

Abstract
Both neurotrophic support and neural activity are required for normal postnatal development and survival of cochlear spiral ganglion (SG) neurons. Previous studies in neonatally deafened cats demonstrated that electrical stimulation (ES) from a cochlear implant can promote improved SG survival but does not completely prevent progressive neural degeneration. Neurotrophic agents combined with an implant may further improve neural survival. Short-term studies in rodents have shown that brain-derived neurotrophic factor (BDNF) promotes SG survival after deafness and may be additive to trophic effects of stimulation. Our recent study in neonatally deafened cats provided the first evidence of BDNF neurotrophic effects in the developing auditory system over a prolonged duration Leake et al. (J Comp Neurol 519:1526–1545, 2011). Ten weeks of intracochlear BDNF infusion starting at 4 weeks of age elicited significant improvement in SG survival and larger soma size compared to contralateral. In the present study, the same deafening and BDNF infusion procedures were combined with several months of ES from an implant. After combined BDNF + ES, a highly significant increase in SG numerical density (>50 % improvement re: contralateral) was observed, which was significantly greater than the neurotrophic effect seen with ES-only over comparable durations. Combined BDNF + ES also resulted in a higher density of myelinated radial nerve fibers within the osseous spiral lamina. However, substantial ectopic and disorganized sprouting of these fibers into the scala tympani also occurred, which may be deleterious to implant function. EABR thresholds improved (re: initial thresholds at time of implantation) on the chronically stimulated channels of the implant. Terminal electrophysiological studies recording in the inferior colliculus (IC) revealed that the basic cochleotopic organization was intact in the midbrain in all studied groups. In deafened controls or after ES-only, lower IC thresholds were correlated with more selective activation widths as expected, but no such correlation was seen after BDNF + ES due to much greater variability in both measures.
Keywords: auditory deprivation, auditory nerve, BDNF, cochlear implant, cochlear spiral ganglion, electrical stimulation, neonatal deafness, primary afferents, neurotrophins
Introduction
The cochlear spiral ganglion (SG) cells are bipolar primary afferent neurons that relay auditory information from the hair cells to the central auditory system. In cats, each inner hair cell is innervated by a single synapse from ten to 20 type I SG neurons (Kiang et al. 1982; Liberman et al. 1990; Spoendlin 1969, 1975, 1981). Type I cells comprise roughly 95 % of the SG population; they provide the main information pathway to the central auditory system (Kiang et al. 1982) and therefore represent the neural substrate for cochlear implant (CI) function. In sensorineural hearing loss, the hair cells degenerate, the distal dendrites of the SG neurons (radial nerve fibers) begin to retract, and subsequently the ganglion cell somata in the cochlear modiolus also degenerate.
In cultured SG neurons, depolarization elicited by elevated potassium strongly promotes the survival of SG neurons (Hansen et al. 2001, 2003; Hegarty et al. 1997; Roehm and Hansen 2005). Electrical stimulation from a CI has also been shown to promote SG survival in vivo in deafened adult guinea pigs (Hartshorn et al. 1991; Kanzaki et al. 2002; Lousteau 1987; Miller 2001; Miller and Altschuler 1995; Mitchell et al. 1997) and in cats deafened early in life (Leake et al. 1999, 2007, 2008b). In our laboratory, studies of cats deafened prior to hearing onset have shown that multichannel CI electrical stimulation applied over several months results in substantial improvement in SG neural survival (Leake et al. 1999, 2007, 2008a) above that seen in the contralateral deafened cochleae. Other studies, however, have not found evidence of trophic effects of electrical stimulation (Araki et al. 1998; Coco et al. 2007; Li et al. 1999; Shepherd et al. 1994). These disparities in findings suggest that differences among animal models, methods of stimulation (e.g., the position of stimulating electrodes, applied signals, and efficacy in exciting neurons across a broad sector of the spiral ganglion), and/or methods of analysis of SG density are critically important. But even in studies showing highly significant trophic effects, electrical stimulation only partly prevents SG degeneration after deafness (Leake et al. 2008b). Thus, there has been considerable recent interest in neurotrophic agents that might be used in conjunction with cochlear implantation to enhance SG and auditory nerve survival (see Staecker et al. 2010). Of particular interest is the nerve growth factor (NGF) family of proteins, or neurotrophins (NTs), including NGF, brain-derived neurotrophic factor (BDNF), neurotrophin-3 (NT-3), and neurotrophin-4/5, which bind to specific high-affinity receptors of the Trk family. The intracellular signaling mechanisms underlying the survival-promoting effects of both neurotrophins and depolarization have been elucidated by in vitro studies of SG neurons, which indicate that multiple mechanisms underlie neural protection (Hansen et al. 2001; Hegarty et al. 1997; Wefstaedt et al. 2005; Zha et al. 2001). Specifically, L-type voltage-gated Ca2+ channels mediate the survival-promoting effect of depolarization through multiple distinct signaling pathways, including an autocrine neurotrophin mechanism, cAMP production, and Ca2+/calmodulin-dependent protein kinase-mediated phosphorylation of the transcription factor CREB. In addition, the neurotrophins BDNF and NT-3 are expressed by immature SG neurons and promote their survival by an autocrine mechanism that is additive with depolarization effects. Thus, we assume that electrical stimulation in our prior animal studies engaged the same mechanisms in vivo, and given that exogenous NTs activate independent signaling pathways, we hypothesize that intracochlear infusion of NTs may elicit trophic effects that are additive to survival-promoting effects of electrical stimulation.
Many previous studies have reported that exogenous NTs delivered directly to the cochlea in adult animals can protect SG neurons and promote their survival after various insults (Ramekers et al. 2012). Recent studies have shown highly significant effects of BDNF in promoting SG survival (Agterberg et al. 2008; Glueckert et al. 2008; Miller et al. 2007; Shepherd et al. 2008; Wise et al. 2005), and trophic effects also have been reported with other neurotrophic factors such as glial cell line-derived neurotrophic factor (GDNF) (Kanzaki et al. 2002; Maruyama et al. 2008; Yagi et al. 2000; Ylikoski et al. 1998) and fibroblast growth factor (FGF) (Glueckert et al. 2008). Other studies have reported that the survival-promoting effects of NTs are enhanced by concomitant electrical stimulation and that these effects can be maintained by stimulation after cessation of NT infusion (Kanzaki et al. 2002; Shepherd et al. 2005, 2008).
Because neurotrophins regulate neuronal differentiation and survival during development (Fariñas et al. 2001; Fritzsch et al. 1999; Gao et al. 1995; Korsching 1993; Rubel and Fritzsch 2002; Tessarollo et al. 2004; Yang et al. 2011), we hypothesized that exogenous NTs might be particularly effective in our developing animals, after early-onset hearing loss. In our initial study, cats were deafened as neonates, implanted unilaterally and received 10 weeks of intracochlear BDNF starting at 4 weeks of age. This elicited significant improvement in SG survival compared to contralateral (Leake et al. 2011). In fact, BDNF treatment maintained neural survival equivalent to deafened controls examined at 4 weeks of age (age at start of treatment). Thus, the neurotrophic effects of BDNF previously reported in rodents also occur in developing animals and in a non-rodent species that shows a considerably slower rate of SG neuronal cell loss (Leake and Hradek 1988) and may better model the slow SG degeneration in the human cochlea.
Although these findings in animal studies are encouraging, a key issue for potential clinical application of NTs is whether neurotrophic effects persist after terminating delivery of exogenous NTs. One previous study in guinea pigs reported accelerated degeneration of SG neurons after cessation of BDNF administration (Gillespie et al. 2003), but other reports have shown maintenance of improved neural survival after NT treatment is terminated (Agterberg et al. 2009). Another study reported that electrical stimulation maintained improved SG survival for up to 6 weeks after cessation of BDNF delivery in guinea pigs, but neurotrophic effects were observed only in the basal cochlea near the stimulating electrode, and accelerated neural degeneration was seen in the upper cochlear turns (Shepherd et al. 2008). Because NTs would be used with a CI, it is of particular interest to determine whether electrical stimulation (ES) can provide long-term trophic support after a limited duration of intracochlear NT infusion. In the present study, we replicated our earlier BDNF experiment, by deafening cats as neonates and delivering BDNF for 10 weeks starting at 4 weeks of age, but animals in the current study also received ES beginning at about 8 weeks of age and continuing for several months after termination of BDNF delivery.
Our data demonstrate that this combined BDNF infusion and ES (BDNF + ES) elicited highly significant neurotrophic effects, with substantially higher SG cell numerical densities and larger SG cell somata observed in implanted cochleae compared to the contralateral deafened ears. Other findings after combined BDNF + ES included a higher density of radial nerve fibers in the osseous spiral lamina, ectopic and disorganized sprouting of fibers into the scala tympani, angiogenesis within the SG in Rosenthal’s canal, and an increase in the thickness of the fibrotic tissue encapsulating the CI electrode. Interestingly, a reduction in electrically evoked auditory brainstem response (EABR) thresholds relative to initial thresholds was observed for the two chronically stimulated channels of the CI but not for a non-stimulated control channel. Functional effects of BDNF treatment were assessed in terminal electrophysiological experiments, recording from the inferior colliculus (IC) in these same animals. These data demonstrated that final EABR thresholds were correlated with minimum neural thresholds in the IC. Moreover, the fundamental cochleotopic (frequency) organization of the IC was clearly maintained not only in animals studied as adults after BDNF and/or ES, but also in untreated controls, despite deafening prior to hearing onset. Finally, IC thresholds were correlated with activation widths measured at 2 dB above threshold in deafened controls and in subjects receiving only ES, but not in BDNF-treated subjects in which much greater variability was observed. We suggest that this may be due to the ectopic sprouting of radial nerve fibers and is likely to be deleterious to optimum CI function.
Methods
All procedures involving animals were approved by the Institutional Animal Care and Use Committee at the University of California, San Francisco and conformed to all NIH guidelines. The animals included in this study were bred in a closed colony maintained at the University.
Experimental groups and deafening histories
The deafened animals included in this study received daily injections of an ototoxic aminoglycoside antibiotic, neomycin sulfate (60 mg/kg, SQ, SID). Neomycin administration started 1 to 3 days after birth (depending on birth weight) and continued until kittens were 16 days old, at which time auditory brainstem response (ABR) testing was performed as previously described (Leake et al. 2011). Briefly, acoustic stimuli (100 μs clicks, 20/s) were delivered by a canister headphone (STAX, model SMR-1/MK-2) through a hollow ear bar inserted into the external ear canal. If a profound hearing loss was documented (absence of ABR at 90 dB peak SPL), neomycin injections were terminated. If residual hearing was observed, neomycin injections were continued in increments of 2 to 3 days, and ABR testing repeated until the hearing loss was profound. Age at termination of neomycin injections ranged from 16 to 21 days in individual subjects (Table 1).
TABLE 1.
Detailed experimental histories and SG numerical density data are shown for individual cats in the BDNF + ES and ES-only groups for which histological data are presented
| Subject | No. of days (neomycin) | Age at implant (weeks) | Age at Stimulation (weeks) | Type of stimulation | Duration of stimulation (weeks) | Age at study (weeks) | SG density (% normal) | |
|---|---|---|---|---|---|---|---|---|
| Implant | Contralateral | |||||||
| Neonatally deafened cats: BDNF infusion and ES | ||||||||
| #1–K313 | 16 | 5 | 10 | SPa | 14 | 24 | 75 % | 50 % |
| #2–K318 | 16 | 5 | 8 | SPa | 17 | 25 | 76 % | 43 % |
| #3–K319b | 16 | 5 | 8 | SPa | 21 | 29 | 57 % | 37 % |
| #4–K324 | 15 | 4.5 | 7 | SPa | 26 | 33 | 55 % | 40 % |
| #5–K329 | 21 | 4 | 7 | 300 pps/30 Hzc | 22.5 | 30 | 88 % | 56 % |
| Mean (n = 5) | 17 | 5 | 8 | 20 | 28.2 | 70 % | 45 % | |
| Neonatally deafened cats: ES-only controls | ||||||||
| #1–K352 | 16 | 6 | 8 | SPa | 15.5 | 24 | 69 % | 56 % |
| #2–K335 | 16 | 7 | 9 | SPa | 16.5 | 26 | 78 % | 57 % |
| #3–K83 | 19 | 9 | 10 | 80 pps | 22 | 32 | 66 % | 45 % |
| #4–K101 | 18 | 9 | 8 | 300 pps/30 Hzc | 29 | 37 | 50 % | 34 % |
| #5–K89 | 19 | 9 | 10 | 300 pps/30 Hzc | 22 | 31 | 53 % | 43 % |
| Mean (n = 5) | 18 | 8 | 9 | 21 | 30 | 63 % | 47 % | |
All animals received electrical stimulation simultaneously on two bipolar channels of the cochlear implant (see text). Two subjects in the ES-only control group (K352, K335) were implanted and stimulated concurrently with the BDNF + ES group. The remaining three subjects were selected from a previous study to match as closely as possible the type and duration of stimulation and age at study of the subjects in the BDNF + ES group. The subjects in the ES-only group are listed in the same order as the subjects they were selected to match in the BDNF + ES group
aSP, chronic stimulation was delivered by two channels of a CII™ Speech Processor (see text)
bIn subject K319, electrode wire #5 failed 1 month after implantation, and stimulation was subsequently delivered on electrodes #4 and #6
c300 pps/30 Hz, chronic stimulation was a continuous train of biphasic pulses (200 μs/phase) delivered at 300 pps and 100 % sinusoidally amplitude modulated at 30 Hz
BDNF, osmotic pumps, and cochlear implant electrodes
The cochlear implant electrodes used in this study contained an integrated miniature cannula for intracochlear drug delivery and have been described in detail previously (Rebscher et al. 2007). In our initial BDNF study (Leake et al. 2011), we used a shortened version of this electrode that extended only about 6 mm into the scala tympani and had two to four stimulating contacts. In the experiments reported here, we used a later generation, longer electrode array with a spiral-shaped silicone carrier molded to fit within the cat scala tympani (left cochlea) and specifically designed for combining drug delivery with multichannel electrical stimulation. The port of the drug delivery cannula was positioned at the apical tip of the electrode about 360 ° from the round window or about 12–13 mm from the cochlear base, which is ideal because it is the approximate midpoint of the 24–25 mm cat cochlea. This facilitated diffusion of BDNF throughout the cochlea. Electrodes contained six platinum–iridium lead wires terminating in a linear array of six stimulation sites (250–300 μm in diameter) activated as three bipolar pairs (interelectrode spacing of 1 mm). The most apical stimulation site, electrode #1, was positioned near the electrode tip and cannula port. Thus, the deep insertion enabled by the custom design of these implants distributed electrodes along the full circumference of the basal cochlear coil, encircling the auditory nerve, and ensuring electrical activation across a broad range of frequencies. The intracochlear electrode was secured in place with a dacron cuff that was fixed to the inferior aspect of the round window with tissue adhesive (Tissuemend II™, Veterinary Products Laboratories, Phoenix, AZ). The drug delivery cannula within the cochlear implant (0.13 mm/0.005 in. ID) was connected to vinyl tubing (0.69 mm/0.027 in. ID; 1.143 mm /0.008 in. OD) that was in turn attached to the regulator of the osmotic pump, which was implanted behind the right pinna.
Human recombinant BDNF for this study was provided by Amgen, Inc. of Thousand Oaks, CA. Osmotic pumps (Alzet, model #1002 or #2004; infusion rate 0.25 μL/h) were filled and primed as follows: Artificial perilymph (125 mM NaCl, 4.2 mM KCl, 1.3 mM CaCl2, 21 mM NaCHO3, pH 7.4, osmolarity 285–295 mOsm) was prepared from stock solutions. Blood was drawn from each animal, centrifuged, and the supernatant serum was added to the artificial perilymph (final concentration of 200 mg/dL) to provide normal autologous protein. The solution was then passed through a sterile filter and divided into several sterile microtubes that were frozen and stored at −80 °C until use. Osmotic pumps were filled in a UV sterile environment. BDNF (94 μg/mL) was added to the sterile perilymph, and the solution (100 or 200 μL for 14- and 28-day pumps, respectively) was loaded into a pump according to the manufacturer’s directions. Allowing for 20 % loss in the pump and cannula, the 14-day pump delivered about 7.5 μg of BDNF and the 28-day pumps delivered 15 μg, for a total amount of 37.5 μg over the 10-week infusion period. The pump was then primed in a sterile saline bath in an incubation oven (37 °C, 5 % CO2) along with the remaining BDNF/perilymph solution for about 24 h prior to implantation. The extra solution was used during the surgical implantation procedure to pre-fill the vinyl tubing and intracochlear cannula before attaching the osmotic pump. In two animals in the BDNF + ES group, dexamethasone was added to the BDNF solution in the cannula only (1.5 mg/mL for a total dose 45–50 mg/mL over 5 days) in an attempt to reduce inflammation and fibrosis associated with implantation. Because the animals were small at the time of implantation (mean body weight, 560 g; range, 520–610 g), a smaller osmotic pump (model #1002) that delivered 14 days of BDNF was implanted initially. Two weeks later, a brief surgical procedure was performed to replace the initial pump with a larger one containing a 28-day supply of BDNF; this was replaced 1 month later with a final 28-day pump. The pumps were examined upon removal from the animals and appeared to be fully depleted. The electrode components, pump, and cannula appeared intact in all subjects at the time of euthanasia, and there was no evidence of leakage.
Chronic electrical stimulation
Stimulation was delivered on two bipolar channels of the CI and was initiated when animals were 7–10 weeks old, depending on their recovery from surgery and weaning. The apical and basal channels were both activated for 4 h/day, 5 days/week, whereas the middle channel was maintained as a non-stimulated control channel and was activated only for the collection of longitudinal EABR threshold data and during the terminal electrophysiology experiment. In all animals, with the exception of K329, the applied stimulation was delivered by a Clarion™ CII speech processor (loaned by Advanced Bionics, Inc.) using the Bionic Ear Programming System software. Each channel delivered trains of 90 μs/phase pulses at 1,114 pps. Levels of stimulation were controlled by the loudness of ambient environmental sounds, including vocalization by the two to three animals receiving stimulation and sounds from a nearby television playing varied video programming at a comfortable listening volume. The minimum stimulation levels were set at 3 dB below EABR thresholds (0 peak values) and maximum stimulation levels at 6 dB above EABR thresholds, but not exceeding 500 μA. One subject, K329, received concurrent stimulation on the two channels of the implant using computer-generated trains of pulses (200 μsec/phase biphasic pulses) with a carrier rate of 300 pps and sinusoidally amplitude modulated at 30 Hz (100 % modulation). Stimulation intensity for this subject was set at 2 dB above the EABR threshold as determined for the individual stimulated channels. For all animals, the signals delivered on the two CI channels were offset in time such that pulses were interleaved and did not occur simultaneously, modeling current clinical processors. EABR thresholds were assessed every 2 weeks, and stimulation levels on each channel were adjusted accordingly.
In one animal (K319), electrode wire #5 of the basal channel failed (very high impedance) about 1 month after implantation. Stimulation was then delivered on a broader bipolar channel between electrodes #6 and #4; however, 3 weeks before the final electrophysiology experiment, the impedance of this channel also became elevated, and stimulation was delivered only on the apical channel. At the time of the final experiment, connectivity to these electrodes was restored by connecting to the wires in the implanted section of the cable. It should be noted that a sixth subject originally included in this BDNF + ES group (K349) was eliminated from the histological studies because electrode failure after only 1 month resulted in intermittent stimulation on the apical channel. However, at the time of the final experiment, these electrodes were restored by connecting to the wires in the implanted section of the cable and electrophysiological data for K349 are included.
Table 1 also shows deafening and stimulation histories of a comparison group of five neonatally deafened animals that received only chronic ES, no BDNF. These ES-only animals were matched to the BDNF + ES group as closely as possible with regard to the age at study because the duration of deafness has been shown in prior studies to be an important factor in SG survival (Leake et al. 2007). Two of the ES-only subjects received stimulation from the CII speech processor and were stimulated along with the BDNF + ES group. A third subject (K327) was initially included in this ES-only control group, but examination of cochlear histology subsequently showed severe electrode insertion trauma which clearly adversely affected SG survival over several millimeters. Therefore, this animal was eliminated from the histological studies of ES effects on SG survival because these data could have biased our results toward seeing less of an SG survival-promoting effect in the ES-only group as compared to the BDNF + ES group, which was one hypothesis we wanted to test. Three additional animals were selected from an earlier study of ES-only (Leake et al. 2007) to best match the duration of stimulation and deafness of the BDNF + ES group. Two of these animals received computer-generated pulse trains at 300 pps that were sinusoidally amplitude modulated at 30 HZ, and one animal received stimulation at 80 pps (unmodulated).
Electrically evoked auditory brainstem response thresholds
Auditory brainstem response thresholds to electrical pulses (EABR thresholds) were determined bi-weekly or monthly in all animals after implantation of the intracochlear electrode. EABR measurements were made with animals lightly anesthetized with isofluorane. Electrical stimuli were charge-balanced biphasic square wave pulses (200 μs/phase) generated by a real-time processor and attenuated using a programmable attenuator (Tucker Davis RP2.1 and PA5). Pulses were delivered at 20 pps to the selected bipolar pair of CI electrodes via a custom-designed optically isolated and capacitively coupled voltage-to-current converter that was calibrated prior to each recording session. Responses were recorded differentially from silver wires inserted subcutaneously (vertex, active; ipsilateral mastoid, reference; contralateral mastoid, ground), amplified (10,000×) and bandpass-filtered (0.3–10 kHz) using a battery-powered preamplifier (DAM-50), and digitized to 12-bit resolution at 20 kHz (National Instruments PCI-6071E). At each stimulus current level (1-dB increments), responses to 1,000 electrical stimulus pulses were averaged, and thresholds were evaluated visually and confirmed by quantitative online threshold analysis based upon EABR waveform variance (Don and Elberling 1994). Thresholds were determined for each of three bipolar pairs or channels of the implant: the apical channel (electrodes 1, 2) was positioned approximately 12–12.5 mm from the cochlear base, the middle channel (electrodes 3, 4) was centered at about 8 mm and the basal channel at about 5–6 mm (electrodes 5,6). These cochlear locations correspond to roughly 4–4.5, 8–9, and 15 kHz in the normal cat cochlea based upon percentage distance from the base (Liberman 1982).
Electrophysiological recording
At the end of predetermined survival intervals, the animals were studied in acute electrophysiological experiments in which neural responses were recorded from the auditory midbrain or inferior colliculus (IC) in order to evaluate potential functional consequences of BDNF treatment in the central auditory system. These recording methods have been described in detail in previous publications (Rebscher et al. 2007; Snyder et al. 2008). Briefly, under sodium pentobarbital anesthesia, the right IC was exposed, and a 32-channel silicon probe (32 sites, 100-μm spacing on a single shank; NeuroNexus Technologies, Ann Arbor, MI) was inserted along a standardized trajectory parallel to the tonotopic axis of the IC. When the probe reached a suitable depth (i.e., responses to all six intracochlear electrodes could be recorded within the central nucleus of the IC (ICC)), responses of multineuronal clusters and single neurons were recorded. Two to four probe penetrations were made in each subject, recording responses to charge-balanced biphasic current pulses (200 μs/phase), delivered by each of the six individual intracochlear CI electrodes in monopolar mode, and also for all five contiguous bipolar electrode combinations (1–2, 2–3, 3–4, 4–5, 5–6). Minimum response thresholds and response widths were determined by evaluating spatial tuning curves (STCs; Snyder et al. 2008) for each recording probe penetration within the ICC and each CI channel. For a given stimulating channel, the STC describes the strength of response on each recording site (abscissa) for different stimulus levels (ordinate). Strength of response for each recording site was computed by measuring activity within a 20-ms time window (5 to 25 ms following stimulus onset), then subtracting the spontaneous rate (determined by measuring activity in the absence of stimulation) and normalizing by the maximum response evoked among all current levels. A Poisson model was created for each recording site using the spontaneous activity as the rate parameter. For each recording site, the lowest level stimulus that elicited activity significantly greater than the spontaneous rate (p < 0.01) according to the Poisson model was identified as the minimum response threshold, subject to the constraint that all higher stimulation levels also elicited a significant response. For each CI channel, the overall minimum response threshold was identified as the lowest stimulus level that evoked a significant response on any recording site for any probe penetration. The STC width was determined as the distance between the most superficial and deepest sites exhibiting a significant response for stimulation 2 dB above the overall minimum response threshold in the same probe penetration. These STC widths provided an estimate of the extent of excitation across the tonotopic gradient of the ICC (spatial selectivity or tuning) for each channel. The cochleotopic organization of the ICC was inferred by observing the progressive increase in depth of the recording site with the lowest response threshold (i.e., the “best site”) for CI stimulating electrodes located progressively further from the cochlear apex.
It should be noted that the animal groups for the electrophysiological studies differed slightly from those shown in Table 1, which shows the experimental groups for histological studies. As noted previously, one BDNF + ES subject (K349) was eliminated from histological studies due to electrode failure and intermittent stimulation, but this animal was studied in a terminal electrophysiological study, making a total of six in the BDNF + ES group. In the ES-only group, electrophysiological data were obtained for three animals that were stimulated concomitantly with the BDNF + ES group, K352, K335, and K327 (K327 was eliminated from the histological studies due to severe insertion trauma; see “Discussion”). The three other animals in the ES-only cohort (K83, K101, K89) were selected from an earlier study in which electrophysiological experiments were conducted using tungsten microelectrodes, and IC data cannot be compared directly to the probe data presented here. Additional data from recent probe experiments were obtained from three deafened animals implanted with identical intracochlear electrodes to those used in the current experiments. These three animals received 10 weeks of infusion of artificial perilymph (AP) rather than BDNF and received no chronic electrical stimulation; they were described fully in a prior study, in which histological data were reported (Leake et al. 2011). Together the three ES-only and three AP animals comprised a control group of six animals.
Histological preparation of cochlear specimens
Methods for preparation of cochlear specimens were as described previously (Leake et al. 1999, 2007, 2011). Briefly, with the animal deeply anesthetized (sodium pentobarbital, I.V.), both cochleae were perfused directly with mixed aldehyde fixative (1.5 % glutaraldehyde and 2.5 % paraformaldehyde in 0.1 M phosphate buffer, pH 7.4) after removing the stapes and opening the round window. In the cochleae with an implanted electrode, the stapes was removed and perfusion first was attempted through the drug delivery cannula. Subsequently, the electrode was carefully withdrawn from the scala tympani, and the perfusion was performed as usual. If return of fluid was not evident or was limited (e.g., due to fibrosis around the implant), a small opening was drilled in the cochlear apex, and perfusion was carried out between the apex and the oval window. An overdose of sodium pentobarbital was administered, and a transcardiac perfusion was performed with the same fixative solution. The temporal bones were placed in fixative overnight, transferred to phosphate buffer, and the cochlea was isolated and the otic capsule bone was thinned using diamond dental burrs. In implanted cochleae, the bony capsule over the scala vestibuli of the basal turn was opened to facilitate decalcification and infiltration of the basal turn. Next, the specimens were post-fixed in 1 % phosphate-buffered osmium tetroxide with 1.5 % potassium ferricyanide (added to enhance contrast), decalcified in 0.2 M EDTA for 36–48 h, dehydrated and embedded in LX™ epoxy resin. Thick block “surface preparations” containing both the organ of Corti and spiral ganglion were prepared and mounted with epoxy resin on glass slides. The basilar membrane was measured, and small segments (~1 mm) of the organ of Corti along with the adjacent spiral ganglion were excised in each 10 % sector of the cochlea. These specimens were remounted and oriented for sectioning in the radial plane. For morphometric analyses, sections were cut at a thickness of 5 μm, stained with toluidine blue and examined with a Zeiss Axioskop II. At selected locations, additional semi-thin sections were cut at a thickness of 1–2 μm for photomicroscopy to illustrate histological findings. The organ of Corti was evaluated for the presence of hair cells, condition of supporting cells, evidence of chronic infection and/or inflammatory response, and trauma from surgical insertion of the electrode. Morphometric analyses were carried out to assess SG cell survival and cell size, radial nerve fiber density and size, cross-sectional area of blood vessels within Rosenthal’s canal and thickness of the tissue encapsulating the electrode within the scala tympani.
Histological and morphometric analyses
Unbiased stereology to assess spiral ganglion cell numerical density
Survival of the SG neurons was assessed by morphometric analysis of the density of cell somata in Rosenthal's canal as a function of cochlear location, in paired comparisons between the implanted cochlea and the opposite side. Because several previous studies (Agterberg et al. 2008, 2009; Glueckert et al. 2008; Leake et al. 2011; Shepherd et al. 2005, 2008) have reported that BDNF results in increased SG cell soma size, we utilized a physical disector stereological method that accurately estimates the number of objects (nucleoli of SG neurons) in a given volume of tissue, independent of their size (Mouton 2002) as described previously (Leake et al. 2011). Four sets of six serial sections (5 μm thickness) were collected at each of 9 locations representing 10 % sectors of the cochlea. In adjacent serial sections, counts were made to determine the number of “new” nucleoli appearing only in the second section. Images were acquired using a Zeiss Axioskop 2, an Axiocam MRc5 camera, and a 40X/0.95 NA lens. In the native files a 330 by 250 μm field was equal to 2584 by 1936 pixels, and the system acquired images at 150 dpi (resolution, 7 pixels/μm). Pairs of serial sections were imaged at a resolution of 3.2 pixels/μm, 300 dpi, and a screen magnification of ~3000X. In Adobe Photoshop™, the “wand” tool was used to select all cells with a visible nucleolus in the reference section, preserving cytoplasmic boundaries but excluding the nucleus, and other reference structures (e.g., vessels) were also selected. The selected structures were copied, creating a template that was inserted into a file with the image of the second serial section. The two images were then matched and aligned, and the “new” nucleoli in the second section were counted. A minimum of four pairs of serial sections (~20 μm of separation between serial pairs) were analyzed at each location to obtain a data set of at least 50 cells. In addition, the “magnetic lasso” tool was used to outline Rosenthal’s canal. The canal outlines and images of cells were imported into the NIH Image J (version 1.37v) and the canal areas (used to calculate numerical density) and cell areas were determined. Image selection procedures for these and all other morphometric analyses were performed with the observer blinded to experimental conditions.
For overall group comparisons to assess the effects of BDNF and ES on SG cell density, the mean number of nucleoli in a [100 μm]3 volume of Rosenthal’s canal was calculated for each of the nine cochlear sectors (0–90 % from the base).
Measurements of cross-sectional areas of spiral ganglion cells and Rosenthal’s canal
Measurements of SG cell somata were made at the same time and in the same images as the physical disector analysis of SG cell numerical densities described above. The Photoshop images were imported as Tiff files into Image J. The “analyze particles" function was used to estimate cross-sectional areas of all SG cell somata containing a nucleolus, as previously selected for the disector analysis. This analysis measured the perimeter of the SG cell cytoplasmic boundary, excluding the somatic myelin and satellite cells. Approximately 50 cells were measured for each cochlear sector.
At four selected cochlear locations (10–20 %, 30–40 %, 50–60 %, and 70–80 % intervals) separate measurements of the cross-sectional area of Rosenthal’s canal were made in ten serial sections selected randomly from the original 24 serial images used for the numerical density analysis. In Photoshopc™ Tiff images, the boundary of Rosenthal’s canal was traced with the “magnetic lasso” tool, the canal outline images were converted to monochrome, imported into Image J, and canal area was measured using the “particle analysis” function.
SG cell area and numerical density data for normal adult cats were obtained previously using these same methods (Leake et al. 2011) and were used to express data as percentages of normal.
Radial nerve fiber counts and cross-sectional areas
In order to evaluate radial nerve fiber survival and size, separate series of sections (5 μm thickness) were cut transversely through the osseous spiral lamina, orthogonal to the radial plane. Three cochlear regions in the basal turn at about 3–6, 7–9, and 11–13 mm (matched between the left and right sides for each animal) were selected in which the blocks used for cell counts still had remaining tissue. Serial 5 μm sections were cut starting at the habenula and proceeding 135 μm toward the modiolus. Every third section was imaged (7.6 pixels per μm, 150 dpi), providing a total of 9 samples per location. In Photoshop™ the “rectangular marquee” tool (fixed size) was used to count the number of cross-sectional profiles of fibers in a 100 μm length of the osseous spiral lamina. A reference structure was identified in all 9 sections of the series to ensure that counts sampled about the same region. Data were averaged for the nine sections and multiplied by ten to provide an estimate of the number of fibers per millimeter length of the osseous spiral lamina. Fiber size was assessed in these same nine images per cochlear sector, using the “wand” tool to select the axonal profiles of the first ten fibers at one side of each image that had a roughly cross-sectional (round) profile. The selected fiber profiles were imported into Image J and their areas measured using the “wand” tool to outline the outer perimeter of the myelin by demarcating the boundary between the higher pixel density of the myelin as compared to the adjacent tissue. The values were averaged for the nine sections at each cochlear location.
Measurements of vessels in Rosenthal’s canal
To assess the potential angiogenic effects of BDNF (and/or electrical stimulation) within the SG, the cross-sectional areas of vessels within Rosenthal’s canal were measured. At three cochlear locations, about 3–6 mm, 7–9 mm and 11–13 mm from the base, Rosenthal’s canal was imaged (Photoshop) in four sections that were separated from each other by about 25 μm. The “wand” or “lasso” tools were used to select profiles of all clearly identifiable vessels (profiles with clear endothelial lining and/or containing red blood cells), excluding those with diameters smaller than ~10 μm. Tiff files of vessel outlines were imported into Image J and the cross-sectional areas of vessels were measured. The cross-sectional area of Rosenthal's canal was measured in the same sections, and the area occupied by vessels was estimated as a percentage of the overall canal area.
Measurements of tissue encapsulating the cochlear implant
To evaluate the extent of fibrosis around the CI electrode, two cochlear locations were selected where a clear fibrous encapsulation had formed around the electrode in the scala tympani (usually about 6 and 10 mm from the base). Sections were imaged (40× objective), and the thickness of the tissue was determined at two locations in each section in measurements made orthogonal to the basilar membrane and osseous spiral lamina and within 200 μm of the habenula perforata.
Statistical analyses
For all statistical analyses performed, the data were normally distributed, and in all but one instance, the data also showed equal variance among the samples and thus did not violate the assumptions of the tests. The cell size data for the ES-only vs. contralateral comparison failed the equal variance test; therefore, a nonparametric version of two-way ANOVA (Friedman’s test) was used. All statistical analyses were performed using the Sigma Stat™ (version 2.03) or MatLab™ software (Friedman’s test).
Results
Experimental groups
Table 1 shows the individual histories for the experimental animals for which histological data are included in the study. Data are shown for the main experimental group of five neonatally deafened cats that received intracochlear BDNF combined with several months of ES delivered by a cochlear implant. Also shown are data for five additional early-deafened animals that received no BDNF but were studied after similar periods of chronic ES for comparison (termed ES-only group). The number of days of neomycin injections required to induce profound hearing loss ranged from 15 to 21. Age at initial implantation ranged from 31 to 36 days for the BDNF + ES group, and intracochlear infusion of BDNF continued for 10 weeks in all subjects. In the BDNF + ES group, chronic stimulation was initiated at a mean age of 8 weeks, continued for 14 to 26 weeks, and animals were about 28 weeks old at the time of study. In the ES-only group, chronic ES was initiated at ~9 weeks of age, continued for 15–29 weeks, and animals were on average 30 weeks old when studied. SG numerical density values (averaged over the entire cochlea) are shown as percent of normal for individual subjects (see below).
EABR thresholds
Initial auditory brainstem response thresholds elicited by electrical stimulation (EABRs) on bipolar CI channels were determined either at the time or surgery or within 1–2 weeks after surgery. However, in one subject (K313) initial testing was delayed until 3 weeks after surgery due to slow postsurgical recovery. EABR thresholds then were tracked longitudinally at 2- to 4-week intervals over the duration of study in both BDNF + ES (Fig. 1A, B) and ES-only experimental groups (Fig. 1C). The longitudinal EABR threshold data show considerable intersubject variability, especially in initial thresholds obtained near the time of implantation. Careful review of the longitudinal data for individual subjects shows that the threshold for the basal CI channel (electrodes 5, 6, dashed line) tended to be higher than that of the apical channel (1,2). In one subject in the BDNF + ES group (K319), data for the basal CI channel were not available for this analysis due to wire breakage. Statistical comparison of the mean initial and final EABR thresholds in the BDNF + ES animals (Fig. 1D) revealed a significant decrease (i.e., improved sensitivity) for the combined data from the two stimulated channels (Students t test, paired; t(8) = 2.39, p < 0.001). The threshold shift on the apical channel averaged −71 μA, − 2 dB and was similar to the basal channel (mean −77 μA; −2 dB). It is also interesting to note that the single animal (K313) in which EABR testing was delayed until 3 weeks post-op exhibited initial EABR thresholds that were already quite low, and in this animal no subsequent improvement was seen. In contrast to the stimulated CI channels, the middle channel that served as an inactive control channel (except for collection of threshold data) showed no significant change in threshold (shift of +7 μ A 0 dB; t(4) = 0.01; p = 0.99). Further, no significant difference in threshold was observed for the stimulated channels in the ES-only control group (t(10) = −0.67, p = 0.52).
FIG. 1.

Electrically evoked auditory brainstem response (EABR) threshold data. A Longitudinal EABR thresholds are shown for the two stimulated channels of the cochlear implant (solid lines, apical bipolar electrode pair 1,2; dashed lines, basal electrodes 5, 6) recorded in neonatally deafened animals during 10 weeks of intracochlear BDNF infusion and several months of chronic electrical stimulation. B Longitudinal EABR data for the non-stimulated control channel (electrodes 3, 4) in the five BDNF + ES animals. C EABR thresholds for the two stimulated channels in the ES-only animals. D Mean summary data show a significant improvement in threshold (initial vs. final threshold) on the stimulated channels in the BDNF + ES group. In contrast, no significant change over time is seen for the unstimulated middle channel (3,4) of the BDNF + ES group or for the stimulated channels of the ES-only group. (Data shown are means and standard deviations; ***p < 0.001.)
Effects of BDNF + ES on spiral ganglion cell soma size
Evaluation of the organ of Corti confirmed that no intact hair cells were present in any of the deafened animals included in the study. In the group of five animals examined after combined BDNF + ES, the mean cross-sectional areas of the cell somata of SG neurons in the implanted cochleae were consistently larger than cells on the contralateral side in all cochlear sectors (Fig. 2A). An overall trend toward decreasing cell size from base to apex was apparent in both the BDNF + ES and control cochleae, as reported previously in normal cats and in deafened cats examined at shorter durations of deafness (Leake et al. 2011). Averaged for all cochlear regions, the mean cell area following BDNF delivery and ES was about 235 μm2, as compared to about 200 μm2 for cells in the contralateral cochlea, and this difference was highly statistically significant (two-way ANOVA: F(1,72) = 35.75, p < 0.001). When compared to data for normal adult cats, mean cell areas on both sides were significantly smaller than normal, with a mean of 88 % of normal for BDNF + ES and 75 % for deafened contralateral cochleae.
FIG. 2.

Cross-sectional areas of the somata of spiral ganglion (SG) neurons. A Data are shown in the BDNF + ES group for nine cochlear sectors representing 10 % intervals from base to apex. Cell areas throughout the cochlea are significantly larger in the BDNF + ES cochleae as compared to the contralateral deafened cochleae. After BDNF + ES, the mean SG cell area is about 235 μm2, which is significantly smaller than normal but also significantly larger than cells contralaterally, which on average measured about 200 μm2. B Data for neonatally deafened animals that received chronic ES only (no BDNF) and were selected to match the BDNF + ES group for duration of deafness and duration and type of applied ES. Again, cell areas are larger in the implanted and stimulated cochleae than contralateral, and values are quite similar to the BDNF + ES group. (Data presented are means and standard errors.)
In the comparison group of neonatally deafened cats examined after comparable periods of ES-only with no BDNF infusion, SG cell somata were also consistently larger on the stimulated side as shown in Figure 2B (Friedman’s test: χ2 (1,N = 45) = 13.97, p < 0.001). Cell areas averaged 85 % of normal in the stimulated ears and 75 % on the contralateral side, similar to values in the BDNF + ES group. Moreover, statistical comparison of cell size in the BDNF + ES cochleae showed no significant difference from the ES-only ears (two-way ANOVA: F(1,72) = 1.70, p = 0.197).
Effects of BDNF on survival of spiral ganglion neurons
The physical disector method (counts of the nucleoli of SG neurons in adjacent serial sections) was applied to estimate the numerical density of SG neurons in Rosenthal’s canal for nine cochlear sectors from the base to the apex in all the animals included in this study. Figure 3A presents data for the five animals examined after combined BDNF + ES. The numerical density of SG cells in the BDNF + ES cochleae was consistently higher than that on the other side in all cochlear sectors, with the largest differences seen in middle cochlear sectors where cell loss was greatest contralaterally. Averaged for the entire cochlea and expressed as percentage of normal, the mean cell density after BDNF + ES was about 70 % of normal, compared to about 45 % in the contralateral deafened cochleae, and this difference was highly significant (two-way ANOVA: F(1,72) = 45.30, p < 0.001). When calculated as percentage increase, normalized to survival on the contralateral side, the BDNF + ES cochleae showed a 56 % increase in SG survival.
FIG. 3.

A Numerical densities of SG neurons. Data are shown for 10 % cochlear sectors from base to apex, with data expressed as percentage of normal for the same BDNF + ES group shown as in Figure 3A. The BDNF + ES cochleae show higher densities for all cochlear intervals than the paired data for the contralateral ears, although significant neural degeneration occurred on both sides. Averaged over all cochlear sectors, SG cell survival is substantially improved at about 70 % of normal after BDNF + ES, as compared to about 45 % on the opposite side. B Data from the ES-only group also show a significantly higher numerical density in the implanted and stimulated cochleae, compared to contralateral deafened ears. C To directly compare the effects of treatment in the two groups, the difference in numerical density (increase re: contralateral deafened) is shown for both the BDNF + ES group and the ES-only group. (Data presented are means and standard errors.)
Figure 3B presents comparable data for the ES-only group. As in the combined BDNF + ES data, the numerical density of SG cells in the implanted ears was consistently higher than that on the opposite side, with the single exception of the 30–40 % sector (where slight electrode insertion trauma often occurred; see below). Overall, the mean SG cell density in the ES-only cochleae was about 63 % of normal, compared to 47 % on the contralateral side, and this difference was highly significant (two-way ANOVA: F(1,72) = 15.68, p < 0.001). When normalized to the survival in the deafened control ears, this represents an increase of more than 30 % in SG survival in the ES-only cochleae. Direct comparison of SG density in the implanted cochleae from BDNF + ES and ES-only groups showed no significant difference. However, when we calculated the differences in SG density (treated minus control), the 25 % of normal increase in SG survival resulting from combined BDNF + ES was significantly greater that the difference of 16 % seen in the ES-only group (two-way ANOVA: F(1,72) = 4.56, p = 0.036; Fig. 3C).
A spurious factor that could potentially affect SG neuronal density is a systematic difference in the size of Rosenthal’s canal that might occur between sides or across groups. Figure 4A shows the data obtained from direct measurements of the cross-sectional area of Rosenthal’s canal in a subset of four cochlear intervals in the BDNF + ES cochleae and matched data from the contralateral cochleae. The canal area in the BDNF + ES-treated side was equivalent to that of the normal adult cochlea (average, 97 % of normal), and the canal in the contralateral untreated ears was slightly smaller (90 % of normal), but this difference was not statistically significant (two-way ANOVA: F(1,32) = 3.41, p = 0.074). Note that the trend toward larger canal size in the treated cochleae would tend to bias our data toward a lower cell density in the BDNF + ES data compared to the contralateral side and thus only increases confidence in the finding of increased neuronal survival in the treated ears. Figure 4B shows measurements of Rosenthal’s canal in the ES-only group, in which the canal was 100 % of normal for the stimulated ears compared to 93 % of normal on the control side, and again this small difference between sides was not statistically significant (two-way ANOVA: F(1,32) = 3.63, p = 0.066).
FIG. 4.

A The mean cross-sectional area of Rosenthal’s canal in neonatally deafened cats after combined BDNF + ES, sampled in four cochlear sectors from the base to the apex. Canal area in the BDNF + ES cochleae averaged 97 % of normal and was slightly larger than on the contralateral side (90 % normal), but this difference was not statistically significant. B Rosenthal’s canal area data for the same four cochlear sectors in the ES-only group. Canal area averaged exactly 100 % of normal in the implanted, stimulated cochleae, and was slightly but not significantly larger than that in the contralateral deafened cochleae (93 % of normal).
The histological sections in Figure 5 illustrate the effects of combined BDNF + ES. In light microscopic images taken from a basal cochlear sector (20–30 % from the base), the higher density of SG cell somata within Rosenthal’s canal is evident in the BDNF + ES treated ear (Fig. 5A, C) as compared to the same region in the contralateral ear (Fig. 5B, D). These images also illustrate the markedly higher density of radial nerve fibers within the osseous spiral lamina noted in the implanted cochlea as compared to the opposite side (arrows). In the higher magnification images, the larger cross-sectional areas of SG cell somata in the BDNF + ES cochleae (Fig. 5C) also can be appreciated, compared to cells on the contralateral side (Fig. 5D).
FIG. 5.

Light microscopic images of histological sections of the cochleae from a neonatally deafened animal (K313) illustrating the marked neurotrophic effects observed following combined BDNF + ES. The 20–30 % sector of the implanted left BDNF + ES cochlea (A, C) and the paired region from the contralateral deafened control cochleae (B, D) illustrate the higher density of SG cells in Rosenthal’s canal and the maintenance of a greater number of radial nerve fibers within the osseous spiral lamina (filled arrows in A, B) after BDNF infusion. In the implanted ear, the scala tympani aspect of the osseous spiral lamina (just below the radial nerve fibers) is thickened by ectopic neo-osteoneogenesis, and below this, the fibrous tissue that encapsulated the CI electrode is evident in the scala tympani (as measured for the data presented in Figure 9B). The higher magnification images in C and D illustrate the quality of preservation, staining of the tissue after osmium post-fixation and the resolution of images used for morphometric analyses in the study. The higher density and size of SG perikarya in the BDNF-treated cochlea as compared to neurons on the opposite side are evident. Open arrows in panel C indicate enlarged blood vessels after BDNF + ES as measured for data in Figure 9A. Scale bars = 50 μm.
It is of interest to note in Figure 5A that although no intact hair cells were present in the degenerated organ of Corti throughout the cochleae in these early-deafened animals, some remnants of supporting cells usually were present (see also Fig. 8A). The condition of the organ of Corti ranged from complete resorption and formation of a single layer of flattened cells (Fig. 5B), to full maintenance of the cytoarchitecture of the organ of Corti with well-formed phalangeal scars where hair cells had degenerated. As reported previously (Leake et al. 2007), the degeneration of supporting cells tended to be most severe in the cochlear base and to improve progressively in more apical regions (see “Discussion”).
FIG. 8.

A Light microscopic image of the organ of Corti (20–30 % cochlear sector) in a neonatally deafened animal (K319) after combined BDNF + ES, illustrating the ectopic sprouting of radial nerve fibers exiting from the osseous spiral lamina into the scala tympani (arrows) and forming small bundles (arrowheads) that take a spiral course within the loose fibrotic tissue matrix above the cochlear implant electrode. B Transmission electron micrograph image of these sprouted peripheral fibers in the scala tympani of this same cochlea illustrates several large, well-myelinated axonal profiles (m) as well as numerous unmyelinated fibers (u). Scale bar in A = 50 μm; scale bar in B = 0.5 μm.
Radial nerve fiber number, size and distribution
The dendrites of the SG neurons (radial nerve fibers) were assessed in tangential cross-sections of the osseous spiral lamina (orthogonal to the radial plane) cut at three cochlear locations within the basal half of the cochlear spiral as described previously (Leake et al. 2011). The number of cross-sectional profiles of radial nerve fibers was consistently higher in the BDNF + ES cochleae in all three cochlear sectors examined, as compared to the contralateral deafened ears (Fig. 6A). The highest density was observed in the most basal cochlear sector, where nearly 60 % of the normal fiber population was maintained with BDNF + ES; values were about 30 % of normal in the middle and apical sectors. Substantially lower fiber counts (~20 % decrease) were observed in the contralateral ears in the basal cochlear sector (20–30 %) adjacent to the basal stimulating CI channel (electrodes 5, 6), and also at the apical location (50–60 %) adjacent to the apical stimulating channel of the CI (electrodes 1, 2). A more modest increase in the implanted ears (less than 10 %) was observed in the middle cochlear sector (30–40 %) near the non-stimulated CI channel (electrodes 3, 4). Given the consistent difference in all three cochlear regions, overall fiber counts were significantly higher in the combined BDNF + ES cochleae compared to contralateral (two-way ANOVA: F(1,24) = 8.64, p = 0.007). In contrast, fiber counts from the same three cochlear regions in the ES only group showed a left–right difference only in the most basal sector, and overall there was no statistically significant difference between sides (two-way ANOVA: F(1,24) = 1.19; p = 0.286, Fig. 6B). Further, direct statistical comparison indicated that fiber numbers in BDNF + ES cochleae were significantly higher than the ES-only ears (two-way ANOVA: F(1,24) = 6.11, p = 0.021).
FIG. 6.

Light microscopic images of histological section of the cochleae from a neonatally deafened animal (K313) illustrating the marked neurotrophic effects observed following combined BDNF + ES. The 20–30 % sector of the implanted left BDNF + ES cochlea (A, C) and the paired region from the contralateral deafened control cochleae (B, D) illustrate the higher density of SG cells in Rosenthal’s canal and the maintenance of a greater number of radial nerve fibers within the osseous spiral lamina (filled arrows in A, B) after BDNF infusion. In the implanted ear, the scala tympani aspect of the osseous spiral lamina (just below the radial nerve fibers) is thickened by ectopic neo-osteoneogenesis, and below this, the fibrous tissue that encapsulated the CI electrode is evident in the scala tympani (as measured for the data presented in Figure 10B). The higher magnification images in C and D illustrate the quality of preservation, staining of the tissue after osmium post-fixation and the resolution of images used for morphometric analyses in the study. The higher density and size of SG perikarya in the BDNF-treated cochlea as compared to neurons on the opposite side are evident. Open arrows in panel C indicate enlarged blood vessels after BDNF + ES as measured for data in Figure 10A. Scale bars = 50 μ0.
Figure 6C shows the calculated ratio of the fiber counts (in percent of normal) to SG cell numerical density (percent of normal) obtained from the same cochlear sectors. This ratio provides a rough estimate of the relative percentage of the surviving SG cells that have maintained their radial nerve fibers. After BDNF + ES, the basal sector had about 80 % of the surviving SG cells apparently associated with fibers, approximately 50 % in the middle region and 75 % in the more apical sector. Calculating these same ratios for animals studied after ES alone shows lower values in all three cochlear sectors (65 %, 30 %, and 33 % of normal, respectively). Two-way analysis of variance gave a main effect for treatment, F(1,24) = 6.97, p = 0.014, indicating that the cell-to-fiber ratio was significantly higher for the BDNF + ES cochleae than for the ES-only ears.
In addition to the greater number of radial nerve fibers observed, measurements of their cross-sectional areas (outer perimeter of myelin) revealed that fibers were substantially larger in the BDNF + ES cochleae than on the contralateral side (Fig. 7A, B, C; one-way ANOVA: F(1,28) = 12.35, p = 0.002). In fact, fibers averaged about 140 % of normal after BDNF + ES, whereas those in the contralateral deafened ears were not significantly different from normal. In the ES-only group, fiber areas were also larger on the implanted side but the difference (106 % vs. 93 % of normal) was much less than in the BDNF + ES group (Fig. 7D; Student’s paired t test, t(4) = 3.42; p = 0.027). It should be noted that because the fiber counts and area measurements were made at the light microscopic level, no attempt was made to assess potential pathological alterations in the myelin or the axoplasm of these fibers. Therefore, some fibers that were undergoing initial stages of degeneration may have been included in both fiber counts and area data.
FIG. 7.

The cross-sectional areas of the radial nerve fibers were also evaluated at the same three cochlear regions where fiber counts were performed. These measurements were made in higher magnification images, as illustrated here in sections taken from the implanted cochlea (A) and contralateral deafened cochlea (B) in one combined BDNF + ES subject (K318). No systematic difference across regions was observed, and the data were averaged for all regions. C Fibers in the BDNF + ES cochleae are significantly larger than fibers in the contralateral cochleae and also markedly larger than normal. D Fiber areas in cochleae examined after chronic ES-only also are larger than fibers in the contralateral deafened cochleae, but not larger than normal.
Another notable finding after BDNF + ES was that myelinated radial nerve fibers had sprouted through the lower bony plate of the osseous spiral lamina and had grown into the scala tympani where they were observed under the osseous spiral lamina and basilar membrane and within the fibrous tissue encapsulating the cochlear implant electrode (Fig. 8A). Such ectopic sprouted neuronal processes were observed in the BDNF + ES cochleae of all five animals examined but were not observed in deafened control ears or in the ES only group. These fibers were largely limited to the cochlear sector adjacent to the cochlear implant electrode, and within this region, the number and distribution of fibers were quite variable among the individual cochleae. These sprouting neurons passed through the osseous spiral lamina to enter the scala tympani, but such fibers were seldom, if ever, seen in regions of insertion trauma, even when the osseous spiral lamina was disrupted by fracture. Because SG survival often was clearly compromised in these limited damaged areas (as compared to adjacent regions), few radial nerve fibers were present, and sprouted fibers were not observed. In two animals K313, K318, sprouted fibers in the implanted ears were seen in locations apical to the implant, as far as 16 and 18 mm from the base, respectively. It is interesting to note that the Schwann cells associated with these sprouted fibers often appeared hypertrophied and/or more numerous than usual as illustrated in Figure 7A and also apparent in Figure 8A and B. Note particularly in Figure 8A, the large and numerous Schwann cells associated with fibers within and exiting the osseous spiral lamina and in the area under the basilar membrane (arrowheads). Transmission electron microscopy of these fibers in three of the BDNF + ES cochleae demonstrated the presence of both large myelinated and smaller unmyelinated profiles (Fig. 8B) that sometimes aggregated into small fascicles surrounded by fibrotic tissue. The TEM image in Figure 8B also illustrates the very large and prominent Schwann cells associated with these sprouting neurons.
Angiogenesis in Rosenthal’s canal and fibrotic encapsulation of CI
Figure 9A presents data on the cross-sectional area of vessels within Rosenthal’s canal after combined BDNF + ES. Vessel area measured in the same cochlear sectors as the fiber density/area measurements (shown as percent normal) was markedly greater in the BDNF + ES animals than in the contralateral deafened cochleae (one-way ANOVA, F(2,57 = 25.07, p < 0.001) and was almost triple the normal vessel area (see Fig. 5A, C for histology). Interestingly, data from animals studied immediately after BDNF delivery at ~14 weeks of age (Leake et al. 2011) showed findings very similar to the present data from the BDNF + ES group, suggesting a relatively long-term effect. In contrast, after ES alone and in the contralateral deaf ears, vessel area was not significantly different from normal. Examination of histological sections in regions adjacent to the CI also suggested increased peri-implant fibrosis in the BDNF-treated ears. Measurements of the thickness of the fibrotic tissue layer encapsulating the electrode array (Fig. 9B) revealed thicker fibrosis in the BDNF + ES group (190 μm) as compared to ES alone subjects (120 μm; [Students t -test, t(15) = −2.81, p = 0.013]), in a group that included the animals in the current study and several additional ES-only subjects from prior studies (Leake et al. 1999, 2007). An example of this peri-implant fibrous tissue is illustrated in Figures 5A and 8A shows a contrasting example where only very loose areolar tissue formed around the implanted electrode.
FIG. 9.

A The cross-sectional area of the vessels within Rosenthal’s canal, normalized to the canal area are shown as percent normal. The area occupied by vessels is markedly larger than normal in the BDNF + ES group and also substantially larger than in the contralateral deafened cochleae and in animals receiving equivalent periods of ES without BDNF. B Measurements of the thickness of the fibrotic tissue encapsulating the cochlear implant revealed thicker fibrosis in the BDNF + ES group as compared to the ES-only group.
Functional effects of BDNF and/or electrical stimulation
Figure 10 presents data from the terminal electrophysiology experiments using 32-channel probes to record from the central nucleus of the IC (ICC) in these animals. Minimum thresholds for electrical activation of bipolar CI channels are plotted as a function of the EABR thresholds for the same channels. The ICC thresholds were usually lower than EABR thresholds, and the two values showed a strong correlation accounting for roughly 70 % of the variance in the data (linear regression, R 2 = 0.710, F(1,31) = 73.46, p < 0.001). No obvious trends were noted in the data for the different deafened groups represented, although all but one of the data points falling above the equal intensity line (i.e., indicating that ICC threshold was higher than the corresponding EABR threshold) were from chronically stimulated cats.
FIG. 10.

In terminal electrophysiology experiments, 32-channel probes were used to record neuronal responses from the ICC. Minimum thresholds for electrical activation of individual bipolar CI channels are plotted as a function of the EABR thresholds for the same channels, and the two values show a strong correlation.
To test for a relationship between threshold and selectivity of stimulation, the IC threshold data for bipolar CI channels were plotted as a function of activation widths measured at 2 dB above minimum threshold for the same bipolar channel of the CI (Fig. 11). In control subjects that received ES alone or intracochlear infusion of artificial perilymph (no BDNF or chronic ES), lower ICC thresholds were significantly correlated with smaller, more selective ICC activation widths as expected (Fig. 11A; linear regression: R 2 = 0.35, F(1,24) = 12.67, p = 0.0016). This relationship accounts for roughly one third of the variance in the data for the control group. In contrast, in animals studied immediately after combined BDNF + ES, no such correlation was observed (Fig. 11B; linear regression: R 2 = 0.033, F(1,22) = 0.76, p = 0.39). Careful inspection of the data in these figures suggests that both activation widths and thresholds were more variable with BDNF treatment. For example, for very low IC thresholds of less than 25 dB STCs in different subjects varied from a highly selective 2-dB activation width of 0.2 mm to extremely broad activation of >2.5 mm. This failure to observe the expected relationship of minimum threshold to selectivity of activation may relate to the observed BDNF effects of ectopic, disorganized sprouting in these ears (see “Discussion”).
FIG. 11.
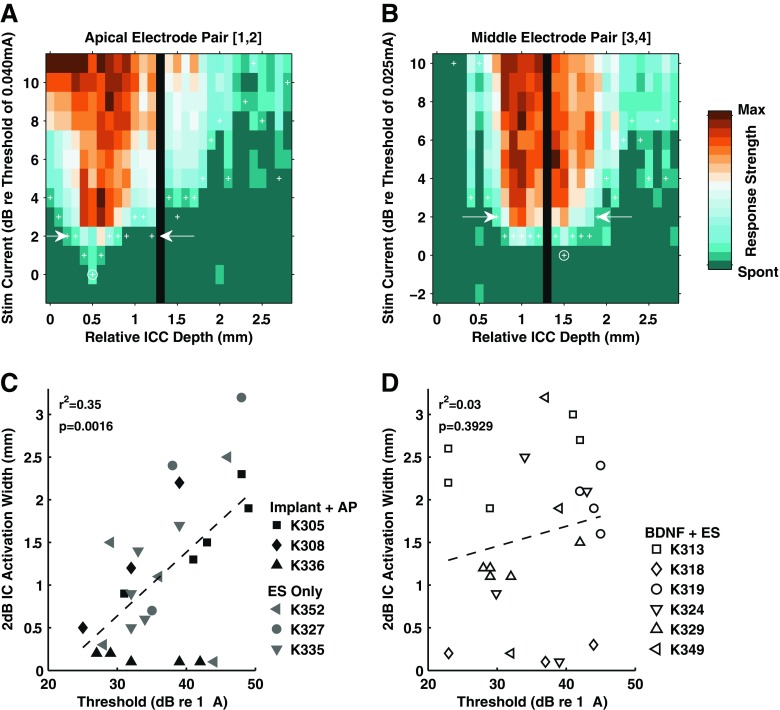
In terminal electrophysiology experiments, 32-channel probes were used to record neuronal responses from the central nucleus of the ICC. Minimum thresholds for electrical activation of individual bipolar CI channels are plotted as a function of the EABR thresholds for the same channels, and the two values show a strong correlation.
Figure 12 presents additional data from these IC probe recordings. Each of the first five panels illustrates data from an individual cat implanted with a six-wire CI electrode and studied after combined BDNF + ES. Each line represents a single probe placement, plotting best depths for monopolar stimulation of electrodes 1 (most apical) through 6 (most basal). Although bipolar channels were used for chronic stimulation, examination of best depths for individual monopolar electrodes was used for this study to provide a higher resolution evaluation of the ICC cochleotopic organization. The normal cochleotopic gradient is apparent in a monotonic relationship between the site of cochlear stimulation and ICC depth for all penetrations. That is, threshold level responses at superficial ICC sites are evoked by stimulation of low frequency (apical) electrodes, whereas threshold level responses at deeper ICC sites are evoked by stimulation of high frequency (basal) electrodes, as appropriate for the well-known tonotopic organization of the ICC.
FIG. 12.

Data from probe recordings in the ICC in individual animals in the combined BDNF + ES group are shown in panels A–E. Each line represents a single probe placement and plots best-depths (lowest threshold location) for monopolar stimulation of the cochlear implant electrodes #1 (most apical) through #6 (most basal). The normal cochleotopic gradient is apparent in a monotonic relationship between the site of cochlear stimulation and ICC depth for all penetrations. Lower frequency (apical) electrodes elicit responses more superficially and higher frequency (basal) best-depths are found deeper, as appropriate for the well-known tonotopic organization of the ICC. (Data are normalized with best-depth for electrode 1 = 0.) For reference, similar plots for three untreated deaf control subjects are shown in panel F.
Discussion
Longitudinal EABR threshold data
Previous studies in adult-deafened guinea pigs have reported reduction in EABR thresholds (Shepherd et al. 2005; Shinohara et al. 2002) and larger amplitude of the suprathreshold EABR responses (Agterberg et al. 2009) after 4 weeks of intracochlear infusion of BDNF. Improved EABR thresholds have also been reported after intracochlear overexpression of BDNF by adenovirus (Chikar et al. 2008) and intracochlear infusion of CTNF (Yamagata et al. 2004) or GDNF (Maruyama et al. 2008; Scheper et al. 2009). Our previous study demonstrated a marked 6-dB improvement in EABR thresholds over 10 weeks of BDNF infusion in neonatally deafened, developing cats (Leake et al. 2011). In the present study, a longitudinal reduction in EABR thresholds also was observed, but only for the two chronically stimulated bipolar channels of the CI, and not for a middle non-stimulated control channel. The specific mechanism(s) underlying these changes in threshold are unclear. It is possible that the larger size and number of the radial nerve fibers or the ectopic sprouting into the scala tympani above the stimulating CI electrodes could provide an improved electrode-to-neuron coupling resulting in improved thresholds. However, none of these findings showed specific regional differences in the three basal cochlear sectors examined (corresponding to the locations of the three CI channels) that might account for the observed differences across channels. Alteration in the SG neuronal membrane properties or channels also could account for the observed EABR threshold reductions. Alternatively, it should also be noted that previous animal studies have shown that lower EABR thresholds are correlated with better SG survival in the cochlear region near the stimulating electrodes (Leake et al. 2008b). Thus, it may be possible that some neurons initially unresponsive to electrical stimulation, due to ototoxic drug administration effects, become viable and responsive during BDNF treatment. From the standpoint of clinical application of CIs, lower thresholds to electrical stimulation may be advantageous, potentially resulting in larger dynamic ranges for electrical stimulation, reduced power consumption and improved battery life for a speech processor. Importantly, a recent detailed study of a large cohort of CI recipients demonstrated that lower CI sound-field threshold levels (frequency modulated tones) were significantly related to better CI performance (Holden et al. 2013).
Effects of deafness, BDNF, and ES on SG cell soma size
Profound deafness has been reported to result in reduction in the size of SG neuron perikarya in many previous studies and in various deaf animal models (Agterberg et al. 2008, 2009; Araki et al. 1998; Elverland and Mair 1980; Glueckert et al. 2008; Leake et al. 1999, 2007, 2011; Leake and Hradek 1988; McGuinness and Shepherd 2005; Shepherd et al. 2005; Wise et al. 2005). Presumably, reduction in somatic size reflects reduced metabolic requirements of these neurons due to reduced spike activity after deafness (Hartmann et al. 1984; Liberman and Kiang 1978; Shepherd and Javel 1997). When electrical stimulation from a CI is applied to reactivate these neurons, modest but significant increases in SG soma area in the stimulated cochleae have been reported (Araki et al. 1998; Coco et al. 2007; Leake et al. 1999, 2007). However, the effects on soma area associated with neurotrophin administration are much greater than the cell size changes elicited with electrical stimulation. In fact, several previous studies in guinea pigs have shown that 4 weeks of intracochlear infusion of BDNF resulted in large increases in SG perikaryal size compared to deafened cochleae and cross-sectional areas that were even significantly larger than in normal controls (Agterberg et al. 2008; Glueckert et al. 2008; McGuinness and Shepherd 2005; Shepherd et al. 2005, 2008). The mechanism(s) underlying this alteration are unclear, but we agree with the suggestion by (Shepherd et al. 2008) that it may be related to the high concentrations of exogenous neurotrophin applied in these studies using osmotic pumps.
Consistent with these previous studies, our data indicate that BDNF + ES promoted growth and maintenance of a significantly larger SG cell soma size as compared to the contralateral deafened cochleae in these developing animals (p < 0.001). However, the difference was relatively modest, with cross-sectional areas of cells in the BDNF + ES cochleae averaging about 88 % of normal adult size, compared to about 75 % of normal for cells on the contralateral side. Romand (Romand and Romand 1990; Romand et al. 1980) previously reported that SG cell somata in normal cats are full adult size and well-myelinated by 1 month of age, the time at which BDNF treatment was initiated in the deafened kittens in our study. In contrast, data from our previous study showed that cells in neonatally deafened animals at about 1 month of age were significantly smaller than normal, averaging only 71 % of normal (Leake et al. 2011). The present data indicate that after 10 weeks of BDNF combined with 5 months of ES, SG cells were significantly larger than those in 4-week-old deafened animals, but also smaller than normal. It is interesting to note that data from a previous study showed that immediately after 10 weeks of BDNF infusion, cells had normal mean areas in the treated ears, compared to 80 % of normal for cells on the contralateral side. The current data indicate that a significant decrease in mean cell area from normal to 88 % of normal occurred after cessation of BDNF, despite ongoing electrical stimulation. A smaller, but still significant decrement from 80 % to 75 % of normal was seen in the contralateral untreated ears, presumably due to the longer duration of deafness in the current study.
Together, findings indicate that BDNF alone elicits a substantial effect on neuronal somatic size, but this effect is not maintained over the long-term with ongoing CI stimulation. Consistent with this interpretation are the similar findings in the ES-only group, in which cell areas in the stimulated cochleae averaged 85 % of normal as compared to 75 % contralateral.
Combined BDNF and ES promote survival of spiral ganglion neurons
A highly significant neurotrophic effect was observed with intracochlear infusion of BDNF and 5 months of ES. Combined BDNF + ES resulted in a mean SG numerical density of about 70 % of normal as compared to 45 % on the contralateral side (p < 0.001), and systematically higher SG survival was maintained throughout the cochlea (Fig. 4). The greatest difference in neural density appeared to be in the apical half of the cochlea, in the region ~50–90 % from the base, largely because this is the area where progressive neuronal loss was greatest on the contralateral side. As reported previously, the trend for greater SG neuronal loss to occur in the middle cochlear sectors seems to be unique to animals deafened as neonates and is interesting because the hair cell degeneration in these animals progresses from base to apex (Leake et al. 1997), which is the typical pattern of aminoglycoside ototoxicity in adult cats (Leake and Hradek 1988). The initial SG neural degeneration in these young animals apparently is not simply secondary to sensory cell degeneration, and neurons in the middle cochlear sectors appear to be more sensitive to direct ototoxic damage than in other regions, presumably due to some regionally varying aspect of cochlear maturation or onset of function.
In our previous study, neonatally deafened cats were examined immediately after 10 weeks infusion of BDNF, and SG density was 80 % of normal, equivalent to values seen in control deafened subjects studied at about 4 weeks of age, when BDNF treatment was initiated (Leake et al. 2011). Thus, over the 10-week period of infusion, BDNF maintained virtually all the SG neurons that were present initially. In the current study, when we terminated BDNF infusion and continued ES for several months, SG survival was not maintained quite at this same level but was reduced by roughly 10 %, to 70 % of normal. One factor that may have contributed to this loss was the fact that the present study used longer electrodes that extended to the 50–60 % cochlear region, whereas the earlier study used mainly shorter electrodes that reached only the 30 % cochlear region. The longer electrodes often cause slight insertion trauma in the middle cochlear sectors that may have reduced survival there. In point of fact, comparison of initial BDNF SG data with the BDNF + ES data show that most of the difference between the two groups occurred in these middle cochlear sectors. Alternatively, although this 10 % reduction was statistically significant, with the relatively small number of subjects in our experimental groups, we cannot rule out the possibility that this small difference is due to individual variability in ototoxic drug effects across groups.
If the overall SG numerical density seen after BDNF + ES (mean ~70 % of normal) is normalized to the density in the contralateral deafened cochleae (45 %), the resulting percentage increase in neuronal survival is more than 55 %. In contrast, our previous study of deafened animals studied immediately after 10 weeks of BDNF infusion showed only a 25 % mean increase in SG numerical density, relative to contralateral with SG survival of 80 % of normal vs. 65 % (Leake et al. 2011). The greater effect in the present study indicates that combined BDNF + ES was highly effective in promoting neuronal survival, while at the same time much more severe neural degeneration occurred in the non-implanted ears over the longer duration of deafness. Together, the findings suggest that the neurotrophic effects of exogenous neurotrophins can be maintained over a prolonged period and that these “rescued” neurons are viable to support ongoing electrical stimulation delivered by a CI. This is important because our prior study of SG neurononal survival immediately after 10 weeks of BDNF also showed that many neurons in the BDNF-treated ears do not have normal somatic myelin (Leake et al. 2011). Thus, it is noteworthy in the present study that most of these neurons survive over the long-term, even when subjected to the ongoing stress of chronic ES. Moreover, a direct comparison of the neurotophic effects of combined BDNF + ES to data from well-matched subjects that receive only ES demonstrated that the combined treatment elicited a significantly greater effect in promoting survival of the primary afferent SG neurons. BDNF + ES maintained an additional 25 % of the normal SG neuronal population compared to contralateral, whereas ES-only improved survival by 16 % of normal (Fig. 4C). On the other hand, our study does not allow us to define the relative contributions of BDNF vs. ES to the overall effects observed. In future work, it would be of interest to examine the potential long-term effects of this same BDNF treatment after a similar duration of deafness, but without ongoing electrical activation. Although our results suggest that BDNF + ES promotes improved SG survival over ES alone, it remains to be determined whether BDNF treatment effects may persist and promote long-term neuronal survival, without subsequent chronic ES.
Another factor that could influence accurate assessment of neuronal survival is a systematic difference in the size of Rosenthal’s canal between sides or among experimental groups. A reduction in the cross-sectional area of Rosenthal’s canal was reported previously in guinea pigs after electrical stimulation delivered by a cochlear implant (Li et al. 1999). This issue is potentially important in the present study because of the young age at which the animals were implanted and possible alterations elicited by BDNF during a period when growth is still occurring. In the present study, however, Rosenthal’s canal was 99 % of normal adult size, both in the BDNF + ES cochleae and in the ES-only ears, and the smaller canal area observed in the contralateral cochleae (i.e., about 90 % of normal, both groups) actually would have biased results toward a higher SG density in the contralateral ears and less difference between the two sides. It is interesting that the small yet significant differences were so similar in both BDNF + ES and ES-only experimental groups, leading the authors to speculate that perhaps the larger population of surviving SG neurons during postnatal development tends to maintain a more normal canal size compared to the more severely degenerated contralateral ears.
Another interesting aspect of the morphological findings in cochleae from these deafened animals was the condition of the supporting cells in the organ of Corti after the hair cells had degenerated. As shown previously in cats deafened neonatally by administration of neomycin, the supporting cells were most severely degenerated in the cochlear base, where it was often difficult to recognize pillar cells (see Fig. 5A, B; Fig. 8A). Better maintenance of supporting cell cytoarchitecture was seen at progressively more apical regions. Previously, it has been reported that long-term neuronal survival is enhanced when supporting cells in the inner hair cell area remain intact (Sugarawara et al. 2005), and recently it has been suggested that the supporting cells may be the main source of SG survival factors (Zilberstein et al. 2012). In the present study, however, it is unclear what role, if any, the supporting cells play in the observed effects of BDNF + ES and ES-only in these developing animals. The pattern of supporting cell degeneration follows the basal-to-apical progression that is typical of aminoglycoside ototoxicity as reported previously (Leake et al. 1997, 2007). In contrast, the degeneration of SG neurons is most severe in the middle cochlear sectors and the survival-promoting effects of treatment are seen throughout the cochlea, both in the BDNF + ES and the ES-only ears. A previous study of neurotrophic effects of GM1 ganglioside and ES in neomycin-deafened cats (Leake et al. 2007) showed a rank–order correlation of supporting cell condition to the overall SG survival in control, deafened cochleae. In contrast, no such correlation occurred in the implanted/stimulated ears, where ES elicited a significant SG protective effect that appeared to be independent of supporting cell condition.
Radial nerve fiber size, distribution and sprouting
Our data on radial nerve fiber density within the osseous spiral lamina demonstrated significantly greater numbers of fibers throughout the basal cochlear turn following BDNF + ES as compared to both the contralateral deaf ears and ES-only cochleae. The highest fiber density in both groups was observed in the most basal cochlear sector, about 3–5 mm from the base. In the other two regions examined (about 7–9 and 11–13 mm), survival decreased markedly, likely due to the more pronounced SG loss in those middle cochlear sectors, as mentioned previously. In the BDNF + ES group, if we view the data in terms of the difference between treated and control, the basal and apical regions both showed a substantial increase in fiber survival (~20 %) over contralateral, but the middle sector showed a more modest difference (7 %). The areas examined were selected to be adjacent to the three bipolar channels of the CI, and it is tempting to speculate that the more substantial differences in the basal and apical cochlear sectors were trophic effects of the chronic electrical stimulation delivered on the adjacent CI channels (electrodes 1, 2 and 5, 6, respectively), whereas the middle channel remained inactive as a control (except for collection of EABR data). In the ES-only group, there was no significant difference between sides.
The finding of an increase in the number of surviving radial nerve fibers after BDNF treatment is consistent with the results of Wise et al. (2005) who reported increased fiber survival after infusion of BDNF and NT-3 in adult guinea pigs and also the findings reported by Glueckert et al. (2008) after combined BDNF and aFGF infusion. Importantly, the latter study also used immunohistochemical techniques to demonstrate that both the fibers in the osseous spiral lamina and ectopic sprouting fibers were afferent peripheral processes of SG neurons. Interesting recent work has shown that adenoviral-mediated expression of BDNF in the cochlea, presumably eliciting much lower concentrations of neurotrophin, also can induce radial nerve fiber regrowth (Shibata et al. 2010; Wise et al. 2010).
Our findings also showed that the larger population of radial nerve fibers surviving in the BDNF + ES cochleae had substantially larger cross-sectional areas than fibers in the contralateral deafened ears. After combined BDNF + ES, fiber areas were significantly larger than normal (~140 %) in the three sampled areas of the basal cochlea, whereas the surviving fibers contralateral were normal in size. On the other hand, the fibers measured in these BDNF + ES ears were substantially smaller than fibers in our earlier study of ears examined immediately after 10 weeks of BDNF infusion, in which fiber areas averaged 175 % of normal (Leake et al. 2011). Thus, our new data suggest a return toward more normal values after terminating BDNF and continuing ES over these prolonged periods.
Along with the improved survival and larger size of the SG radial nerve fibers in the BDNF + ES cochleae, the marked ectopic and disorganized sprouting of these neurons into the scala tympani is also very important. The hypertrophy of the Schwann cells seen in the osseous spiral lamina and in proximity to sprouting fibers suggest that glial cell activation may be associated with the ectopic neural remodeling in the BDNF-treated ears. Djalali et al. (2005) reported that BDNF is required for the maintenance of glial cells and production of myelin basic protein. Given that the high concentration of BDNF delivered in the present study also resulted in increased fibrotic response to the CI and angiogenesis within Rosenthal’s canal observed (see below), it seems likely that a glial cell response may also have occurred in the auditory nerve in these animals. Although no clear evidence of this was seen in the present histological studies, additional work (e.g., immunohistochemistry to assess myelin basic protein) would be of interest in addressing this issue in the future.
Angiogenesis and fibrosis
It has been reported previously that BDNF promotes endothelial cell survival and induces neoangiogenesis (Kermani and Hempstead 2007) in many tissues and organs, during development and also in response to injury. In the present study, we documented significant angiogenesis within Rosenthal’s canal in the BDNF + ES cochleae, where vessel area was more than doubled, in contrast to the normal vessel areas observed both in the contralateral deaf ears and in the cochleae examined after equivalent durations of ES alone. This must be considered a significant pathological finding, although the significance of such marked angiogenesis is unclear. Similarly, the fibrotic encapsulation of the CI electrode was significantly thicker after combined BDNF + ES ears as compared to ES only. This is a concern because thicker fibrosis would tend to displace the electrode away from the osseous spiral lamina and modiolus, which could increase thresholds to electrical stimulation and result in limited dynamic range. It seems likely that both of these findings may be related to the relatively high concentrations of neurotrophin delivered by direct intracochlear infusion using osmotic pumps in the present study.
Functional consequences of deafness, BDNF, and ES
The potential functional consequences of BDNF and improved neuronal survival are obviously extremely important in terms of the implications for potential clinical neurotrophin therapies. Data from acute terminal electrophysiological experiments in both BDNF + ES and control groups (ES-only or Implant + AP) demonstrated that final EABR thresholds were well-correlated with minimum neural thresholds in the IC, as expected. Further, the fundamental cochleotopic (frequency) organization of the IC was maintained both in animals studied as adults after BDNF and/or ES, and also in untreated controls, despite deafening prior to hearing onset, and also despite any potential distortion of the cochleotopic map due to SG fiber sprouting. That is, the minimum thresholds for apical to basal channels of the CI occurred in a clear superficial-to-deep order, reflecting the normal low-to-high frequency organization of the IC. We interpret these findings to indicate that the basic cochleotopic organization at the level of the midbrain is relatively stable, and its persistence in our animal model of congenital deafness is presumably why a CI can be effective even in a congenitally deaf child. However, it is important to remember that our ability to examine the frequency resolution of the ICC is significantly limited by the 1-mm separation of CI stimulation sites in deafened animals, as compared to the precise frequency selectivity in a normal hearing animal.
Moreover, additional findings suggest potentially deleterious effects of BDNF treatment. Specifically, data in subjects receiving only ES and in other deafened control animals showed that central auditory neuronal thresholds were correlated with relative selectivity of activation (i.e., IC activation widths at 2 dB above threshold). In contrast, no such correlation was seen in BDNF-treated subjects, and this group also exhibited a much greater intersubject variability in both IC thresholds and activation widths. The finding of lower thresholds associated with a much wider range of 2 dB activation widths indicates that improvement in—or even equivalent—selectivity of stimulation may not be achieved with the improved neural survival in the BDNF-treated ears. In fact, it seems likely that this finding is a direct result of the ectopic disorganized sprouting seen in these ears. Sprouted fibers would produce exceptionally low thresholds due to the extremely close coupling of some myelinated axons directly adjacent to the stimulating contacts of the CI. However, the excited SG cells from which these fibers originate could be located at some distance away, representing a significant offset in cochlear place frequency. Thus, the variability in activation widths after BDNF and the disorganized sprouting are of particular concern for the potential application of neurotrophin therapy in the human cochlea.
Clinical implications
Contemporary cochlear implants are generally very effective. On average, CI users with the latest technology enjoy speech recognition scores of about 80 % correct on high-context sentences, the majority are able to use a telephone (Zeng et al. 2008), and those with the best outcomes even enjoy music (Drennan and Rubinstein 2008; Won et al. 2011). However, results on more difficult tasks such as word recognition show that there is great variability among CI recipients (Firszt et al. 2004; Holden et al. 2013). This variability in outcomes has refocused attention on the importance of auditory nerve survival for CI function and on neurotrophic agents that might be used with a CI to improve auditory nerve survival (for review, see Staecker et al. 2010). In fact, human CIs modified for intracochlear drug delivery already have been designed (Hochmair et al. 2006) and tested in animals (Paasche et al. 2003). However, animal studies examining the long-term effects of potential neurotrophic agents have been quite limited to date.
The issue of maintaining auditory nerve survival seems particularly important in very young deaf children and congenitally deaf infants who are now receiving CIs (Nicholas and Geers 2007) because they will have to acquire speech and language and depend on electrical hearing for many decades. With pediatric implants, there is evidence that restoring auditory input early is more effective in allowing the immature auditory system to adapt to electrical stimulation, and it is encouraging that many children with CIs eventually are mainstreamed into public education settings (Geers 2004; Nicholas and Geers 2007; Svirsky et al. 2000). However, the same studies show that many other pediatric CI recipients lag far behind in language development. Thus, we suggest that it is critically important to better understand the specific factors and mechanisms underlying SG degeneration following early onset deafness and the role of neurotrophins (for review see Ramekers et al. 2012). To our knowledge, our studies are the first to report on the effects of exogenous neurotrophins combined with electrical stimulation in deafened, developing animals. Our findings demonstrate that exogenous intracochlear BDNF combined with chronic ES can result in substantially improved survival of the cochlear SG neurons in an animal model of congenital deafness. Together with the many previous studies showing SG protection with BDNF in adult animals, findings suggest that neurotrophic agents can be successfully combined with CI stimulation and may offer promise for clinical therapies to promote improved cochlear neuronal survival, both in the adult and developing auditory system. However, the use of osmotic pumps to deliver neurotrophic factors clearly is not a good option for clinical application, particularly due to concerns about infection (Shepherd et al. 2008; Staecker et al. 2010). Numerous recent studies have explored alternative strategies for cochlear delivery of neurotrophins (Hendricks et al. 2008; Richardson et al. 2008), including cell-based therapies (Warnecke et al. 2007; Pettingill et al. 2008; Wise et al. 2011), hydrogels (Endo et al. 2005), and gene therapy using adenovirus-mediated expression of neurotrophic factors (Nakaizumi et al. 2004; Rejali et al. 2007; Chikar et al. 2008; Wise et al. 2010). These methods may provide better alternatives in the future, but they are still under development and concerns about potential side effects and risks have not yet been adequately addressed. For recent reviews see: Ramekers et al. (2012) and Shibata et al. (2010).
Finally, the goal of neurotrophin therapy is to improve neural survival for the purpose of supporting optimized function of a CI, and it is important to emphasize that recent advances in CI technologies, such as current, focusing, virtual channel stimulation, and other more complex stimulation strategies, rely on highly spatially restricted current fields. We suggest that it is a serious concern that the ectopic and disorganized sprouting of the radial nerve fibers induced by exogenous neurotrophins may be deleterious to these objectives. In fact, in the cochleae studied in the present BDNF + ES experiments, fascicles of these fibers were traced in a spiral course for several hundred microns, probably because of the longer durations of treatment and survival studied. These fibers appear to survive over the long-term, and they clearly could blur or distort the precise cochlear frequency map, which would be detrimental to optimum implant function. We hypothesize that with improved methods for delivering neurotrophic agents in more physiological concentrations (e.g., viral-mediated neurotrophin expression), it may be possible to elicit improved neuronal and radial nerve fiber survival, and also maintain more normal cochlear neuroanatomy of the radial nerve fibers by avoiding the ectopic and disorganized exuberant sprouting seen with high concentrations of neurotrophins.
Acknowledgments
The authors thank Chantale Dore for assistance in surgery, care of chronic animals, and measurements of radial nerve fibers; Mr. Larry Ackerman for expert sectioning of the specimens for TEM; and Mr. Paolo Machado for help in the sectioning and counting of the radial nerve fibers. This work was supported by the U.S. National Institutes of Health, the National Institute on Deafness and Other Communication Disorders, Contract #HHS-N-263-2007-00054-C, the Epstein Fund and Hearing Research, Inc., Human recombinant BDNF for these studies was donated by Amgen, Inc., Thousand Oaks, CA.
References
- Agterberg MJH, Versnel H, de Groot JCMJ, et al. Morphological changes in spiral ganglion cells after intracochlear application of brain-derived neurotrophic factor in deafened guinea pigs. Hear Res. 2008;244:25–34. doi: 10.1016/j.heares.2008.07.004. [DOI] [PubMed] [Google Scholar]
- Agterberg MJH, Versnel H, van Dijk LM, et al. Enhanced survival of spiral ganglion cells after cessation of treatment with brain-derived neurotrophic factor in deafened guinea pigs. J Assoc Res Otolaryngol. 2009;10:355–367. doi: 10.1007/s10162-009-0170-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Araki S, Kawano A, Seldon L, et al. Effects of chronic electrical stimulation on spiral ganglion neuron survival and size in deafened kittens. Laryngoscope. 1998;108:687–695. doi: 10.1097/00005537-199805000-00012. [DOI] [PubMed] [Google Scholar]
- Chikar JA, Colesa DJ, Swiderski DL, et al. Over-expression of BDNF by adenovirus with concurrent electrical stimulation improves cochlear implant thresholds and survival of auditory neurons. Hear Res. 2008;245:24–34. doi: 10.1016/j.heares.2008.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coco A, Epp SB, Fallon JB, et al. Does cochlear implantation and electrical stimulation affect residual hair cells and spiral ganglion neurons? Hear Res. 2007;225:60–70. doi: 10.1016/j.heares.2006.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Djalali S, Holtje M, Grosse G et al (2005) Effects of brain-derived neurotrophic factor (BDNF) on glial cells and serotonergic neurons during development. J Neurochem 92:616–627 [DOI] [PubMed]
- Don M, Elberling C. Evaluating residual background noise in human auditory brain-stem responses. J Acoust Soc Am. 1994;96:2746–2757. doi: 10.1121/1.411281. [DOI] [PubMed] [Google Scholar]
- Drennan WR, Rubinstein JT. Music perception in cochlear implant users and its relationship with psychophysical capabilities. J Rehabil Res Dev. 2008;45:779–789. doi: 10.1682/JRRD.2007.08.0118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elverland HH, Mair IW. Hereditary deafness in the cat. An electron microscopic study of the spiral ganglion. Acta Otolaryngol. 1980;90:360–369. doi: 10.3109/00016488009131737. [DOI] [PubMed] [Google Scholar]
- Endo T, Nakagawa T, Kita T, et al. Novel strategy for treatment of inner ears using a biodegradable gel. Laryngoscope. 2005;115:2016–2020. doi: 10.1097/01.mlg.0000183020.32435.59. [DOI] [PubMed] [Google Scholar]
- Fariñas I, Jones KR, Tessarollo L, et al. Spatial shaping of cochlear innervation by temporally regulated neurotrophin expression. J Neurosci. 2001;21:6170–6180. doi: 10.1523/JNEUROSCI.21-16-06170.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Firszt JB, Holden LK, Skinner MW, et al. Recognition of speech presented at soft to loud levels by adult cochlear implant recipients of three cochlear implant systems. Ear Hear. 2004;25:375–387. doi: 10.1097/01.AUD.0000134552.22205.EE. [DOI] [PubMed] [Google Scholar]
- Fritzsch B, Pirvola U, Ylikoski J. Making and breaking the innervation of the ear: neurotrophic support during ear development and its clinical implications. Cell Tissue Res. 1999;295:369–382. doi: 10.1007/s004410051244. [DOI] [PubMed] [Google Scholar]
- Gao WQ, Zheng JL, Karihaloo M. Neurotrophin-4/5 (NT-4/5) and brain-derived neurotrophic factor (BDNF) act at later stages of cerebellar granule cell differentiation. J Neurosci. 1995;15:2656–2667. doi: 10.1523/JNEUROSCI.15-04-02656.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geers AE. Speech, language, and reading skills after early cochlear implantation. Arch Otolaryngol Head Neck Surg. 2004;130:634–638. doi: 10.1001/archotol.130.5.634. [DOI] [PubMed] [Google Scholar]
- Gillespie LN, Clark GM, Bartlett PF, Marzella PL. BDNF-induced survival of auditory neurons in vivo: Cessation of treatment leads to accelerated loss of survival effects. J Neurosci Res. 2003;71:785–790. doi: 10.1002/jnr.10542. [DOI] [PubMed] [Google Scholar]
- Glueckert R, Bitsche M, Miller JM, et al. Deafferentation-associated changes in afferent and efferent processes in the guinea pig cochlea and afferent regeneration with chronic intrascalar brain-derived neurotrophic factor and acidic fibroblast growth factor. J Comp Neurol. 2008;507:1602–1621. doi: 10.1002/cne.21619. [DOI] [PubMed] [Google Scholar]
- Hansen MR, Zha XM, Bok J, Green SH. Multiple distinct signal pathways, including an autocrine neurotrophic mechanism, contribute to the survival-promoting effect of depolarization on spiral ganglion neurons in vitro. J Neurosci. 2001;21:2256–2267. doi: 10.1523/JNEUROSCI.21-07-02256.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen MR, Bok J, Devaiah AK, et al. Ca2+/calmodulin-dependent protein kinases II and IV both promote survival but differ in their effects on axon growth in spiral ganglion neurons. J Neurosci Res. 2003;72:169–184. doi: 10.1002/jnr.10551. [DOI] [PubMed] [Google Scholar]
- Hartmann R, Topp G, Klinke R. Discharge patterns of cat primary auditory fibers with electrical stimulation of the cochlea. Hear Res. 1984;13:47–62. doi: 10.1016/0378-5955(84)90094-7. [DOI] [PubMed] [Google Scholar]
- Hartshorn DO, Miller JM, Altschuler RA. Protective effect of electrical stimulation in the deafened guinea pig cochlea. Otolaryngol Head Neck Surg. 1991;104:311–319. doi: 10.1177/019459989110400305. [DOI] [PubMed] [Google Scholar]
- Hegarty JL, Kay AR, Green SH. Trophic support of cultured spiral ganglion neurons by depolarization exceeds and is additive with that by neurotrophins or cAMP and requires elevation of [Ca2+]i within a set range. J Neurosci. 1997;17:1959–1970. doi: 10.1523/JNEUROSCI.17-06-01959.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hendricks JL, Chikar JA, Crumling MA, et al. Localized cell and drug delivery for auditory prostheses. Hear Res. 2008;242:117–131. doi: 10.1016/j.heares.2008.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hochmair I, Nopp P, Jolly C, et al. MED-EL Cochlear implants: state of the art and a glimpse into the future. Trends Amplif. 2006;10:201–219. doi: 10.1177/1084713806296720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holden, LK, Finley, CC, Firszt JB, et al (2013) Factors affecting open-set word recognition in adults with cochlear implants. Ear Hear. doi:10.1097/AUD.0b013e3182741aa7 [DOI] [PMC free article] [PubMed]
- Kanzaki S, Stöver T, Kawamoto K, et al. Glial cell line-derived neurotrophic factor and chronic electrical stimulation prevent VIII cranial nerve degeneration following denervation. J Comp Neurol. 2002;454:350–360. doi: 10.1002/cne.10480. [DOI] [PubMed] [Google Scholar]
- Kermani P, Hempstead B. Brain-derived neurotrophic factor: a newly described mediator of angiogenesis. Trends Cardiovasc Med. 2007;17:140–143. doi: 10.1016/j.tcm.2007.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiang NY, Rho JM, Northrop CC, et al. Hair-cell innervation by spiral ganglion cells in adult cats. Science. 1982;217:175–177. doi: 10.1126/science.7089553. [DOI] [PubMed] [Google Scholar]
- Korsching S. The neurotrophic factor concept: a reexamination. J Neurosci. 1993;13:2739–2748. doi: 10.1523/JNEUROSCI.13-07-02739.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leake PA, Hradek GT. Cochlear pathology of long term neomycin induced deafness in cats. Hear Res. 1988;33:11–33. doi: 10.1016/0378-5955(88)90018-4. [DOI] [PubMed] [Google Scholar]
- Leake PA, Kuntz AL, Moore CM, Chambers PL. Cochlear pathology induced by aminoglycoside ototoxicity during postnatal maturation in cats. Hear Res. 1997;113:117–132. doi: 10.1016/S0378-5955(97)00133-0. [DOI] [PubMed] [Google Scholar]
- Leake PA, Hradek GT, Snyder RL. Chronic electrical stimulation by a cochlear implant promotes survival of spiral ganglion neurons after neonatal deafness. J Comp Neurol. 1999;412:543–562. doi: 10.1002/(SICI)1096-9861(19991004)412:4<543::AID-CNE1>3.0.CO;2-3. [DOI] [PubMed] [Google Scholar]
- Leake PA, Hradek GT, Vollmer M, Rebscher SJ. Neurotrophic effects of GM1 ganglioside and electrical stimulation on cochlear spiral ganglion neurons in cats deafened as neonates. J Comp Neurol. 2007;501:837–853. doi: 10.1002/cne.21275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leake PA, Hradek GT, Bonham BH, Snyder RL. Topography of auditory nerve projections to the cochlear nucleus in cats after neonatal deafness and electrical stimulation by a cochlear implant. J Assoc Res Otolaryngol. 2008;9:349–372. doi: 10.1007/s10162-008-0127-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leake PA, Stakhovskaya O, Hradek GT, Hetherington AM. Factors influencing neurotrophic effects of electrical stimulation in the deafened developing auditory system. Hear Res. 2008;242:86–99. doi: 10.1016/j.heares.2008.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leake PA, Hradek GT, Hetherington AM, Stakhovskaya O. Brain-derived neurotrophic factor promotes cochlear spiral ganglion cell survival and function in deafened, developing cats. J Comp Neurol. 2011;519:1526–1545. doi: 10.1002/cne.22582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Parkins CW, Webster DB. Does electrical stimulation of deaf cochleae prevent spiral ganglion degeneration? Hear Res. 1999;133:27–39. doi: 10.1016/S0378-5955(99)00043-X. [DOI] [PubMed] [Google Scholar]
- Liberman MC. The cochlear frequency map for the cat: labeling auditory-nerve fibers of known characteristic frequency. J Acoust Soc Am. 1982;72:1441–1449. doi: 10.1121/1.388677. [DOI] [PubMed] [Google Scholar]
- Liberman MC, Kiang NY. Acoustic trauma in cats. Cochlear pathology and auditory-nerve activity. Acta Otolaryngol Suppl. 1978;358:1–63. [PubMed] [Google Scholar]
- Liberman MC, Dodds LW, Pierce S. Afferent and efferent innervation of the cat cochlea: quantitative analysis with light and electron microscopy. J Comp Neurol. 1990;301:443–460. doi: 10.1002/cne.903010309. [DOI] [PubMed] [Google Scholar]
- Lousteau RJ. Increased spiral ganglion cell survival in electrically stimulated, deafened guinea pig cochleae. Laryngoscope. 1987;97:836–842. doi: 10.1288/00005537-198707000-00012. [DOI] [PubMed] [Google Scholar]
- Maruyama J, Miller JM, Ulfendahl M. Glial cell line-derived neurotrophic factor and antioxidants preserve the electrical responsiveness of the spiral ganglion neurons after experimentally induced deafness. Neurobiol Dis. 2008;29:14–21. doi: 10.1016/j.nbd.2007.07.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGuinness SL, Shepherd RK. Exogenous BDNF rescues rat spiral ganglion neurons in vivo. Otol Neurotol. 2005;26:1064–1072. doi: 10.1097/01.mao.0000185063.20081.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller AL. Effects of chronic stimulation on auditory nerve survival in ototoxically deafened animals. Hear Res. 2001;151:1–14. doi: 10.1016/S0378-5955(00)00226-4. [DOI] [PubMed] [Google Scholar]
- Miller JM, Altschuler RA. Effectiveness of different electrical stimulation conditions in preservation of spiral ganglion cells following deafness. Ann Otol Rhinol Laryngol Suppl. 1995;166:57–60. [PubMed] [Google Scholar]
- Miller JM, Le Prell CG, Prieskorn DM, et al. Delayed neurotrophin treatment following deafness rescues spiral ganglion cells from death and promotes regrowth of auditory nerve peripheral processes: effects of brain-derived neurotrophic factor and fibroblast growth factor. J Neurosci Res. 2007;85:1959–1969. doi: 10.1002/jnr.21320. [DOI] [PubMed] [Google Scholar]
- Mitchell A, Miller JM, Finger PA, et al. Effects of chronic high-rate electrical stimulation on the cochlea and eighth nerve in the deafened guinea pig. Hear Res. 1997;105:30–43. doi: 10.1016/S0378-5955(96)00202-X. [DOI] [PubMed] [Google Scholar]
- Mouton PR. Principles and practices of unbiased stereology: an introduction for bioscientists. Baltimore: The Johns Hopkins University Press; 2002. [Google Scholar]
- Nakaizumi T, Kawamoto K, Minoda R, Raphael Y. Adenovirus-mediated expression of brain-derived neurotrophic factor protects spiral ganglion neurons from ototoxic damage. Audiol Neurootol. 2004;9:135–143. doi: 10.1159/000077264. [DOI] [PubMed] [Google Scholar]
- Nicholas JG, Geers AE. Will they catch up? The role of age at cochlear implantation in the spoken language development of children with severe to profound hearing loss. J Speech Lang Hear Res. 2007;50:1048–1062. doi: 10.1044/1092-4388(2007/073). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paasche G, Gibson P, Averbeck T, et al. Technical report: modification of a cochlear implant electrode for drug delivery to the inner ear. Otol Neurotol. 2003;24:222–227. doi: 10.1097/00129492-200303000-00016. [DOI] [PubMed] [Google Scholar]
- Pettingill LN, Minter RL, Shepherd RK. Schwann cells genetically modified to express neurotrophins promote spiral ganglion neuron survival in vitro. Neuroscience. 2008;152:821–828. doi: 10.1016/j.neuroscience.2007.11.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramekers D, Versnel H, Grolman W, Klis SFL (2012) Neurotrophins and their role in the cochlea. Hear Res 288:19–33. doi:10.1016/j.heares.2012.03.002 [DOI] [PubMed]
- Rebscher SJ, Hetherington AM, Snyder RL, et al. Design and fabrication of multichannel cochlear implants for animal research. J Neurosci Methods. 2007;166:1–12. doi: 10.1016/j.jneumeth.2007.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rejali D, Lee VA, Abrashkin KA, et al. Cochlear implants and ex vivo BDNF gene therapy protect spiral ganglion neurons. Hear Res. 2007;228:180–187. doi: 10.1016/j.heares.2007.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson RT, Wise AK, Andrew JK, O’Leary SJ. Novel drug delivery systems for inner ear protection and regeneration after hearing loss. Expert Opin Drug Deliv. 2008;5:1059–1076. doi: 10.1517/17425247.5.10.1059. [DOI] [PubMed] [Google Scholar]
- Roehm PC, Hansen MR. Strategies to preserve or regenerate spiral ganglion neurons. Curr Opin Otolaryngol Head Neck Surg. 2005;13:294–300. doi: 10.1097/01.moo.0000180919.68812.b9. [DOI] [PubMed] [Google Scholar]
- Romand MR, Romand R. Development of spiral ganglion cells in mammalian cochlea. J Electron Microsc Tech. 1990;15:144–154. doi: 10.1002/jemt.1060150206. [DOI] [PubMed] [Google Scholar]
- Romand R, Romand MR, Mulle C, Marty R. Early stages of myelination in the spiral ganglion cells of the kitten during development. Acta Otolaryngol. 1980;90:391–397. doi: 10.3109/00016488009131740. [DOI] [PubMed] [Google Scholar]
- Rubel EW, Fritzsch B. Auditory system development: primary auditory neurons and their targets. Annu Rev Neurosci. 2002;25:51–101. doi: 10.1146/annurev.neuro.25.112701.142849. [DOI] [PubMed] [Google Scholar]
- Scheper V, Paasche G, Miller JM, et al. Effects of delayed treatment with combined GDNF and continuous electrical stimulation on spiral ganglion cell survival in deafened guinea pigs. J Neurosci Res. 2009;87:1389–1399. doi: 10.1002/jnr.21964. [DOI] [PubMed] [Google Scholar]
- Shepherd RK, Javel E. Electrical stimulation of the auditory nerve. I. Correlation of physiological responses with cochlear status. Hear Res. 1997;108:112–144. doi: 10.1016/S0378-5955(97)00046-4. [DOI] [PubMed] [Google Scholar]
- Shepherd RK, Matsushima J, Martin RL, Clark GM. Cochlear pathology following chronic electrical stimulation of the auditory nerve: II. Deafened kittens. Hear Res. 1994;81:150–166. doi: 10.1016/0378-5955(94)90162-7. [DOI] [PubMed] [Google Scholar]
- Shepherd RK, Coco A, Epp SB, Crook JM. Chronic depolarization enhances the trophic effects of brain-derived neurotrophic factor in rescuing auditory neurons following a sensorineural hearing loss. J Comp Neurol. 2005;486:145–158. doi: 10.1002/cne.20564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shepherd RK, Coco A, Epp SB. Neurotrophins and electrical stimulation for protection and repair of spiral ganglion neurons following sensorineural hearing loss. Hear Res. 2008;242:100–109. doi: 10.1016/j.heares.2007.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shibata SB, Cortez SR, Beyer LA, et al. Transgenic BDNF induces nerve fiber regrowth into the auditory epithelium in deaf cochleae. Exp Neurol. 2010;223:464–472. doi: 10.1016/j.expneurol.2010.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinohara T, Bredberg G, Ulfendahl M, et al. Neurotrophic factor intervention restores auditory function in deafened animals. Proc Natl Acad Sci U S A. 2002;99:1657–1660. doi: 10.1073/pnas.032677999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snyder RL, Bonham BH, Sinex DG. Acute changes in frequency responses of inferior colliculus central nucleus (ICC) neurons following progressively enlarged restricted spiral ganglion lesions. Hear Res. 2008;246:59–78. doi: 10.1016/j.heares.2008.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spoendlin H. Innervation patterns in the organ of corti of the cat. Acta Otolaryngol. 1969;67:239–254. doi: 10.3109/00016486909125448. [DOI] [PubMed] [Google Scholar]
- Spoendlin H. Retrograde degeneration of the cochlear nerve. Acta Otolaryngol. 1975;79:266–275. doi: 10.3109/00016487509124683. [DOI] [PubMed] [Google Scholar]
- Spoendlin H. Differentiation of cochlear afferent neurons. Acta Otolaryngol. 1981;91:451–456. doi: 10.3109/00016488109138527. [DOI] [PubMed] [Google Scholar]
- Staecker H, Jolly C, Garnham C. Cochlear implantation: an opportunity for drug development. Drug Discov Today. 2010;15:314–321. doi: 10.1016/j.drudis.2010.02.005. [DOI] [PubMed] [Google Scholar]
- Sugarawara M, Corfas G, Liberman MC. Influence of supporting cells on neuronal degeneration after hair cell loss. JARO. 2005;6:136–147. doi: 10.1007/s10162-004-5050-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svirsky MA, Robbins AM, Kirk KI, et al. Language development in profoundly deaf children with cochlear implants. Psychol Sci. 2000;11:153–158. doi: 10.1111/1467-9280.00231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tessarollo L, Coppola V, Fritzsch B. NT-3 replacement with brain-derived neurotrophic factor redirects vestibular nerve fibers to the cochlea. J Neurosci. 2004;24:2575–2584. doi: 10.1523/JNEUROSCI.5514-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warnecke A, Wissel K, Hoffmann A, et al. The biological effects of cell-delivered brain-derived neurotrophic factor on cultured spiral ganglion cells. Neuroreport. 2007;18:1683–1686. doi: 10.1097/WNR.0b013e3282f0b5d7. [DOI] [PubMed] [Google Scholar]
- Wefstaedt P, Scheper V, Lenarz T, Stöver T. Brain-derived neurotrophic factor/glial cell line-derived neurotrophic factor survival effects on auditory neurons are not limited by dexamethasone. Neuroreport. 2005;16:2011–2014. doi: 10.1097/00001756-200512190-00008. [DOI] [PubMed] [Google Scholar]
- Wise AK, Richardson R, Hardman J, et al. Resprouting and survival of guinea pig cochlear neurons in response to the administration of the neurotrophins brain-derived neurotrophic factor and neurotrophin-3. J Comp Neurol. 2005;487:147–165. doi: 10.1002/cne.20563. [DOI] [PubMed] [Google Scholar]
- Wise AK, Hume CR, Flynn BO, et al. Effects of localized neurotrophin gene expression on spiral ganglion neuron resprouting in the deafened cochlea. Mol Ther. 2010;18:1111–1122. doi: 10.1038/mt.2010.28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wise AK, Fallon JB, Neil AJ, et al. Combining cell-based therapies and neural prostheses to promote neural survival. Neurotherapeutics. 2011;8:774–787. doi: 10.1007/s13311-011-0070-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Won JH, Drennan WR, Nie K, et al. Acoustic temporal modulation detection and speech perception in cochlear implant listeners. J Acoust Soc Am. 2011;130:376–388. doi: 10.1121/1.3592521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yagi M, Kanzaki S, Kawamoto K, et al. Spiral ganglion neurons are protected from degeneration by GDNF gene therapy. J Assoc Res Otolaryngol. 2000;1:315–325. doi: 10.1007/s101620010011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamagata T, Miller JM, Ulfendahl M, et al. Delayed neurotrophic treatment preserves nerve survival and electrophysiological responsiveness in neomycin-deafened guinea pigs. J Neurosci Res. 2004;78:75–86. doi: 10.1002/jnr.20239. [DOI] [PubMed] [Google Scholar]
- Yang T, Kersigo J, Jahan I, et al. The molecular basis of making spiral ganglion neurons and connecting them to hair cells of the organ of Corti. Hear Res. 2011;278:21–33. doi: 10.1016/j.heares.2011.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ylikoski J, Pirvola U, Virkkala J, et al. Guinea pig auditory neurons are protected by glial cell line-derived growth factor from degeneration after noise trauma. Hear Res. 1998;124:17–26. doi: 10.1016/S0378-5955(98)00095-1. [DOI] [PubMed] [Google Scholar]
- Zeng F-G, Rebscher S, Harrison W, et al. Cochlear implants: system design, integration, and evaluation. IEEE Rev Biomed Eng. 2008;1:115–142. doi: 10.1109/RBME.2008.2008250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zha XM, Bishop JF, Hansen MR, et al. BDNF synthesis in spiral ganglion neurons is constitutive and CREB-dependent. Hear Res. 2001;156:53–68. doi: 10.1016/S0378-5955(01)00267-2. [DOI] [PubMed] [Google Scholar]
- Zilberstein Y, Liberman MC, Corfas G. Inner hair cells are not required for survival of spiral ganglion neurons in the adult cochlea. J Neurosci. 2012;32(2):405–410. doi: 10.1523/JNEUROSCI.4678-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]