

Abstract
Stem cell transplantation is a promising approach for improving cardiac function after severe myocardial damage, for which the use of autologous donor cells has been preferred to avoid immune rejection. Recently, however, rodent as well as human mesenchymal stem cells have been reported to be uniquely immune-tolerant, in both in vitro and in vivo transplant models. In this review, we explore in detail the current understanding of the underlying immunologic mechanisms, which can facilitate the use of such cells as “universal donor cells” with fascinating clinical implications.
Keywords: Immunogenicity, Immunosuppression, Marrow stromal stem cells, Mesenchymal stem cells, Stem cell transplantation
Introduction
Myocardial infarction remains a widespread and important cause of morbidity and mortality among adults, accounting for >15 million new cases worldwide every year [1]. The loss of cardiomyocytes that results, combined with the limited endogenous repair mechanism, sets into play the remodeling process that ultimately leads to progressive heart failure. End-stage heart failure still has a grave prognosis, with an estimated 5-year mortality of 70%.
Although current medical and surgical treatments have significantly altered the survival of these patients, none of these modalities contributes new contractile tissue to replace the myocardial scar. A promising approach is cellular transplantation, which is directly aimed at overcoming the problem of myocardial cell loss and improving the function of the injured myocardium through several mechanisms, including myogenesis [2], angiogenesis, and paracrine effects [3, 4], which may attenuate left ventricular function. The observed beneficial effects of cell transplantation have led to numerous human clinical trials in the past several years [5].
The current preferred approach of using autologous stem cells aims to avoid immune rejection of donor cells, which can be expected after allogeneic or xenogeneic transplantation. Despite the promising early results, harvesting autologous cells from individual patients still poses significant logistic, economic, and timing constraints. Furthermore, most of the patients who could benefit from such therapy are elderly patients with multiple medical comorbidities. Unfortunately a number of recent studies have documented that mesenchymal stem cells (MSCs) obtained from elderly donors, and those with diabetes, renal failure, or severe ischemic heart disease, demonstrate significantly reduced capacity for proliferation, differentiation, and neovascularization, with increased levels of apoptosis in vitro and in vivo [6, 7]. Such impaired autologous donor cells from sick elderly patients could therefore limit their therapeutic potential. Thus there would be obvious clinical advantages if “universal donor cells” from healthy young donors could be used for stem cell allotransplantation without the need for immunosuppressive therapies.
In the last several years, increasing experimental findings have pointed toward a unique immunomodulatory property of the MSCs in both the in vitro and in vivo settings [8]. One intriguing property of MSCs is their ability to escape immune recognition and even actively inhibit immune responses. In this chapter, we will briefly review the general properties of MSCs. We will then discuss in depth the evidence behind the role of MSCs in immunomodulation, both in vivo and in vitro, and describe our current understanding of the possible underlying mechanisms by which it occurs and by which MSCs could be viewed as universal donor cells.
MSCs As Adult Stem Cells
Mesenchymal stem cells, also known as bone marrow stem cells, stromal stem cells, marrow progenitor cells, and marrow-derived adult stem cells, essentially represent a heterogeneous population of fibroblast-like cells, which can be found in the bone marrow stroma. Unlike hematopoietic cells, MSCs are CD34− and CD45−. Although still not fully identified, some other typical MSCs markers include CD29, CD44, CD71, CD90, CD106, CD124, SH2, and SH3. A full description of their properties was previously published in great detail by Pittenger et al. [9] and is summarized in Table 1.
Table 1.
Phenotype characterization of human MSCs

Abbreviations: FGFR, fibroblast growth factor receptor; HLA, human leukocyte antigen; ICAM, intercellular adhesion molecule; IL, interleukin; INF, interferon; PDGFR, platelet-derived growth factor receptor; TNF, tumor necrosis factor; VCAM, vascular cell adhesion molecule; VLA, very late activation antigen.
Although the extent of the plasticity of MSCs is still under investigation, studies within the last few years have demonstrated the capacity of these MSCs to differentiate into cells of different lineages [8, 9]. Because these cells do not have the ethical or tumorigenicity problems of embryonic stem cells, their plasticity has generated much excitement, giving hope for their therapeutic use in a wide range of diseases.
MSCs As Universal Donor Cells
Recently there has been an explosive advance in our knowledge of adult stem cells for their use as donor cells for regenerative therapies. Associated with such advances are some unexpected and controversial findings that defy current scientific dogmas. One such dilemma is a series of observations indicating that MSCs are immune-privileged, able to survive and differentiate in immunocompatibility-mismatched allogeneic or even xenogeneic transplant recipients [8]. In fact, a number of laboratories have recently reported that MSCs may have a unique immunological property capable of inducing tolerance in immunocompetent allotransplants or even xenotransplant recipients [10, 11]. The mechanisms of such immunotolerance have been the subject of intense study, and three interrelated candidate mechanisms are emerging [12, 13]. MSCs appear to evade rejection by (a) being hypoimmunogenic, (b) modulating T-cell phenotype, and (c) immunosuppressing the local environment.
Evidence from In Vitro Studies
A large body of in vitro experiments involving coculture mixed lymphocyte reactions supports the view that MSCs avoid allogeneic response [8, 11, 13, 14] and that the use of mismatched MSCs does not induce a proliferative T-cell response in allogeneic and xenogeneic coculture studies [10, 12, 15, 16]. Largely responsible for being hypoimmunogenic is a unique expression pattern of cell-surface antigens on the MSCs. Although there is a continuous controversy surrounding the exact composition of these cell-surface markers, most studies describe MSCs as major histocompatibility complex (MHC) class I positive and MHC class II negative [9]. The expression of class I MHC is important because it protects these cells from certain natural killer (NK)-cell-mediated deletion. As MHC class II proteins are potent alloantigens, their lack of expression on MSCs allows them to escape recognition by effector CD4+ T cells. In addition to this, MSCs do not appear to express Fas-ligand or costimulatory molecules such as B7-1 (CD80), B7-2 (CD86), or CD40 for effector T-cell induction [9]. The presence of these cell-surface markers, along with the finding that MSCs are customary residents of the bone marrow stroma, suggests that MSCs are hypoimmunogenic cells that play an important role in the immunoregulation provided by the bone marrow microenvironment by evading the recognition of alloreactive cells [13].
There is also good in vitro evidence that MSCs can directly modulate the function of T cells. Pittenger and Aggarwal reported that human MSCs constitutively secrete PGE2, hence altering the cytokine secretion profile of dendritic cells, naïve and cytotoxic T lymphocytes, and NK cells, namely, by inhibiting tumor necrosis factor-α and interferon-γ (INF-γ) and by stimulating interleukin 10 (IL-10) secretion to modulate the immune cell response [17]. By doing so, they inhibit the maturation and migration of various antigen-presenting cells, suppress B-cell activation, induce suppressor T-cell formation, and alter the expression of several receptors necessary for antigen capture and processing [13, 17]. Furthermore, through the release of IL-4, they accelerate a shift from a majority of proinflammatory Th1 cells toward an increase in the anti-inflammatory Th2 cells [17]. We suspect that this anti-inflammatory property of MSCs may play an important role in the so-called paracrine effects associated with MSCs, as postulated by several investigators [3, 4].
Although still controversial, there is some evidence that MSCs do also exhibit immunosuppressive properties. Some reports show that MSCs do express mRNA for cytokines such as IL-1, -6, -7, -8, -11, -12, -14, -15, -27, leukemia inhibitory factor, macrophage colony stimulating factor, and stem cell factor [14, 17]. Although their role is still not fully understood, some of these cytokines provide critical cell-cell interactions and promote hemopoietic stem cell differentiation. Furthermore, IL-10 seems to be constitutively expressed by MSCs. This interleukin has a well-documented role in T-cell regulation and in the promotion of the suppressor phenotype and can also antagonize the action of IL-12 during induction of the inflammatory immune responses [17].
In addition to the proposed role of indoleamine 2,3-dioxygenase-mediated tryptophan degradation [14], MSCs can also secrete other peptides, such as hepatocyte growth factor (HGF), which may contribute to the creation of a local immunosuppressive environment [12]. Similarly, transforming growth factor β1 (TGF-β1) also seems involved in T-cell suppression by working with HGF in promoting the allo-escaping phenotype. In fact, Di Nicola et al. showed that neutralizing antibodies to HGF and TGF-β1 restored the proliferative response in mixed lymphocyte reactions [12].
Zhang et al. provided further evidence that MSCs interfere with dendritic cell (DC) maturation, differentiation, and function by downregulating the expression of CD1a, CD40, CD80, CD86, and human leukocyte antigen-DR [18]. These findings were confirmed by Beyth et al. who suggested that human MSCs convert the DC into a suppressor cell, thus locking it into an immature state and thereby inducing peripheral tolerance [19]. All these results suggest that MSCs may be mediating allogeneic tolerance by directing antigen-presenting cells toward a suppressor phenotype that ultimately results in an attenuated T-cell response [13, 14].
Most studies have also shown that these immunosuppressive properties are broad, increase with increased concentration of MSCs [14], and are effective whether the stimulation is specific or nonspecific [12, 15, 16, 20], across species [20, 21], and across different populations of lymphocytes [18, 19, 21]. Together, these results suggest that these immunosuppressive mechanisms may even cross species barriers.
In addition, increasing evidence has emerged that MSCs can also interact directly with T cells to suppress alloreactivity and direct CD4+ T cells to a suppressive phenotype. Di Nicola et al. [12] and Tse et al. [16] showed that MSCs strongly suppressed CD4+ T cells in mixed lymphocyte reactions and attenuated the proliferation of T-cell subsets.
Some studies have also shown that MSCs can influence control over cell division cycle pathways in cells of immunological relevance. Glennie et al. have shown that T cells stimulated in cocultures with MSCs exhibited an extensive inhibition of cyclin-D2 and an upregulation of the cyclin-dependent kinase inhibitor p27kip1 [23].
Furthermore, the role of MSCs on CD8+ T cells and NK cells has also been addressed. There is evidence that MSCs inhibit the formation of CD8+ T cells and appear to evade NK-cell targeting mechanisms [23]. Rasmusson et al. showed that NK cells in cocultures did not recognize MSCs although their lytic properties were still present [22]. Moreover, it has been suggested that MSCs reduce IL-2-induced NK-cell proliferation, and INF-γ production [17, 22, 24, 25].
Finally, another level at which MSCs may modulate immune responses is through the inhibition of B-cell proliferation, as well as their chemotactic behavior and antibody production [8, 23]. Taken together, numerous studies provide strong evidence that MSCs are able to modulate the function of different immune cells in vitro through several interrelated mechanisms, as summarized in Figure 1.
Figure 1.
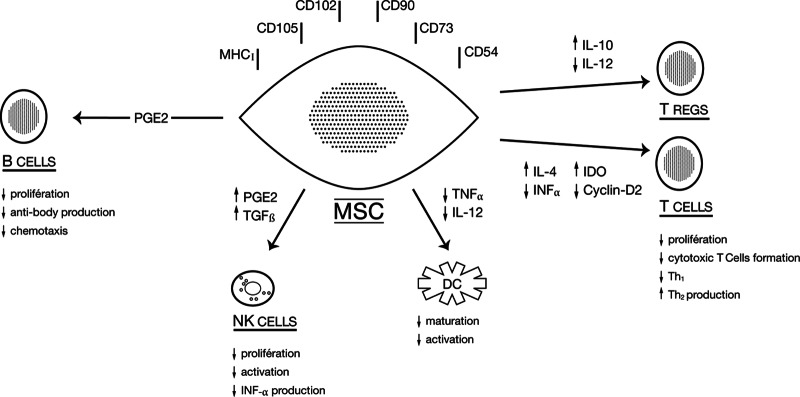
Proposed mechanisms of the immunomodulatory effects of MSCs. Abbreviations: DC, dendritic cell; IDO, indoleamine 2,3-dioxygenase; IL, interleukin; MSC, mesenchymal stem cell; NK, natural killer; PGE2, prostaglandin E2; TGF-β, transforming growth factor β; TNF-α, tumor necrosis factor-α; Tregs, regulatory T cells. Adapted from [8] with permission from Blood.
Evidence from In Vivo Studies
Although considerable data of in vitro findings support the immunomodulatory properties of MSCs, relatively little evidence is available on the immunogenicity of MSCs in vivo. Despite this, there is growing evidence that the in vitro observations may translate to the in vivo setting. Bartholomew et al. [15] first demonstrated that the in vivo administration of allogeneic MSCs prolonged third-party skin graft survival in immunocompetent baboons. This study, as well as many others, paved the way for the use of these cells in immune-mediated disorders [8, 11]. For example, Koç et al. showed no evidence of alloreactive T cells and no incidence of graft versus host disease (GVHD) when allogeneic MSCs were infused into patients with Hurler's syndrome or metachromatic leukodystrophy [26]. Horwitz et al. reported that donor MSCs contributed to bone remodeling after allogeneic stem cell transplantation in three children with osteogenesis imperfecta [27]. Other groups have reported that MSC transplantation can also prevent the rejection of allogeneic B16 mouse melanoma cells in immunocompetent mice [21], successfully engraft in brains of albino rats, lead to significant improvement in symptoms in mice with autoimmune encephalomyelitis through the induction of peripheral tolerance [28], and attenuate GVHD in humans with grade IV acute GVHD [29]. It is important to note that several mechanisms involving specific factors have been proposed, mostly based on in vitro findings, but a clear consensus, based on in vivo experiments specifically targeting these particles, is still lacking [13].
Allogeneic MSC transplants into the myocardium between unrelated porcine donors and recipients was reported by Caparelli et al. [30] and by Makkar et al. [31]. The recipients underwent coronary ligation before allogeneic MSC implantation and received no immunosuppression. The implanted cells remained viable and differentiated without being rejected. Furthermore, the cardiac functions of transplanted hearts were significantly improved. In a swine and a rat model, Amado et al. [32] and Dai et al. [3], respectively, reported the survival of allogeneic MSCs in infarcted myocardium without immunosuppression. Moreover, these cells were shown to differentiate and contribute to the functional improvement of the host myocardium [33].
In November 2000, a fascinating study by Liechty et al. was published in Nature Medicine [34]. Well-characterized human MSCs were implanted into fetal sheep early in gestation. In this xenogeneic system, the human MSCs engrafted and persisted in multiple tissues for as long as 13 months after transplantation, even after maturation of the fetal immune system. Furthermore, these cells underwent site-specific differentiation into multiple cell lineages, including cardiomyocytes. Nevertheless, these experiments were carried out in fetal recipients.
In a series of studies at our laboratory, Saito et al. injected intravenously labeled mouse MSCs into fully immunocompetent adult rats, successfully producing stable cardiac chimeras for at least 12 weeks without any immunosuppression and with no evidence of rejection [35]. It was confirmed that within days these mouse cells had homed into the bone marrow of the rats and, upon coronary artery ligation, were recruited to the peri-infarcted myocardium. In the following 4–6 weeks, the labeled cells were seen to differentiate into various phenotypes. In subsequent studies, MacDonald et al. showed that not only were stable chimeras formed but also the overall ventricular function was significantly improved [36]. These findings were once again replicated by Luo et al., who confirmed the survival of pig MSCs implanted into fully immunocompetent rat myocardium for up to 6 months after xenotransplantation [37]. More recently, Atoui et al. were able to confirm the engraftment of human MSCs within the rat myocardium for at least 8 weeks after myocardial infarction, without the use of any immunosuppression [38]. Such xenotransplant significantly contributed to the improvement in the overall cardiac function and in attenuating left ventricular remodeling.
However, tolerance of MSCs across the MHC barrier might not be absolute. Grinnemo et al. [39] demonstrated that, although the MSCs successfully engraft across allogeneic barriers, rejection occurs when a xenotransplant model is used. In their follow-up study, the same group demonstrated that the survival of human MSCs into ischemic rat myocardium is possible only when immunosuppression is used [40]. These findings were in direct contrast to those obtained in our laboratory. Despite the similarities between our two studies, nonetheless there seems to be subtle differences in the experimental designs. For instance, in their study, MSCs were harvested from the sternum of patients undergoing cardiac surgery. These cells, taken from elderly donors, were previously shown to have a significantly lower capacity for differentiation, angiogenesis, survival, and even proliferation [6, 7, 41]. It is of interest to note that, in the in vitro studies, human MSCs used were instead harvested from young healthy donors. Furthermore, other experimental differences linked to the amount of fetal calf serum present in the culture media could also partially explain these differences [42]. Still, further studies are needed to better clarify these contradictory findings.
Other opposing findings were also reported showing that, despite retaining their immunosuppressive properties in vitro, allogeneic murine MSCs can be immunogenic in immunocompetent animals [43, 44]. The discrepancy observed could be observed by differences in the experimental conditions such as the level of INF-γ [45], different stages of differentiation, or species diversity [13]. Furthermore, it is also important to note that there is typically a low retention yield when MSCs are directly injected within the myocardium. This is mostly attributed to mechanical loss secondary to leakage and washout [46]. This could also partially explain the different results sometimes observed among different studies. Finally, it should also be mentioned that murine and human MSCs differ in their immunosuppressive property. In fact, it was shown that the immunosuppressive effect of human MSCs, at least in vitro, is much stronger than that of murine MSCs [12].
Such contradictory findings are perplexing but not unique in this rapidly developing field of stem cell biology and regenerative medicine. Despite the substantive body of evidence from the in vitro literature confirming the immunomodulatory properties of MSCs, their importance in the in vivo setting remains controversial. Nevertheless, MSCs have already been introduced to clinical practice, especially in the autoimmune and hematological fields [47].
Furthermore, although the immunomodulatory effects of MSCs are now well-documented, they provide no explanation as to why such tolerance persists even after the implanted stem cells differentiate into their targeted tissue phenotypes. In an attempt to explain this phenomenon, Chiu proposed the “stealth immune tolerance” hypothesis [33], which is in fact an application of the “danger model” theory described earlier by Matzinger [48], who suggested that the immune rejection of a transplanted organ is not due to the mismatch of MHC antigens alone but due to the presence of a “danger signal” serving as a costimulant factor. Thus, although it is still hypothetical, the stealth immune tolerance hypothesis is based on the fact that the expression of new foreign recognition antigens (that is, MHC antigens) on the gradually differentiating cells is dissociated in timing from the danger signals derived from the injury inflicted by the invasive implantation procedure. In other words, it takes weeks for the implanted cells to mature and fully express their MHC antigens. Thus, by the time these implanted cells differentiate, the effects on tissue injury would have subsided, so that the immune synapsis receives only the first “recognition” signal without the second “activation” signal [48]. According to the two-signal theory for immune synapsis, recognition without activation could then lead to T-cell anergy, such that the implanted cells, now fully differentiated, are tolerated and allowed to survive. It is important to note that presently this view remains hypothetical and needs to be further confirmed.
Clinical Trials Involving the Allogeneic Use of MSCs
Interestingly, and based on the clinical and experimental data discussed previously, there have been two clinical trials so far that looked at the allogeneic use of MSCs in patients post myocardial infarction. The FDA recently approved an Osiris Inc. sponsored phase I multicenter clinical trial in which allogeneic human MSCs were given intravenously, without immunosuppression, to patients following an acute myocardial infarction. The preliminary results after 6 months were presented and are highly encouraging [49], with evidence of significant improvement in the ventricular function. Similarly, the Revascor trial is another randomized, placebo-controlled multicenter trial that assesses the safety and feasibility of three different doses of allogeneic MSCs delivered into the myocardium of patients with chronic heart failure. After 3 months, their preliminary results were also reported online (http://www.angioblast.com) and described a significant improvement in the overall ventricular function [50]. More importantly, promising results of the phase II trial were recently reported at the 2011 meeting of the American Heart Association (held in Orlando, FL). On the basis of this, phase III trials are expected to start within the upcoming 6 months.
Conclusion
The potential importance of these findings for the treatment of ischemic heart disease is apparent. In addition to their powerful replicative capacity, MSCs can easily be harvested from bone marrows, expanded ex vivo, and differentiated into many cell type lineages, if desired. Because of their immunotolerance property, the establishment of MSCs as effective universal donor cells [13] could then dramatically expand the therapeutic potential for cellular cardiomyoplasty. From a clinical perspective, these cells could be isolated and expanded from donors irrespective of their MHC haplotype, tested for their functional capabilities well in advance, and stored as an “off-the-shelf” cell population for immediate use when needed on any patient after an acute myocardial infarction. Such logistical advantages are not available with the use of autologous MSCs, which is currently the cell source of choice. Perhaps more importantly, since such allogeneic MSCs can be obtained from young healthy donors, they could have great value in patients with genetic cardiomyopathies and in elderly patients with multiple medical comorbidities whose own MSCs could be dysfunctional.
Despite the exciting preliminary results, further investigations are required to address many of the remaining controversial findings, as well as the important question of chronic rejection after cell transplantation. Although the preliminary results of allogeneic MSC transplantation described seem quite promising, the trials enrolled only a limited number of patients who were evaluated relatively shortly after the treatment. Furthermore, further mechanistic studies and more quantitative assessment of MSCs engraftment are still needed before the therapeutic promise of these cells can be fully achieved.
Author Contributions
R.A.: conception and design, financial support, collection of data, data interpretation, manuscript writing, final approval; R.C.J.C.: financial support, data interpretation, final approval.
Disclosure of Potential Conflicts of Interest
The authors indicate no potential conflicts of interest.
References
- 1.Orlic D, Hill J, Arai A. Stem cells for myocardial regeneration. Circ Res. 2002;91:1092–1102. doi: 10.1161/01.res.0000046045.00846.b0. [DOI] [PubMed] [Google Scholar]
- 2.Orlic D, Kajstura J, Chimenti S, et al. Bone marrow cells regenerate infarcted myocardium. Nature. 2001;401:701–705. doi: 10.1038/35070587. [DOI] [PubMed] [Google Scholar]
- 3.Dai W, Hale S, Martin B, et al. Allogeneic mesenchymal stem cell transplantation in postinfarcted rat myocardium: Short and long-term effects. Circulation. 2005:112–214-23. doi: 10.1161/CIRCULATIONAHA.104.527937. [DOI] [PubMed] [Google Scholar]
- 4.Tang Y, Zhao Q, Qin X, et al. Paracrine action enhances the effects of autologous mesenchymal stem cell transplantation on vascular regeneration in rat model of myocardial infarction. Ann Thorac Surg. 2005;80:229–237. doi: 10.1016/j.athoracsur.2005.02.072. [DOI] [PubMed] [Google Scholar]
- 5.Schächinger V, Erbs S, Elsasser A, et al. Intra-coronary bone marrow derived progenitor cells in acute myocardial infarction. N Engl J Med. 2006;355:1210–1221. doi: 10.1056/NEJMoa060186. [DOI] [PubMed] [Google Scholar]
- 6.Zhuo Y, Li SH, Chen MS, et al. Aging impairs the angiogenic response to ischemic injury and the activity of implanted cells: Combined consequences for cell therapy in older recipients. J Thorac Cardiovasc Surg. 2010;139:1286–1294. doi: 10.1016/j.jtcvs.2009.08.052. [DOI] [PubMed] [Google Scholar]
- 7.Nayan M, Paul A, Chen G. Superior therapeutic potential of young bone marrow mesenchymal stem cells by direct intramyocardial delivery in aged recipients with acute myocardial infarction: In vitro and in vivo investigation. J Tissue Eng. 2011;2011:741213. doi: 10.4061/2011/741213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nauta AJ, Fibbe WE. Immunomodulatory properties of mesenchymal stromal cells. Blood. 2007;110:3499–3506. doi: 10.1182/blood-2007-02-069716. [DOI] [PubMed] [Google Scholar]
- 9.Pittenger MF, MacKay AM, Beck SC, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143–147. doi: 10.1126/science.284.5411.143. [DOI] [PubMed] [Google Scholar]
- 10.Le Blanc K, Tammik C, Rosendahl K, et al. HLA expression and immunologic properties of differentiated and undifferentiated mesenchymal stem cells. Exp Hematol. 2003;31:890–896. doi: 10.1016/s0301-472x(03)00110-3. [DOI] [PubMed] [Google Scholar]
- 11.Le Blanc K, Ringden O. Immunomodulation by mesenchymal stem cells and clinical experience. J Intern Med. 2007;262:509–525. doi: 10.1111/j.1365-2796.2007.01844.x. [DOI] [PubMed] [Google Scholar]
- 12.Di Nicola M, Carlo-Stella C, Magni M, et al. Human bone marrow stromal cells suppress T lymphocyte proliferation induced by cellular or non-specific mitogenic stimuli. Blood. 2002;99:3838–3843. doi: 10.1182/blood.v99.10.3838. [DOI] [PubMed] [Google Scholar]
- 13.Atoui R, Shum-Tim D, Chiu RCJ. Myocardial regenerative therapy: Immunologic basis for the potential “universal donor cells.”. Ann Thorac Surg. 2008;86:327–334. doi: 10.1016/j.athoracsur.2008.03.038. [DOI] [PubMed] [Google Scholar]
- 14.Rasmusson I. Immune modulation by mesenchymal stem cells. Exp Cell Res. 2006;312:2169–2179. doi: 10.1016/j.yexcr.2006.03.019. [DOI] [PubMed] [Google Scholar]
- 15.Bartholomew A, Sturgeon C, Satskas M, et al. Mesenchymal stem cells suppress lymphocyte proliferation in vitro and prolong skin graft survival in vivo. Exp Hematol. 2002;30:42–48. doi: 10.1016/s0301-472x(01)00769-x. [DOI] [PubMed] [Google Scholar]
- 16.Tse WT, Pendleton JD, Beyer WM, et al. Suppression of allogeneic T-cell proliferation by human marrow stromal cells: Implication in transplantation. Transplantation. 2003;75:389–397. doi: 10.1097/01.TP.0000045055.63901.A9. [DOI] [PubMed] [Google Scholar]
- 17.Aggarwal S, Pittenger M. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood. 2005;105:1815–1822. doi: 10.1182/blood-2004-04-1559. [DOI] [PubMed] [Google Scholar]
- 18.Zhang W, Ge W, Li C, et al. Effects of mesenchymal stem cells on differentiation, maturation, and function of human monocyte-derived dendritic cells. Stem Cell Dev. 2004;13:263–271. doi: 10.1089/154732804323099190. [DOI] [PubMed] [Google Scholar]
- 19.Beyth S, Borovsky Z, Mevorach D, et al. Human mesenchymal stem cells alter antigen-presenting cell maturation and induce T cell unresponsiveness. Blood. 2005;105:2214–2219. doi: 10.1182/blood-2004-07-2921. [DOI] [PubMed] [Google Scholar]
- 20.Maccario R, Podesta M, Moretta A, et al. Interaction of human mesenchymal stem cells with cells involved in alloantigen-specific immune response favors the differentiation of CD4+ T-cell subsets expressing a regulatory/suppressive phenotype. Haematologica. 2005;90:516–525. [PubMed] [Google Scholar]
- 21.Djouad F, Plence P, Bony C, et al. Immunosuppressive effect of mesenchymal stem cells favors tumor growth in allogeneic animals. Blood. 2003;102:3837–3844. doi: 10.1182/blood-2003-04-1193. [DOI] [PubMed] [Google Scholar]
- 22.Rasmusson I, Ringden O, Sundberg B, et al. Mesenchymal stem cells inhibit the formation of cytotoxic T lymphocytes, but not activated cytotoxic T lymphocytes or natural killer cells. Transplantation. 2003;76:1208–1213. doi: 10.1097/01.TP.0000082540.43730.80. [DOI] [PubMed] [Google Scholar]
- 23.Glennie S, Soeiro I, Dyson PJ, et al. Bone marrow mesenchymal stem cells induce division arrest anergy of activated T cells. Blood. 2005;105:2821–2827. doi: 10.1182/blood-2004-09-3696. [DOI] [PubMed] [Google Scholar]
- 24.Kassis I, Vaknin-Dembinsky A, Karussis D. Bone marrow mesenchymal stem cells: Agents of immunomodulation and neuroprotection. Curr Stem Cell Res Ther. 2011;6:63–68. doi: 10.2174/157488811794480762. [DOI] [PubMed] [Google Scholar]
- 25.Spaggiari GM, Capobianco A, Becchetti S, et al. Mesenchymal stem cell-natural killer cell interactions: Evidence that activated NK cells are capable of killing MSCs, whereas MSCs can inhibit IL-2 induced NK-cell proliferation. Blood. 2006;107:1484–1490. doi: 10.1182/blood-2005-07-2775. [DOI] [PubMed] [Google Scholar]
- 26.Koç ON, Day J, Nieder M, et al. Allogeneic mesenchymal stem cell infusion for treatment of metachromatic leukodystrophy and Hurler syndrome. Bone Marrow Transplant. 2002;30:215–222. doi: 10.1038/sj.bmt.1703650. [DOI] [PubMed] [Google Scholar]
- 27.Horwitz EM, Gordon PL, Koo WK, et al. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: Implications for cell therapy of bone. Proc Natl Acad Sci U S A. 2002;99:8932–8937. doi: 10.1073/pnas.132252399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zappia E, Casazza S, Pedemonte E, et al. Mesenchymal stem cells ameliorate experimental autoimmune encephalomyelitis inducing T-cell anergy. Blood. 2005;106:1755–1761. doi: 10.1182/blood-2005-04-1496. [DOI] [PubMed] [Google Scholar]
- 29.Le Blanc K, Rasmusson J, Sundberg B, et al. Treatment of severe acute graft-versus-host disease with third party haploidentical mesenchymal stem cells. Lancet. 2004;363:1439–1441. doi: 10.1016/S0140-6736(04)16104-7. [DOI] [PubMed] [Google Scholar]
- 30.Caparelli DJ, Cattaneo SM, Shake JG, et al. Cellular cardiomyoplasty with allogeneic mesenchymal stem cells results in improved cardiac performance in a swine model of myocardial infarction. Circulation. 2001;104:II–599. [Google Scholar]
- 31.Makkar RR, Price M, Lill M, et al. Intramyocardial injection of allogeneic bone marrow-derived mesenchymal stem cells without immunosuppression preserves cardiac function in a porcine model of myocardial infarction. J Cardiovasc Pharmacol Ther. 2005;10:225–233. doi: 10.1177/107424840501000403. [DOI] [PubMed] [Google Scholar]
- 32.Amado LC, Saliaris AP, Schuleri KH, et al. Cardiac repair with intramyocardial injection of allogeneic mesenchymal stem cells after myocardial infarction. Proc Natl Acad Sci U S A, 2005;102:11474–11479. doi: 10.1073/pnas.0504388102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chiu R. “Stealth immune tolerance” in stem cell transplantation: Potential for “universal donors” in myocardial regenerative therapy. J Heart Lung Transplant. 2005;24:511–516. doi: 10.1016/j.healun.2004.11.010. [DOI] [PubMed] [Google Scholar]
- 34.Liechty KW, MacKenzie TC, Shaaban AF, et al. Human mesenchymal stem cells engraft and demonstrate site-specific differentiation after in utero transplantation in sheep. Nat Med. 2000;6:1282–1286. doi: 10.1038/81395. [DOI] [PubMed] [Google Scholar]
- 35.Saito T, Kuang JQ, Bittira B, et al. Xenotransplant cardiac chimera: Immune tolerance of adult stem cells. Ann Thorac Surg. 2002;74:19–24. doi: 10.1016/s0003-4975(02)03591-9. [DOI] [PubMed] [Google Scholar]
- 36.MacDonald D, Saito T, Shum-Tim D, et al. Persistence of marrow stromal cells implanted into acutely infarcted myocardium: Observations in a xenotransplant model. J Thorac Cardiovasc Surg. 2005;130:1114–1121. doi: 10.1016/j.jtcvs.2005.04.033. [DOI] [PubMed] [Google Scholar]
- 37.Luo J, Shum-Tim D, Chiu RCJ. Marrow stromal cells as universal donor cells for cardiac regenerative therapy: Fact or fancy? In: Dimarakis I, et al., editors. Stem Cell Therapy in Ischemic Heart Disease. London: Imperial College Press; 2009. pp. 117–137. [Google Scholar]
- 38.Atoui R, Asenjo JF, Duong M, et al. Marrow stromal cells as “universal donor cells” for myocardial regenerative therapy: Their unique immune tolerance. Ann Thorac Surg. 2008;85:571–580. doi: 10.1016/j.athoracsur.2007.10.034. [DOI] [PubMed] [Google Scholar]
- 39.Grinnemo K, Mansson A, Dellgren D, et al. Xenoreactivity and engraftment of human mesenchymal stem cells transplanted into infarcted rat myocardium. J Thorac Cardiovasc Surg. 2004;127:1293–1300. doi: 10.1016/j.jtcvs.2003.07.037. [DOI] [PubMed] [Google Scholar]
- 40.Grinnemo KH, Mansson-Broberg A, Leblanc K, et al. Human mesenchymal stem cells do not differentiate into cardiomyocytes in a cardiac ischemic xenomodel. Ann Med. 2006;38:144–153. doi: 10.1080/07853890500422982. [DOI] [PubMed] [Google Scholar]
- 41.Stolzing A, Scutt A. Age-related impairment of mesenchymal progenitor cell function. Aging Cell. 2006;5:213–224. doi: 10.1111/j.1474-9726.2006.00213.x. [DOI] [PubMed] [Google Scholar]
- 42.Javazon EH, Beggs KJ, Flake AW. Mesenchymal stem cells: Paradoxes of passaging. Exp Hematol. 2004;32:414–425. doi: 10.1016/j.exphem.2004.02.004. [DOI] [PubMed] [Google Scholar]
- 43.Eliopoulos N, Stagg J, Lejeune L, et al. Allogeneic marrow stromal cells are immune rejected by MHC class I-and class II-mismatched recipient mice. Blood. 2005;106:4057–4065. doi: 10.1182/blood-2005-03-1004. [DOI] [PubMed] [Google Scholar]
- 44.Nauta AJ, Westerhuis G, Kruisselbrink AB, et al. Donor-derived mesenchymal stem cells are immunogenic in an allogeneic host and stimulate donor graft rejection in a nonmyeloablative setting. Blood. 2006;108:2114–2120. doi: 10.1182/blood-2005-11-011650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Stagg J, Pommey S, Eliopoulos N, et al. Interferon-γ-stimuated marrow stromal cells: A new type of nonhematopoietuc antigen-presenting cell. Blood. 2006;107:2570–2577. doi: 10.1182/blood-2005-07-2793. [DOI] [PubMed] [Google Scholar]
- 46.Teng CJ, Luo J, Chiu RC, et al. Massive mechanical loss of microspheres with direct intramyocardial injection in the beating heart: Implications for cellular cardiomyoplasty. J Thorac Cardiovasc Surg. 2006;132:628–632. doi: 10.1016/j.jtcvs.2006.05.034. [DOI] [PubMed] [Google Scholar]
- 47.Prasad VK, Lucas KG, Kleiner GI, et al. Efficacy and safety of ex vivo cultured adult human mesenchymal stem cells (Prochymal) in pediatric patients with severe refractory acute graft-versus-host disease in a compassionate use study. Biol Blood Marrow Transplant. 2011;17:534–541. doi: 10.1016/j.bbmt.2010.04.014. [DOI] [PubMed] [Google Scholar]
- 48.Matzinger P. The danger model: A renewed sense of self. Science. 2002;296:301–305. doi: 10.1126/science.1071059. [DOI] [PubMed] [Google Scholar]
- 49.Zambrano J, Traverse J, Henry T, et al. The impact of intravenous human mesenchymal stem cells (Provacel) on ejection fraction in patients with myocardial infarction. Circulation. 2007;116:II–202. [Google Scholar]
- 50.Kinkaid HY, Huang XP, Li RK, et al. What's new in cardiac cell therapy? Allogeneic bone marrow stromal cells as “universal donor cells.”. J Card Surg. 2010;25:359–366. doi: 10.1111/j.1540-8191.2009.00984.x. [DOI] [PubMed] [Google Scholar]