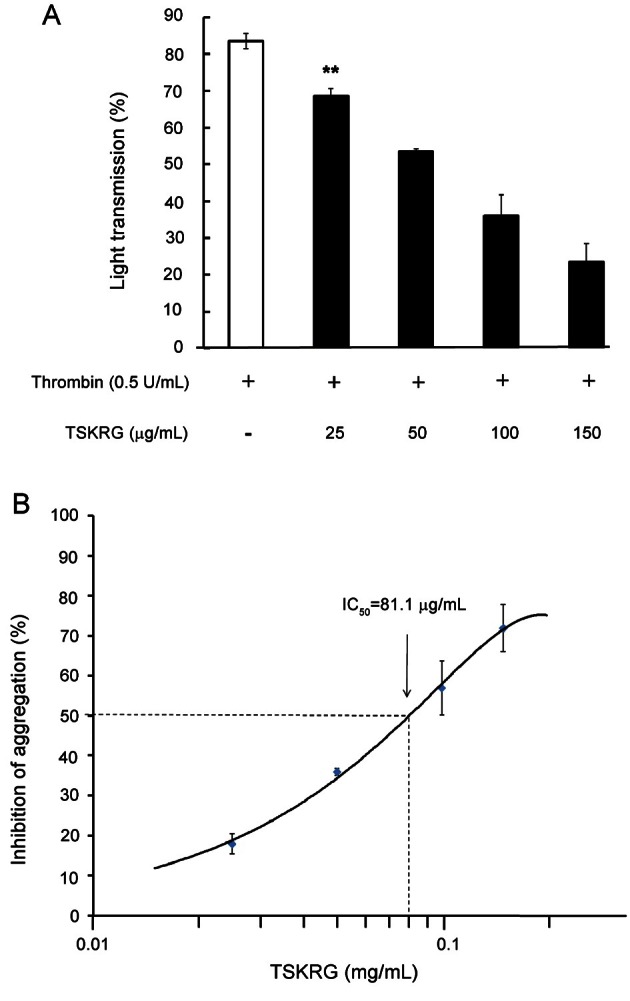
Effects of total saponin from Korean red ginseng on the down-regulation of aggregation-inducing molecule, cytosolic-free Ca2+
When washed platelets were stimulated by thrombin, the level of [Ca2+]i increased from 121.3 nM, the basal level, to 477.5±48.7 nM (Fig. 2). However, this was significantly reduced by various concentrations (25 to 150 μg/mL) of TSKRG in a dose-dependent manner. Thrombin-elevated [Ca2+]i (477.5±48.7 nM) was decreased to 218.0±11.3 nM by 150 μg/mL of TSKRG, and its inhibitory degree was 54.3% as compared with that by thrombin (Fig. 2, small table). We next investigated whether TSKRG up-regulates the level of intracellular cAMP and cGMP, Ca2+-antagonistic molecules, in thrombin-induced platelet aggregation.
Fig. 2. Effect of total saponin from Korean red ginseng (TSKRG) on thrombin-induced cytosolic-free Ca2+ ([Ca2+]i) mobilization. Fura 2-loaded platelets (108/mL) were preincubated with or without various concentrations of TSKRG in the presence of 2 mM CaCl2 for 3 min at 37℃, and then thrombin (0.5 U/mL) was added. [Ca2+]i was determined as described in Materials and Methods section. Data are expressed as means±SEM (n=3). *p<0.05.
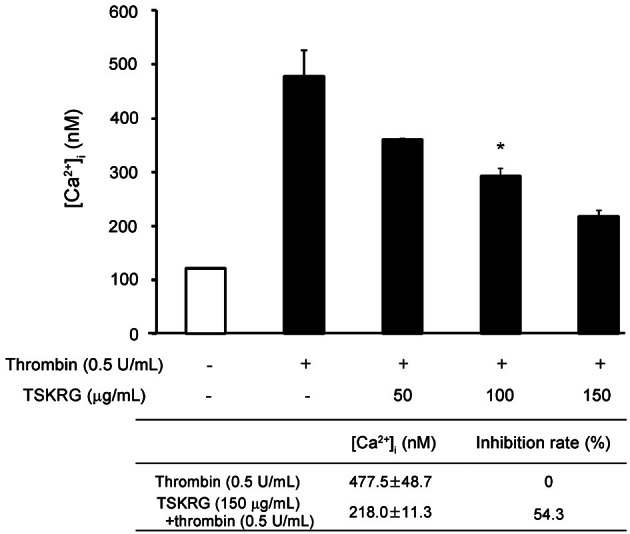
Effects of total saponin from Korean red ginseng on the up-regulation of aggregation-inhibiting molecules, cyclic adenosine monophosphate and cyclic guanosine monophosphate
As shown in Fig. 3A, thrombin decreased intracellular cAMP level from 3.82±0.12 pmol/109 platelets (basal level) to 3.02±0.27 pmol/109 platelets, which was reduced to 20.9% as compared with that of basal level (Fig. 3A, small table). When the platelets, however, were incubated in the presence of both TSKRG and thrombin, TSKRG increased cAMP level in a dose dependent, and 150 μg/mL of TSKRG increased from 3.02±0.27 pmol/109 platelets to 9.42±0.68 pmol/109 platelets (Fig. 3A). This result suggests that TSKRG increased thrombin-decreased cAMP level to 211.9% (Fig. 3A, small table).
Fig. 3. Effects of total saponin from Korean red ginseng (TSKRG) on cyclic adenosine monophosphate (cAMP) and cyclic guanosine monophosphate (cGMP) production in resting and thrombin-stimulated platelets. (A) Effects of TSKRG on cAMP production in resting or thrombin-stimulated platelets. (B) Effects of TSKRG on cGMP production in resting or thrombin-stimulated platelets. Washed platelets (108/mL) were preincubated with or without various concentrations of TSKRG for 3 min in the presence of 2 mM CaCl2 and then stimulated with thrombin (0.5 U/mL) for 5 min at 37℃. The reactions were terminated by adding 80% ice-cold ethanol. cAMP and cGMP contents were measured using enzyme immunoassay kits. Data are expressed as means±SEM (n=4). NS, not significant. *p<0.05.
On the other hand, thrombin decreased intracellular cGMP level from 2.24±0.14 pmol/109 platelets (basal level) to 1.16±0.23 pmol/109 platelets. This means that thrombin reduced basal cGMP level to 25.9% to aggregate platelets (Fig. 3B, small table). When the platelets, however, were incubated in the presence of both TSKRG and thrombin, the cGMP level was not significantly increased (Fig. 3B).
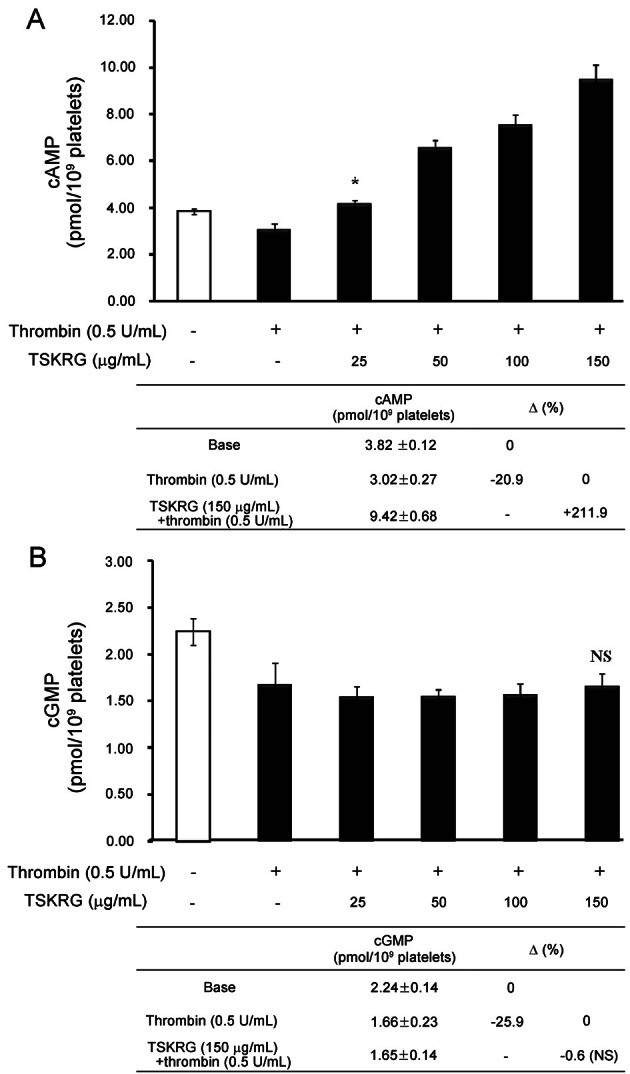
Effects of total saponin from Korean red ginseng on adenosine triphosphate release
Because ATP release out of dense body in platelets is critical marker of platelet aggregation [26], which reflects inhibition of intracellular cAMP production, we investigated the effects of TSKRG on thrombin-elevated ATP release. As shown in Fig. 4A, ATP level in supernatant from the thrombin-activated platelets was 3.48±0.03 μM, which is 69.6 fold as compared with that (0.05±0.01 μM) in intact cell, control. This reflects that thrombin-elevated ATP release is resulted from thrombin-aggregated platelets (Fig. 4B-a). However, TSKRG (25 to 150 μg/mL) dose dependently inhibited thrombin-elevated ATP release (3.48±0.03 μM) (Fig. 4A), and TSKRG (150 μg/mL) inhibited ATP release to 80.2% (0.69±0.02 μM) as compared with that (3.48±0.03 μM ) by thrombin (Fig. 4A, small table). In addition, the dose dependent-inhibition of ATP release by TSKRG (25 to 150 μg/mL) was accompanied with dose dependent-inhibition of platelet aggregation (Fig. 4B-b, c, d, and e).
Fig. 4. Effects of total saponin from Korean red ginseng (TSKRG) on adenosine triphosphate (ATP) release in resting and thrombin-stimulated platelets and light transmission (%) coincidentally. (A) Effects of TSKRG on ATP release in resting and thrombin-stimulated platelets. (B) Light transmission (%) with ATP release on thrombin-stimulated platelets. a, thrombin; b, thrombin+TSKRG (25 μg/mL); c, thrombin+TSKRG (50 μg/mL); d, thrombin+TSKRG (100 μg/mL); and e, thrombin+TSKRG (150 μg/mL). Washed platelets (108/mL) were preincubated with or without various concentrations of TSKRG for 3 min in the presence of 2 mM CaCl2 and then stimulated with thrombin (0.5 U/mL) for 5 min at 37℃. The reaction was terminated by the addition of ice-cold 2mM EDTA, and centrifuged and supernatant were used for the assay of ATP release. ATP release was measured in a luminometer using an ATP assay kit. Data are expressed as means±SEM (n=4). **p<0.001.
Effects of total saponin from Korean red ginseng on vasodilator-stimulated phosphoprotein phosphorylation
In intact platelets, basal 46 kDa dephosphoprotein of VASP only was observed (Fig. 5A, lane 1) and was shifted to 50 kDa phosphoprotein of VASP in thrombin-induced platelet aggregation (Fig. 5A, lane 2). Thrombin increased weakly the p-VASP (Ser157) at 50 kDa phosphoprotein of VASP (Fig. 5A, lane 2). It is known that thrombin and collagen, agonists of platelets, involve in a feedback inhibition by elevating p-VASP (Ser157 and Ser239) [27]. The ratio of VASP (46+50 kDa) to β-actin was dose dependently increased in the presence of both thrombin and TSKRG (Fig. 5B). The p-VASP (Ser157) at 50 kDa phosphoprotein of VASP (Fig. 5A; lanes 3, 4), and the ratio of p-VASP (Ser157)-50 kDa to β-actin (Fig. 5C) were dose dependently increased in the presence of both thrombin and TSKRG. Even though the ratio of VASP (46+50 kDa) to β-actin was dose dependently increased in the presence of both thrombin and TSKRG (Fig. 5B), the ratio of p-VASP (Ser239)-50 kDa to β-actin (Fig. 5D) was not significantly increased in the presence of both thrombin and TSKRG.
Fig. 5. Effects of total saponin from Korean red ginseng (TSKRG) on vasodilator-stimulated phosphoprotein (VASP) phosphorylation of resting or thrombin-stimulated platelets. (A) Effects of TSKRG on phosphorylations of VASP, VASP (Ser157), VASP (Ser239). Lane 1, intact platelets (base); lane 2, thrombin; lane 3, thrombin+TSKRG (100 μg/mL); lane 4, thrombin+TSKRG (150 μg/mL). (B) The ratio of VASP (46+50 kDa) to β-actin by TSKRG. (C) The ratio of phosphorylation of VASP (p-VASP) (Ser157)-50 kDa to β-actin by TSKRG. (D) The ratio of p-VASP (Ser239)-50 kDa to β-actin by TSKRG. Washed platelets (108/mL) were preincubated with or without TSKRG for 3 min in the presence of 2 mM CaCl2 and then stimulated with thrombin (0.5 U/mL) for 5 min at 37℃ in an aggregometer. The reactions were terminated by adding an equal volume of lysis buffer. Proteins were extracted, separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) transferred to polyvinylidene difluoride and immunoblotted with the indicated corresponding antidodies, anti-phosphor-VASP Ser239 or Ser157. Blots were visualized by ECL plus kit (Amersham, Buckinghamshire, UK) and the immunoblot panels are representative of 3-4 similar experiments. Blots were analyzed by using the Quantity One ver. 4.5 program, and units were expressed density/mm2.

Ginsenoside composition of total saponin from Korean red ginseng
As shown in Fig. 6, in HPLC-ELSD, eight internal standards, ginsenosides Rg1, Re, Rb1, Rc, Rg2, Rb2, Rd, and Rg3 in order. (Fig. 6A), and TSKRG (Fig. 6B) were separated within 60 min. Various peaks from TSKRG were observed in HPLC-ELSD chromatograms (Fig. 6B). The retention time of eight peaks was in accord with those of internal standard ginsenosides, which were 20(S)-protopanaxadiol group of ginsenoside (Rb1, Rb2, Rc, Rd, and Rg3), and 20(S)-protopanaxatriol (PPT) group of ginsenoside (Re, Rg1, and Rg2) (Fig. 6B).
Fig. 6. HPLC-evaporative light scattering detection (ELSD) chromatograms of standard ginsenosides and total saponin from Korean red ginseng (TSKRG). (A) The chromatograms of standard ginsenosides. (B) The chromatograms of ginsenosides in TSKRG. HPLC was performed on a Zorbax ODS C18 column (250 mm×4.6 mm id, 5 μm) and a Zorbax ODS C18 guard column (12.5 mm×4.6 mm id, 5 μm) were used at a column of 35℃. The mobile phase consisted of water (a) and ACN (b) using the following gradient program: 0 to 30 min, 18% to 19% b; 30 to 40 min, 19% to 31% b; 40 to 60 min, 31% to 56% b. The flow rate was at 1.5 mL/min and sample injection volume was 10 μL. ELSD was set to a probe temperature of 70℃ and nebulizer nitrogen gas flow rate was at 1.4 L/min. 1 to 8, ginsenoside Rg1, Re, Rb1, Rc, Rg2, Rb2, Rd, and Rg3 in order.

Calibration curves were linear over the range of 125 to 1,000 μg/mL for Rg1, Re, Rb1, Rc, Rg2, Rb2, Rd, and 25 to 200 μg/mL for Rg3 with r2>0.99 (Table 1). In contents of ginsenosides calculated from calibration curve, as shown in Table 1, the total contents of Rb1, Rb2, Rc, Rd, and Rg3 were 186.80±4.65 mg/g, and contents of Re, Rg1, and Rg2 were 109.37±2.29 mg/g.
Table 1.
Calibration curves and contents of eight ginsenosides in TSKRG
| Ginsenosides | RT (min) | Calibration curve1) | r2 | Test range (μg/mL) | x 2) (μg/mL) | Contents (mg/g-TSKRG) | |
|---|---|---|---|---|---|---|---|
|
| |||||||
| PPD | Rb1 | 47.1 | y=0.0542x-0.008 | 0.9929 | 125-1,000 | 700.23±33.39 | 70.02±3.34 |
| Rb2 | 48.6 | y=0.0517x-0.0072 | 0.9935 | 125-1,000 | 339.70±4.97 | 33.97±0.50 | |
| Rc | 47.9 | y=0.134x-0.017 | 0.9957 | 125-1,000 | 390.13±3.50 | 39.01±0.35 | |
| Rd | 50.3 | y=0.0778x-0.010 | 0.9965 | 125-1,000 | 229.40±1.54 | 22.94±0.15 | |
| Rg3 | 58.9 | y=0.0555x-0.0017 | 0.9904 | 25-200 | 207.63±3.10 | 20.76±0.31 | |
| Sum | 186.80±4.65 | ||||||
| PPT | Re | 38.5 | y=0.103x-0.0135 | 0.9957 | 125-1,000 | 148.73±0.46 | 14.87±0.05 |
| Rg1 | 38.2 | y=0.139x-0.0171 | 0.9973 | 125-1,000 | 129.90±0.17 | 12.99±0.02 | |
| Rg2 | 48.3 | y=0.0165x-0.0023 | 0.9941 | 125-1,000 | 815.13±22.15 | 81.51±2.22 | |
| Sum | 109.37±2.29 | ||||||
TSKRG, total saponin from Korean red ginseng; RT, retention time; PPD, 20(S)-protopanaxadiol; PPT, 20(S)-protopanaxatriol.
1,2) y, peak area of analyte; x, concentration of ginsenoside in 10 mg/mL TSKRG (μg/mL).
DISCUSSION
IC50 value of about 81.1 μg/mL of TSKRG that inhibited thrombin-induced platelet aggregation is about 21 fold lower than that (1.7 mg/mL) of aqueous extract from of P. notoginseng [28]. Of several aggregation-inducing molecules, Ca2+ and TXA2 are known to be essential for platelet aggregation [29]. Since TSKRG significantly blocked [Ca2+]i elevation (Fig. 2) and TXA2 production [20], but increased cAMP production (Fig. 3A), it is suggested that TSKRG-elevated Ca2+-antagonistic cAMP (Fig. 3B) may involve in the inhibition of TXA2 production by suppressing [Ca2+]i-mobilization. In special, as shown in Fig. 7A, TSKRG that dose dependently elevated cAMP level attenuated [Ca2+]i level, which means that TSKRG regulates cAMP and Ca2+ as mutual antagonistic molecules to inhibit platelet aggregation. TSKRG that elevated cAMP level attenuated ATP release in a dose dependently manner (Fig. 7B), which means that TSKRG used ATP to produce cAMP (Fig. 3A), and subsequently involves in reduction of [Ca2+]i-mobilization (Fig. 2) to inhibit platelet aggregation (Fig. 1A). cAMP and cGMP involve in inhibition of platelet aggregation by phosphorylating IP3 receptor or VASP via A-kinase or G-kinase pathway [9,10]. In present study, it is unknown whether the inhibitory effect of [Ca2+]i by TSKRG dues to phosphorylation of IP3 receptor by cAMP or cGMP. Because, however, panaxatriol, which is known to form by acid hydrolysis of ginsenosides (ex. ginsenoside Rg1), inhibited thrombin-elevated PIP2 breakdown in platelets [30], and TSKRG contains PPT (Rg1, Rg2, and Re) having antiplatelet effect [31-33] (Table 1), it is thought that the inhibition of [Ca2+]i–mobilization by TSKRG (Fig. 2) may be due to the inhibition of PIP2 breakdown.
Fig. 7. Relationship between the cyclic adenosine monophosphate (cAMP) production and aggregation-related molecules ([Ca2+]i and adenosine triphosphate [ATP] release) in thrombin-induced platelet aggregation. (A) Relationship between the cAMP production and [Ca2+]i in thrombin-induced platelet aggregation from Fig. 2 and Fig. 3A. (B) Relationship between the cAMP production and ATP release in thrombin-induced platelet aggregation from Fig. 3A and Fig. 4A. TSKRG, total saponin from Korean red ginseng.
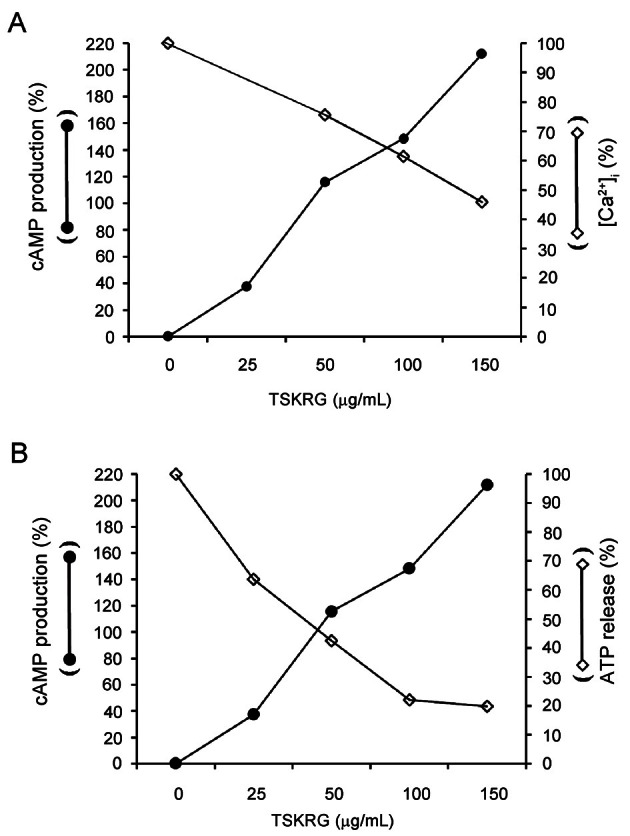
In VASP phosphorylation, TSKRG phosphorylated potently A-kinase substrate VASP (Ser157) (Fig. 5A, C), but did not phosphorylated G-kinase substrate VASP (Ser239) (Fig. 5A, D). These results are in accord with the result that TSKRG potently enhanced cAMP only in thrombin-stimulated platelets (Fig. 3A). Eigenthaler et al. [34] reported that small elevation in cAMP level is enough to activate most of A-kinase, whereas even several fold elevation in cGMP level may stimulate only a small fraction of total G-kinase. This previous report [34] reflects that even a little cAMP is enough to phosphorylate VASP (Ser157), a A-kinase substrate, and a lot of cGMP is required to phosphorylate VASP (Ser239), a G-kinase substrate. In our other report [35], using epigallocatechin-3-gallate, we reported a similar result to our present finding, and suggested that its antiplatelet effect is mediated through an increase of cAMP level, A-kinase activity, and VASP-(Ser157) phosphorylation. The levels of intracellular cAMP and cGMP are regulated by the balance between cyclic nucleotide-producing enzymes, adenylate/guanylate cyclases, and hydrolyzing enzymes, cAMP/cGMP phosphodiesterses (PDEs). If TSKRG inhibited the activity of PDE2 to produce cAMP and cGMP in thrombin-stimulated platelets, because PDE2 hydrolyzes both cAMP and cGMP [36], TSKRG would increase the level of both cAMP and cGMP in thrombin-stimulated platelets, however, TSKRG potently increased the cAMP level only, but did not increased quietly the cGMP level (Fig. 3B). Accordingly, it is thought that TSKRG-elevated cAMP (Fig. 3A) is not due to the regulation of cGMP-stimulated PDE2, cGMP-inhibited PDE3, and cGMP-binding-cGMP-specific PDE5. If so, these results suggest that TSKRG produced cAMP from ATP by activating adenylate cyclase, and phosphorylated VASP (Ser157)-50 kDa by activating A-kinase to inhibit thrombin-induced platelet aggregation.
TXA2 is produced via COX-1 and TXAS pathway from arachidonic acid (20:4). In our previous report, TSKRG inhibited the microsomal COX-1 and TXAS activities in thrombin-induced platelet aggregation. It is thought that TSKRG has multiple actions that reduce [Ca2+]i mobilization (Fig. 2) by elevating cAMP, and inhibit TXA2 production by suppressing activities of both COX-1 and TXAS [20]. 20:4 is also metabolized to prostaglandin E2, an inflammatory mediator, by cyclooxygenase-2 (COX-2). Because 20:4 is a substrate of both COX-1 and COX-2, it is nature that TSKRG and ginsenoside Rp1 have an anti-inflammatory effect by inhibiting COX-2 activity [37-39]. Because both platelet aggregation and inflammation are the cause of atherosclerosis, it is thought that TSKRG could contribute to treatment of cardiovascular disease. It is reported that collagen-induced platelet aggregation, and blood coagulation were inhibited in the subjects who have taken red ginseng products (e.g., water extract, tea, and drink) containing TSKRG as compared with those of control subjects, who did not take ginseng products [40]. Using Korean red ginseng extract, Jin et al. [41] reported similar results to effects of dietary red ginseng products, which inhibited U46619-, 20:4-, collagen-, and thrombin-induced platelet aggregation, prolonged prothrombin time and activated partial thromboplastin time, coagulation indicators. In addition, TSKRG contains Rg1, Rg2, and Rg3 (Table 1) known to inhibit platelet aggregation [31-33]. However, saponins in American ginseng do not contain Rg2 and Rg3 [42]. Accordingly, with regard to antiplatelet effects, it is thought that Korean red ginseng would be outstanding as compared with American.
In conclusion, the most important result of this study is that TSKRG significantly decreases the level of [Ca2+]i, platelet-aggregating molecule, and inhibition of [Ca2+]i by TSKRG depends on up-regulation of Ca2+-antagonistic intracellular cAMP level. Furthermore, our findings showed that TSKRG-elevated cAMP stimulates the phosphorylation of an A-kinase substrate VASP (Ser157)-50 kDa, which may contribute to attenuating [Ca2+]i level to inhibit thrombin-induced platelet aggregation. Therefore, these results suggest that TSKRG may be a physiologically effective negative feedback regulator during platelet aggregation, a cause of thrombosis, atherosclerosis, and myocardial infarction.
Acknowledgments
This study was supported by a grant (2012-0002802 to Hwa-Jin Park) from the Basic Science Research Program via the National Research Foundation of Korea funded by the Ministry of Education, Science and Technology.
References
- 1.Schwartz SM, Heimark RL, Majesky MW. Developmental mechanisms underlying pathology of arteries. Physiol Rev. 1990;70:1177–1209. doi: 10.1152/physrev.1990.70.4.1177. [DOI] [PubMed] [Google Scholar]
- 2.Berridge MJ, Irvine RF. Inositol trisphosphate, a novel second messenger in cellular signal transduction. Nature. 1984;312:315–321. doi: 10.1038/312315a0. [DOI] [PubMed] [Google Scholar]
- 3.Jennings LK. Role of platelets in atherothrombosis. Am J Cardiol. 2009;103:4A–10A. doi: 10.1016/j.amjcard.2008.11.017. [DOI] [PubMed] [Google Scholar]
- 4.Pasqui AL, Capecchi PL, Ceccatelli L, Mazza S, Gistri A, Laghi Pasini F, Di Perri T. Nitroprusside in vitro inhibits platelet aggregation and intracellular calcium translocation. Effect of haemoglobin. Thromb Res. 1991;61:113–122. doi: 10.1016/0049-3848(91)90238-r. [DOI] [PubMed] [Google Scholar]
- 5.Nishikawa M, Tanaka T, Hidaka H. Ca2+-calmodulin-dependent phosphorylation and platelet secretion. Nature. 1980;287:863–865. doi: 10.1038/287863a0. [DOI] [PubMed] [Google Scholar]
- 6.Halbrugge M, Walter U. Purification of a vasodilator-regulated phosphoprotein from human platelets. Eur J Biochem. 1989;185:41–50. doi: 10.1111/j.1432-1033.1989.tb15079.x. [DOI] [PubMed] [Google Scholar]
- 7.Halbrugge M, Friedrich C, Eigenthaler M, Schanzenbacher P, Walter U. Stoichiometric and reversible phosphorylation of a 46-kDa protein in human platelets in response to cGMP- and cAMP-elevating vasodilators. J Biol Chem. 1990;265:3088–3093. [PubMed] [Google Scholar]
- 8.Butt E, Abel K, Krieger M, Palm D, Hoppe V, Hoppe J, Walter U. cAMP- and cGMP-dependent protein kinase phosphorylation sites of the focal adhesion vasodilator-stimulated phosphoprotein (VASP) in vitro and in intact human platelets. J Biol Chem. 1994;269:14509–14517. [PubMed] [Google Scholar]
- 9.Homer KL, Wanstall JC. Inhibition of rat platelet aggregation by the diazeniumdiolate nitric oxide donor MAHMA NONOate. Br J Pharmacol. 2002;137:1071–1081. doi: 10.1038/sj.bjp.0704971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Park WH, Kim HK, Nam KS, Shon YH, Jeon BH, Moon SK, Kim MG, Kim CH. Inhibitory effect of GBH on platelet aggregation through inhibition of intracellular Ca2+ mobilization in activated human platelets. Life Sci. 2004;75:3063–3076. doi: 10.1016/j.lfs.2004.07.010. [DOI] [PubMed] [Google Scholar]
- 11.Laurent V, Loisel TP, Harbeck B, Wehman A, Grobe L, Jockusch BM, Wehland J, Gertler FB, Carlier MF. Role of proteins of the Ena/VASP family in actin-based motility of Listeria monocytogenes. J Cell Biol. 1999;144:1245–1258. doi: 10.1083/jcb.144.6.1245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Smolenski A, Bachmann C, Reinhard K, Honig-Liedl P, Jarchau T, Hoschuetzky H, Walter U. Analysis and regulation of vasodilator-stimulated phosphoprotein serine 239 phosphorylation in vitro and in intact cells using a phosphospecific monoclonal antibody. J Biol Chem. 1998;273:20029–20035. doi: 10.1074/jbc.273.32.20029. [DOI] [PubMed] [Google Scholar]
- 13.Horstrup K, Jablonka B, Honig-Liedl P, Just M, Kochsiek K, Walter U. Phosphorylation of focal adhesion vasodilator-stimulated phosphoprotein at Ser157 in intact human platelets correlates with fibrinogen receptor inhibition. Eur J Biochem. 1994;225:21–27. doi: 10.1111/j.1432-1033.1994.00021.x. [DOI] [PubMed] [Google Scholar]
- 14.Ernst E. Panax ginseng: an overview of the clinical evidence. J Ginseng Res. 2010;34:259–263. [Google Scholar]
- 15.Kim SK, Park JH. Trends in ginseng research in 2010. J Ginseng Res. 2011;35:389–398. doi: 10.5142/jgr.2011.35.4.389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chung IM, Lim JW, Pyun WB, Kim H. Korean red ginseng improves vascular stiffness in patients with coronary artery disease. J Ginseng Res. 2010;34:212–218. [Google Scholar]
- 17.Wee JJ, Kim YS, Kyung JS, Song YB, Do JH, Kim DC, Lee SD. Identification of anticoagulant components in Korean red ginseng. J Ginseng Res. 2010;34:355–362. [Google Scholar]
- 18.Jung YH, Park KY, Jeon JH, Kwak YS, Song YB, Wee JJ, Rhee MH, Kim TW. Red ginseng saponin fraction A isolated from Korean red ginseng by ultrafiltration on the porcine coronary artery. J Ginseng Res. 2011;35:325–330. doi: 10.5142/jgr.2011.35.3.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hwang SY, Son DJ, Kim IW, Kim DM, Sohn SH, Lee JJ, Kim SK. Korean red ginseng attenuates hypercholesterolemia-enhanced platelet aggregation through suppression of diacylglycerol liberation in high-cholesterol-diet-fed rabbits. Phytother Res. 2008;22:778–783. doi: 10.1002/ptr.2363. [DOI] [PubMed] [Google Scholar]
- 20.Lee DH, Cho HJ, Kang HY, Rhee MH, Park HJ. Total saponin from Korean red ginseng inhibits thromboxane A2 production associated microsomal enzyme activity in platelets. J Ginseng Res. 2012;36:40–46. doi: 10.5142/jgr.2012.36.1.40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Schaeffer J, Blaustein MP. Platelet free calcium concentrations measured with fura-2 are influenced by the transmembrane sodium gradient. Cell Calcium. 1989;10:101–113. doi: 10.1016/0143-4160(89)90050-x. [DOI] [PubMed] [Google Scholar]
- 22.Kim SN, Ha YW, Shin H, Son SH, Wu SJ, Kim YS. Simultaneous quantification of 14 ginsenosides in Panax ginseng C.A. Meyer (Korean red ginseng) by HPLC-ELSD and its application to quality control. J Pharm Biomed Anal. 2007;45:164–170. doi: 10.1016/j.jpba.2007.05.001. [DOI] [PubMed] [Google Scholar]
- 23.Wan JB, Lai CM, Li SP, Lee MY, Kong LY, Wang YT. Simultaneous determination of nine saponins from Panax notoginseng using HPLC and pressurized liquid extraction. J Pharm Biomed Anal. 2006;41:274–279. doi: 10.1016/j.jpba.2005.10.023. [DOI] [PubMed] [Google Scholar]
- 24.Wan JB, Li P, Li S, Wang Y, Dong TT, Tsim KW. Simultaneous determination of 11 saponins in Panax notoginseng using HPLC-ELSD and pressurized liquid extraction. J Sep Sci. 2006;29:2190–2196. doi: 10.1002/jssc.200600103. [DOI] [PubMed] [Google Scholar]
- 25.Cao H, Yu R, Choi Y, Ma ZZ, Zhang H, Xiang W, Lee DY, Berman BM, Moudgil KD, Fong HH, et al. Discovery of cyclooxygenase inhibitors from medicinal plants used to treat inflammation. Pharmacol Res. 2010;61:519–524. doi: 10.1016/j.phrs.2010.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mackman N. Triggers, targets and treatments for thrombosis. Nature. 2008;451:914–918. doi: 10.1038/nature06797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gambaryan S, Kobsar A, Rukoyatkina N, Herterich S, Geiger J, Smolenski A, Lohmann SM, Walter U. Thrombin and collagen induce a feedback inhibitory signaling pathway in platelets involving dissociation of the catalytic subunit of protein kinase A from an NFkappaB-IkappaB complex. J Biol Chem. 2010;285:18352–18363. doi: 10.1074/jbc.M109.077602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wang J, Huang ZG, Cao H, Wang YT, Hui P, Hoo C, Li SP. Screening of anti-platelet aggregation agents from Panax notoginseng using human platelet extraction and HPLC-DAD-ESI-MS/MS. J Sep Sci. 2008;31:1173–1180. doi: 10.1002/jssc.200700507. [DOI] [PubMed] [Google Scholar]
- 29.Charo IF, Feinman RD, Detwiler TC. Interrelations of platelet aggregation and secretion. J Clin Invest. 1977;60:866–873. doi: 10.1172/JCI108841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Park KM, Rhee MH, Shin HJ, Song YB, Hyun HC, Park KH, Cho HJ, Choi SA, Kang HC, Kim KJ, et al. Inhibitory effects of panaxtriol from Panax ginseng C.A. Meyer on phosphoinositide breakdown induced by thrombin in platelets. J Ginseng Res. 2008;32:107–113. [Google Scholar]
- 31.Kimura Y, Okuda H, Arichi S. Effects of various ginseng saponins on 5-hydroxytryptamine release and aggregation in human platelets. J Pharm Pharmacol. 1988;40:838–843. doi: 10.1111/j.2042-7158.1988.tb06285.x. [DOI] [PubMed] [Google Scholar]
- 32.Kuo SC, Teng CM, Lee JC, Ko FN, Chen SC, Wu TS. Antiplatelet components in Panax ginseng. Planta Med. 1990;56:164–167. doi: 10.1055/s-2006-960916. [DOI] [PubMed] [Google Scholar]
- 33.Lee SR, Park JH, Choi KJ, Kim ND. Inhibitory effects of ginsenoside Rg3 on platelet aggregation and its mechanism of action. Korean J Ginseng Sci. 1997;21:132–140. [Google Scholar]
- 34.Eigenthaler M, Nolte C, Halbrugge M, Walter U. Concentration and regulation of cyclic nucleotides, cyclic-nucleotide-dependent protein kinases and one of their major substrates in human platelets. Estimating the rate of cAMP-regulated and cGMP-regulated protein phosphorylation in intact cells. Eur J Biochem. 1992;205:471–481. doi: 10.1111/j.1432-1033.1992.tb16803.x. [DOI] [PubMed] [Google Scholar]
- 35.Ok WJ, Cho HJ, Kim HH, Lee DH, Kang HY, Kwon HW, Rhee MH, Kim M, Park HJ. Epigallocatechin-3-gallate has an anti-platelet effect in a cyclic AMP-dependent manner. J Atheroscler Thromb. 2012;19:337–348. doi: 10.5551/jat.10363. [DOI] [PubMed] [Google Scholar]
- 36.Schwarz UR, Walter U, Eigenthaler M. Taming platelets with cyclic nucleotides. Biochem Pharmacol. 2001;62:1153–1161. doi: 10.1016/s0006-2952(01)00760-2. [DOI] [PubMed] [Google Scholar]
- 37.Kim S, Shim S, Choi DS, Kim JH, Kwon YB, Kwon J. Modulation of LPS-stimulated astroglial activation by ginseng total saponins. J Ginseng Res. 2011;35:80–85. [Google Scholar]
- 38.Shen T, Lee J, Park MH, Lee YG, Rho HS, Kwak YS, Rhee MH, Park YC, Cho JY. Ginsenoside Rp1, a ginsenoside derivative, blocks promoter activation of iNOS and COX-2 genes by suppression of an IKKb-mediated NF-κB pathway in HEK293 cells. J Ginseng Res. 2011;35:200–208. doi: 10.5142/jgr.2011.35.2.200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yayeh T, Jung KH, Jeong HY, Park JH, Song YB, Kwak YS, Kang HS, Cho JY, Oh JW, Kim SK, et al. Korean red ginseng saponin fraction downregulates proinflammatory mediators in LPS stimulated RAW264.7 cells and protects mice against endotoxic shock. J Ginseng Res. 2012;36:263–269. doi: 10.5142/jgr.2012.36.3.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lee JH, Park HJ. Effects of intaking of red ginseng products on human platelet aggregation and blood lipids. J Ginseng Res. 1998;22:173–180. [Google Scholar]
- 41.Jin YR, Yu JY, Lee JJ, You SH, Chung JH, Noh JY, Im JH, Han XH, Kim TJ, Shin KS, et al. Antithrombotic and antiplatelet activities of Korean red ginseng extract. Basic Clin Pharmacol Toxicol. 2007;100:170–175. doi: 10.1111/j.1742-7843.2006.00033.x. [DOI] [PubMed] [Google Scholar]
- 42.Yuan CS, Wang CZ, Wicks SM, Qi LW. Chemical and pharmacological studies of saponins with a focus on American ginseng. J Ginseng Res. 2010;34:160–167. doi: 10.5142/jgr.2010.34.3.160. [DOI] [PMC free article] [PubMed] [Google Scholar]